Introduction
Allergic rhinitis (AR) is a chronic inflammatory
disease of the upper airway with a high worldwide prevalence
(10–40%). AR is characterized by sneezing, pruritus, rhinorrhea,
retronasal drainage and nasal congestion (1–3). AR is
triggered by allergen-specific IgE-mediated reactions, and type 2
helper T (Th2) cells further drive this inflammatory response
against inhaled allergens in the environment (4). These inhaled allergens include
pollens, dust mites, animal dander and mold (5). Multiple inflammatory cells, such as B
and T cells, basophils and mast cells, are involved in this
inflammatory process (6). AR has a
severe impact on the quality of life of patients. Children or
adults with AR may suffer from learning impairments, and sleep or
emotional disturbances (7). In
recent years, the prevalence of AR has increased at an alarming
rate in adults and children worldwide (8). In addition, the presence of AR could
significantly increase the risk of asthma (9); according to previous studies, ≤40% of
individuals with AR either have or will get asthma (5,9–11).
Furthermore, patients with AR usually suffer from allergic
conjunctivitis (12). AR presents a
great challenge to humans, due to its high prevalence and other
complications. Pharmacotherapy is one of the major approaches to
the management of AR, and nasal corticosteroids and antihistamines
are the most common medications. Effective pharmacotherapy requires
regular use of medications so that the symptoms do not recur
following discontinuation (13–15).
MicroRNAs (miRNAs/miRs) are single-stranded
non-coding RNA molecules ~22 nucleotides in length. miRNAs mediate
silencing of target genes by binding to target mRNA (16). Mature miRNAs bind to the
3′-untranslated region (3′-UTR) of their target mRNA transcripts
with the help of the RNA-induced silencing complex, and then induce
the degradation of target mRNA or inhibit protein translation
(17,18). miRNAs serve an important role in a
variety of physiological and pathological processes. Abnormal miRNA
expression has been reported to be involved in the occurrence and
development of various human diseases, including AR (19,20).
Suojalehto et al (21)
reported that miR-205-5p was upregulated in the nasal mucosa of
symptomatic patients with AR. The significance of miR-205-5p in
regulating inflammation has also been demonstrated. Notably, IL-32α
has been shown to suppress vascular inflammation and
atherosclerosis by inhibiting miR-205-5p biogenesis in mice
(22). Furthermore, miR-205-5p has
been demonstrated to alleviate hip fracture-induced rat lung injury
by decreasing inflammatory response through inhibition of the
expression of high mobility group box 1 (23). These studies suggested that
miR-205-5p may have different functions in different pathological
environments, which can be explained by its target gene or
regulatory mechanism. However, to the best of our knowledge, the
role of miR-205-5p in AR remains unknown and requires further
investigation.
B-cell lymphoma 6 (BCL6) is a sequence-specific
transcriptional regulator that inhibits the transcription of target
genes by binding to a specific DNA sequence in the promoter region
(24,25). BCL6 serves an important role in the
development of B cells, follicular Th cells and T regulatory cells,
indicating its potential function in regulation of the immune
response (26–28). BCL6 has also been reported to be
downregulated in the nasal mucosa of patients with AR (29). Hiromura et al (30) reported that IL-21 upregulated BCL6
and relieved AR. In addition, BCL6 may inhibit the inflammatory
response in human renal tubular epithelial cells and attenuate
renal inflammation by negatively regulating the transcription of
nucleotide-binding oligomerization domain-like receptor family
pyrin domain-containing 3 (NLRP3) (31). These findings suggested the
importance of BCL6 in the regulation of inflammation in human
diseases. Therefore, it was hypothesized that BCL6 could also be
involved in AR-related inflammation.
With the rising prevalence of AR, it is necessary to
develop novel and effective approaches to AR treatment. In the
present study, the functions of miR-205-5p and its underlying
regulatory mechanism in AR were explored. The present study aimed
to improve understanding of AR pathogenesis and a potential
therapeutic target for AR.
Materials and methods
Animal model
Female wild-type (WT) BALB/c mice (age, 4 weeks;
weight, 18–22 g) were purchased from Liaoning Changsheng
Biotechnology Co., Ltd. A total of 168 mice were used in the
present study. All animal experiments were approved by the Ethics
Committee of The Second Affiliated Hospital of Shenyang Medical
College (approval no. 2020005; Shenyang, China). Mice were
maintained under a 12-h dark/light cycle, at 45–55% humidity and
22±1°C room temperature with free access to food and water. After a
1-week acclimation period, all mice were randomly divided into the
Sham, AR, AR + negative control (NC) and AR + Lv-anti-miR-205-5p
sponge groups. A mouse model of AR was established according to a
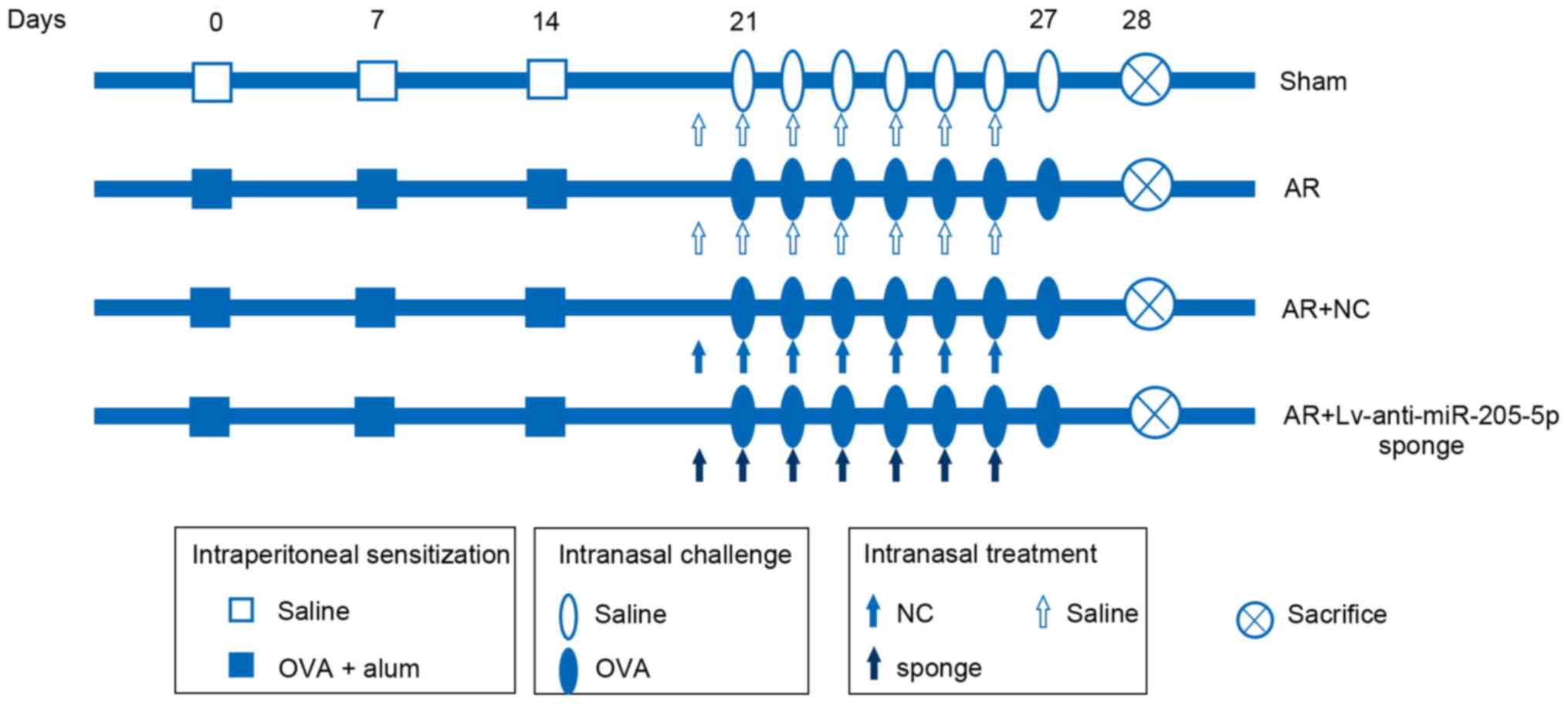
previous study (32). As shown in
Fig. 1, in the AR group, the mice
were sensitized via an intraperitoneal (i.p.) injection of 200 µl
saline containing 25 µg ovalbumin (OVA; Shanghai Aladdin
Biochemical Technology Co., Ltd.) and 2 mg aluminum hydroxide on
days 0, 7 and 14. The mice were then subjected to continuous
intranasal challenges with 100 µg OVA (50 µl; 2 mg/ml) on days
21–27. The sham group was administered an equivalent amount of
saline without OVA or aluminum hydroxide on days 0, 7 and 14, and
on days 21–27. A total of 24 h before each OVA challenge on days
21–27, a recombinant lentiviral vector carrying an anti-miR-205-5p
sponge or NC (empty vector) was constructed and intranasally
administered to the mice (20 µl; 1×108 TU/ml).
