Introduction
Septicemia remains an all too common pathology that
is associated with multiple abnormalities and heterogeneous
responses, but particularly with cardiovascular dysfunction
(1). Due to a lack of effective
interventions, sepsis is one of the primary causes of high
morbidity and mortality rates worldwide, with confirmed mortality
rates of ≤47% in patients with sepsis (2). Despite efforts to improve diagnosis
and treatment, sepsis remains a major medical challenge. Cardiac
dysfunction is considered to be a widely accepted manifestation of
sepsis, which is closely related to sepsis-associated dilemma.
Studies have indicated that 40–50% of patients with sepsis exhibit
varying degrees of myocardial dysfunction, with a mortality rate of
70–90% in those with myocardial injury, compared with 20% in those
without it (3,4). Therefore, the determination of novel
interventions for sepsis-induced myocardial injury (SIMI) is an
urgent requirement for reducing the morbidity and mortality rates
of individuals with sepsis.
MicroRNAs (miRNAs/miRs) are small endogenous
non-coding single-stranded RNAs (~22 nucleotides in length), which
are transcribed and transported to the cytoplasm from the nucleus
(5). miRNAs negatively regulate
gene expression by interacting with the 3′untranslated regions of
target mRNAs, resulting in mRNA inhibition or degradation (6). It has been demonstrated that miRNAs
may be involved in the regulation of almost all major cellular
functions, including development, differentiation, proliferation
and apoptosis (7). Previous studies
have shown that miRNAs are also associated with a diverse array of
disorders, including sepsis, heart disease and diabetes (8–10).
Studies have confirmed that the dysregulation of multiple miRNAs
during sepsis (including miRNA-146a, miRNA-499-5p, miRNA-297,
miRNA-574-5p, miRNA-122 and miRNA-133a) accelerates
sepsis-associated disorders (11–15).
Studies have also shown that loss of the miRNA-144/451 cluster
impairs ischemic preconditioning-mediated cardio-protection
(16–18). Furthermore, overexpression of
miRNA-494 has a protective effect on ischemia/reperfusion-induced
cardiac injuries, while elevation of miRNA-320 has a detrimental
effect (19–21). miR-214 overexpression also protects
against SIMI (22) and the same
results obtained by Ge et al (22) also confirmed that miR-214
overexpression showed ameliorative effect against SIMI (data not
published) and the protective effects miRNA-335 on
H2O2-induced cardiomyocyte injury have been
demonstrated (23). However,
whether miRNA-335 expression is also associated with SIMI remains
to be elucidated. As myocardial damage is the leading cause of
sepsis-related mortality, the present study aimed to determine the
association between miRNA-335 upregulation and SIMI and the effect
of miRNA-335 on the progression of SIMI.
Materials and methods
Animals
All protocols were approved by the Medical Animal
Ethics Committee of Hunan Academy of Chinese Medicine (Xiang
20200015). All animals were handled in strict accordance with good
animal practice as defined by the Hunan Academy of Chinese Medicine
animal welfare bodies. Healthy male Kunming mice (age 7–8 weeks;
weight, 25–30 g; n=100) purchased from the Experimental Animal
Center of Central South University were kept under standardized
laboratory conditions of a maintained on a 12-h light/dark cycle,
temperature-controlled environment. Mice were housed 4–5 per cage
in an enriched environment with an ambient temperature of 22±1°C
and humidity of 50±5% for with ad libitum access to food and
water for 1 week before the experiments started. All of the mice
were randomly divided (15 mice per group) into 6 groups: Normal
group (NC); Sham group; CLP group (cecal ligation and puncture);
pre-miR-335 group; anti-miR-335 group and wild-type group (WT). The
experiment continued for one week and all surgical procedures were
performed under anesthesia with an initial intraperitoneal
injection of sodium pentobarbital (30 mg/kg body weight) to
minimize animal suffering until they lost consciousness. All the
animals were sacrificed following anesthesia by exsanguination
(24,25). During all the experiments, the care
and treatment of the animals received prior institutional approval
from the Ethical Commission on Animal Research of the Hunan Academy
of Chinese Medicine.
Establishment of septicemia
Sepsis was established by cecal ligation and
puncture (CLP) as previously described (26) and with slight modifications.
Following anesthesia with an intraperitoneal injection of sodium
pentobarbital (30 mg/kg) to minimize animal suffering, the cecum
was exposed via a 1–2 cm midline incision across the abdomen, the
cecum was exposed via a 1–2 cm midline incision across the abdomen.
The cecum was ligated with a 5-0 sterile silk thread just below the
ileocecal valve. The area between the ligation point and the tip of
the cecum was punctured twice with a 18-gauge needle (with a 1-cm
drainage tube attached). A small amount of feces was gently
squeezed from the cecum, which was then returned to its anatomical
position. Warm saline (0.05 ml/g) was then subcutaneously injected
into each mouse. Mice in the Sham group underwent cecum exposure
only.
Determination of cardiac function by
Langendorff perfusion
Mice were anesthetized with an intraperitoneal
injection of sodium pentobarbital (30 mg/kg) to minimize animal
suffering at 3, 6, 12 and 24 h post-sham or -CLP surgery. A
midsternal sternotomy was performed and the heart was mounted on a
Langendorff isolated cardiac perfusion system. The aorta was
intubated with a 30-gauge cannula and perfusion was initiated by
intubating in retrograde mode. The perfusion solution consisted of
7 mmol/l glucose, 0.4 mmol/l oleate, 1% BSA and a low concentration
of insulin (10 µU/ml). Pressure was monitored by inserting a
water-filled latex balloon into the left ventricle, which was
connected to a pressure sensor. The balloon volume was adjusted to
a constant baseline left ventricular end-diastolic pressure of 5 mm
Hg for 15 min. The signal acquisition system (cat. no. RM6240B;
Chengdu Biological Instruments) was used to record the left
ventricular development pressure (LVDP), maximum pressure increase
rate (+dp/dt), maximum pressure reduction rate (-dp/dt), heart
rate, cardiac output and stroke output.
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
TRIzol® (Thermo Fisher Scientific, Inc.)
was used to isolate total RNA from the heart tissues of mice that
underwent sham or CLP surgery at 6, 12 and 24 h. RNA purity was
determined using a microplate reader at a wavelength of 260 nm.
Cardiac mRNA expression levels were determined by RT-qPCR (Applied
Biosystems; Thermo Fisher Scientific, Inc.). First, the extracted
total RNA was purified with 75% ethanol and its concentration was
determined by spectrophotometry. The RNA (200 ng per sample) was
then used as a template for reverse transcription using the
PrimeScript® RT reagent kit (cat. no. RP1201, BioTeke
Corporation) with U6 as the internal standardized control. In the
reaction system with a total amount of 10 µl, 2X SYBR-Green Mixture
5.0, 0.5 µl 5 µm/l positive and reverse primers, 1 µl cDNA and
ddH2O replenishment volume to 10.0 µl. The conditions of
reverse transcription reaction were: 50°C for 15 min, 85°C for 5
min. Reaction conditions were denaturation at 95°C for 5 min, 95°C
for 15 sec, 60°C for 1 min, for 40 cycles and were performed on
Bio-Rad CFX96 Real-time Quantitative Fluorescence PCR (Bio-Rad
Laboratories, Inc.). The following primers were provided by
GeneCopoeia Inc.: miR-335 forward, 5′-GCGGTCAAGAGCAATAACGAA−3′ and
reverse, 5′-GTGCAGGGTCCGAGGTATTC-3′; U6 (internal control) forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′. To
determine a ΔCt value, the Ct value of the target gene was
normalized by subtracting the U6 Ct value. The relative expression
level between treatments was then calculated using the following
equation: relative gene expression=2-(ΔCt sample-ΔCt control)
(27).
Transfection and survival
Transfection and survival were performed as
previously described (28–31). Mice were intravenously injected with
miR-335 precursor (5′-UCAAGAGCAAUAACGAAAAAUGU-3′) or inhibitor
(5′-ACAUUUUUCGUUAUUGCUCUUGA-3′; purchased from Guangzhou RiboBio
Co., Ltd.) and corresponding controls (negative control:
5′-UUCUCCGAACGUGUCACGUTT−3′; negative control inhibitor
5′-CAGUACUUUUGUGUAGUACAA-3′; Thermo Fisher Scientific, Inc.) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at a final concentration of 50 nM according to
the manufacturer's protocols and transfection efficiency was
assessed after 12 h. CLP was performed 4 days post-transfection.
