Introduction
Colorectal cancer (CRC) is one of the most common
malignant tumors. The incidence rate of CRC in women ranks third
only to lung cancer and breast cancer, and ranks fourth only to
lung cancer, stomach cancer and liver cancer in men (1). According to the World Health
Organization's International Agency for Research on Cancer, in 2018
there were ~1.8 million new CRC diagnoses, accounting for 10% of
all patients with cancer, and ~860,000 CRC related-deaths,
accounting for 9.9% of all cancer-related mortalities (1). The prevalence of CRC has been
increasing over the last 5–10 years, with an estimated 2.2 million
new CRC cases and 1.1 million deaths worldwide predicted by 2030
(2). In 1990, CRC was ranked 21st
among the leading causes of death in China, but in 2017 it rose to
the 11th, with a CRC mortality rate of 13.24 per 100,000 people
(3). In 2017, CRC was therefore
accountable for 1.79% of all deaths, seriously threatening human
health and imposing a huge economic burden on society (3). Thus, it is important to find a new,
safe and effective treatment for CRC.
According to the Gene Expression Omnibus database
(dataset, GSE10950), the expression of mitochondrial fission
regulator (MTFR) 2 was significantly upregulated in patients with
CRC (4). MTFR2 is a gene that
encodes mitochondrial proteins and belongs to the MTFR1/family with
sequence similarity 54 member A family. MTFR2 induces fission of
mitochondria in eukaryotic cells and provides energy for aerobic
respiration (5). Wang et al
(6) demonstrated that MTFR2 is
highly expressed in glioblastoma and is associated with a poor
prognosis (6). MTFR2 has also been
reported to be highly expressed in oral squamous cell carcinoma
tissues and to be negatively related to the overall survival (OS)
of patients (7). MTFR2 may also
aggravate proliferation, migration and invasion of oral squamous
carcinoma cells by switching oxidative phosphorylation to
glycolysis (7). A previous study
revealed that high MTFR2 expression is related to poor prognosis in
patients with breast cancer (BC) and is more prevalent in patients
with aggressive tumors (8). The
invasion, migration and epithelial-mesenchymal transition of BC
cells has also been shown to be suppressed by the knockdown of
MTFR2 (9). However, the biological
function of MTFR2 in CRC remains unclear.
A previous study revealed that HOXC10 expression is
significantly higher in CRC samples than in normal adjacent
tissues, and may be used as a biomarker for the diagnosis of CRC
and as a potential therapeutic target (10). HOXC10 is also considered a key
modulator in ovarian cancer (OC), which is associated with poor
prognosis in patients with OC, actively regulating Slug
transcription to promote OC metastasis (11). Furthermore, knockdown of HOXC10 has
been reported to suppress the proliferation, migration and invasion
of gastric cancer (GC) cells in vitro, and to suppress tumor
growth and induce apoptosis in vivo (12). HOXC10 expression is also markedly
upregulated in glioblastoma tissues and cells, resulting in a poor
OS in patients with glioblastoma. Notably, inhibition of HOXC10
could inhibit the proliferation, migration and invasion of
glioblastoma cells (13).
Therefore, it is speculated that HOXC10 could also regulate CRC
progression.
The aim of the present study was to investigate
whether HOXC10 binding to MTFR2 could regulate the proliferation,
invasion and migration of CRC cells.
Materials and methods
Human tumor samples
The present study was approved by the Clinical
Research Ethics Committee of Nanjing Medical University (Nanjing,
China; approval no. 2021–448) and written informed consent was
obtained from each patient. In total, CRC tissues and
paraneoplastic tissues (>2 cm away from the edge of the tumor)
were obtained from 16 patients with CRC in the Jiangsu Cancer
Hospital (Nanjing, China) between May 2019 and June 2020. None of
the patients had received cancer treatment beforehand. The tumor
samples were stored at −80°C until further use. The
histopathological characteristics of the patients are shown in
Table I.
 | Table I.Histopathological characteristics of
the patients. |
Table I.
Histopathological characteristics of
the patients.
| No. | Age, years | Sex | Number of LNM | Grading | Tumor location | Differentiation
degree | Distant
metastasis |
|---|
| 1 | 63 | Female | 1 | II | Sigmoid colon | Middle-low | M1 |
| 2 | 16 | Male | 1 | IV | Middle segment of
transverse colon | Low | M1 |
| 3 | 69 | Female | 0 | II | Rectum | Middle | M0 |
| 4 | 39 | Male | 0 | II | Transverse
colon | Middle | M0 |
| 5 | 66 | Male | 1 | III | Junction of rectum
and sigmoid colon | Middle | M0 |
| 6 | 43 | Female | 2 | III | Sigmoid colon | Middle | M0 |
| 7 | 60 | Male | 0 | I | Rectum | Middle | M0 |
| 8 | 75 | Female | 0 | II | Ascending
colon | Middle | M0 |
| 9 | 67 | Female | 0 | I | Rectum | Middle | M0 |
| 10 | 63 | Female | 0 | II | Transverse colon
(proximal to hepatic curvature) | Middle | M0 |
| 11 | 48 | Female | 0 | I | Rectum | Middle | M0 |
| 12 | 54 | Male | 2 | III | Sigmoid colon | Middle | M0 |
| 13 | 54 | Female | 1 | III | Colon | Middle | M0 |
| 14 | 51 | Male | 2 | IV | Sigmoid colon | Middle-low | M1 |
| 15 | 66 | Female | 0 | IV | Ileocecal
junction | Middle-low | M1 |
| 16 | 55 | Male | 1 | III | Rectum | Middle | M0 |
Cell culture and transfection
The human intestinal epithelial cell line (HIEC-6)
and colorectal cancer cell lines (Caco-2, HCT116, SW480 and LoVo)
were all obtained from the American Type Culture Collection. Cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc.) containing 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin solution at 37°C with 5% CO2.