The frequency of nose-rubbing or sneezing within 15
min from the final OVA challenge was recorded. A total of 24 h
after the final OVA challenge, mice were euthanized with an
overdose of sodium pentobarbital (150 mg/kg, i.p.). Blood (~1 ml)
was collected from the retrobulbar vein immediately and serum was
isolated for use (33,34). Nasal lavage fluid (NLF) was
harvested by cannulating the upper part of the trachea into nasal
cavity and lavaging with 1 ml PBS, followed by centrifugation at
277 g for 10 min at 4°C. The collected serum, NLF and some nasal
mucosa tissues were frozen in liquid nitrogen and then stored at
−70°C for Gimesa staining, ELISA, reverse
transcription-quantitative (RT-q)PCR and western blot analysis.
Some nasal mucosa tissues were fixed in 4% paraformaldehyde at room
temperature for 24 h for histological analysis.
RT-qPCR
Total RNA was isolated from the nasal mucosa using
TRIpure (BioTeke Corporation) and reverse transcribed into
first-strand cDNA at 25°C for 10 min, 42°C for 50 min and 80°C for
10 min using Super M-MLV Reverse Transcriptase (BioTeke
Corporation). qPCR was then performed using an Exicycler™ 96
Real-time PCR system (Bioneer Corporation) and SYBR Green kit
(Merck KGaA) according to the manufacturer's instructions. The
reaction conditions were as follows: 94°C for 5 min, 94°C for 15
sec, 60°C for 25 sec and 72°C for 30 sec; followed by 40 cycles of
72°C for 5 min 30 sec, 40°C for 5 min 30 sec; melting at 60–94°C
for 34 sec and final extension for 2 min at 25°C.
The transcript levels of genes were analyzed
quantitatively using the 2−ΔΔCq method and normalized to
5S or β-actin (35). The primer
sequences used were as follows: mmu-miR-205-5p, forward,
5′-TCCTTCATTCCACCGGAGTCTG-3′ and reverse, 5′-GCAGGGTCCGAGGTATTC-3′;
5S, forward, 5′-CTAAAGATTTCCGTGGAGAG-3′ and reverse,
5′-TGGTGCAGGGTCCGAGGTAT-3′; BCL6, forward, 5′-CGGAAGGGTCTGGTAGT-3′
and reverse, 5′-CATTCTGATTGAGGCTGTTG-3′; β-actin, forward,
5′-CTGTGCCCATCTACGAGGGCTAT-3′ and reverse,
5′-TTTGATGTCACGCACGATTTCC-3′.
Histological analysis
H&E staining was performed to assess
histopathological alterations in nasal mucosa tissues. Briefly,
mouse nasal mucosa tissues fixed with 4% paraformaldehyde at room
temperature for 24 h were dehydrated in increasing ethanol
gradients and embedded in paraffin. The tissues were cut into 5-µm
slices and stained with 0.2% hematoxylin (Beijing Solarbio Science
& Technology Co., Ltd.) for 5 min and 0.35% eosin (Sangon
Biotech Co., Ltd.) for 3 min at room temperature for
histopathological examination. Histopathological changes were
observed under a BX53 light microscope (Olympus Corporation) at a
magnification of ×200. The replicates of histological analysis came
from seven different mice in each group and only the representative
image was displayed.
Semi-quantification of inflammatory
cells in NLF
Inflammatory cells in NLF were measured by
cytochemical staining with Wright's-Giemsa. Mouse NLF was collected
and centrifuged at 300 × g for 10 min at the room temperature.
Nasal lavage smears were stained with Wright's-Giemsa (cat. no.
D010; Nanjing Jiancheng Bioengineering Institute) at room
temperature according to the manufacturer's instructions, and
inflammatory cells were identified as leukocytes, eosinophils,
neutrophils, lymphocytes and macrophages, according to their
morphology. NLF cell differentials were counted under a light
microscope at a magnification of ×200. In total, five random fields
of each sample were captured for average counting.
Western blot analysis
The nasal mucosa tissue of each mouse was ground in
liquid nitrogen with protein lysis buffer (Beyotime Institute of
Biotechnology) containing 1 mM PMSF and placed on ice for 5 min.
After centrifugation at 10,000 × g and 4°C for 5 min, the
supernatant was collected and quantified using a BCA kit (Beyotime
Institute of Biotechnology). The mean protein concentration was
3.46 µg/µl (Sham group), 3.31 µg/µl (AR group), 3.81 µg/µl (AR + NC
group) and 3.88 µg/µl (AR + Lv-anti-miR-205-5p sponge group). The
amount of protein loaded for BCL6 detection was 20 µg per sample,
and that for the detection of IL-1β, caspase-1, ASC or NLRP3 was 40
µg per sample. The proteins were separated by SDS-PAGE on 10% gels,
transferred to polyvinylidene difluoride membranes
(MilliporeSigma), and then blocked with 5% skimmed milk solution
for 1 h at room temperature. The membranes were incubated with
primary antibodies (Table I)
overnight at 4°C. After washing with TBS-Tween-20 (1.5% m/v)
buffer, the membranes were incubated with goat anti-rabbit IgG-HRP
(dilution, 1:5,000; cat. no. A0208; Beyotime Institute of
Biotechnology) or goat anti-mouse IgG-HRP secondary antibodies
(dilution, 1:5,000; cat. no. A0216; Beyotime Institute of
Biotechnology) for 45 min at 37°C. β-actin was used as an internal
reference. All antibodies used for western blot analysis were
polyclonal antibodies. Protein bands were visualized and analyzed
using enhanced chemiluminescence reagent (Beyotime Institute of
Biotechnology) under a gel imaging Gel-Pro-Analyzer system
(WD-9413B; Beijing Liuyi Biotechnology Co., Ltd.). Image-Pro Plus
software 6.0 (Media Cybernetics, Inc.) was used for
semi-quantification. The western blot analysis replicates were from
seven different mice in each group.
 | Table I.Primary antibodies used in western
blot analysis. |
Table I.
Primary antibodies used in western
blot analysis.
| Primary
antibody | Dilution | Supplier | Cat. no. |
|---|
| BCL6 | 1:500 | ProteinTech Group,
Inc. | 21187-1-AP |
| IL-1βa | 1:400 | Affinity
Biosciences, Ltd. | AF5103 |
|
Caspase-1b | 1:1,000 | Affinity
Biosciences, Ltd. | AF5418 |
| ASC | 1:500 | ABclonal Biotech
Co., Ltd. | A1170 |
| NLRP3 | 1:1,000 | ABclonal Biotech
Co., Ltd. | A5652 |
| β-actin | 1:1,000 | Santa Cruz
Biotechnology Co., Ltd. | sc-47778 |
ELISA
ELISA was used to measure the serum levels of total
and OVA-specific IgE, and those of IL-4, IL-5 and IL-13 in nasal
mucosa tissues. The serum levels of total and OVA-specific IgE were
detected using a Mouse IgE ELISA kit [cat. no. EK275; Hangzhou
Multisciences (Lianke) Biotech Co., Ltd.] and Mouse OVA-sIgE ELISA
kit (cat. no. JL46328; Jianglaibio Biology), respectively,
according to the manufacturer's instructions. To detect the levels
of IL-4, IL-5 and IL-13, nasal mucosa tissues were homogenized in
saline and centrifuged at 430 × g for 10 min at the room
temperature. The concentration of proteins in the supernatant was
quantified using a BCA kit (Beyotime Institute of Biotechnology).
The concentration of IL-4 (cat. no. EK0405), IL-5 (cat. no. EK0408)
or IL-13 (cat. no. EK0425) was assessed using the corresponding
ELISA kit (Boster Biological Technology). The levels of IL-4, IL-5
or IL-13 in nasal mucosa tissues were expressed as the ratio of
cytokine concentration to isolated protein concentration (pg/mg
protein). Replicates were from seven different mice in each
group.
Luciferase reporter assay
Luciferase reporter assay was performed to verify
the binding of miR-205-5p to its target mRNA. The putative target
was predicted according to the prediction website (TargetScanHuman;
targetscan.org/vert_72/); BCL6 was
revealed to be a potential target of miR-205-5p. The wild-type (WT)
3′-UTR of BCL6 and its mutant (MT) sequence, synthesized by
GenScript (Nanjing) Co., Ltd., were cloned into the pmirGLO vector
[GenScript (Nanjing) Co., Ltd.]. 293T cells (Shanghai Zhongqiao
Xinzhou Biotechnology Co., Ltd.) at the density of 70% were
co-transfected with the recombinant vector (1.5 µg, 0.5 µg/ml)
containing the WT or MT 3′-UTR of BCL6 plus miR-205 or NC mimics
(75 pmol, 25 pmol/ml) at 37°C for 4 h using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The sequences, synthesized by JTS scientific,
were as follows: miR-205-5p mimics, 5′-UCCUUCAUUCCACCGGAGUCUG-3′
and 5′-GACUCCGGUGGAAUGAAGGAUU-3′; NC mimics,
5′-UUCUCCGAACGUGUCACGUTT-3′ and 5′-ACGUGACACGUUCGGAGAATT-3′. Cells
were grown in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
containing 10% FBS (Cytiva) and collected 48 h after transfection
to determine luciferase signals using a dual-luciferase assay
reporter kit (Promega Corporation). The relative luciferase
activities represented the firefly/Renilla luciferase
ratios. The experiment was repeated at least three times.
