Introduction
Allergic rhinitis is a non-infectious disease of the
nasal mucosa, which is characterized by paroxysmal sneezing, a
runny nose, nasal itching and nasal congestion (1). Patients with allergic rhinitis may
have an atopic constitution, usually showing familial aggregation
with some associated genetic alterations (2). With regards to external factors,
exposure to allergens, such as mites, pollen, fungal spores and
animal dander, often serves as the trigger for the development of
allergic rhinitis (3,4). Drug therapy, such as
H1-antihistamines and intranasal corticosteroids, may
control the symptoms of allergic rhinitis (5); however, there is currently no
definitive cure (6). This disease
although not fatal, does hinder general health, for example
symptoms include sleep disruption and poor concentration, which can
interfere with work and education (1,7).
Hence, medical researchers continue to search for a cure for
allergic rhinitis to improve the quality of life of patients.
Formononetin is an active ingredient of a
traditional Chinese herb, Radix Astragali, and may also be
extracted from the inflorescence and flowered branches and leaves
of the leguminous plant, Trifolium pratense L. and the whole
leaf of Ononis spinosa L (8,9). The
anticancer properties of formononetin have been investigated in
numerous studies (10–12). An increasing number of studies have
also begun to focus on its potential in treating other disease
types. For example, formononetin was previously found to protect
against airway inflammation and oxidative stress in a mouse model
of murine allergic asthma (13).
Formononetin was also shown to alleviate atopic dermatitis by
activating G protein-coupled estrogen receptors and upregulating
A20 expression (14). However, to
the best of our knowledge, there has been no report to date on
whether formononetin exerts therapeutic effects in allergic
rhinitis. Sirtuin 1 (SIRT1) is an universal gene regulator that
affects multiple inflammation-related signaling pathways and has
also been reported to play a role in attenuating ovalbumin-induced
allergic symptoms in vivo (15). Upregulation of the nuclear factor
erythroid 2-related factor 2 (Nrf2)/heme oxygenase (HO)-1 signaling
pathway by mangiferin was previously demonstrated to be associated
with the improvement of the ovalbumin-induced inflammatory response
in mice with allergic rhinitis (16). Moreover, the regulatory effect of
formononetin on SIRT1 expression has already been identified in
various types of disease. For example, formononetin was shown to
ameliorate cholestasis by regulating hepatic SIRT1 and peroxisome
proliferator-activated receptor α expression (17).
IL-13 is a typical T helper cell 2 cytokine and
serves a prominent role in the regulation of numerous types of
allergic disease by activating its receptor and the associated
STAT6 (18). IL-13 is often used to
simulate pathological changes observed in a number of airway
allergic diseases, such as eosinophil recruitment, mucus cell
metaplasia, subepithelial fibrosis and smooth muscle hypertrophy
(19). On this basis, IL-13
induction was used in the present study to establish an in
vitro allergic rhinitis model.
The present study hypothesized that formononetin may
exert protective effects in an allergic rhinitis model established
with IL-13 via regulating the SIRT1/Nrf2 signaling pathway to
inhibit inflammatory cytokine secretion and mucus formation. The
present study was undertaken to verify this hypothesis, with the
aim of providing a novel agent for the effective treatment or
alleviation of allergic rhinitis.
Materials and methods
Cell culture and treatment
The JME/CF15 human nasal epithelial cell line was
acquired from The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences. The cells were cultured in DMEM/F12
(cat. no. 11320033; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (cat. no. 12484010; Gibco; Thermo Fisher
Scientific, Inc.), 2 mM L-glutamine (cat. no. A2916801; Gibco;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin (cat. no.
V900929; Sigma-Aldrich; Merck KGaA) and 100 mg/ml streptomycin
(cat. no. HY-B0472; MedChemExpress). The cells were maintained in
95% air and 5% CO2 at 37°C.
The allergic rhinitis model was established by
stimulating the JME/CF15 cells with 10 ng/ml IL-13 (cat. no.
HY-P7033; MedChemExpress) at 37°C for 30 min as previously
described (20), or pretreatment
with formononetin [cat. no. CN00152; ChemeGen (Shanghai)
Biotechnology Co., Ltd.] at doses of 0.1, 1 and 10 µM (14,21) or
EX527 (SIRT1 inhibitor; cat. no. E7034; Sigma-Aldrich; Merck KGaA)
at a dose of 10 µM at 37°C for 24 h (22). Normal JME/CF15 cells without any
treatment were used as the model group.