Short hairpin RNA (sh)-MTFR2-1/2 (its negative control was sh-NC)
and overexpression (Oe)-HOXC10 (its negative control was Oe-NC)
plasmids were provided by Shanghai GenePharma Co., Ltd. HCT116
cells were seeded at 1.5×105 cells/well in 6-well plates
for 24 h at 37°C. When cell density reached 60% confluence, HCT116
cells were transfected with sh-NC (40 nM), sh-MTFR2-1/2 (40 nM),
Oe-NC (60 nM) and Oe-HOXC10 (60 nM) using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Cells were then cultured in normal medium
at 37°C for 48 h and subsequent experiments were conducted.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells and tissue
samples using TRIzol® reagent (Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Total RNA (1 µg)
was reverse transcribed into cDNA at 42°C for 30 min, according to
the manufacturer's protocol of the PrimeScript RT Reagent Kit (cat.
no. RR037A; Takara Biotechnology Co., Ltd.). qPCR was performed
using SYBR Premix Ex Taq (Takara Biotechnology Co., Ltd.) on an
Applied Biosystems 7900 PCR system (Thermo Fisher Scientific,
Inc.). The thermocycling conditions were used: Initial denaturation
at 95°C for 10 min; followed by 40 cycles of denaturation at 95°C
for 15 sec and annealing at 60°C for 1 min; and a final extension
of 10 min at 72°C. The following primer pairs were used for qPCR:
MTFR2 forward, 5′-AGGGCTACGGGCCAATTTGA-3′ and reverse,
5′-TTCCTAAATAAAGTTTGGTCCAC-3′; HOXC10 forward,
5′-CTCGGATAACGAAGCGAAAG-3′ and reverse, 5′-CGCTCTCGCGTCAAATACAT-3′;
and GAPDH forward, 5′-TGTGGGCATCAATGGATTTGG−3′ and reverse,
5′-ACACCATGTATTCCGGGTCAAT-3′. MTFR2 and HOXC10 mRNA expression
levels were quantified using the 2−ΔΔCq method and
normalized to the internal reference gene GAPDH (14).
Western blotting
Total protein was extracted from cells using cold
RIPA buffer (cat. no. P0013C; Beyotime Institute of Biotechnology),
and were centrifuged at 3,000 × g for 15 min at 4°C. The
non-transfected cells were used as the control group. Total protein
was quantified using a BCA kit (Beyotime Institute of
Biotechnology) and 20 µg protein/lane was separated by SDS-PAGE on
a 12% gel. The separated proteins were transferred to PVDF
membranes and blocked with 5% non-fat milk with TBS-0.1% Tween-20
(TBST) for 1 h at room temperature. The membranes were incubated
overnight at 4°C with primary antibodies against the following:
MTFR2 (1:500; cat. no. ab155678; Abcam), MMP2 (1:1,000; cat. no.
ab92536; Abcam), MMP9 (1:1,000; cat. no. ab76003; Abcam), HOXC10
(1:500; cat. no. ab153904; Abcam) and GAPDH (1:2,500; cat. no.
ab9485; Abcam). Following the primary incubation, membranes were
washed with TBST and incubated with an appropriate HRP-conjugated
secondary antibody (1:1,000; cat. no. 7074; Cell Signaling
Technology, Inc.) for 1 h at room temperature. Protein bands were
observed using a chemiluminescence reagent (ECL) kit (Beyotime
Institute of Biotechnology). The gray values of bands were
semi-quantified using ImageJ software (version 1.0; National
Institutes of Health) with GAPDH as the loading control.
Cell Counting Kit-8 (CCK-8) assay
The proliferation of HCT116 cells was detected by
performing the CCK-8 assay. Following transfection, cells
(2×103 cells/well) were seeded into 96-well plates. The
non-transfected cells were used as the control group. Cell
proliferation was detected at 24, 48 and 72 h using the CCK-8
reagent (Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. Cells were incubated with 10 µl CCK-8
reagent for 2 h before the absorbance value at 450 nm was detected
via a microplate reader (Bio-Rad Laboratories, Inc.).
Clone formation assay
Following transfection, HCT116 cells
(1×103 cells/well) were seeded into a 6-well plate and
cultured in DMEM at 37°C for 2 weeks. The non-transfected cells
were used as the control group. Cell culture was stopped when
clones were visible to the naked eye in the culture dish. The cells
were fixed with 10% formaldehyde for 10 min at room temperature and
stained with 0.5% crystal violet for 5 min at room temperature. The
number of cloned cells was quantified by eye.
Wound healing assay
The migratory ability of HCT116 cells was detected
by the wound healing assay. Following transfection, HCT116 cells
(5×105 cells/well) were seeded into 6-well plates and
cultured until cells reached 100% confluence. The non-transfected
cells were used as the control group. Serum-free medium replaced
normal medium and a sterile 100-µl pipette tip was used to scratch
the cells (0 h). Unattached cells were removed and the remaining
cells were cultured at 37°C for 24 h. The wound gap was observed
under an inverted light microscope (magnification, ×100) and the
cell migration rate was calculated using ImageJ (version 1.52r;
National Institutes of Health).
Transwell assay
The invasive ability of HCT116 cells was detected
via the Transwell assay. Following transfection, HCT116 cells
(1×105 cells/well) in 200 µl DMEM were placedin the
upper Transwell chamber with the 8-µm pore inserted membranes
pre-coated with Matrigel (BD Biosciences) overnight at 37°C. The
non-transfected cells were used as the control group. In the lower
chamber, 600 µl DMEM containing 10% FBS was added. After cells were
cultured at 37°C for 24 h, cells in the upper side of the chamber
were removed and those in the lower side of the chamber were fixed
with methanol for 20 min at room temperature and then stained with
crystal violet for 10 min at room temperature. Cells were observed
and imaged using an inverted light microscope (magnification,
×100), and the cell invasion rate was calculated using ImageJ
(version 1.52r; National Institutes of Health). The number of
invasive cells were counted from five randomly selected fields.
Bioinformatics analysis
Japan Automotive Software Platform and Architecture
[JASPAR, 8th (2020); http://jaspar.genereg.net/] predicted that HOXC10
could bind to the promoter sequence of MTFR2.
Dual-luciferase reporter assay
The interaction between HOXC10 and the MTFR2
promoter in HCT116 cells was determined via the dual-luciferase
reporter assay. The wild-type (WT) and mutant (MUT) sequences of
MTFR2 untranslated region were amplified by Shanghai GenePharma
Co., Ltd., cloned into a pGL3 luciferase vector (Promega
Corporation) and respectively named MTFR2-WT and MTFR2-MUT. HCT116
cells at 80% confluence were co-transfected with MTFR2-WT (50 nM)
or MTFR2-MUT (50 nM) and Oe-HOXC10 (50 nM) or Oe-NC (50 nM) using
Lipofectamine 2000. Following incubation for 48 h at 37°C, cells
were collected and firefly and Renilla luciferase activities
were detected using a Dual-Luciferase Reporter Assay System
(Promega Corporation) according to the manufacturer's protocol.