Immunohistochemistry
The fixed and paraffin-embedded nasal mucosa tissue
slides were deparaffinized, rehydrated and subjected to boiling
antigen retrieval for 10 min using 10% citrate buffer solution.
Subsequently, the slides were exposed to 3%
H2O2 (Sinopharm Chemical Reagent Co., Ltd.)
for 15 min and then blocked with 100% goat serum (Beijing Solarbio
Science & Technology Co., Ltd.) for 15 min at room temperature.
These slides were stained with a polyclonal anti-BCL6 primary
antibody (dilution, 1:100; cat. no. Bs-2734R; Beijing Biosynthesis
Biotechnology Co., Ltd.) overnight at 4°C. The slides were then
washed with PBS three times, followed by incubation with polyclonal
HRP-conjugated secondary antibody (dilution, 1:500; cat. no. 31460;
Thermo Fisher Scientific, Inc.) at 37°C for 60 min. Chromogenic
detection was performed using a DAB Substrate kit (Beijing Solarbio
Science & Technology Co., Ltd.), followed by counterstaining
with hematoxylin. Immunohistochemistry images were captured at a
magnification of ×400 under a light microscope. The replicates came
from seven different mice in each group and only the representative
image was displayed. For semi-quantification of
immunohistochemistry, three random fields were captured, and
integrated optical density and area of each image were measured by
Image-Pro Plus software 6.0 (Media Cybernetics, Inc.). The mean
density represented BCL6 expression in the nasal mucosa.
Immunofluorescence staining
After dewaxing and rehydration, the fixed and
paraffin-embedded nasal mucosa tissue slides underwent antigen
retrieval as aforementioned, followed by blocking with 100% goat
serum (Beijing Solarbio Science & Technology Co., Ltd.) for 15
min at room temperature. The slides were then incubated with a
mixture of polyclonal anti-BCL6 (dilution, 1:100; cat. no.
Bs-2734R; Beijing Biosynthesis Biotechnology Co., Ltd.) and
monoclonal anti-CD4 (dilution, 1:50; cat. no. Sc-20079; Santa Cruz
Biotechnology, Inc.) or anti-F4/80 (dilution, 1:50; cat. no.
Sc-377009; Santa Cruz Biotechnology, Inc.) antibodies overnight at
4°C. Subsequently, the sections were incubated with a mixture of
Cy3-conjugated goat anti-rabbit IgG (dilution, 1:200; cat. no.
A0516; Beyotime Institute of Biotechnology) and fluorescein
isothiocyanate-labelled goat anti-mouse IgG (dilution, 1:200; cat.
no. A0568; Beyotime Institute of Biotechnology) antibodies at room
temperature for 90 min. Nuclei were counterstained with DAPI
(Shanghai Aladdin Biochemical Technology Co., Ltd.). The slides
were visually examined under a BX53 immunofluorescence microscope
(Olympus Corporation) at a magnification of ×400. The replicates
came from seven different mice in each group, and only the
representative image was displayed.
Statistical analysis
All experiments were performed with seven biological
replicates. Data are presented as the mean ± SD. Statistical
analysis was performed using GraphPad Prism 7.0 software (GraphPad
Software, Inc.). One-way ANOVA followed by Tukey's post-hoc test
was used for multiple comparisons among groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-205-5p knockdown alleviates
OVA-induced AR
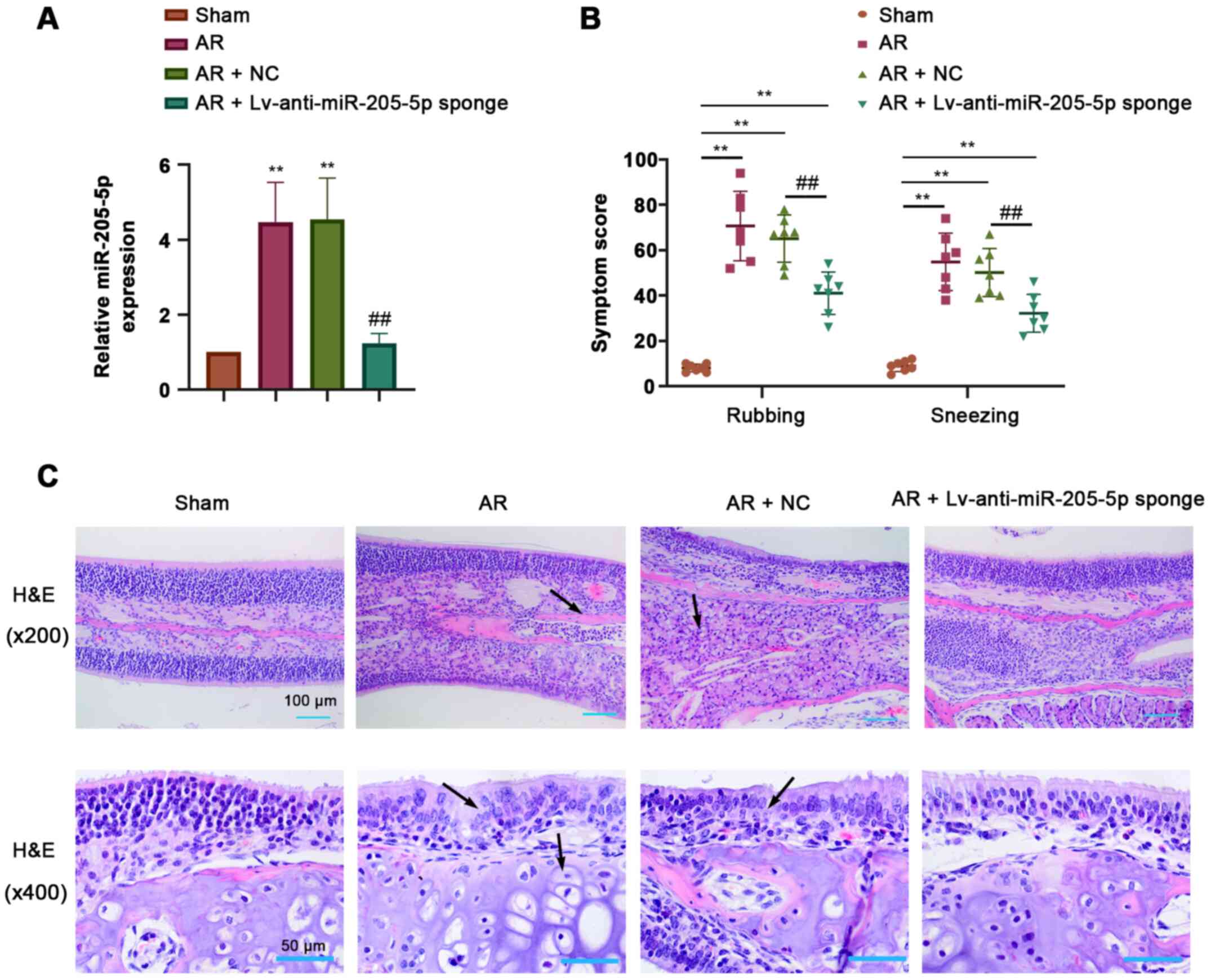
To determine the role of miR-205-5p in AR, the
expression levels of miR-205-5p were detected by RT-qPCR in nasal
mucosa tissues from AR mice sensitized by OVA. miR-205-5p
expression was significantly increased in response to OVA-induced
AR compared with that in the sham group, and was efficiently
downregulated by the intranasal administration of a lentiviral
sponge for miR-205-5p (Fig. 2A).
The frequency of nose-rubbing or sneezing within 15 min of the
final OVA challenge was recorded to evaluate the OVA-triggered AR
response. The frequency of nose-rubbing or sneezing of AR mice was
markedly increased compared with that of sham mice but was
significantly alleviated by miR-205-5p knockdown (Fig. 2B). H&E staining showed apparent
pathological alterations in the nasal mucosa of AR mice, including
disarrangement of the epidermis, capillary edema and inflammatory
cell infiltration. miR-205-5p knockdown attenuated hyperemia and
inflammatory cell infiltration in the nasal mucosa of AR mice
(Fig. 2C). These results suggested
that miR-205-5p knockdown may alleviate OVA-induced AR.
miR-205-5p knockdown attenuates
OVA-induced inflammatory response
ELISA was performed to measure the levels of total
and OVA-specific IgE, IL-4, IL-5 and IL-13 during AR. The
concentrations of both total and OVA-specific IgE in the serum were
increased in AR mice compared with those detected in sham mice, and
were decreased by miR-205-5p knockdown (Fig. 3A). Wright's-Giemsa staining showed
that the number of inflammatory cells, including leukocytes,
eosinophils, neutrophils, lymphocytes and macrophages, was
significantly higher in AR mice than that in sham mice. Conversely,
the AR-induced increase in the number of inflammatory cells was
significantly reduced by miR-205-5p knockdown (Fig. 3B). In addition, the levels of IL-4,
IL-5 and IL-13 in the nasal mucosa were increased in AR mice
compared with those in sham mice, but were decreased following
miR-205-5p knockdown, as determined by ELISA (Fig. 3C). These data indicated that
miR-205-5p knockdown may attenuate OVA-induced inflammatory
reaction by reducing the levels of inflammatory cells and the
production of proinflammatory cytokines.