For the wild-type (WT), pre-miR-335 negative control, anti-miR-335
negative control, pre-miR-335 and anti-miR-335 mice (10 mice per
group), survival rates were observed every 2 h for 36 h.
The mice were transfected with miR-335 precursor or
anti-miR-335 using adenovirus (2×1011 pfu; GenePharma,
Shanghai, China) for 4 days according to the manufacturer's
protocol. The mice were anesthetized and 200 µl adenovirus was
injected into tail veins.
Detection of inflammatory and
diagnostic markers
The mice were anesthetized intraperitoneally with an
intraperitoneal injection of sodium pentobarbital (30 mg/kg) to
minimize animal suffering. Retro-orbital puncture was performed to
take a 100 µl blood sample with a Brand micropipette, where the eye
was pulled out to collect at ≥500 µl blood. At 3, 6, 12 and 24 h
post-sham or -CLP surgery, the animals were anesthetized with an
intraperitoneal injection of sodium pentobarbital (30 mg/kg) till
loss of consciousness and sacrificed following anesthesia by
exsanguination. The heart tissues were collected and homogenized.
The blood samples and tissue homogenates were centrifuged at 500 ×
g for 10 min at 4°C and the supernatants were collected for storage
at −80°C. The levels of TNF-α, IL-6 and IL-10 and troponin (cTNI;
cat. no. MAB3152; Sigma-Aldrich; Merck KGaA), brain natriuretic
peptide (BNP; cat. no. AF6336; Sigma-Aldrich; Merck KGaA), creatine
kinase (CK; cat. no. MAK116; Sigma-Aldrich; Merck KGaA), lactate
dehydrogenase (LDH; cat. no. MAK066; Sigma-Aldrich; Merck KGaA) and
aspartate aminotransferase (AST; cat. no. MAK055; Sigma-Aldrich:
Merck KGaA) were determined using commercially available kits.
Flow cytometric detection of
apoptosis
Heart tissues were collected from the WT,
pre-miR-335 and anti-miR-335 mice 12 h post-CLP. The animals were
anesthetized with an initial intraperitoneal injection of sodium
pentobarbital (30 mg/kg) prior to exsanguination till loss of
consciousness (24,25). Then the chest cavity was opened and
the heart was removed and washed with 0°C calcium solution (140 mM
NaCl, 1 mM CaCl2, 1 mM MgCl2, 0.33 mM
NaH2PO4, 10 mM glucose, and 10 mM HEPES, pH
7.2). The aorta was isolated and filled with calcium-free solution
(140 mM NaCl, 1 mM CaCl2, 0.33 mM
NaH2PO4, 10 mM glucose, and 10 mM HEPES, pH
7.2) until the heart stopped beating. The heart was then placed in
a pre-prepared solution of collagen and albumin (4°C), after which
the tissues were minced and digested in KB solution for 1 h (4°C)
(32). A cardiomyocyte sample was
collected from one mouse in each group and an elemental suspension
was prepared. Following the protocols of Annexin V-fluorescein
isothiocyanate (FITC) apoptosis detection kit (cat. no. C1065;
Beyotime Institute of Biotechnology), Annexin V-FITC, propidium
iodide (PI), hydroxyethyl piperazine ethylsulfonic acid buffer was
added to Annexin V-FITC/PI staining solution in the proportion of
1:2:50. Cells at a concentration of 1×106 cells/100 µl
of the staining solution were resuspended, incubated at ambient
temperature for 15 min in the dark and added with 1 ml of
4-(2-hydroxyethyl)-1-piperazineëthanesulfonic acid buffer and then
analyzed using BD Accuri C6 software (version 5.0; BD Biosciences).
A flow cytometer (BD Accuri C6; BD Biosciences) was employed to
determine the excitation wavelength at 488 nm. The excitation
wavelength at 525 or 620 nm was employed to detect FITC or PI
fluorescence for cell apoptosis, respectively. The samples (n=10)
were randomly selected in each group. The apoptotic rate was
calculated as the percentage of early apoptotic cells, or the
percentage of late apoptotic cells.
Histological examination
At 24 h post-CLP, all animals received cardiac
perfusion under anesthesia. The heart specimens were then fixed in
4% paraformaldehyde for 24 h at room temperature, dehydrated,
cleared and embedded in paraffin. A microtome was used to cut the
specimens into 5-µm sections and each section was stained with
hematoxylin and eosin at room temperature for 5 min. Changes in the