Firefly luciferase was normalized to Renilla luciferase
activity.
Chromatin immunoprecipitation
(ChIP)
The interaction between HOXC10 and the MTFR2
promoter in HCT116 cells was also determined by ChIP. ChIP was
conducted using the ChIP Assay Kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. Protein
and DNA in HCT116 cells were cross-linked in 1% formaldehyde for 10
min at room temperature, extracted by SDS lysis buffer, and
separated by ultrasonication to obtain DNA fragments. The cell
lysate was then immunoprecipitated using anti-HOXC10 (1:500; cat.
no. ab153904; Abcam) or IgG (1:1,000; cat. no. 7074; Cell Signaling
Technology, Inc.) antibodies. Immunoprecipitated DNA was analyzed
via RT-qPCR.
Statistical analysis
All experiments were repeated independently in
triplicate. All statistical analyses were performed using GraphPad
Prism 8.0 (GraphPad Software, Inc.). Data are presented as the mean
± SD. Comparisons among two groups were analyzed using a paired or
unpaired Student's t-test, whereas comparisons among multiple
groups were analyzed using one-way ANOVA, followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
mRNA expression levels of HOXC10 and
MTFR2 are upregulated in CRC tissues
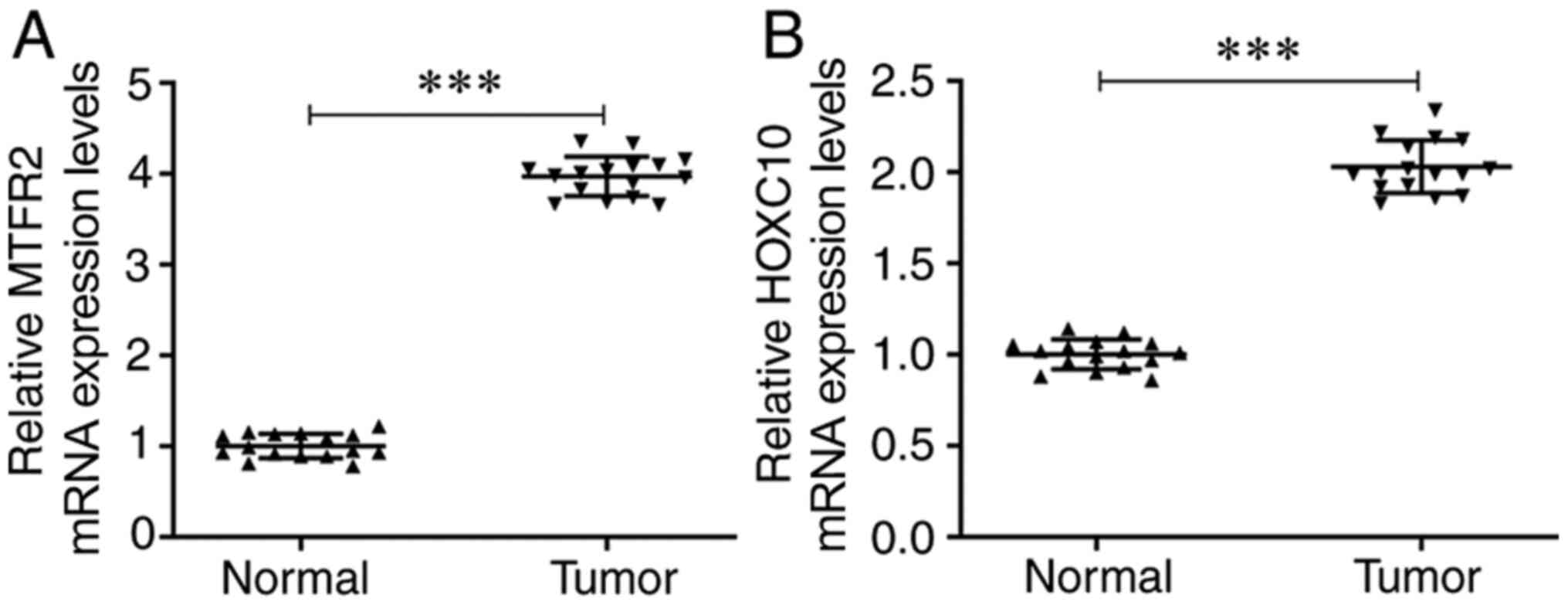
The mRNA expression levels of MTFR2 were
significantly increased in CRC tissues compared with those in the
matched normal adjacent tissues (Fig.
1A). Furthermore, the mRNA expression levels of HOXC10 were
also significantly upregulated in CRC tissues compared with those
in the matched adjacent tissues (Fig.
1B).