Cell Counting Kit-8 (CCK-8) assay
The cytotoxicity of formononetin was detected using
a CCK-8 assay (cat. no. C0037; Beyotime Institute of Biotechnology)
as previously described (23).
Briefly, 100 µl cell suspension was added into a 96-well plate
using a density of 2×103 cells/well, with incubation at
37°C for 24 h. Cells were then treated accordingly, and after 24 h,
10 µl CCK-8 solution was added to each well. Subsequently, the
cells were incubated with CCK-8 solution at 37°C for 1.5 h. The
absorbance was measured at a wavelength of 450 nm using a
microplate reader.
ELISA
ELISAs were performed to examine the proinflammatory
cytokine secretion in IL-13-stimulated JME/CF15 cells as previously
described (24). The levels of
histamine, IgE, TNF-α, IL-1β, IL-6, granulocyte-macrophage
colony-stimulating factor (GM-CSF) and eotaxin were detected using
histamine (cat. no. ab213975; Abcam), IgE (cat. no. ab195216;
Abcam), TNF-α (cat. no. PT518; Beyotime Institute of
Biotechnology), IL-1β (cat. no. PI305; Beyotime Institute of
Biotechnology), IL-6 (cat. no. PI330; Beyotime Institute of
Biotechnology) ELISA kits, GM-CSF (cat. no. PG355; Beyotime
Institute of Biotechnology) and eotaxin (cat. no. PC115; Beyotime
Institute of Biotechnology), respectively. The optical density
value was measured at a wavelength of 450 nm using an ELISA
detector.
Western blotting
Whole cell protein extracts were extracted using
RIPA lysis buffer, while nuclear and cytoplasmic proteins were
extracted using Nuclear and Cytoplasmic Protein Extraction Kit
(cat. no. P0027; Beyotime Institute of Biotechnology), and all
protein samples were quantified using the BCA method. The sample
proteins (20 µg/lane) were separated via 10% SDS-PAGE and
subsequently transferred onto PVDF membranes. The membrane was
removed from the transfer apparatus and soaked in TBS with 10%
Tween-20 (TBST) washing buffer (Shanghai Aladdin Biochemical
Technology Co., Ltd.) twice for 15 min each time. The membrane was
then blocked with 10% non-fat dried milk freshly diluted in TBST
and rocked on a rotating shaker for 15 min at room temperature.
After rinsing with TBST, the membranes were incubated with the
following primary antibodies diluted in 1% non-fat dried milk at
4°C overnight: Anti-phosphorylated (p)-NF-κB p65 (1:1,000; cat. no.
ab239882; Abcam), anti-NF-κB p65 (1:1,000; cat. no. ab207297;
Abcam), anti-cyclooxygenase 2 (Cox-2; 1:1,000; cat. no. 12282; Cell
Signaling Technology, Inc.), anti-mucin 5AC oligomeric
mucus/gel-forming (MUC5AC; 1:20,000; cat. no. ab198294; Abcam),
anti-Nrf2 (1:1,000; cat. no. ab62352; Abcam), anti-SIRT1 (1:1,000;
cat. no. ab189494; Abcam), anti-β-actin (1:1,000; cat. no. ab8227;
Abcam) and anti-lamin B1 (1:1,000; cat. no. ab229025; Abcam).
Following the primary antibody incubation, the membranes were
rinsed three times in TBST and incubated with a HRP-conjugated
secondary antibody (1:2,000; cat. no. ab97051; Abcam) diluted in 1%
non-fat dried milk for 30 min at room temperature. Excess secondary
antibody was then removed from the membrane with three rinses in 20
ml TBST for 5 min each time. Protein bands were analyzed using a
standard chemiluminescence detection reagent (cat. no. P0018S;
Beyotime Institute of Biotechnology) and ImageJ software (version
1.48v; National Institutes of Health) was used for
semi-quantification as previously described (23).
Reverse transcription-quantitative PCR
(RT-qPCR)
mRNA expression levels of MUC5AC in IL-13-stimulated
JME/CF15 cells were detected using RT-qPCR as previously described
(23). Total RNA from JME/CF15
cells was extracted using TRIzol® reagent (cat. no.