myocardial tissue structure were observed under an light microscope
(magnification, ×400).
Statistical analysis
SPSS 21.0 (IBM Corp) was used for statistical
analysis. All data are presented as the mean ± standard deviation.
Measurements at single time point were performed by the Student's
unpaired t-test. One-way analysis of variance was used for
comparison between multiple groups, followed by Tukey's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
CLP accelerates myocardial injury in
mice
Compared with the Sham group, CLP led to reduced
animal activity and slow response to stimuli with increasing time,
as well as a decreased appetite. In a pre-study experiment, the
mortality rates of the septic mice following CLP were 50% at 48 h,
rising to 80% at 72 h and 90% at 96 h. Over time, cardiac
functions, including heart rate, cardiac output and stroke output,
showed an increase and then decreased. Additionally, cardiac
evaluations from the perfused CLP-mice showed that LVDP, +dP/dt and
-dP/dt were significantly reduced compared with those of the WT
group. By contrast, inflammatory factors (TNF-α, IL-6 and IL-10)
and myocardial enzyme indicators (cTNI, BNP, AST, LDH and CK) were
elevated in the septic mice at various time points (Fig. 1; P<0.05).
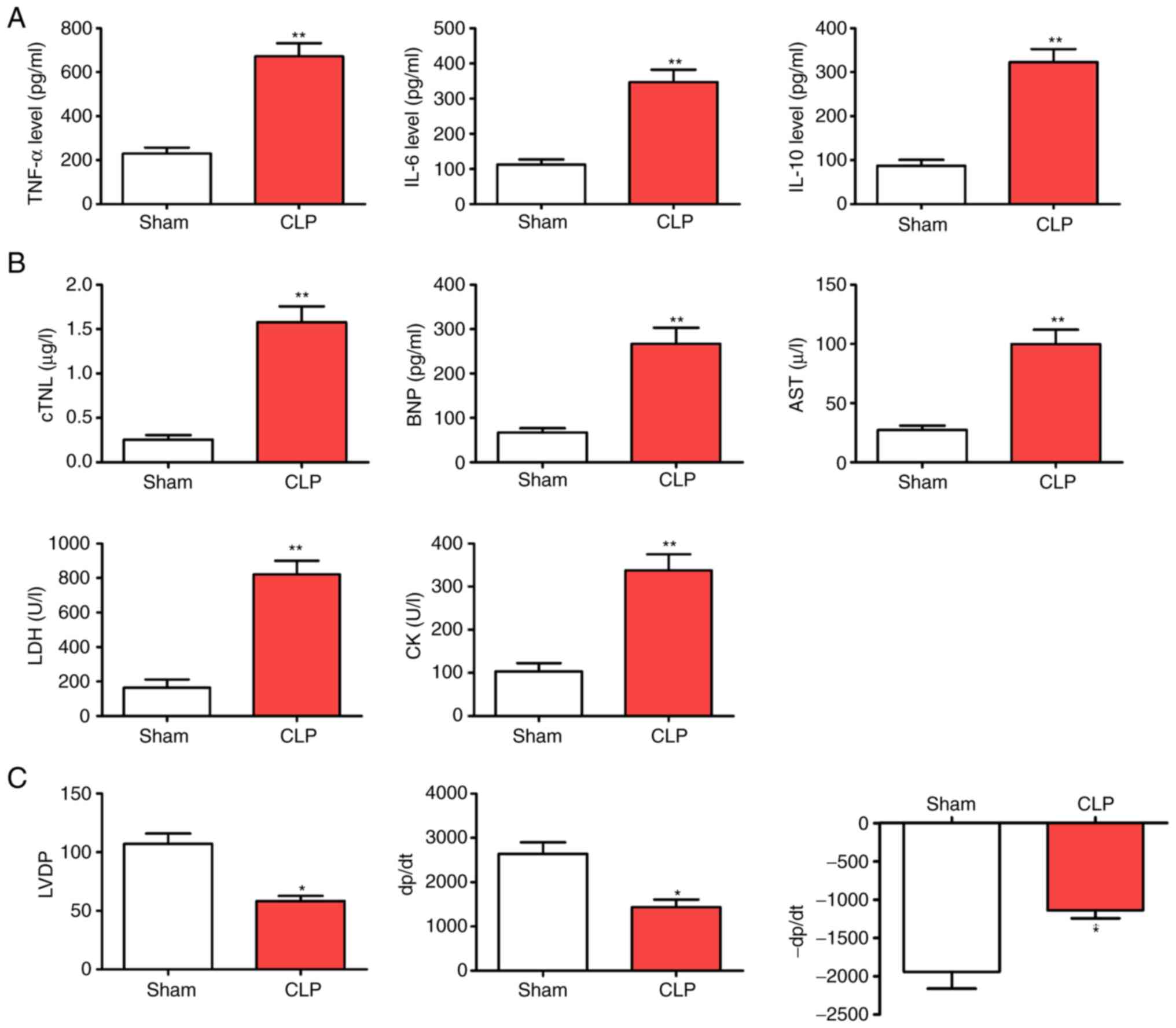 | Figure 1.CLP accelerates myocardial injury
following CLP surgery. (A) Levels of inflammatory factors (TNF-α,
IL-6 and IL-10). (B) Myocardial enzyme indicators (cTNI, BNP, AST,
LDH and CK). (C) Alterations in cardiac function (LVDP, +dP/dt and
-dP/dt). *P<0.05 and **P<0.01 vs. the Sham group; n=5. CLP,
cecal ligation and puncture; cTNI, troponin; BNP, brain natriuretic
peptide; AST, aspartate aminotransferase; LDH, lactate
dehydrogenase; CK creatine kinase; LVDP, left ventricular
development pressure; +dp/dt, maximum pressure increase rate;
-dp/dt maximum pressure reduction rate. |
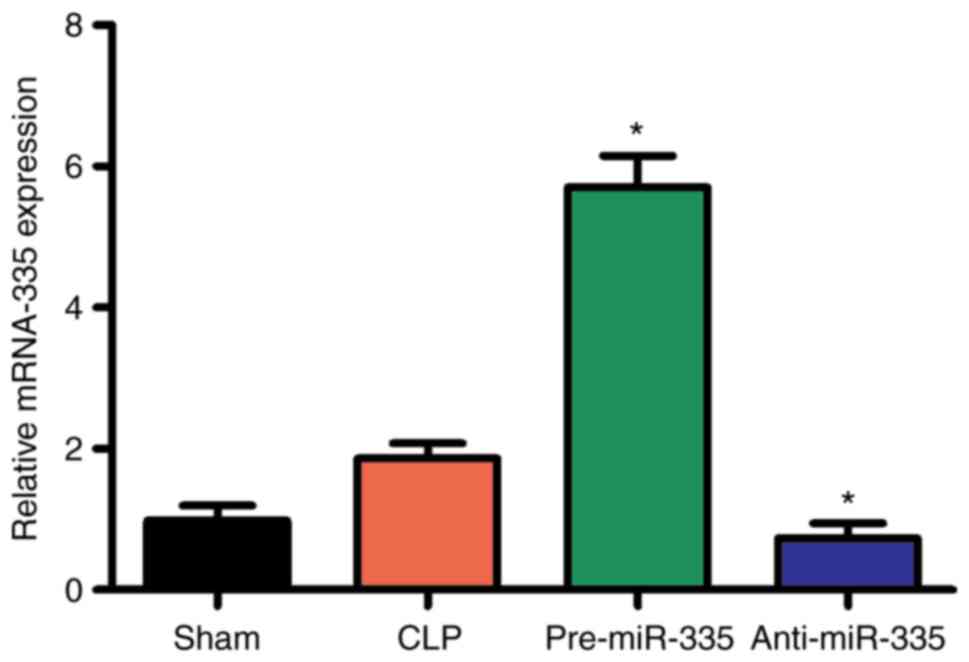
miR-335 is upregulated in myocardial
tissues following septic assault
Differential expression of multiple different miRNAs
has been reported in diseases of the heart (33). In particular, the expression of
miR-335 has been shown to significantly increase, which has been
demonstrated to protect cardiomyocytes from oxidative stress
(34,35). Therefore, it was hypothesized that
miR-335 may also be involved in SIMI. To confirm this hypothesis,
RT-qPCR was used to detect the expression of miR-335 at 6, 12 and
24 h post-CLP. The results showed that the expression of miR-335 in
the myocardial tissues of septic mice was significantly higher than
that of the sham-operated mice at 6, 12 and 24 h (Fig. 2; P<0.05).
Regulation of miR-335 expression in
mouse cardiac tissues
miR overexpression and silencing were performed to
further investigate the expression of miR-335 in cardiac tissue.
Preliminary experiments showed that miR-335 transfection efficiency
is highly effective (Fig. S1).
Compared with the CLP group, miR-335 expression was increased in
heart tissues after transfection with miR-335 precursors peaked at
12 h and then decreased after 24 h (Figs. 3 and S2). By contrast, the miR-335 inhibitor
caused a decrease in miR-335 expression. The effects of the
precursors and inhibitor were both specific to miR-335 and no
off-target effects on other miRNAs were observed. In addition, all
transfected mice were healthy and without significant cardiac
dysfunction (Fig. 3).
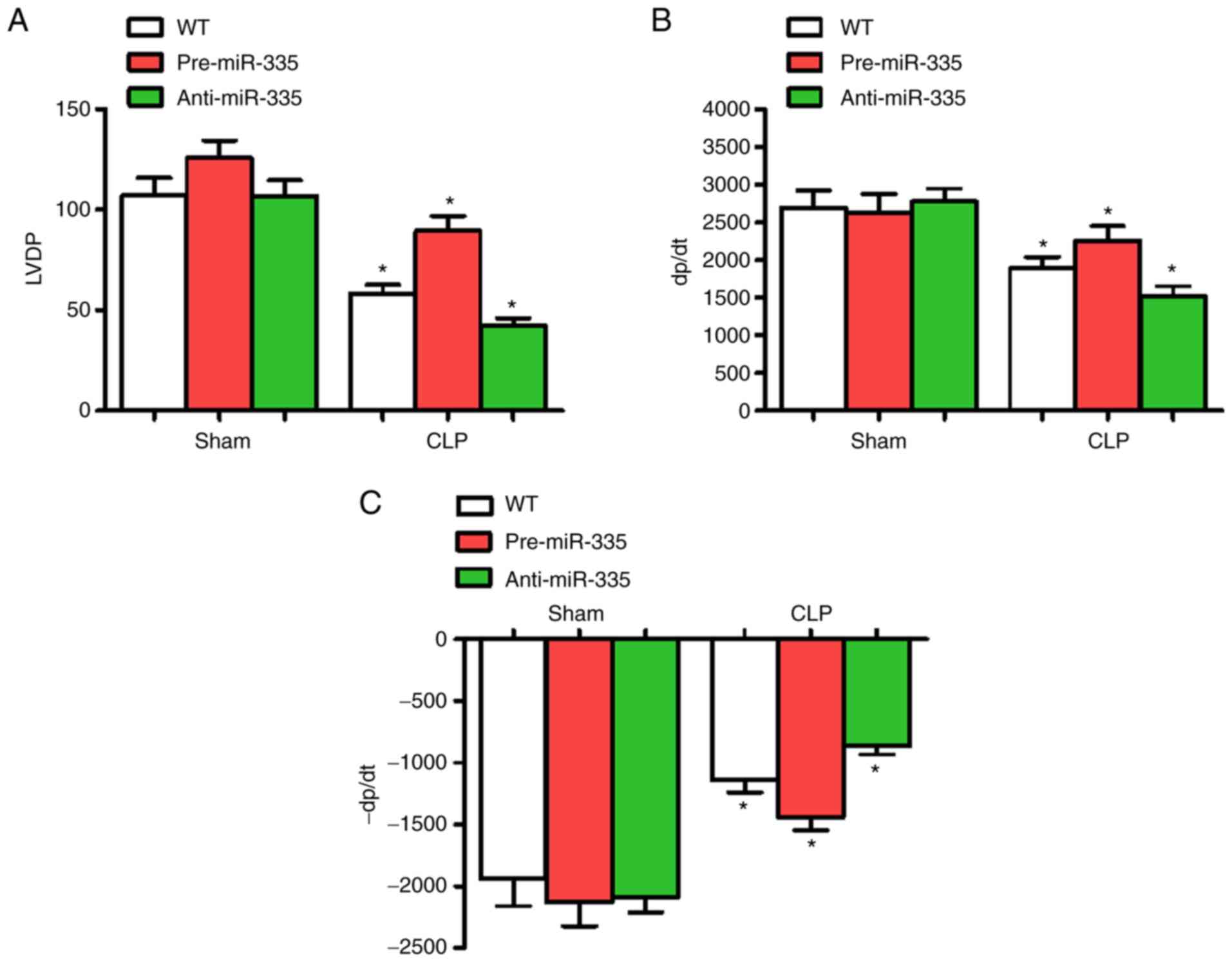
Effect of miR-335 on CLP-induced
myocardial dysfunction
At 6 h post-CLP, LVDP, +dP/dt and -dP/dt were
significantly altered, thus cardiac functions at 6 h were
determined in each group. Compared with the WT group, the
administration of miRNA-335 precursors 6 h post-CLP significantly
improved cardiac LVDP and ±dP/dt (n=6; P<0.05), while
anti-miR-335-pretreated mice experienced a decline in these
parameters (n=6; P<0.05). Compared with the WT group, miR-335
expression level had no significant effect on cardiac contractile
function (Fig. 4).