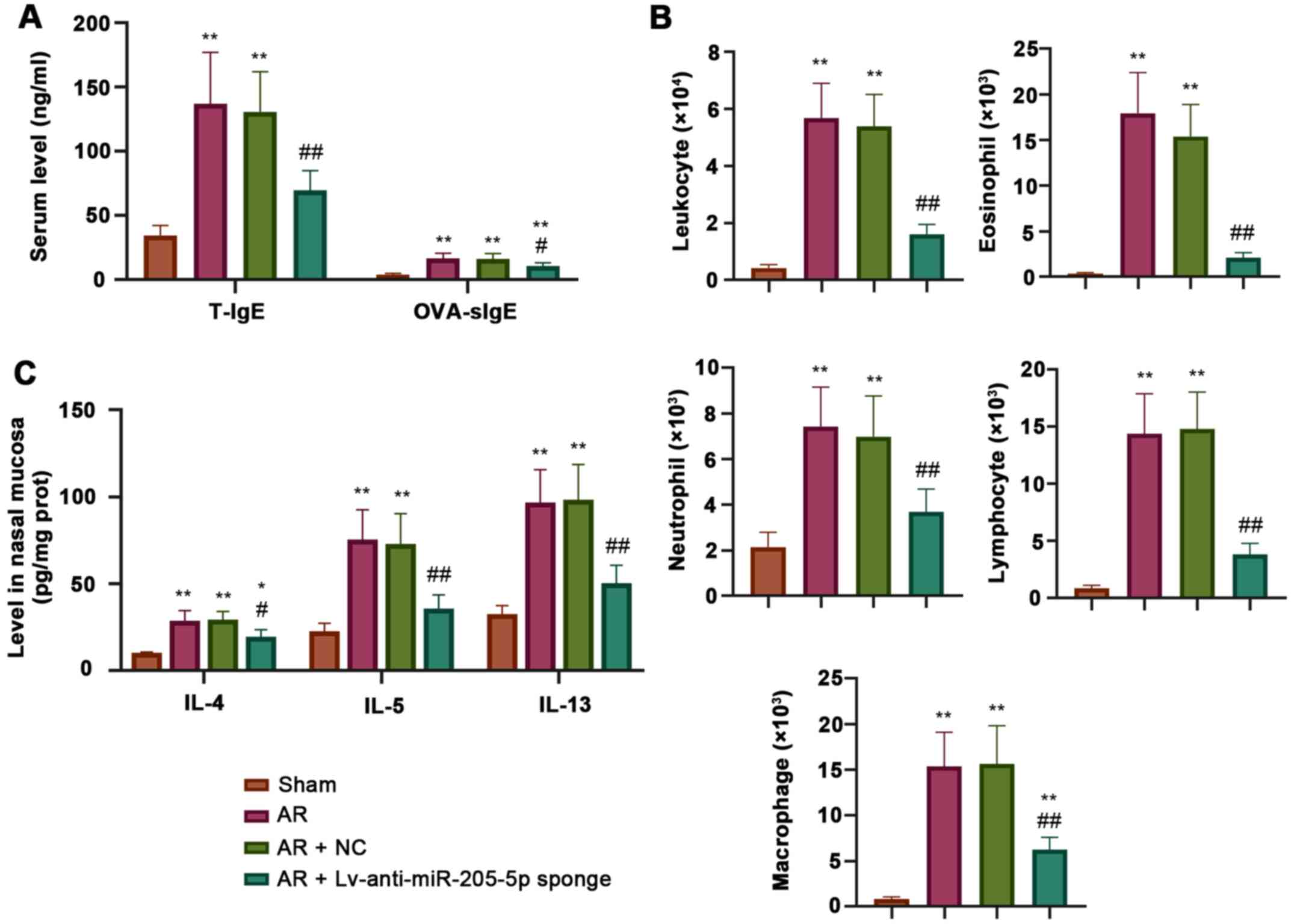 | Figure 3.miR-205-5p knockdown decreases
OVA-induced inflammatory response. (A) Levels of T-IgE and
OVA-specific IgE in serum. (B) Number of inflammatory cells,
including leukocytes, eosinophils, neutrophils, lymphocytes and
macrophages in nasal lavage fluid. (C) Levels of IL-4, IL-5 and
IL-13 in the nasal mucosa. Data are presented as the mean ± SD
(n=7). *P<0.05, **P<0.01 vs. Sham; #P<0.05,
##P<0.01 vs. AR + NC. AR, allergic rhinitis; miR,
microRNA; NC, negative control; OVA, ovalbumin; T-IgE, total IgE;
OVA-sIgE, ovalbumin-specific serum immunoglobulin E. |
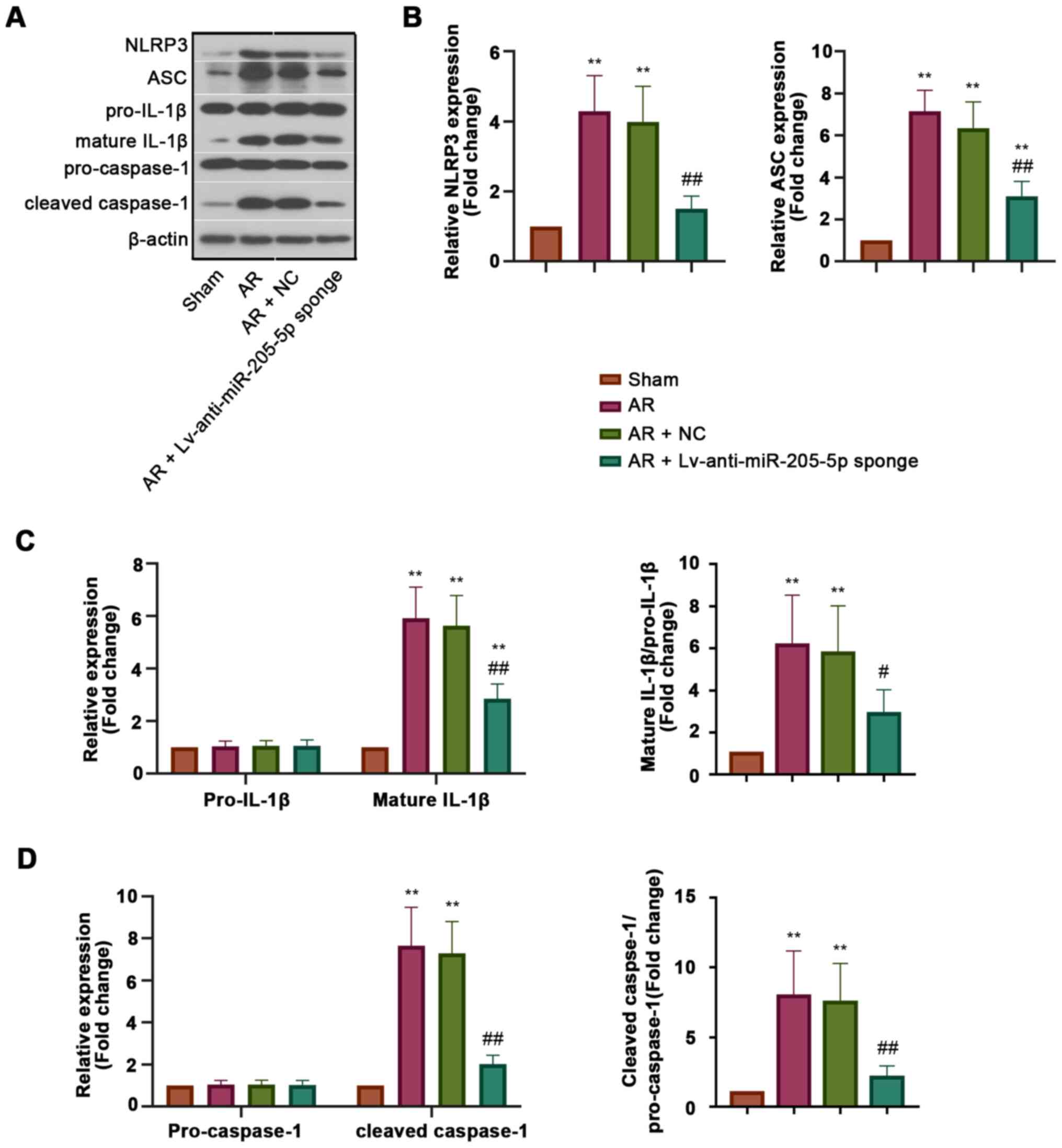
miR-205-5p knockdown inhibits NLRP3
inflammasome activation in AR
To explore the effect of miR-205-5p on NLRP3
inflammasome activation during AR, the expression levels of
proteins associated with NLRP3 inflammasome activation, including
NLRP3, apoptosisassociated specklike protein containing a CARD
(ASC), cleaved caspase-1 and mature IL-1β, were detected in the
nasal mucosa (Fig. 4A). The results
of western blot analysis revealed that the protein expression
levels of NLRP3 and ASC were increased in AR mice compared with
those in sham mice, but were inhibited by miR-205-5p knockdown
(Fig. 4B). The expression levels of
pro-caspase-1 and pro-IL-1β in all groups exhibited no significant
difference, whereas those of cleaved caspase-1 and mature IL-1β,
which were recruited and matured by activated NLRP3 inflammasome,
were decreased by miR-205-5p knockdown in AR mice (Fig. 4C and D). These results indicated
that miR-205-5p knockdown may inhibit NLRP3 inflammasome activation
in AR, which could help to explain the regulatory effect of
miR-205-5p on inflammation.