15596018; Invitrogen; Thermo Fisher Scientific, Inc.) and the
quality was assessed using a NanoDrop 2000 spectrophotometer
(Thermo Fisher Scientific, Inc.). Total RNA was reverse transcribed
into cDNA using a Sensiscript RT kit (cat. no. 205213; Qiagen,
Inc.) according to the manufacturer's protocol. qPCR was
subsequently performed using a TaqMan Universal Master mix II (cat.
no. 4440048; Applied Biosystems; Thermo Fisher Scientific, Inc.) on
an ABI Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The following thermocycling conditions were used
for the qPCR: Initial denaturation for 3 min at 95°C; followed by
40 cycles of denaturation for 15 sec at 95°C, and annealing and
extension for 30 sec at 60°C. The primers used for the qPCR were as
follows: MUC5AC forward, 5′-GCTTCCTGCTCCGAGATGT-3′ and reverse,
5′-AAGACGCAGCCCTCATAGAA-3′; and GAPDH forward,
5′-CACCCACTCCTCCACCTTTG-3′ and reverse, 5′-CCACCACCCTGTTGCTGTAG-3′.
The relative expression levels were calculated using the
2−ΔΔCq method (25).
Immunofluorescence (IF)
IF was employed to further verify the expression of
MUC5AC in IL-13-stimulated JME/CF15 cells as previously described
(26). Briefly, following fixation
with 4% paraformaldehyde (Shanghai Aladdin Biochemical Technology
Co., Ltd.) for 10 min at room temperature, the cells were incubated
with 0.5% Triton X-100 (cat. no. HFH10; Invitrogen; Thermo Fisher
Scientific, Inc.) at 25°C for 20 min. Subsequently, 5% BSA (cat.
no. B265993; Shanghai Aladdin Biochemical Technology Co., Ltd.)
diluted in PBS (Beyotime Institute of Biotechnology) was used to
block the cells at room temperature for 1 h before incubation with
an anti-MUC5AC antibody (1:250) overnight at 4°C. Following the
primary antibody incubation, the cells were washed with PBS and
incubated with a FITC-conjugated goat anti-rabbit IgG secondary
antibody (1:200; cat. no. P0186; Beyotime Institute of
Biotechnology) at 25°C for 1 h, followed by staining with DAPI
(MedChemExpress) for 15 min at room temperature. Stained cells were
visualized using a fluorescence microscope (magnification,
×200).
Statistical analysis
Independent experiments were performed at least
three times. The data are presented as the mean ± SD and were
analyzed using a one-way ANOVA followed by a Tukey's post hoc test.
GraphPad Prism 6 software (GraphPad Software, Inc.) was used for
the statistical analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Formononetin does not affect JME/CF15
cell viability
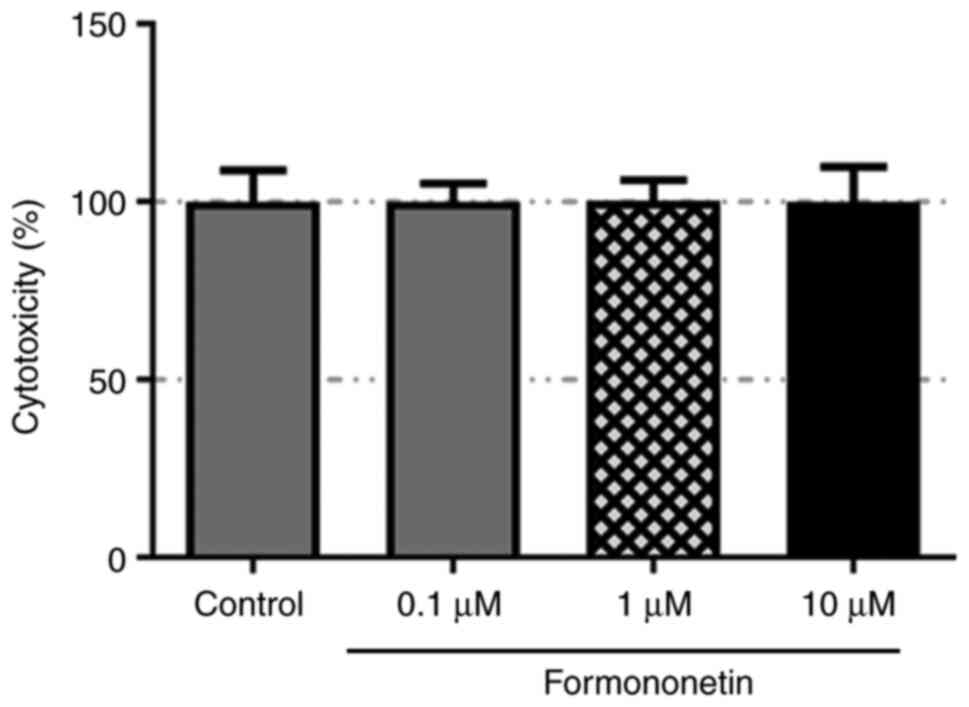
The cytotoxicity of formononetin was examined using
a CCK-8 assay, which demonstrated that the viability of JME/CF15
cells was not significantly altered following formononetin
treatment at doses of 0.1, 1 or 10 µM (Fig. 1). These results suggested that
formononetin may exert no cytotoxic effect on JME/CF15 cells.