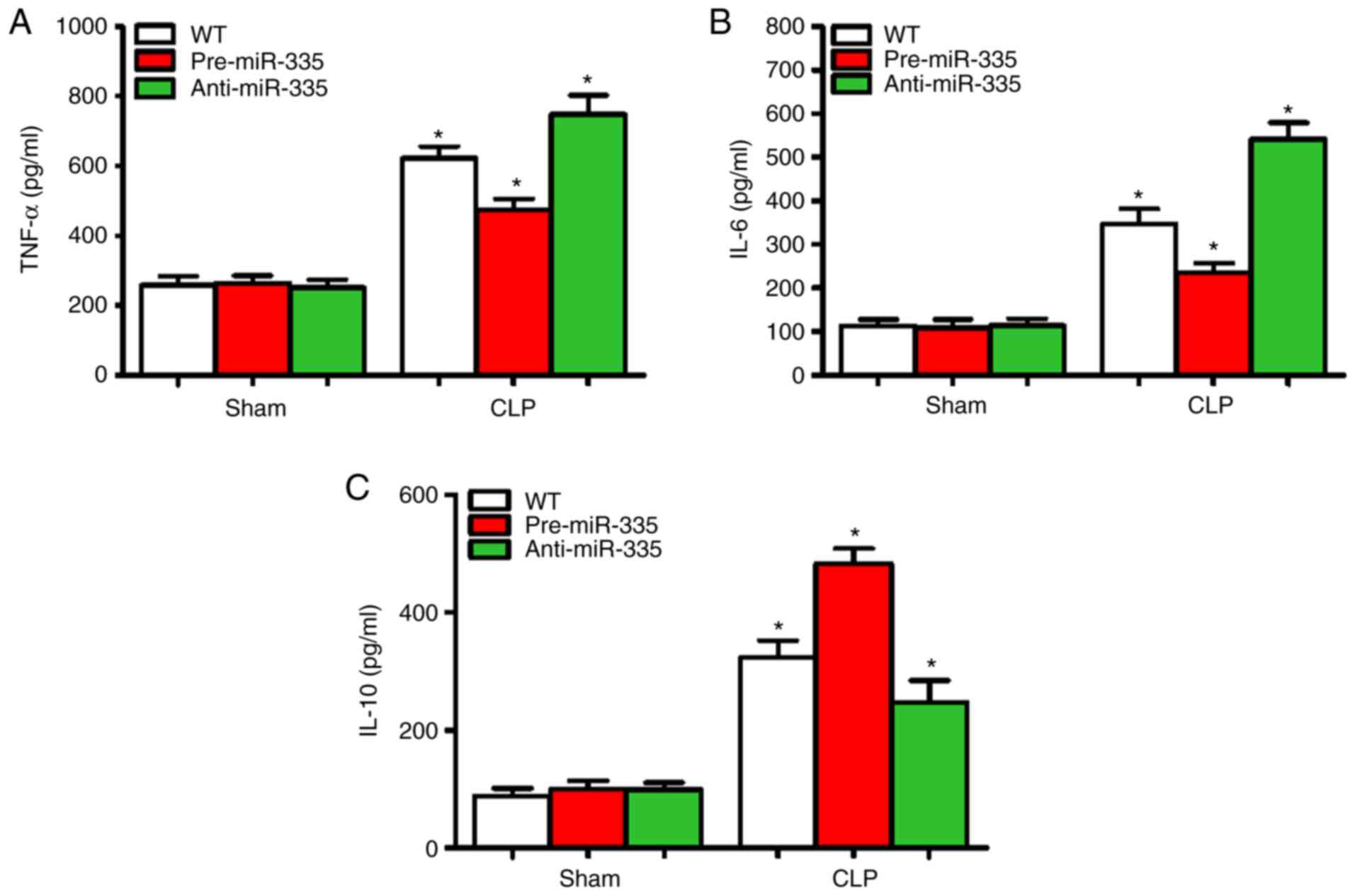
miR-335 inhibits myocardial
inflammation in septic mice
Sepsis-induced myocardial suppression may be
associated with increased cardiac inflammation (36). Therefore, the expression levels of
inflammatory cytokines (TNF-α, IL-6 and IL-10) were detected in the
myocardial tissues of mice 12 h after CLP. Compared with the WT
group, no changes in myocardial inflammatory cytokine levels
(TNF-α, IL-6 and IL-10) were observed in the pre-miR-335 and
anti-miR-335 mice (Fig. 5A-C).
Compared with the CLP-treated WT mice, pre-miR-335 caused a
decrease in TNF-α and IL-6 expression, while inflammation was
significantly aggravated in anti-miR-335 mice (Fig. 5A and B; n=6; P<0.05). Changes in
IL-10 showed the opposite trend (Fig.
5C). Compared with the WT Sham group, the levels of TNF-α in
the CLP-treated group were increased by 1.33 times in the
pre-miR-335 group and 2.69 times in the anti-miR-335 group
(Fig. 5A; n=6; P<0.05). It is
worth noting that compared with the sham-operated mice, CLP caused
an increase in IL-6 levels in the WT and pre-miR-335 mice, but
particularly in the anti-miR-335-treated mice (Fig. 5B; n=6; P<0.05). By contrast,
compared with the sham-operated WT group, IL-10 levels increased by
5.93 times in the pre-miRNA-335 mice post-CLP and were 2.58-fold
higher in anti-miRNA-335 than in pre-miRNA-335 mice (Fig. 5C; n=6; P<0.05).
Upregulation of miR-335 attenuates
SIMI
Detection of myocardial enzymes is critical for the
assessment of myocardial injury (22,37,38).
Therefore, in the present study, changes in myocardial enzyme
levels were examined 12 h after CLP treatment. Compared with the WT
sham-operated mice, no significant differences were observed in
cTNI, BNP, AST, LDH or CK in the sham-operated mice treated with
miRNA-335 precursors or miRNA-335 inhibitors (Fig. 6A-E; n=6; P>0.05). By contrast,
the levels of myocardial enzymes increased significantly in WT,
pre-miRNA-335 and anti-miRNA-335 mice following CLP. miRNA-335
precursors were then used to treat septic mice 4 days before
surgery. Compared with WT mice receiving CLP, significantly reduced
levels of serum cTNI, BNP, AST, LDH and CK were observed, while the
miRNA-335 inhibitor increased these levels (Fig. 6A-E; n=6; P<0.05).