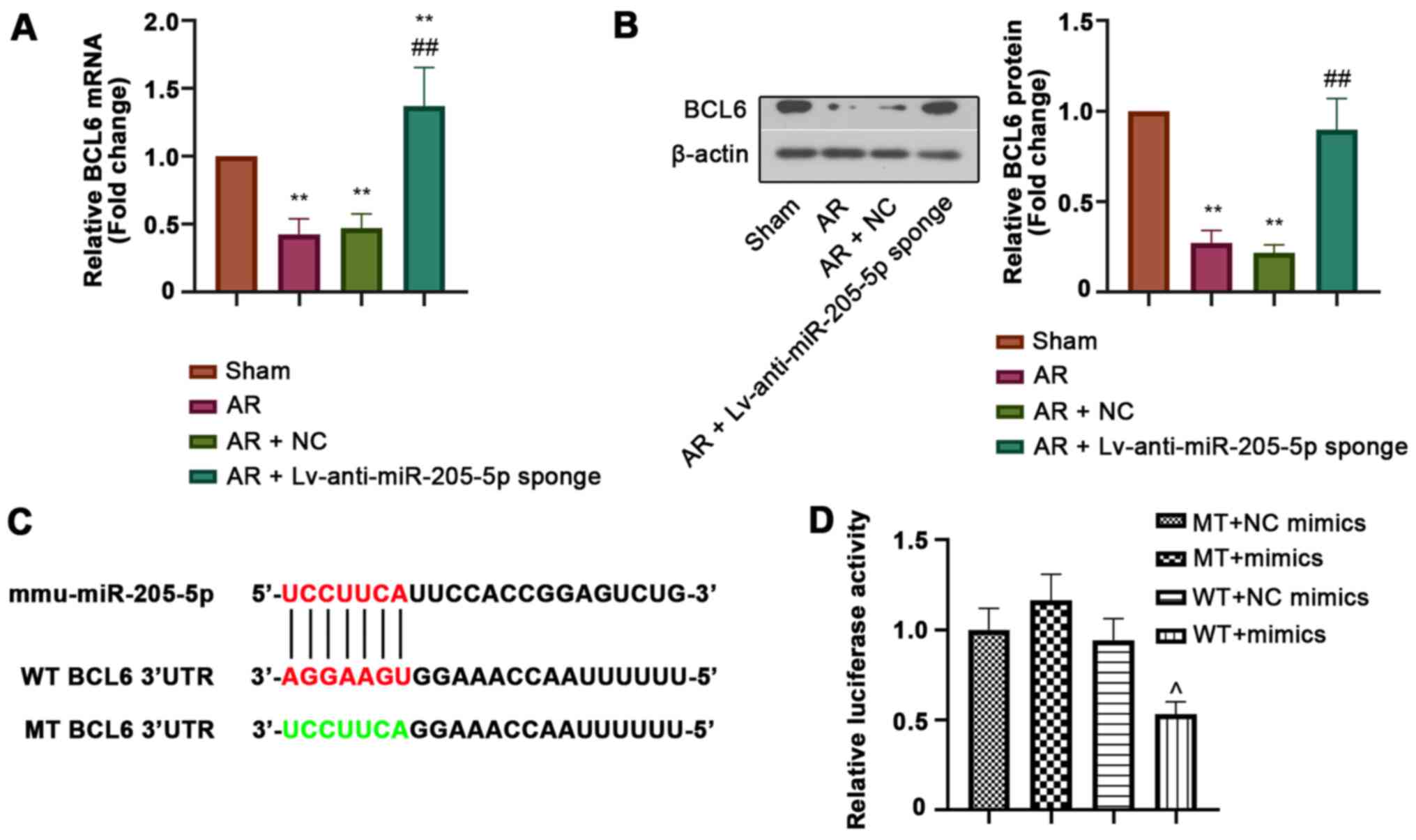
BCL6 is a target of miR-205-5p
BCL6 was predicted as a potential target gene of
miR-205-5p. The relative mRNA and protein expression levels of BCL6
in the nasal mucosa were detected by RT-qPCR and western blot
analysis, respectively. Both the mRNA and protein expression levels
of BCL6 were downregulated in the nasal mucosa of AR mice compared
with those in sham mice, but were markedly increased by miR-205-5p
knockdown (Fig. 5A and B). In
addition, luciferase reporter assay was performed to confirm the
binding of miR-205-5p to BCL6. The WT or MT 3′-UTR sequence of BCL6
was cloned into the vector and then co-transfected into cells with
miR-205-5p mimics or NC mimics (Fig.
5C). The decreased luciferase activity in WT cells transfected
with miR-205-5p mimics indicated that miR-205-5p bound to the
3′-UTR of BCL6 (Fig. 5D). These
results suggested that BCL6 was a target of miR-205-5p.
BCL6 is expressed in epithelial cells,
T cells and macrophages within the nasal mucosa
The possible type of cells expressing BCL6 within
the nasal mucosa tissue was investigated. Immunohistochemical
images showed a clear epithelial structure and indicated that BCL6
was likely expressed in the epithelial cells of mouse nasal mucosa
(Fig. 6A). The expression of BCL6
was clearly reduced in OVA-induced AR mice, but was recovered by
miR-205-5p knockdown (Fig. 6B);
this was consistent with the results of RT-qPCR and western
blotting. Furthermore, double-immunofluorescence staining was
performed for BCL6 and CD4 or F4/80 in the mouse nasal mucosa from
the AR group (Fig. 6C). The results
showed a co-localization of BCL6 and CD4+ T cells,
suggesting that BCL6 was expressed in T cells of the nasal mucosa
in the AR group. Moreover, BCL6 and F4/80 double-positive cells
indicated that BCL6 was also expressed in macrophages from the
nasal mucosa of mice in the AR group. Therefore, it was
demonstrated that BCL6 was likely expressed in epithelial and
immune cells, including T cells and macrophages, in the nasal
mucosa in a mouse model of AR. These findings suggested that the
function of BCL6 in OVA-induced inflammation may be linked to its
expression in these cells.
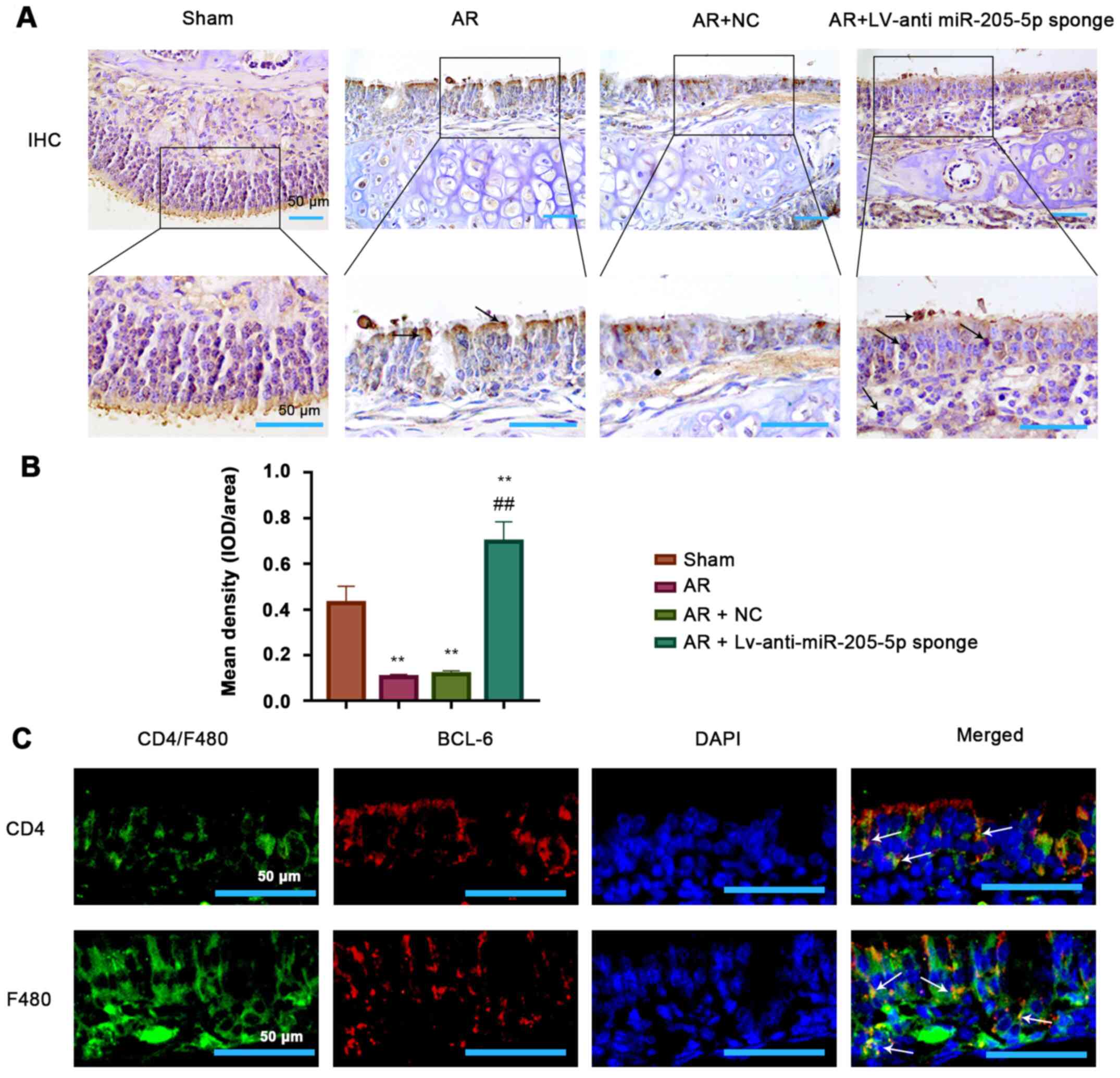 | Figure 6.Presence of BCL6 in epithelial cells,
T cells and macrophages. (A) Representative images (magnification,
×200 and ×400; scale bars, 100 and 50 µm) and (B)
semi-quantification of immunohistochemical staining for BCL6 in the
epidermis of nasal mucosa tissue. (C) Double-immunofluorescence
staining of BCL6 (red) and CD4+ T cell (green) or F4/80
macrophage (green) in the nasal mucosa tissues in the AR group
(magnification, ×400; scale bar, 50 µm). White arrows indicate
double-positive cells (yellow). **P<0.01 vs. Sham;
##P<0.01 vs. AR + NC. AR, allergic rhinitis; BCL6,
B-cell lymphoma 6; IHC, immunohistochemistry; miR, microRNA; NC,
negative control. |
Discussion
miRNA-based therapy is an emerging treatment for AR.