Formononetin inhibits inflammatory
cytokine secretion in IL-13-stimulated JME/CF15 cells
According to the results of the ELISAs (Fig. 2A and B), the levels of the
immunoregulatory autacoid histamine, the allergy index IgE and the
proinflammatory cytokines, TNF-α, IL-1β and IL-6, were
significantly upregulated in IL-13-stimulated JME/CF15 cells
compared with the control group, and were subsequently
downregulated in a dose-dependent manner following pretreatment
with 0.1, 1 or 10 µM formononetin. Furthermore, IL-13 stimulation
significantly elevated the levels of GM-CSF and eotaxin in JME/CF15
cells compared with the control group, which were then
dose-dependently reduced by formononetin treatment at all doses
(Fig. 2C). Western blotting
revealed that the expression levels of inflammation-related Cox-2
and p-NF-κB p65/NF-κB p65 were significantly upregulated in
IL-13-stimulated JME/CF15 cells compared with the control group,
while their expression levels were subsequently downregulated
following formononetin treatment (Fig.
2D). These results suggested an inhibitory effect of
formononetin on the secretion of inflammatory cytokines in
IL-13-stimulated JME/CF15 cells.
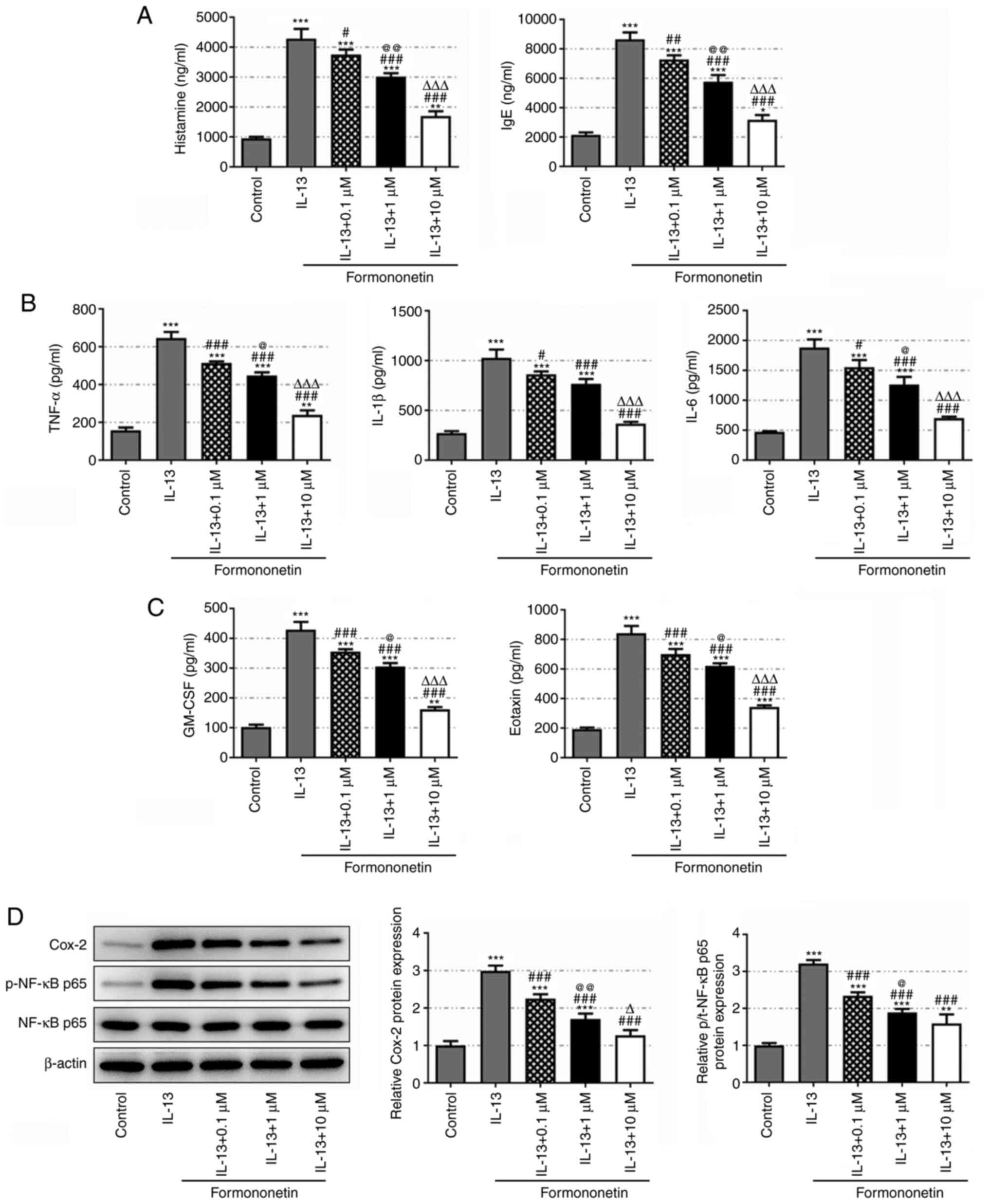 | Figure 2.Formononetin inhibits inflammatory
cytokine secretion in IL-13-stimulated JME/CF15 cells.