MTFR2 knockdown inhibits CRC cell
proliferation
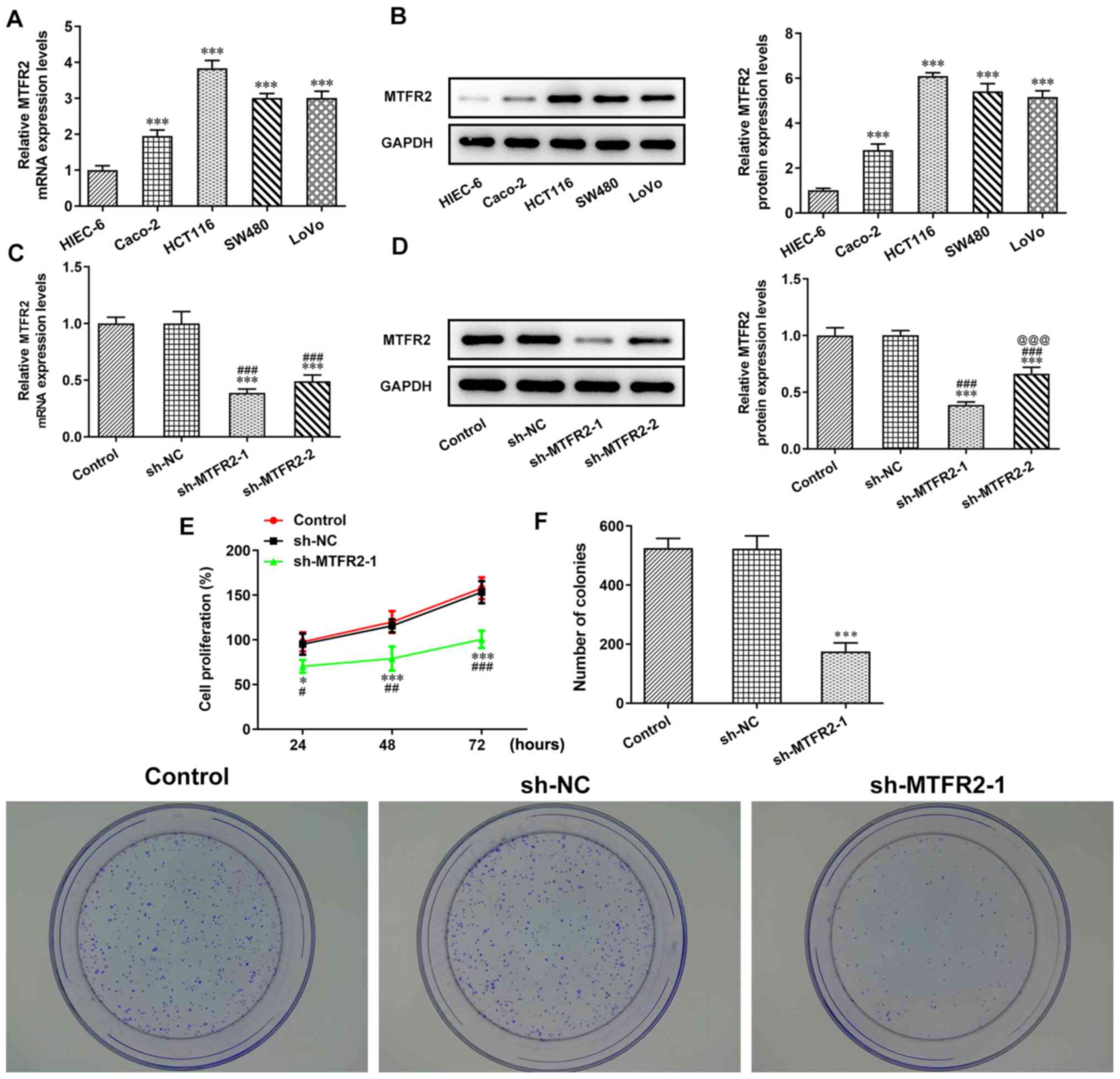
The mRNA and protein expression levels of MTFR2 in
the different CRC cell lines were significantly higher compared
with those in the HIEC-6 cells (Fig. 2A
and B). Notably, the MTFR2 mRNA and protein expression levels
were highest in HCT116 cells, which were therefore chosen for
subsequent experiments. When HCT116 cells were transfected with
sh-MTFR2-1/2, the mRNA and protein expression levels of MTFR2 were
significantly downregulated compared with those in the control and
sh-NC groups (Fig. 2C and D).
Furthermore, MTFR2 protein expression levels in the sh-MTFR2-1
group were significantly lower compared with those in the
sh-MTFR2-2 group, sh-MTFR2-1 was therefore used for subsequent
experiments. Proliferation and clone formation were also shown to
be significantly reduced in response to sh-MTFR2-1 compared with in
the sh-NC group (Fig. 2E and
F).
MTFR2 knockdown inhibits invasion and
migration of CRC cells
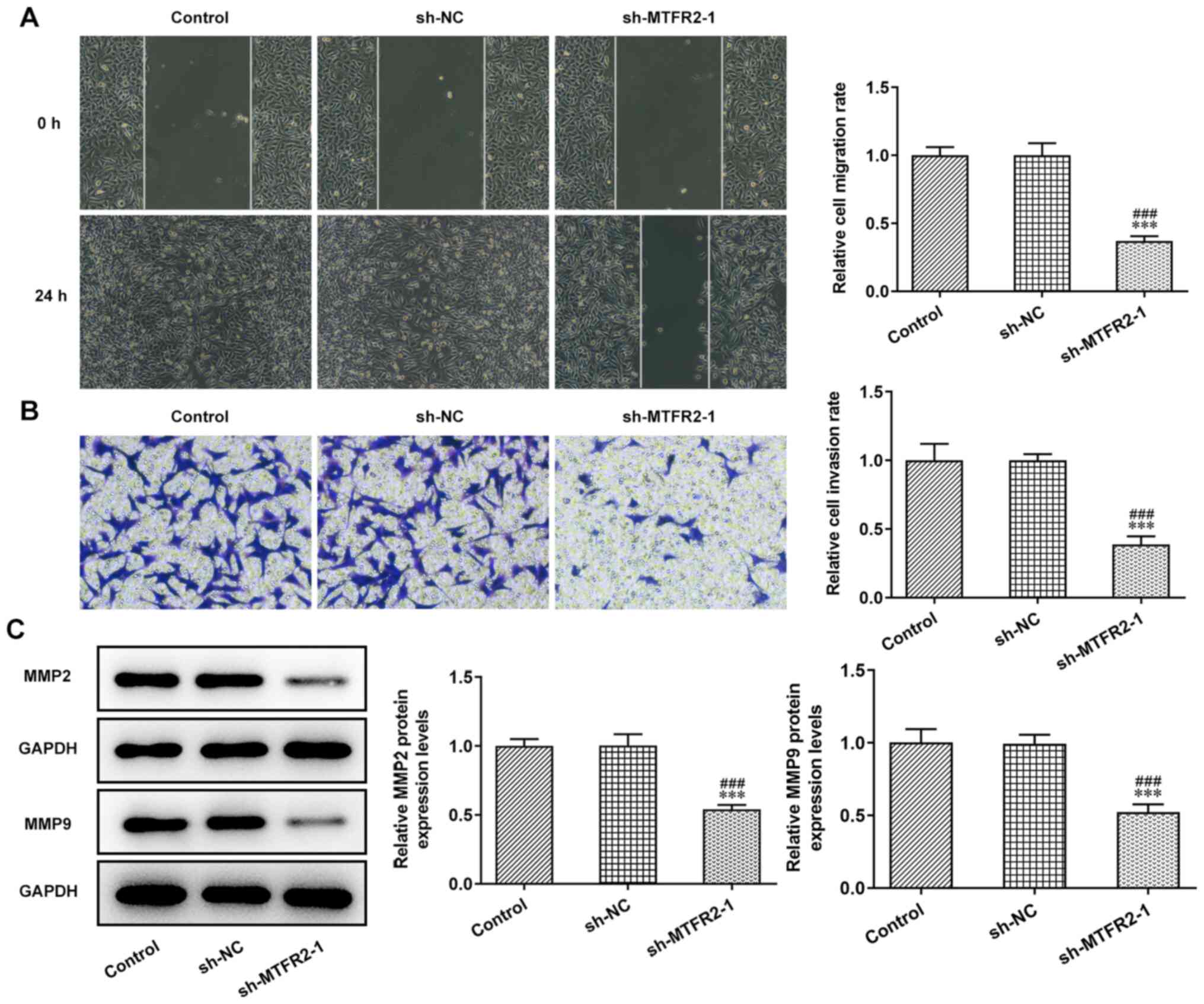
The invasion and migration of HCT116 cells were
significantly inhibited by sh-MTFR2-1 compared with in the control
and sh-NC groups (Fig. 3A and B).