Certain miRNAs have been reported to be differentially expressed in
patients with AR and asthma, rendering them a diagnostic biomarker
for these diseases (36). Several
miRNAs have also been identified by RNA microarray analysis to be
differentially expressed in patients with AR compared with in
healthy participants (37). The
therapeutic effects of miRNAs on AR have also been reported.
miR-466a-3p has been shown to mitigate AR response by inhibiting
Th2 cell priming (38).
Furthermore, miR-133b has been demonstrated to alleviate allergic
inflammation in AR by targeting NLRP3 (39). Overall, these studies supported the
hypothesis that miRNAs are involved in the regulation of AR
development; however, to the best of our knowledge, little research
has been conducted on the effects of miR-205-5p on AR. The
increased level of miR-205 in the nasal mucosa of symptomatic
patients with AR was observed by Suojalehto et al (21). Consistent with this previous study,
the present study reported an increased expression of miR-205-5p in
mice with OVA-induced AR, accompanied by severe symptoms of AR.
Notably, miR-205-5p knockdown alleviated the symptoms of AR by
reducing the inflammatory response. The present study suggested
that miR-205-5p may be involved in AR development and could be
considered a novel therapeutic target for AR treatment.
miR-205-5p has been reported to stimulate MMP
activity and inflammation in abdominal aortic aneurysm development
(40). miR-205-5p has also been
shown to suppress inflammatory responses in
lipopolysaccharide-induced sepsis and lung injury following hip
fracture (23,41). The contradictory function of miR-205
in inflammation may be attributed to different pathological
environments. In the present study, an anti-inflammatory effect of
miR-205-5p knockdown was observed in AR in vivo. Multiple
inflammatory cells and cytokines are vital components of the
inflammatory response in AR. Increased infiltration of inflammatory
cells, such as leukocytes, eosinophils, neutrophils, lymphocytes
and macrophages, is usually observed in AR (42,43).
In the present study, it was revealed that the number of
inflammatory cells in NLF was decreased by miR-205-5p knockdown
following OVA challenge. The Th2 cytokines IL-4, IL-5 and IL-13
play major roles in allergic inflammation (44). In the present study, miR-205-5p
knockdown effectively reduced the serum levels of total and
OVA-specific IgE, as well as the production of IL-4, IL-5 and
IL-13, in the nasal mucosa. These findings suggested that
miR-205-5p knockdown may inhibit the inflammatory response in AR
mice.
The NLRP3 inflammasome is a signaling complex
consisting of the inflammasome sensor molecule NLRP3, the adaptor
ASC and the effector protease caspase-1. The NLRP3 inflammasome has
been well characterized and is a key regulator in the maturation of
pro-inflammatory cytokines IL-1β and IL-18 (45,46).
The activated NLRP3 recruits the adaptor ASC, resulting in the
recruitment and activation of pro-caspase-1 into its cleaved form,
which cleaves and matures IL-1β and IL-18 (47,48).
It has been reported that NLRP3 inflammasome activation may promote
AR development in an IgE-independent manner (49). In a previous study, mice treated
with MCC950, an NLRP3 inflammasome inhibitor, exhibited a reduced
OVA-induced AR response (50). The
NLRP3 inflammasome has been demonstrated to be a potential
therapeutic target for AR (51).
Therefore, the regulatory effect of miR-205-5p on NLRP3
inflammasome activation in AR was further investigated. NLRP3
inflammasome activation is represented by increased NLRP3
expression, ASC speck formation, and caspase-1 cleavage and
therefore enhanced IL-1β and IL-18 secretion (52). Initiating the transcription of the
NLRP3 gene to increase its expression is essential for activation
of the NLRP3 inflammasome (48).
Notably, AR has been found to contribute to the activation of the
NLRP3 inflammasome by increasing the expression levels of its
components, including NLRP3 and ASC, and via activation of
caspase-1, led to the maturation of inflammatory cytokines IL-1β
and IL-18 (49,53,54).
In the present study, the levels of NLRP3, ASC and cleaved
caspase-1 were decreased by miR-205-5p knockdown in the nasal
mucosa, suggesting the involvement of miR-205-5p in NLRP3
inflammasome activation in AR. The deactivation of NLRP3
inflammasome by miR-205-5p knockdown resulted in the subsequent
reduced production of proinflammatory cytokines, such as IL-1β,
which critically contribute to AR development. In addition, NLRP3
has been reported to function as a Th2 transcription factor and
promote a Th2-dependent allergic response (55). Accumulating evidence has indicated
that caspase-1 and IL-1β are involved in Th2 immune response and
asthma development (56,57). These results suggested that
miR-205-5p knockdown may alleviate the inflammatory response in AR
mice by inhibiting NLRP3 inflammasome activation.
To explore the underlying regulatory mechanism of
miR-205-5p in the development of AR, BCL6 was identified as a
candidate target gene of miR-205-p. A previous study demonstrated
that BCL6 expression was significantly decreased in the nasal
mucosa of patients with AR compared with that in healthy
individuals (29). In the present
study, the mRNA and protein expression levels of BCL6 were lower in
the nasal mucosa of AR mice compared with those in sham mice, which
was consistent with the previous finding. In addition, miR-205-5p
inhibited BCL6 expression by binding to its 3′-UTR sequence, and
miR-205-5p knockdown increased its expression. The expression of
BCL6 was detected in both CD4+ T cells and F4/80
macrophages in the nasal mucosa of OVA-sensitized mice. Notably,
BCL6 has been reported to suppress the expression of Th2
transcription factor GATA-binding protein 3 and Th2 genes including
IL-4, IL-5 and IL-13 to regulate inflammation (58). BCL6 has been suggested to be a
likely key regulator of the production of IL-1β and IL-18, which
are produced by macrophages, inducing Th2 cell differentiation in
allergic diseases. An increased percentage of macrophages producing
IL-1β has been detected in asthmatic submucosa (54). In addition, BCL6 may serve a
negative role in the regulation of key genes that predispose
patients to allergies in a wide range of cells, including T cells,
B cells, macrophages, mast cells and airway epithelial cells,
resulting in the suppressed production of Th2 cytokines and IgE;
therefore, BCL6 may act as a negative regulator in IgE-mediated
allergic response (59,60). Furthermore, BCL6 has been
demonstrated to be highly expressed in epithelial cells of the
mouse nasal mucosa, and can suppress NLRP3 transcription by binding
to the NLRP3 promoter, thus leading to attenuated inflammation in
human renal tubular epithelial cells and in vivo (31). Therefore, the upregulation of BCL6
may have contributed to NLRP3 inflammasome deactivation following
miR-205-5p knockdown in AR mice in the present study. The
expression of NLRP3 and caspase-1 has been identified in human
epithelial bronchus and cells, which are involved in asthma through
the secretion of IL-1β (61,62).
Whether BCL6 regulates the development of AR in an NLRP3
inflammasome-dependent pathway in the present study needs to be
confirmed. The current study verified the likely presence of BCL6
in epithelial cells, T cells and macrophages in the nasal mucosa in
a mouse model of AR. The function of BCL6 in AR inflammation may be
linked to its expression in these cells. Overall, the results of
the present study suggested that BCL6 may alleviate OVA-induced
inflammation through the inhibition of Th2 cell differentiation, as
indicated by decreased levels of Th2 cytokines IL-4, IL-5 and
IL-13, as well as macrophage and NLRP3 inflammasome activation.
However, the function and regulatory mechanism of BCL6 in a
specific cell type in AR remain to be investigated.