Concentrations of (A) histamine and IgE, (B) TNF-α, IL-1β and IL-6
and (C) GM-CSF and eotaxin in IL-13-stimulated JME/CF15 cells
treated with 0, 0.1, 1 or 10 µM formononetin were detected using
ELISAs. (D) Relative expression levels of Cox-2 and p-NF-κB
p65/NF-κB p65 in IL-13-stimulated JME/CF15 cells treated with 0,
0.1, 1 or 10 µM formononetin were detected using western blotting.
*P<0.05, **P<0.01, ***P<0.001 vs. control;
#P<0.05, ##P<0.01,
###P<0.001 vs. IL-13; @P<0.05,
@@P<0.01 vs. IL-13 + 0.1 µM; ∆P<0.05,
∆∆∆P<0.001 vs. IL-13 + 1 µM. GM-CSF,
granulocyte-macrophage colony-stimulating factor; Cox-2,
cyclooxygenase 2; p-, phosphorylated. |
Formononetin inhibits mucus formation
in IL-13-stimulated JME/CF15 cells
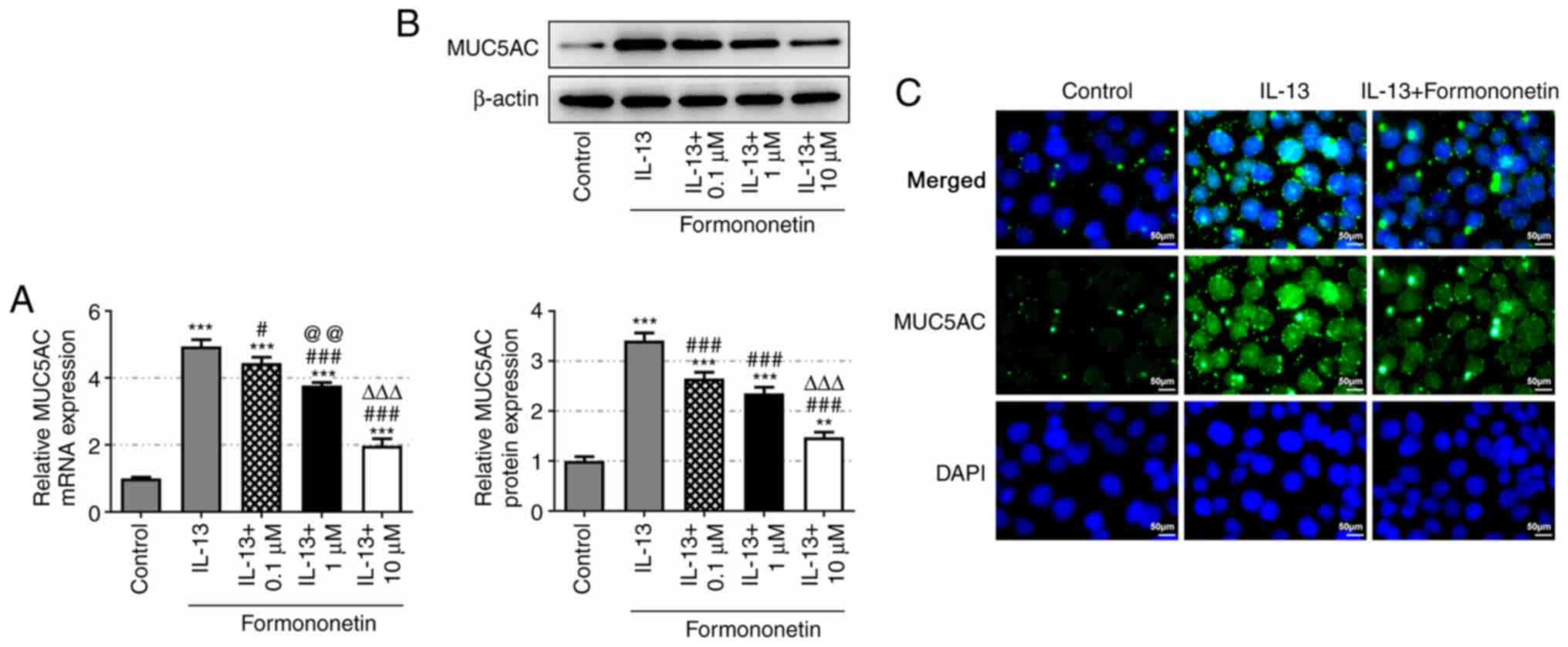
Overexpression of human MUC5AC has been reported to
contribute to the formation of mucus in airway inflammation
(27,28); therefore, its protein expression
levels were detected in the allergic rhinitis model in the present
study. It was observed that, while IL-13 induced high expression of
MUC5AC, pretreatment with formononetin effectively reduced MUC5AC
expression levels in IL-13-stimulated JME/CF15 cells (Fig. 3A and B). Formononetin at 10 µM was
associated with the most significant reduction in MUC5AC expression