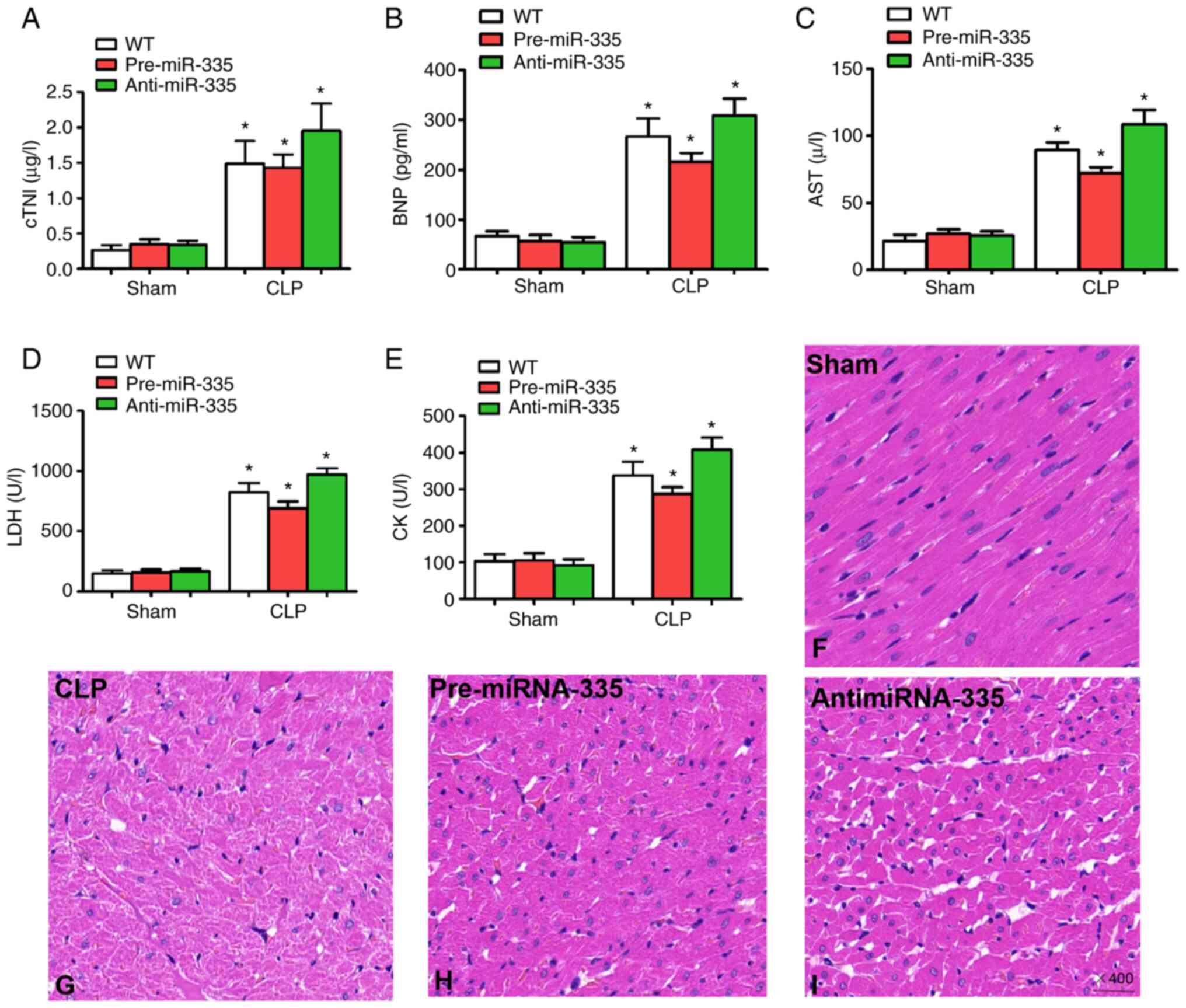 | Figure 6.Upregulation of miR-335 attenuates
sepsis-induced myocardial injury and histological changes. Levels
of (A) cTNI, (B) BNP, (C) AST, (D) LDH and (E) CK 12 h after CLP.
Pathological morphology was assessed by hematoxylin and eosin
staining; (F) Hearts from the Sham surgery group. (G) CLP mouse
heart. (H) Pre-miR-335 and (I) anti-miR-335 mouse heart
(magnification, ×400). *P<0.05 vs. WT. n=5. miRNA/miR, micro
RNA; cTNI, troponin; BNP, brain natriuretic peptide; AST, aspartate
aminotransferase; LDH, lactate dehydrogenase; CK, creatine kinase;
CLP, cecal ligation and puncture; WT, wild-type. |
In addition, the results of histological analysis
revealed myocardial degeneration, myocardial fibrillation and
interstitial edema in CLP-treated mice compared with the Sham group
(Fig. 6F and G; n=6). Pre-miR-335
mice showed reduced myocardial injury following CLP and accelerated
myocardial injury was observed in anti-miRNA-335 mice after CLP
(Fig. 6H and I; n=6). Therefore,
these findings indicated that treatment with miRNA-335 precursors
ameliorates SIMI in septic mice.
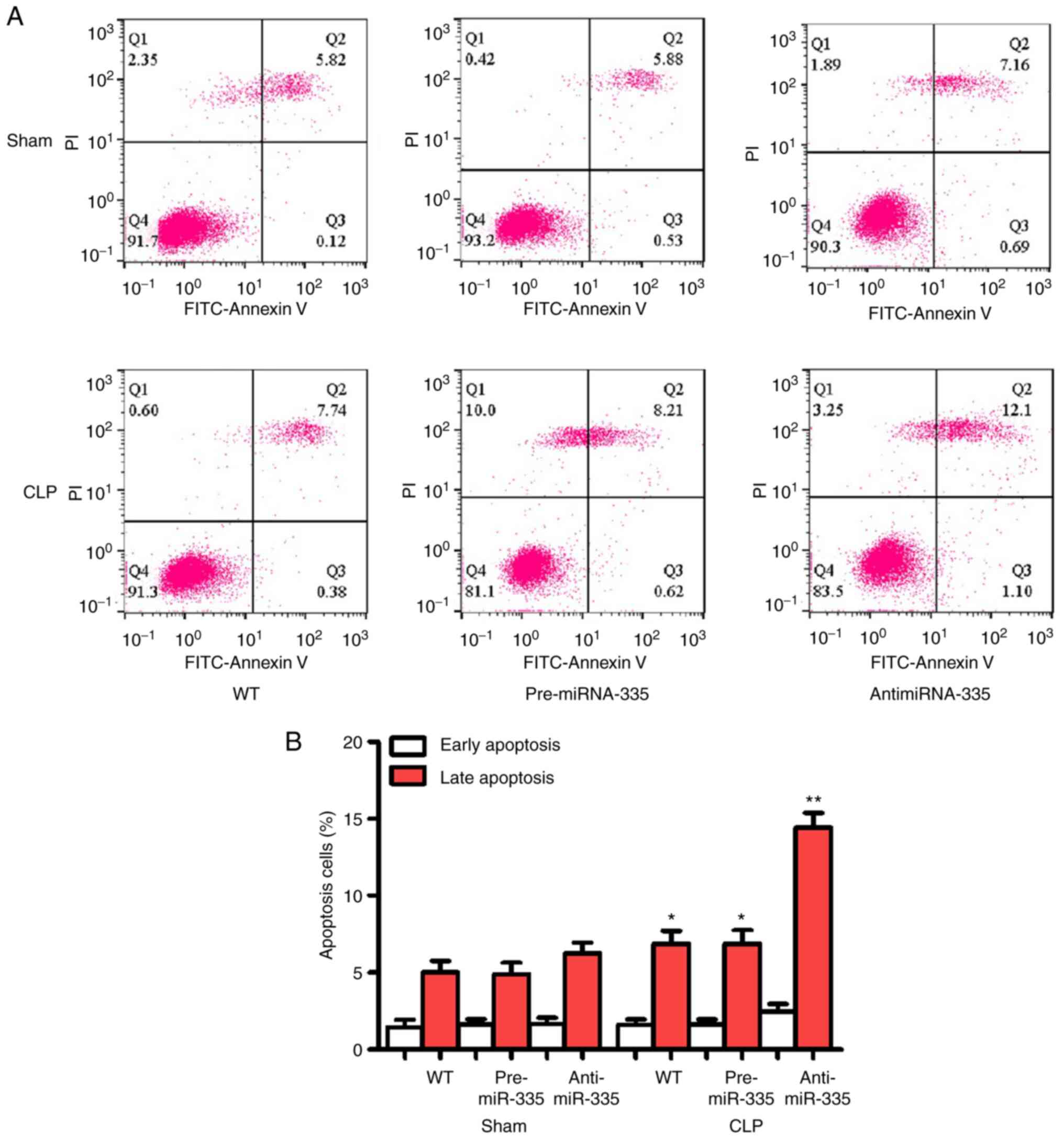
miR-335 inhibits apoptosis in
SIMI
Apoptosis is considered an important aggravator of
myocardial injury and may be an indicator for evaluating this
pathology. As shown in Fig. 7, no
significant differences in apoptosis were observed between the WT,
pre-miR-335 and anti-miR-335 groups following surgery. However, CLP
promoted significant apoptosis, which was reversed by pre-miRNA-335
treatment. Anti-miRNA-335-pretreated mice showed stable myocardial
apoptosis following CLP.