Furthermore, knockdown of MTFR2 significantly downregulated the
protein expression levels of MMP2 and MMP9 in HCT116 cells compared
with those in the control and sh-NC groups (Fig. 3C).
HOXC10 is upregulated in CRC cells and
activates MTFR2 expression
HOXC10 mRNA and protein expression levels in HCT116
cells were significantly higher compared with those in HIEC-6 cells
(Fig. 4A and B). JASPAR predicted
the binding sites between HOXC10 and MTFR2 (Fig. 4C). In HCT116 cells transfected with
Oe-HOXC10, the mRNA and protein expression levels of HOXC10
(Fig. 4D and E) and MTFR2 (Fig. 4F and G) were significantly
upregulated compared with those in the control and Oe-NC groups.
Furthermore, the results demonstrated that relative luciferase
activity was increased in HCT116 cells co-transfected with MTFR2-WT
and Oe-HOXC10 compared with co-transfected with MTFR2-WT and Oe-NC,
while there was no significant change in MTFR2-MUT (Fig. 4H). The ChIP results showed a
significant increase in relative MTFR2 expression in anti-HOXC10
compared with IgG, which further confirmed that HOXC10 bound to
MTFR2 (Fig. 4I).
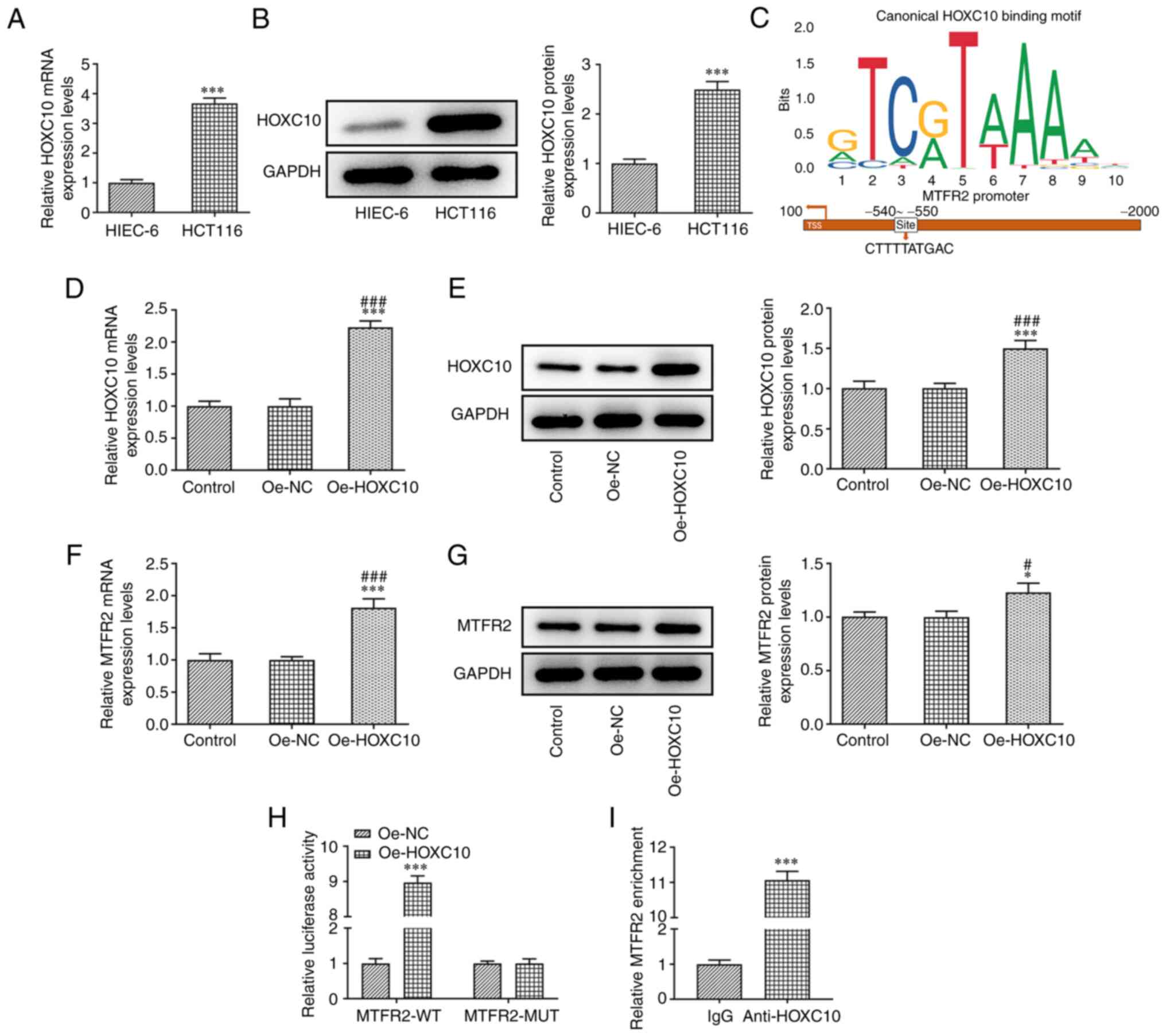 | Figure 4.HOXC10 is upregulated in colorectal
cancer cells, and binds to the MTFR2 promoter to activate MTFR2
expression. (A) mRNA and (B) protein expression levels of HOXC10 in
HCT116 and HIEC-6 cell lines were analyzed by RT-qPCR and western
blotting, respectively. ***P<0.001 vs. HIEC-6. (C) Binding sites
between HOXC10 and MTFR2 were determined using the Japan Automotive
Software Platform and Architecture program. (D) mRNA and (E)
protein expression levels of HOXC10 in HCT116 cells transfected
with Oe-HOXC10 were analyzed by RT-qPCR and western blotting,
respectively. (F) mRNA and (G) protein expression of MTFR2 in
HCT116 cells transfected with Oe-HOXC10 were analyzed by RT-qPCR
and western blotting, respectively. *P<0.05 and ***P<0.001
vs. control group; #P<0.05 and
###P<0.001 vs. Oe-NC group. (H) Relative luciferase
activity was determined using a dual-luciferase reporter assay in
HCT116 cells co-transfected with Oe-HOXC10 or Oe-NC and MTFR2-WT or
MTFR2-MUT. ***P<0.001 vs. Oe-NC group. (I) Chromatin
immunoprecipitation confirmed the interaction between HOXC10 and
MTFR2. ***P<0.001 vs. IgG group. MTFR2, mitochondrial fission