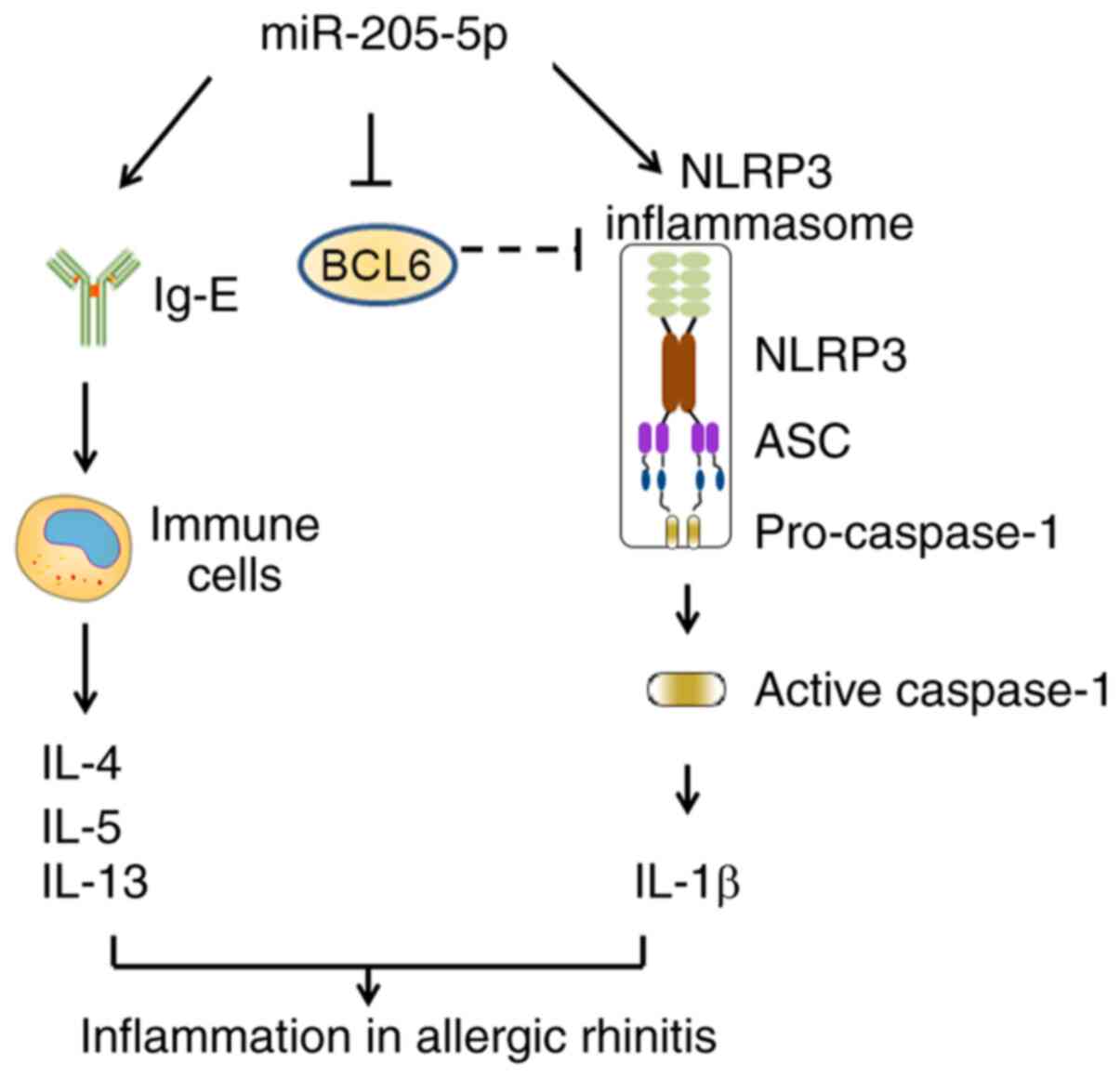
In conclusion, the present study demonstrated that
miR-205-5p knockdown may ameliorate the inflammatory response in AR
by suppressing the production of Th2 cytokines and activation of
NLRP3 inflammasome (Fig. 7). As a
target of miR-205-5p, BCL6 could serve a vital role in this
process. The present study revealed the importance of the
miR-205-5p/BCL6 axis in AR development, providing an improved
understanding of AR pathogenesis and a potential therapeutic target
for AR treatment.
Acknowledgements
Not applicable.
Funding
This work was supported by the Science and
Technology Project of Liaoning Province (grant no.
2019010183-JH8/103).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZY and QT designed the experiments; SZ and SL
performed the experiments and analyzed data; SZ wrote the paper; ZY
and QT revised the paper. SZ and SL confirm the authenticity of all
the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal studies were approved by and performed in
accordance with the Ethics Committee of the Second Affiliated
Hospital of Shenyang Medical College (approval no. 2020005).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bousquet J, Van Cauwenberge P and Khaltaev
N; Aria Workshop Group; World Health Organization, : Allergic
rhinitis and its impact on asthma. J Allergy Clin Immunol. 108
(Suppl 1):S147–S334. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maurer M and Zuberbier T: Undertreatment
of rhinitis symptoms in Europe: Findings from a cross-sectional
questionnaire survey. Allergy. 62:1057–1063. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bernstein DI, Schwartz G and Bernstein JA:
Allergic rhinitis: Mechanisms and treatment. Immunol Allergy Clin
North Am. 36:261–278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wheatley LM and Togias A: Clinical
practice. Allergic rhinitis. N Engl J Med. 372:456–463. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Greiner AN, Hellings PW, Rotiroti G and
Scadding GK: Allergic rhinitis. Lancet. 378:2112–2122. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Han X, Krempski JW and Nadeau K: Advances
and novel developments in mechanisms of allergic inflammation.
Allergy. 75:3100–3111. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Meltzer EO: Quality of life in adults and
children with allergic rhinitis. J Allergy Clin Immunol. 108 (Suppl
1):S45–S53. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schuler Iv CF and Montejo JM: Allergic
rhinitis in children and adolescents. Pediatr Clin North Am.
66:981–993. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guerra S, Sherrill DL, Martinez FD and
Barbee RA: Rhinitis as an independent risk factor for adult-onset
asthma. J Allergy Clin Immunol. 109:419–425. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mastrorilli C, Posa D, Cipriani F and
Caffarelli C: Asthma and allergic rhinitis in childhood: what's
new. Pediatr Allergy Immunol. 27:795–803. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shaaban R, Zureik M, Soussan D, Neukirch
C, Heinrich J, Sunyer J, Wjst M, Cerveri I, Pin I, Bousquet J, et
al: Rhinitis and onset of asthma: A longitudinal population-based
study. Lancet. 372:1049–1057. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bielory L: Allergic conjunctivitis and the
impact of allergic rhinitis. Curr Allergy Asthma Rep. 10:122–134.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cox L: Approach to Patients with Allergic
Rhinitis: Testing and treatment. Med Clin North Am. 104:77–94.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guilbert TW, Morgan WJ, Zeiger RS, Mauger
DT, Boehmer SJ, Szefler SJ, Bacharier LB, Lemanske RF Jr, Strunk
RC, Allen DB, et al: Long-term inhaled corticosteroids in preschool
children at high risk for asthma. N Engl J Med. 354:1985–1997.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Covar RA, Szefler SJ, Martin RJ, Sundstrom
DA, Silkoff PE, Murphy J, Young DA and Spahn JD: Relations between
exhaled nitric oxide and measures of disease activity among
children with mild-to-moderate asthma. J Pediatr. 142:469–475.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hutvágner G and Zamore PD: A microRNA in a
multiple-turnover RNAi enzyme complex. Science. 297:2056–2060.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jinek M and Doudna JA: A three-dimensional
view of the molecular machinery of RNA interference. Nature.
457:405–412. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Coskun M, Bjerrum JT, Seidelin JB and
Nielsen OH: MicroRNAs in inflammatory bowel disease - pathogenesis,
diagnostics and therapeutics. World J Gastroenterol. 18:4629–4634.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang XH, Zhang YN and Liu Z: MicroRNA in
chronic rhinosinusitis and allergic rhinitis. Curr Allergy Asthma
Rep. 14:4152014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Suojalehto H, Toskala E, Kilpeläinen M,
Majuri ML, Mitts C, Lindström I, Puustinen A, Plosila T, Sipilä J,
Wolff H, et al: MicroRNA profiles in nasal mucosa of patients with
allergic and nonallergic rhinitis and asthma. Int Forum Allergy
Rhinol. 3:612–620. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Son DJ, Jung YY, Seo YS, Park H, Lee DH,
Kim S, Roh YS, Han SB, Yoon DY and Hong JT: Interleukin-32α
inhibits endothelial inflammation, vascular smooth muscle cell
activation, and atherosclerosis by upregulating Timp3 and Reck
through suppressing microRNA-205 biogenesis. Theranostics.
7:2186–2203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu X, Chen X and Sun T: MicroRNA-205-5p
Targets HMGB1 to suppress inflammatory responses during lung injury
after hip fracture. BioMed Res Int. 2019:73048952019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Q, Zhou L, Wang L, Li S, Xu G, Gu H, Li
D, Liu M, Fang L, Wang Z, et al: Bcl6 modulates innate immunity by
controlling macrophage activity and plays critical role in
experimental autoimmune encephalomyelitis. Eur J Immunol.