Discussion
The aim of the present study was to determine the
effects and underlying molecular mechanisms of miRNA-335 in SIMI.
The results suggested that pre-miRNA-335 reduced cardiac function,
the inflammatory response, myocardial damage and myocardial
apoptosis. These findings provided insights into the mechanisms
underlying miRNA-335 function and suggest that it may be a novel
target for attenuating SIMI.
Patients with SIMI exhibit higher rates of mortality
and disability worldwide than those without cardiovascular
dysfunction (39). SIMI is
considered to be difficult to treat. Although there are various
available treatments, including early target-oriented remedies,
timely appropriate antibiotic management, boosters and positive
inotropic interventions and mechanical ventilation (40), the development of effective
interventions to decrease mortality rates remains clinically
challenging. Clinical trials have widely focused on regulating
uncontrolled immune responses to suppress SIMI (41). miRNAs are extensively implicated in
biological and pathophysiological processes, including
sepsis-associated dysfunction (42–44).
Studies have shown that miRNA-335 is markedly upregulated in the
infarcted and bordered areas of the heart following acute
myocardial infarction in rats (42). Upregulation of miR-335 also
ameliorates myocardial ischemia/reperfusion injury by inhibiting
hypoxia-inducible factor 1α (45).
Furthermore, miR-335 protects the mouse heart from ischemic injury
by inhibiting Ca2+ overload and apoptosis (46). These studies indicate that miR-335
exerts protective effects in SIMI. In the present study, miR-335
expression was found to increase after CLP in mice, which proved
beneficial to sepsis-associated cardiac dysfunction. The results
confirmed that miR-335 protects against SIMI and further
demonstrated that miR-335 expression/reactivity is first increased
in sepsis, which is a compensatory protective mechanism, followed
by a gradual decrease. Further research confirmed that the decrease
in miR-335 expression was time-dependent. In order to investigate
the potential role of miR-335 in SIMI, miR-335 precursors and
inhibitors were then used to regulate miR-335 expression. The
results indicated that miR-335 preconditioning improves myocardial
dysfunction in septic mice.
miRNAs are a class of small non-coding RNA molecules
which are involved in various biological functions, such as
cellular differentiation, proliferation and transformation. Studies
have demonstrated that the aberrant expression of specific miRNAs
is associated with the occurrence and development of numerous
pathologies, including tumors and cardiovascular diseases (47–49). A
previous study demonstrated that miRNA-335 is involved in the
deterioration of a variety of tumors, as well as Alzheimer's
disease (50). However, to the best
of the authors' knowledge, there have been no reports on the
effects of miRNA-335 expression in SIMI. By establishing a sepsis
model and detecting the miRNA expression profiles of myocardial
tissues, the results of the present study illustrated that
miRNA-335 expression in myocardial tissue was significantly
upregulated following CLP in mice. However, the trend in miRNA-335
expression gradually decreased, which indicated that miRNA-335
downregulation is time-dependent. An miRNA-335 precursor or
inhibitor was then employed to investigate the potential role of
miRNA-335 in SIMI. The results revealed that the levels of cTNI,
BNP, CK, LDH, AST and BNP decreased in pre-miRNA-335 mice, while
the opposite effect was observed in anti-miRNA-335 mice. These
results suggested that miRNA-335 was involved in ameliorating
sepsis-induced myocardial dysfunction.
The inflammatory response is closely associated with
various cardiovascular diseases (51,52).
There is evidence to suggest that the accumulation of different
subsets of inflammatory cytokines occurs in the ischemic heart
(53). As important
pro-inflammatory cytokines, TNF-α and IL-6 are regarded as critical
contributors to the initiation and regulation of inflammatory
responses and their expression is elevated in the majority of
cardiovascular diseases (54). For
instance, TNF-α is a known potent inducer of cardiovascular disease
which accelerates myocardial injury. TNF-α also induces chemokine
expression in cardiac tissues, thus promoting uncontrolled
inflammation via NF-κB (55).
Further evidence supports that TNF-α impairs systolic and diastolic
function following electrical stimulation. In addition to TNF-α,
IL-6 is also involved in pro-inflammatory processes and may
contribute to increases in myocardial injury by promoting
neutrophil influx into the injured cardiac tissue (56). IL-6-deficiency reduces acute
myocardial I/R injury and there is mounting evidence to demonstrate
that the critical role of IL-6 in sepsis-induced cardiac
dysfunction is associated with the PI3K/Akt signaling pathway
(57). These data suggest that the
presence of myocardial inhibitory factors (TNF-α and IL-6) in the
serum, or incubation of cardiomyocytes in vitro may promote
contractile dysfunction in both animals and patients with sepsis
(46). Inhibiting TNF-α and IL-6
with neutralizing antibodies has proven beneficial for treating
sepsis (58). Therefore, the
significant reduction in inflammatory factors following septic
challenge may be one of the most important protective mechanisms of
pre-miR-335-mediated myocardial injury. The present results
confirmed that the upregulation of miR-335 inhibited
pro-inflammatory cytokine release and that its downregulation
exacerbated the release of pro-inflammatory cytokines. In addition,
anti-miR-335-treated CLP mice produced higher levels of the
anti-inflammatory cytokine IL-10, compared with the WT control
group, which may lead to an enlarged myocardial inflammatory
response.
Another cardioprotective mechanism of miR-335 during
sepsis may lie in its ability to reduce apoptosis (59). A large number of studies has
demonstrated that myocardial infarction, myocardial hypertrophy and
heart failure induce excessive apoptosis and that myocardial
apoptosis is involved in SIMI (37,60–62).
Sepsis-associated inflammatory factors induce cardiomyocyte
apoptosis by activating the caspase pathway (63). Additionally, sepsis promotes tissue
hypoxia/ischemia, resulting in a decline in the accumulation and
clearance of oxygen free radicals and an aggravation in
contractility and apoptosis in myocardial cells (64). In the present study, pre-miR-335
treatment inhibited myocardial apoptosis in septic mice. By
contrast, anti-miR-335-treated CLP mice exhibited accelerated
myocardial apoptosis. These results indicated that miRNA-335
reduced myocardial injury by inhibiting apoptosis. Moreover, in
vitro experiments have confirmed that miRNA-335 inhibits
myocardial cell apoptosis by inhibiting the PTEN/PI3K/Akt pathway
(22). These results suggest that
PTEN is a pro-apoptotic protein that negatively regulates the
primary phosphatase of the PI3K/Akt signaling (65). Studies have demonstrated that
miR-335 maintains cell survival by targeting the 3′untranslated
region of PTEN, which leads to the downregulation of PTEN and
activation of the Akt pathway in ovarian cancer and nasopharyngeal
carcinoma (66,67). These studies demonstrate that
miR-335 may relieve SIMI. However, whether miR-335 suppresses
apoptosis in SIMI-induced myocardial injury, which involves the
PTEN/PI3K/Akt pathway, warrants further investigation.
In conclusion, the results of the present study
indicated that miR-335 expression is upregulated in SIMI in CLP
mice, which is associated with myocardial protection during sepsis.
miR-335 reduced the inflammatory response associated with
myocardial injury in sepsis, by inhibiting pro-inflammatory
cytokine release and myocardial apoptosis. However, the absence of
overall inflammation levels of the mice might be a limitation of
the present study. Until now, to the best of the authors'
knowledge, the present study is the first to suggest that the
cardio-protection rendered by miR-335 may represent an effective
intervention for SIMI treatment.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by Hunan Natural Science
Foundation (grant no. 2020JJ7090 to Yongpan Huang).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL, YH, JH, XZ, YZ, YW, YT and LL designed and
performed experiments, and analyzed, interpreted and presented
results for group discussions. XL, YH, JH, XZ, YZ, YW, YT and LL
performed some of the experiments and confirm the authenticity of
all the raw data. JH and LL made substantial contributions to
conception and design. All authors reviewed and approved the final
manuscript.