regulator 2; RT-qPCR, reverse transcription-quantitative PCR; Oe,
overexpression; NC, negative control; WT, wild-type; MUT,
mutant. |
Overexpression of HOXC10 partially
reverses the inhibitory effect of MTFR2 knockdown on CRC cell
proliferation and migration
MTFR2 knockdown significantly suppressed the
proliferation and clone formation of HCT116 cells compared with
sh-NC; however, this effect was significantly reduced by HOXC10
overexpression compared with in the sh-MTFR2-1 + Oe-NC group
(Fig. 5A and B). In the present
study, MTFR2 knockdown significantly inhibited HCT116 cell invasion
and migration (Fig. 5C and D), and
significantly reduced MMP2 and MMP9 protein compression levels
compared with in the sh-NC group (Fig.
5E); however, these effects were reversed by HOXC10
overexpression.
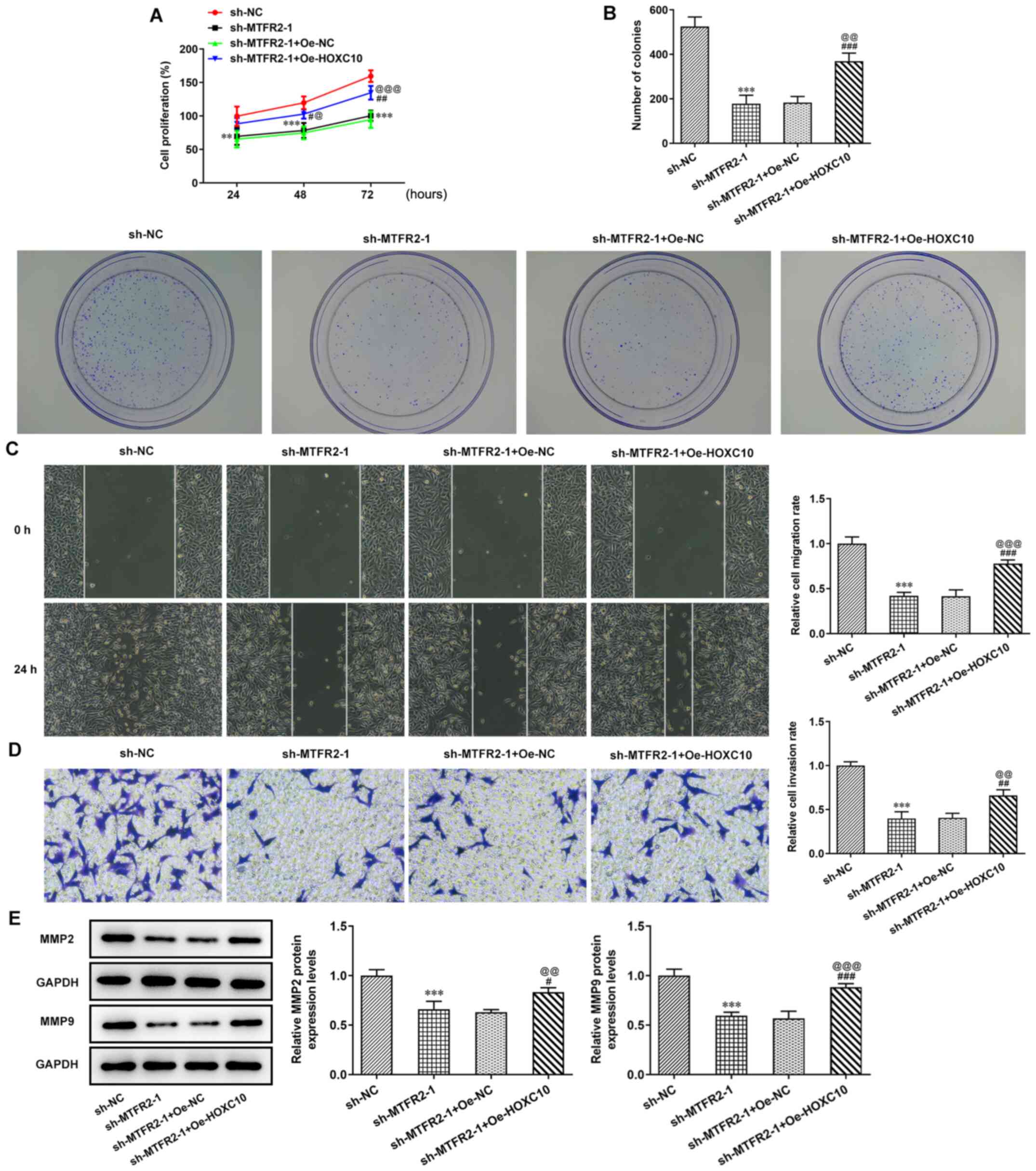 | Figure 5.Overexpression of HOXC10 partially
reverses the inhibitory effect of MTFR2 knockdown on colorectal
cancer cell proliferation and migration. (A) Proliferation, (B)
clone formation, (C) migration and (D) invasion of HCT116 cells
co-transfected with sh-MTFR2-1 and Oe-HOXC10 were in turn detected
by the Cell Counting Kit-8, clone formation, wound healing and
Transwell assays (magnification, ×100). Relative migration and
invasion rates were expressed normalized to the control group. (E)
Protein expression levels of invasion- and migration-related
proteins in HCT116 cells co-transfected with sh-MTFR2-1 and
Oe-HOXC10 were detected by western blotting. **P<0.01 and
***P<0.001 vs. sh-NC; #P<0.05,
##P<0.01 and ###P<0.001 vs. sh-MTFR2-1;
@P<0.05, @@P<0.01 and
@@@P<0.001 vs. sh-MTFR2-1 + Oe-NC. MTFR2,
mitochondrial fission regulator 2; sh, short hairpin RNA; Oe,
overexpression; NC, negative control. |
Discussion
CRC is a malignancy of the digestive system with
high morbidity and mortality rates; consequently, much focus has
been given to CRC at both the medical and societal levels (15). If patients with CRC can be diagnosed
and treated early there is a higher likelihood of a better
prognosis; therefore, early diagnosis and treatment is extremely
important (16). Colonoscopy is the
international gold standard for CRC diagnosis; however, it is
difficult to popularize this type of screening (17,18).