and was thus selected for use in subsequent experiments. As shown
in Fig. 3C, the fluorescence
intensity of MUC5AC was increased in the IL-13 group compared with
the control group, while it was markedly weakened in the IL-13 +
formononetin group. These results indicated that formononetin may
prevent the formation of mucus in IL-13-stimulated JME/CF15
cells.
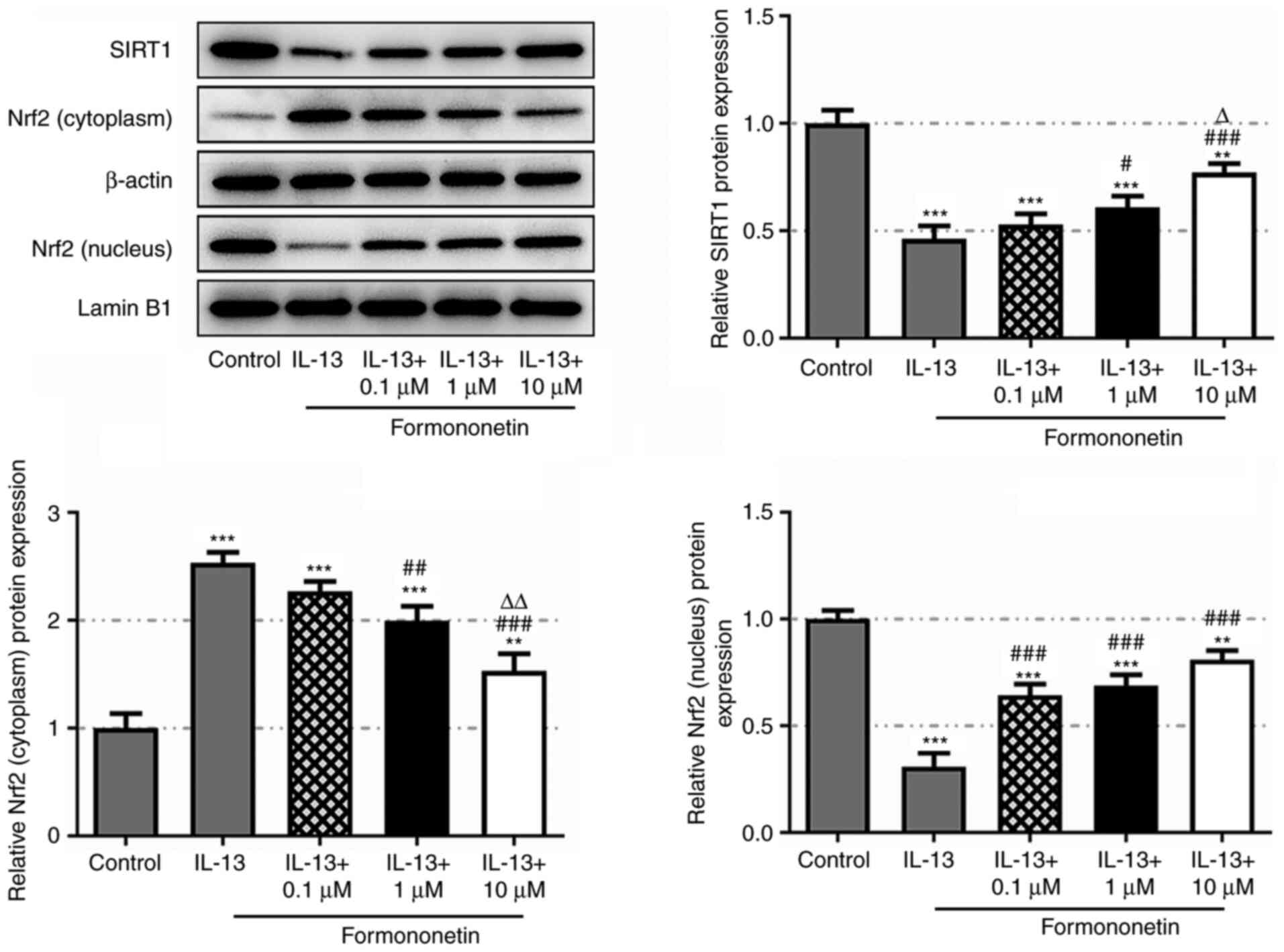
Formononetin activates the SIRT1/Nrf2
signaling pathway
To understand the mechanism of action of
formononetin, it was investigated whether there was an interaction
between formononetin and the SIRT1/Nrf2 signaling pathway. Western
blotting revealed that the expression levels of SIRT1 and nucleic
Nrf2 were downregulated in IL-13-stimulated JME/CF15 cells compared
with the control group, and were subsequently dose-dependently
upregulated following treatment with formononetin (Fig. 4). Conversely, the expression levels
of cytoplasmic Nrf2 exhibited the opposite trends. These findings
indicated that formononetin may activate the SIRT1/Nrf2 signaling
pathway.
SIRT1 inhibitor EX527 reverses the
effects of formononetin on IL-13-stimulated JME/CF15 cells
To verify the role of the SIRT1/Nrf2 axis in the
mechanism of action of formononetin, the expression levels of SIRT1
in IL-13-stimulated JME/CF15 cells were inhibited using EX527. A