Ethics approval and consent to
participate
All protocols were approved by the Medical Animal
Ethics Committee of Hunan Academy of Chinese Medicine (Xiang
20200015). All animals were handled in strict accordance with good
animal practice as defined by the Hunan Academy of Chinese Medicine
animal welfare bodies.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gomez E, Vercauteren M, Kurtz B,
Ouvrard-Pascaud A, Mulder P, Henry JP, Besnier M, Waget A, Hooft
Van Huijsduijnen R, Tremblay ML, et al: Reduction of heart failure
by pharmacological inhibition or gene deletion of protein tyrosine
phosphatase 1B. J Mol Cell Cardiol. 52:1257–1264. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Romero-Bermejo FJ, Ruiz-Bailen M,
Gil-Cebrian J and Huertos-Ranchal MJ: Sepsis-induced
cardiomyopathy. Curr Cardiol Rev. 7:163–183. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hochstadt A, Meroz Y and Landesberg G:
Myocardial dysfunction in severe sepsis and septic shock: More
questions than answers? J Cardiothorac Vasc Anesth. 25:526–535.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Patop IL, Wüst S and Kadener S: Past,
present and future of circRNAs. EMBO J. 38:e1008362019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meng S, Zhou H, Feng Z, Xu Z, Tang Y, Li P
and Wu M: CircRNA: Functions and properties of a novel potential
biomarker for cancer. Mol Cancer. 16:942017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhelankin AV, Vasiliev SV, Stonogina DA,
Babalyan KA, Sharova EI, Doludin YV, Shchekochikhin DY, Generozov
EV and Akselrod AS: Elevated Plasma levels of circulating
extracellular miR-320a-3p in patients with paroxysmal atrial
fibrillation. Int J Mol Sci. 21:34852020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang L, Wang HC, Chen C, Zeng JM, Wang Q,
Zheng L and Yu HD: Differential expression of plasma miR-146a in
sepsis patients compared with non-sepsis-SIRS patients. Exp Ther
Med. 5:1101–1104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ailawadi S, Wang X, Gu H and Fan GC:
Pathologic function and therapeutic potential of exosomes in
cardiovascular disease. Biochim Biophys Acta. 1852:1–11. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Halushka PV, Goodwin AJ and Halushka MK:
Opportunities for microRNAs in the crowded field of cardiovascular
biomarkers. Annu Rev Pathol. 14:211–238. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang HJ, Zhang PJ, Chen WJ, Feng D, Jia YH
and Xie LX: Four serum microRNAs identified as diagnostic
biomarkers of sepsis. J Trauma. 73:850–854. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang HJ: Serum miRNA-574-5p: A prognostic
predictor of sepsis patients. Shock. 37:263–267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang HJ, Deng J, Wang JY, Zhang PJ, Xin Z,
Xiao K, Feng D, Jia YH, Liu YN and Xie LX: Serum miRNA-122 levels
are related to coagulation disorders in sepsis patients. Clin Chem
Lad Med. 52:927–933. 2014.PubMed/NCBI
|
|
14
|
Tacke F, Roderburg C, Benz F, Cardenas DV,
Luedde M, Hippe HJ, Frey N, Vucur M, Gautheron J, Koch A, et al:
Levels of circulating miR-133a are elevated in sepsis and predict
mortality in critically ill patients. Crit Care Med. 42:1096–104.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dong S, Cheng Y, Yang J, Li J, Liu X, Wang
X, Wang D, Krall TJ, Delphin ES and Zhang C: MicroRNA expression
signature and the role of microRNA-21 in the early phase of acute
myocardial infarction. J Biol Chem. 284:29514–29525. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang X, Zhu H, Zhang X, Liu Y, Chen J,
Medvedovic M, Li H, Weiss MJ, Ren X and Fan GC: Loss of the
miR-144/451 cluster impairs ischaemic preconditioning-mediated
cardioprotection by targeting Rac-1. Cardiovasc Res. 94:379–390.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li W, Dong M, Chu L, Feng L and Sun X:
MicroRNA-451 relieves inflammation in cerebral ischemia-reperfusion
via the Toll-like receptor 4/MyD88/NF-κB signaling pathway. Mol Med
Rep. 20:3043–3054. 2019.PubMed/NCBI
|
|
18
|
Kura B, Szeiffova Bacova B, Kalocayova B,
Sykora M and Slezak J: Oxidative stress-responsive MicroRNAs in
heart injury. Int J Mol Sci. 21:3582020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang X, Wang X, Zhu H, Zhu C, Wang Y, Pu
WT, Jegga AG and Fan GC: Synergistic effects of the GATA-4-mediated
miR-144/451 cluster in protection against simulated
ischemia/reperfusion-induced cardiomyocyte death. J Mol Cell
Cardiol. 49:841–850. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang X, Zhang X, Ren XP, Chen J, Liu H,
Yang J, Medvedovic M, Hu Z and Fan GC: MicroRNA-494 targeting both
proapoptotic and antiapoptotic proteins protects against
ischemia/reperfusion induced cardiac injury. Circulation.
122:1308–1318. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu Y, Gao R, Kaul Z, Li L, Kato Y, Zhang
Z, Groden J, Kaul SC and Wadhwa R: Loss-of-function screening to
identify miRNAs involved in senescence: Tumor suppressor activity
of miRNA-335 and its new arget CARF. Sci Rep. 6:301852016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ge C, Liu J and Dong S: miRNA-214 protects
sepsis-induced myocardial injury. Shock. 50:112–118. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Wang H, Ding Y, Li Y, Chen S,
Zhang L, Wu H, Zhou J, Duan K, Wang W, et al: N-peptide of vMIP-II
reverses paclitaxel-resistance by regulating miRNA-335 in breast
cancer. Int J Oncol. 51:918–930. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
O'Shea KM, Ananthakrishnan R, Li Q, Quadri
N, Thiagarajan D, Sreejit G, Wang L, Zirpoli H, Aranda JF, Alberts
AS, et al: The formin, DIAPH1, is a key modulator of myocardial
ischemia/reperfusion injury. EBioMedicine. 26:165–174. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Togashi Y, Shirakawa J, Okuyama T,
Yamazaki S, Kyohara M, Miyazawa A, Suzuki T, Hamada M and Terauchi
Y: Evaluation of the appropriateness of using glucometers for
measuring the blood glucose levels in mice. Sci Rep. 6:254652016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Haak BW, Prescott HC and Wiersinga WJ:
Therapeutic potential of the gut microbiota in the prevention and
treatment of sepsis. Front Immunol. 9:20422018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang SB, Lin SY, Liu M, Liu CC, Ding HH,
Sun Y, Ma C, Guo RX, Lv YY, Wu SL, et al: CircAnks1a in the spinal
cord regulates hypersensitivity in a rodent model of neuropathic
pain. Nat Commun. 10:41192019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liang L, Fu J, Wang S, Cen H, Zhang L,
Mandukhail SR, Du L, Wu Q, Zhang P and Yu X: MiR-142-3p enhances
chemosensitivity of breast cancer cells and inhibits autophagy by
targeting HMGB1. Acta Pharm Sin B. 10:1036–1046. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao Y, Zhao H and Li Y: LncRNA MCM3AP-AS1
regulates miR-142-3p/HMGB1 to promote LPS-induced chondrocyte
apoptosis. BMC Musculoskelet Disord. 20:6052019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liang H, Su X, Wu Q, Shan H, Lv L, Yu T,
Zhao X, Sun J, Yang R, Zhang L, et al: LncRNA 2810403D21Rik/Mirf
promotes ischemic myocardial injury by regulating autophagy through
targeting Mir26a. Autophagy. 16:1077–1091. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang YP, Gao FF, Wang B, Zheng FC, Zhang
YM, Chen YC, Huang ZQ, Zheng YS, Zhong SP and Shi GG: N-n-butyl
haloperidol iodide inhibits H2O2-induced
Na+/Ca2+-exchanger activation via the Na+/H+ exchanger
in rat ventricular myocytes. Drug Des Devel Ther. 8:1257–1267.
2014.PubMed/NCBI
|
|
33
|
Chistiakov DA, Orekhov AN and Bobryshev
YV: Cardiac-specific miRNA in cardiogenesis, heart function and
cardiac pathology (with focus on myocardial infarction). J Mol Cell
Cardiol. 94:107–121. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bai XY, Ma Y, Ding R, Fu B, Shi S and Chen
XM: miR-335 and miR-34a Promote renal senescence by suppressing
mitochondrial antioxidative enzymes. J Am Soc Nephrol.