Therefore, it is necessary to find more appropriate means and
methods for the clinical diagnosis, prognosis, treatment effects
and postoperative survival prediction of CRC.
MTFR2 is located on chromosome 6q23.3 and serves a
key role in mitochondria, promoting mitochondrial division and
aerobic respiration in eukaryotic cells (5,6).
Previous studies have indicated that mitochondrial fission proteins
could promote the cell cycle, proliferation, invasion and migration
(19), and that mitochondrial
fission was related to apoptosis (20). In addition, recent studies have
demonstrated that MTFR2 expression was upregulated in BC and oral
squamous cell carcinoma, which was related to the aggravation of
invasion and migration (7–9). The present study indicated that MTFR2
expression was also significantly upregulated in CRC tissues and
cells. Furthermore, MTFR2 knockdown could suppress the
proliferation, invasion and migration of CRC cells.
In terms of the depth of infiltration, activation of
MMP2 and MMP9 can promote the invasion of tumor cells, and are
closely related to malignant phenotypes (21). Infiltration has been reported to be
positively correlated with MMP2; with the increase in depth of
infiltration, the positive expression intensity and expression rate
of MMP2 were increased (22). It
has also been reported that MMP9 was upregulated in colon cancer
tumors and maintained high expression levels, suggesting that it
may be involved in later invasion and metastasis (23). In the present study, MTFR2 knockdown
significantly decreased MMP2 and MMP9 protein expression, but
HOXC10 overexpression reversed these effects.
HOXC10 is an important member of the HOX family, a
group of evolutionarily conserved genes that control cell
proliferation, differentiation and embryo development (24). It has previously been reported that
the HOX gene family serves an important role in tumorigenesis and
development. In cervical squamous cell carcinoma, the elevation of
HOXC10 was determined to be correlated with increased invasion
(25). Data from The Cancer Genome
Atlas demonstrated that HOXC10 expression was significantly
increased in human thyroid cancer tissues compared with in healthy
thyroid tissues (26). Furthermore,
HOXC10 promoted the migration and invasion of thyroid cancer cells,
suggesting that HOXC10 may be a novel biomarker for the prognosis
of human thyroid cancer (26).
HOXC10 also promoted metastasis of human lung adenocarcinoma and
indicated poor survival outcomes (27). Migration and invasion are important
for tumor cell circulation and establishment of distant metastases
(28). Previous studies
demonstrated that HOXC10 overexpression promoted proliferation,
inhibited apoptosis, and promoted metastasis and invasion of GC
cells (12,29). The present study was consistent with
previous research, whereby HOXC10 was highly expressed in CRC
cancer cells (HCT116) compared with in healthy cells (HIEC-6).
Moreover, HOXC10 overexpression attenuated the effects of MTFR2
knockdown to promote the proliferation, invasion and migration of
CRC cells.
A recent study reported that increased expression of
HOXC10 conferred resistance to radiotherapy and chemotherapy in
esophageal squamous cell carcinoma (ESCC) tumor cells, and
predicted the poor prognosis of patients with ESCC (30). In GC, knockdown of HOXC10 made GC
cells more sensitive to apatinib, solving problems with apatinib
resistance in the treatment of GC (31). Tang et al (27) indicated that HOXC10 expression was
upregulated in lung cancer tissues, especially in tissues with
metastatic potential, and predicted a poor prognosis of patients
with lung cancer. HOXC10 expression has also been reported to be
overexpressed in human thyroid cancer tissues, which may be
positively related to a poor prognosis (26). In future work, the correlation
analyses of HOXC10 expression with the prognosis of patients with
CRC, especially those receiving adjuvant chemoradiotherapy, will be
investigated. Furthermore, the results obtained in the present
study will be confirmed in animal studies.
In conclusion, the present study demonstrated that
HOXC10 overexpression activated the expression of MTFR2 to enhance
the proliferation, clone formation, invasion and migration of CRC
cells. HOXC10 may therefore be considered an ideal therapeutic
target for CRC. However, the study did not perform an animal model,
which was a limitation of this study.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YX, QW and GL conceived and designed the study. RC,
LY and ZJ were responsible for the acquisition, analysis and
interpretation of data. YX, RC and GL was responsible for
manuscript preparation, writing and critical revisions. YX and RC
confirm the authenticity of all the raw data. All authors have read
and approved the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Clinical
Research Ethics Committee of Nanjing Medical University (Nanjing,
China; approval no. 2021-448) and informed consent was obtained
from each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arnold M, Sierra MS, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global patterns and trends in
colorectal cancer incidence and mortality. Gut. 66:683–691. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yin J, Bai Z, Zhang J, Zheng Z, Yao H, Ye
P, Li J, Gao X and Zhang Z: Burden of colorectal cancer in China,
1990–2017: Findings from the Global Burden of disease study 2017.
Chin J Cancer Res. 31:489–498. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Clough E and Barrett T: The gene
expression omnibus database. Methods Mol Biol. 1418:93–110. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Monticone M, Panfoli I, Ravera S, Puglisi
R, Jiang MM, Morello R, Candiani S, Tonachini L, Biticchi R,
Fabiano A, et al: The nuclear genes Mtfr1 and Dufd1 regulate
mitochondrial dynamic and cellular respiration. J Cell Physiol.