significant rise in the levels of histamine, IgE, TNF-α, IL-1β and
IL-6 was observed in IL-13-stimulated formononetin-treated JME/CF15
cells following SIRT1 inhibition (Fig.
5A and B). EX527 treatment also increased the levels of GM-CSF
and eotaxin, which were suppressed by formononetin treatment in
IL-13-stimulated JME/CF15 cells (Fig.
5C). Furthermore, while the expression levels of Cox-2 and
p-NF-κB p65/NF-κB p65 were downregulated by formononetin treatment
in IL-13-stimulated JME/CF15 cells, inhibiting SIRT1 with EX527
partially reversed their expression levels (Fig. 5D). The expression levels of nucleic
Nrf2 were downregulated, while the expression levels of cytoplasmic
Nrf2 were upregulated in IL-13-stimulated JME/CF15 cells. After
pretreatment with formononetin, the expression level of nucleic
Nrf2 was upregulated and cytoplasmic Nrf2 expression was
downregulated, which was partially reversed following EX527
treatment (Fig. 5E). Similarly,
EX527 treatment rescued the expression levels of MUC5AC, which were
downregulated by formononetin treatment in IL-13-stimulated
JME/CF15 cells (Fig. 5F and G).
These results indicated that formononetin may inhibit the secretion
of proinflammatory cytokines and mucus formation in
IL-13-stimulated JME/CF15 cells by activating the SIRT1/Nrf2
signaling pathway.