22:1252–1261. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu Y, Lai P, Deng J, Hao Q, Li X, Yang M,
Wang H and Dong B: Micro-RNA335-5p targeted inhibition of sKlotho
and promoted oxidativestress-mediated aging of endothelial cells.
Biomark Med. 13:457–466. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu ZJ, Chen YF, Wang HD and Gao FH:
Expression of plasma miRNA-497 in children with sepsis-induced
myocardial injury and its clinical significance. Zhongguo Dang Dai
Er Ke Za Zhi. 20:32–36. 2018.(In Chinese). PubMed/NCBI
|
|
37
|
Wang SM, Liu GQ, Xian HB, Si JL, Qi SX and
Yu YP: LcRNA NEAT1 alleviates sepsis-induced myocardial injury by
regulating the TLR2/NF-κB signaling pathway. Eur Rev Med Pharmacol
Sci. 23:4898–4907. 2019.PubMed/NCBI
|
|
38
|
An R, Feng J, Xi C, Xu J and Sun L:
miR-146a attenuates sepsis-induced myocardial dysfunction by
suppressing IRAK1 and TRAF6 via Targeting ErbB4 Expression. Oxid
Med Cell Longev. 2018:71630572018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shang X, Li J, Yu R, Zhu P, Zhang Y, Xu J,
Chen K and Li M: Sepsis-related myocardial injury is associated
with mst1 upregulation, mitochondrial dysfunction and the
Drp1/F-actin signaling pathway. J Mol Histol. 50:91–103. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rudiger A and Singer M: The heart in
sepsis: From basic mechanisms to clinical management. Curr Vasc
Pharmacol. 11:187–195. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Reinhart K, Bauer M, Riedemann NC and
Hartog CS: New approaches to sepsis: Molecular diagnostics and
biomarkers. Clin Microbiol Rev. 25:609–634. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu Y, Liu L and Zhang J: Protective role
of matrine in sepsis-associated cardiac dysfunction through
regulating the lncRNA PTENP1/miR-106b-5p axis. Biomed Pharmacother.
134:1111122021. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sluijter JP and Doevendans PA:
Sepsis-associated cardiac dysfunction is controlled by small RNA
molecules. J Mol Cell Cardiol. 97:67–69. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang H, Bei Y, Shen S, Huang P, Shi J,
Zhang J, Sun Q, Chen Y, Yang Y, Xu T, et al: miR-21-3p controls
sepsis-associated cardiac dysfunction via regulating SORBS2. J Mol
Cell Cardiol. 94:43–53. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wu N, Zhang X, Du S, Chen D and Che R:
Upregulation of miR-335 ameliorates myocardial ischemia reperfusion
injury via targeting hypoxia inducible factor 1-alpha subunit
inhibitor. Am J Transl Res. 10:4082–4094. 2018.PubMed/NCBI
|
|
46
|
Joulin O, Petillot P, Labalette M, Lancel
S and Neviere R: Cytokine profile of human septic shock serum
inducing cardiomyocyte contractile dysfunction. Physiol Res.
56:291–297. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Correia de Sousa M, Gjorgjieva M, Dolicka
D, Sobolewski C and Foti M: Deciphering miRNAs' Action through
miRNA Editing. Int J Mol Sci. 20:62492019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Agnelli L, Bisognin A, Todoerti K, Manzoni
M, Taiana E, Galletti S, Cutrona G, Gaffo E, Bortoluzzi S and Neri
A: Expanding the repertoire of miRNAs and miRNA-offset RNAs
expressed in multiple myeloma by small RNA deep sequencing. Blood
Cancer J. 9:212019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Huang W: MicroRNAs: Biomarkers,
diagnostics, and therapeutics. Methods Mol Biol. 1617:57–67. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Moradifard S, Hoseinbeyki M, Ganji SM and
Minuchehr Z: Analysis of microRNA and gene expression profiles in
Alzheimer's disease: A Meta-analysis approach. Sci Rep. 8:47672018.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Papaioannou V, Pneumatikos I and
Maglaveras N: Association of heart rate variability and
inflammatory response in patients with cardiovascular diseases:
Current strengths and limitations. Front Physiol. 4:1742013.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ruiz-Ortega M, Esteban V and Egido J: The
regulation of the inflammatory response through nuclear
factor-kappab pathway by angiotensin IV extends the role of the
renin angiotensin system in cardiovascular diseases. Trends
Cardiovasc Med. 17:19–25. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Frangogiannis NG: Pathophysiology of
myocardial infarction. Compr Physiol. 5:1841–1875. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Hanna A and Frangogiannis NG: Inflammatory
cytokines and chemokines as therapeutic targets in heart failure.
Cardiovasc Drugs Ther. 34:849–863. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bartekova M, Radosinska J, Jelemensky M
and Dhalla NS: Role of cytokines and inflammation in heart function
during health and disease. Heart Fail Rev. 23:733–758. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bai R, Yin X, Feng X, Cao Y, Wu Y, Zhu Z,
Li C, Tu P and Chai X: Corydalis hendersonii Hemsl. Protects
against myocardial injury by attenuating inflammation and fibrosis
via NF-κB and JAK2-STAT3 signaling pathways. J Ethnopharmacol.
207:174–183. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Qiu Z, He Y, Ming H, Lei S, Leng Y and Xia
ZY: Lipopolysaccharide (LPS) aggravates high glucose- and
hypoxia/reoxygenation-induced injury through activating
ROS-dependent NLRP3 inflammasome-mediated pyroptosis in H9C2
cardiomyocytes. J Diabetes Res. 2019:81518362019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Beutler B, Milsark IW and Cerami AC:
Passive immunization against cachectin/tumor necrosis factor
protects mice from lethal effect of endotoxin. J Immunol. 181:7–9.
2008.PubMed/NCBI
|
|
59
|
Gao XL, Li JQ, Dong YT, Cheng EJ, Gong JN,
Qin YL, Huang YQ, Yang JJ, Wang SJ and An DD: Upregulation of
microRNA-335-5p reduces inflammatory responses by inhibiting FASN
through the activation of AMPK/ULK1 signaling pathway in a septic
mouse model. Cytokine. 110:466–478. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Fadeel B, Orrenius S and Zhivotovsky B:
The potential role of apoptosis in human disease. Med Princ Pract.
9:151–163. 2000. View Article : Google Scholar
|
|
61
|
Kong W, Kang K, Gao Y, Liu H, Meng X, Cao
Y, Yang S, Liu W, Zhang J, Yu K and Zhao M: GTS-21 protected
against LPS-induced sepsis myocardial injury in mice through
α7nAChR. Inflammation. 41:1073–1083. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang X and Yu Y: miR-146b protect against
sepsis induced mice myocardial injury through inhibition of Notch1.
J Mol Histol. 49:411–417. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Meng YY, Liu Y, Hu ZF, Zhang Y, Ni J, Ma
ZG, Liao HH, Wu QQ and Tang QZ: Sanguinarine attenuates
lipopolysaccharide-induced inflammation and apoptosis by inhibiting
the TLR4/NF-κB pathway in H9c2 cardiomyocytes. Curr Med Sci.
38:204–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Falck M, Osredkar D, Wood TR, Maes E,
Flatebø T, Sabir H and Thoresen M: Neonatal systemic inflammation
induces inflammatory reactions and brain apoptosis in a
pathogen-specific manner. Neonatology. 113:212–220. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mocanu MM and Yellon DM: PTEN, the
Achilles' heel of myocardial ischaemia/reperfusion injury? Br J
Pharmacol. 150:833–838. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhou XM, Sun R, Luo DH, Sun J, Zhang MY,
Wang MH, Yang Y, Wang HY and Mai SJ: Upregulated TRIM29 promotes
proliferation and metastasis of nasopharyngeal carcinoma via
PTEN/AKT/mTOR signal pathway. Oncotarget. 7:13634–13650. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Yang H, Kong W, He L, Zhao JJ, O'Donnell
JD, Wang J, Wenham RM, Coppola D, Kruk PA, Nicosia SV and Cheng JQ:
MicroRNA expression profiling in human ovarian cancer: miR-335
induces cell survival and cisplatin resistance by targeting PTEN
MicroRNA expression profiling in human ovarian cancer: miR-335
induces cell survival and cisplatin resistance by targeting PTEN.
Cancer Res. 68:425–433. 2008. View Article : Google Scholar : PubMed/NCBI
|