50:525–536. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Béguelin W, Teater M, Gearhart MD, Calvo
Fernández MT, Goldstein RL, Cárdenas MG, Hatzi K, Rosen M, Shen H,
Corcoran CM, et al: EZH2 and BCL6 cooperate to assemble CBX8-BCOR
complex to repress bivalent promoters, mediate germinal center
formation and lymphomagenesis. Cancer Cell. 30:197–213. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Basso K and Dalla-Favera R: Roles of BCL6
in normal and transformed germinal center B cells. Immunol Rev.
247:172–183. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nurieva RI, Chung Y, Martinez GJ, Yang XO,
Tanaka S, Matskevitch TD, Wang YH and Dong C: Bcl6 mediates the
development of T follicular helper cells. Science. 325:1001–1005.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Koh B, Ulrich BJ, Nelson AS, Panangipalli
G, Kharwadkar R, Wu W, Xie MM, Fu Y, Turner MJ, Paczesny S, et al:
Bcl6 and Blimp1 reciprocally regulate ST2+ Treg-cell
development in the context of allergic airway inflammation. J
Allergy Clin Immunol. 146:1121–1136.e9. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng Y, Li XQ and Qiu QH: Detection of
differentially expressed gene of allergic rhinitis based on RT2
profiler PCR array. Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi.
31:869–872. 2017.(In Chinese). PubMed/NCBI
|
|
30
|
Hiromura Y, Kishida T, Nakano H, Hama T,
Imanishi J, Hisa Y, Mazda O, et al: IL-21 administration into the
nostril alleviates murine allergic rhinitis. J Immunol.
179:7157–7165. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen D, Xiong XQ, Zang YH, Tong Y, Zhou B,
Chen Q, Li YH, Gao XY, Kang YM and Zhu GQ: BCL6 attenuates renal
inflammation via negative regulation of NLRP3 transcription. Cell
Death Dis. 8:e31562017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cho SW, Zhang YL, Ko YK, Shin JM, Lee JH,
Rhee CS and Kim DY: Intranasal Treatment With 1,
25-Dihydroxyvitamin D3 alleviates allergic rhinitis symptoms in a
mouse model. Allergy Asthma Immunol Res. 11:267–279. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kuntz C, Wunsch A, Rosch R, Autschbach F,
Windeler J and Herfarth C: Short- and long-term results after
laparoscopic vs conventional colon resection in a tumor-bearing
small animal model. Surg Endosc. 14:561–567. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Robert R, Nail S, Marot-Leblond A, Cottin
J, Miegeville M, Quenouillere S, Mahaza C and Senet JM: Adherence
of platelets to Candida species in vivo. Infect Immun. 68:570–576.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Panganiban RP, Wang Y, Howrylak J,
Chinchilli VM, Craig TJ, August A and Ishmael FT: Circulating
microRNAs as biomarkers in patients with allergic rhinitis and
asthma. J Allergy Clin Immunol. 137:1423–1432. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shaoqing Y, Ruxin Z, Guojun L, Zhiqiang Y,
Hua H, Shudong Y and Jie Z: Microarray analysis of differentially
expressed microRNAs in allergic rhinitis. Am J Rhinol Allergy.
25:e242–e246. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen Z, Deng Y, Li F, Xiao B, Zhou X and
Tao Z: MicroRNA-466a-3p attenuates allergic nasal inflammation in
mice by targeting GATA3. Clin Exp Immunol. 197:366–375.
2019.PubMed/NCBI
|
|
39
|
Xiao L, Jiang L, Hu Q and Li Y:
MicroRNA-133b Ameliorates Allergic Inflammation and Symptom in
Murine Model of Allergic Rhinitis by Targeting Nlrp3. Cell Physiol
Biochem. 42:901–912. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim CW, Kumar S, Son DJ, Jang IH,
Griendling KK and Jo H: Prevention of abdominal aortic aneurysm by
anti-microRNA-712 or anti-microRNA-205 in angiotensin II-infused
mice. Arterioscler Thromb Vasc Biol. 34:1412–1421. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhou W, Wang J, Li Z, Li J and Sang M:
MicroRNA-205 5b inhibits HMGB1 expression in LPS-induced sepsis.
Int J Mol Med. 38:312–318. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li J, Wang B, Luo Y, Zhang Q, Bian Y and
Wang R: Resveratrol-mediated SIRT1 activation attenuates
ovalbumin-induced allergic rhinitis in mice. Mol Immunol.
122:156–162. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Benson M, Strannegård IL, Strannegård O
and Wennergren G: Topical steroid treatment of allergic rhinitis
decreases nasal fluid TH2 cytokines, eosinophils, eosinophil
cationic protein, and IgE but has no significant effect on
IFN-gamma, IL-1beta, TNF-alpha, or neutrophils. J Allergy Clin
Immunol. 106:307–312. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Barnes PJ: Pathophysiology of allergic
inflammation. Immunol Rev. 242:31–50. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kelley N, Jeltema D, Duan Y and He Y: The
NLRP3 inflammasome: An overview of mechanisms of activation and
regulation. Int J Mol Sci. 20:33282019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhen Y and Zhang H: NLRP3 inflammasome and
inflammatory bowel disease. Front Immunol. 10:2762019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jo EK, Kim JK, Shin DM and Sasakawa C:
Molecular mechanisms regulating NLRP3 inflammasome activation. Cell
Mol Immunol. 13:148–159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mangan MSJ, Olhava EJ, Roush WR, Seidel
HM, Glick GD and Latz E: Targeting the NLRP3 inflammasome in
inflammatory diseases. Nat Rev Drug Discov. 17:588–606. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yang Z, Liang C, Wang T, Zou Q, Zhou M,
Cheng Y, Peng H, Ji Z, Deng Y, Liao J, et al: NLRP3 inflammasome
activation promotes the development of allergic rhinitis via
epithelium pyroptosis. Biochem Biophys Res Commun. 522:61–67. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang W, Ba G, Tang R, Li M and Lin H:
Ameliorative effect of selective NLRP3 inflammasome inhibitor
MCC950 in an ovalbumin-induced allergic rhinitis murine model. Int
Immunopharmacol. 83:1063942020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xiao Y, Xu W and Su W: NLRP3 inflammasome:
A likely target for the treatment of allergic diseases. Clin Exp
Allergy. 48:1080–1091. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao W, Ma L, Cai C and Gong X: Caffeine
inhibits NLRP3 inflammasome activation by suppressing MAPK/NF-κB
and A2aR signaling in LPS-induced THP-1 macrophages. Int J Biol
Sci. 15:1571–1581. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Elliott EI and Sutterwala FS: Initiation
and perpetuation of NLRP3 inflammasome activation and assembly.
Immunol Rev. 265:35–52. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Sousa AR, Lane SJ, Nakhosteen JA, Lee TH
and Poston RN: Expression of interleukin-1 beta (IL-1beta) and
interleukin-1 receptor antagonist (IL-1ra) on asthmatic bronchial
epithelium. Am J Respir Crit Care Med. 154:1061–1066. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bruchard M, Rebé C, Derangère V, Togbé D,
Ryffel B, Boidot R, Humblin E, Hamman A, Chalmin F, Berger H, et
al: The receptor NLRP3 is a transcriptional regulator of TH2
differentiation. Nat Immunol. 16:859–870. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Iwata A, Nishio K, Winn RK, Chi EY,
Henderson WR Jr and Harlan JM: A broad-spectrum caspase inhibitor
attenuates allergic airway inflammation in murine asthma model. J
Immunol. 170:3386–3391. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ben-Sasson SZ, Hu-Li J, Quiel J,
Cauchetaux S, Ratner M, Shapira I, Dinarello CA and Paul WE: IL-1
acts directly on CD4 T cells to enhance their antigen-driven
expansion and differentiation. Proc Natl Acad Sci USA.
106:7119–7124. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Sawant DV, Sehra S, Nguyen ET, Jadhav R,
Englert K, Shinnakasu R, Hangoc G, Broxmeyer HE, Nakayama T,
Perumal NB, et al: Bcl6 controls the Th2 inflammatory activity of
regulatory T cells by repressing Gata3 function. J Immunol.
189:4759–4769. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Takeda N, Arima M, Tsuruoka N, Okada S,
Hatano M, Sakamoto A, Kohno Y and Tokuhisa T: Bcl6 is a
transcriptional repressor for the IL-18 gene. J Immunol.
171:426–431. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Arima M, Fukuda T and Tokuhisa T: Role of
the transcriptional repressor BCL6 in allergic response and
inflammation. World Allergy Organ J. 1:115–122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hirota JA, Hirota SA, Warner SM,
Stefanowicz D, Shaheen F, Beck PL, Macdonald JA, Hackett TL, Sin
DD, Van Eeden S, et al: The airway epithelium nucleotide-binding
domain and leucine-rich repeat protein 3 inflammasome is activated
by urban particulate matter. J Allergy Clin Immunol.
129:1116–25.e6. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Birrell MA and Eltom S: The role of the
NLRP3 inflammasome in the pathogenesis of airway disease. Pharmacol
Ther. 130:364–370. 2011. View Article : Google Scholar : PubMed/NCBI
|