225:767–776. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang J, Xie Y, Bai X, Wang N, Yu H, Deng
Z, Lian M, Yu S, Liu H, Xie W and Wang M: Targeting dual
specificity protein kinase TTK attenuates tumorigenesis of
glioblastoma. Oncotarget. 9:3081–3088. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang W, Xiong M, Jiang L, Chen Z and Shao
Y: MTFR2 promotes the proliferation, migration, and invasion of
oral squamous carcinoma by switching OXPHOS to glycolysis. Front
Oncol. 10:8582020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu W, Zang R, Du Y, Li X, Li H, Liu C,
Song Y, Li Y and Wang Y: Overexpression of MTFR2 predicts poor
prognosis of breast cancer. Cancer Manag Res. 12:11095–11102. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu G, Lai Y, Wang T, Lin W, Lu J, Ma Y,
Chen Y, Ma H, Liu R and Li J: Mitochondrial fission regulator 2
(MTFR2) promotes growth, migration, invasion and tumour progression
in breast cancer cells. Aging (Albany NY). 11:10203–10219. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Enteghami M, Ghorbani M, Zamani M and
Galehdari H: HOXC10 is significantly overexpressed in colorectal
cancer. Biomed Rep. 13:182020.PubMed/NCBI
|
|
11
|
Peng Y, Li Y, Li Y, Wu A, Fan L, Huang W,
Fu C, Deng Z, Wang K, Zhang Y, et al: HOXC10 promotes tumour
metastasis by regulating the EMT-related gene Slug in ovarian
cancer. Aging (Albany NY). 12:19375–19398. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li J, Tong G, Huang C, Luo Y, Wang S,
Zhang Y, Cheng B, Zhang Z, Wu X, Liu Q, et al: HOXC10 promotes cell
migration, invasion, and tumor growth in gastric carcinoma cells
through upregulating proinflammatory cytokines. J Cell Physiol.
235:3579–3591. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guan Y, He Y, Lv S, Hou X, Li L and Song
J: Overexpression of HOXC10 promotes glioblastoma cell progression
to a poor prognosis via the PI3K/AKT signalling pathway. J Drug
Target. 27:60–66. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang PS and Shaukat A: Assessing the
impact of lowering the colorectal cancer screening age to 45 years.
Lancet Gastroenterol Hepatol. 5:523–524. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Song WY, Zhang X, Zhang Q, Zhang PJ and
Zhang R: Clinical value evaluation of serum markers for early
diagnosis of colorectal cancer. World J Gastrointest Oncol.
12:219–227. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang K, Xu H and Li HT: Safety and
efficacy of propofol alone or in combination with other agents for
sedation of patients undergoing colonoscopy: An updated
meta-analysis. Eur Rev Med Pharmacol Sci. 24:4506–4518.
2020.PubMed/NCBI
|
|
18
|
Park SW, Shin SP and Hong JT: Efficacy and
tolerability of prucalopride in bowel preparation for colonoscopy:
A systematic review and Meta-analysis. Adv Ther. 37:2507–2519.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lima AR, Santos L, Correia M, Soares P,
Sobrinho-Simões M, Melo M and Máximo V: Dynamin-related protein 1
at the crossroads of cancer. Genes (Basel). 9:1152018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sheridan C and Martin SJ: Mitochondrial
fission/fusion dynamics and apoptosis. Mitochondrion. 10:640–648.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu D, Duan W, Guo H, Xu X and Bai Y:
Meta-analysis of associations between polymorphisms in the promoter
regions of matrix metalloproteinases and the risk of colorectal
cancer. Int J Colorectal Dis. 26:1099–1105. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong W, Li H, Zhang Y, Yang H, Guo M, Li L
and Liu T: Matrix metalloproteinase 2 promotes cell growth and
invasion in colorectal cancer. Acta Biochim Biophys Sin (Shanghai).
43:840–848. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Morán A, Iniesta P, García-Aranda C, De
Juan C, Díaz-López A, Sánchez-Pernaute A, Torres AJ, Díaz-Rubio E,
Balibrea JL and Benito M: Clinical relevance of MMP-9, MMP-2,
TIMP-1 and TIMP-2 in colorectal cancer. Oncol Rep. 13:115–120.
2005.PubMed/NCBI
|
|
24
|
McGinnis W and Krumlauf R: Homeobox genes
and axial patterning. Cell. 68:283–302. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhai Y, Kuick R, Nan B, Ota I, Weiss SJ,
Trimble CL, Fearon ER and Cho KR: Gene expression analysis of
preinvasive and invasive cervical squamous cell carcinomas
identifies HOXC10 as a key mediator of invasion. Cancer Res.
67:10163–10172. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng X, Li T, Liu Z, Shi Y and Peng Y:
HOXC10 up-regulation contributes to human thyroid cancer and
indicates poor survival outcome. Mol Biosyst. 11:2946–2954. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tang XL, Ding BX, Hua Y, Chen H, Wu T,
Chen ZQ and Yuan CH: HOXC10 promotes the metastasis of human lung
adenocarcinoma and indicates poor survival outcome. Front Physiol.
8:5572017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li S, Zhang W, Wu C, Gao H, Yu J, Wang X,
Li B, Jun Z, Zhang W, Zhou P, et al: HOXC10 promotes proliferation
and invasion and induces immunosuppressive gene expression in
glioma. FEBS J. 285:2278–2291. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zheng J, Ge P, Liu X, Wei J, Wu G and Li
X: miR-136 inhibits gastric cancer-specific peritoneal metastasis
by targeting HOXC10. Tumour Biol. 39:10104283177062072017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Suo D, Wang Z, Li L, Chen Q, Zeng T, Liu
R, Yun J, Guan XY and Li Y: HOXC10 upregulation confers resistance
to chemoradiotherapy in ESCC tumor cells and predicts poor
prognosis. Oncogene. 39:5441–5454. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yu J, Zhang X, Ma Y, Li Z, Tao R, Chen W,
Xiong S and Han X: miR-129-5p restrains apatinib resistance in
human gastric cancer cells via downregulating HOXC10. Cancer
Biother Radiopharm. 36:95–105. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jin M and Frankel WL: Lymph node
metastasis in colorectal cancer. Surg Oncol Clin N Am. 27:401–412.
2018. View Article : Google Scholar : PubMed/NCBI
|