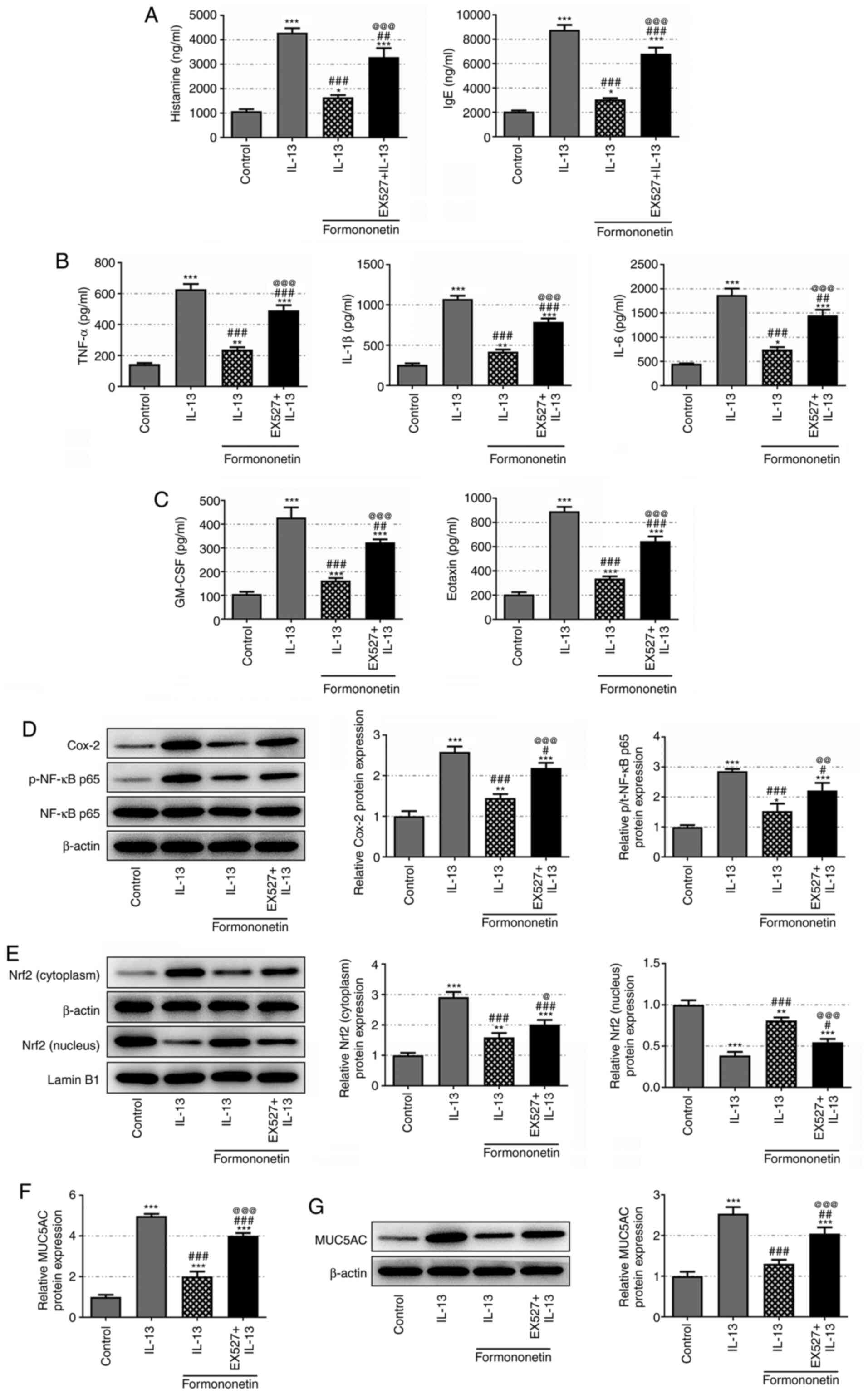 | Figure 5.Sirtuin 1 inhibitor EX527 reverses
the effects of formononetin on IL-13-stimulated JME/CF15 cells.
Concentrations of (A) histamine and IgE, (B) TNF-α, IL-1β and IL-6
and (C) GM-CSF and eotaxin in JME/CF15 cells treated with IL-13,
IL-13 + formononetin or IL-13 + formononetin + EX527 were detected
using ELISAs. (D) Relative protein expression levels of Cox-2 and
p-NF-κB p65/NF-κB p65 in JME/CF15 cells treated with IL-13, IL-13 +
formononetin or IL-13 + formononetin + EX527 were detected using
western blotting. (E) Relative protein expression levels of nucleic
and cytoplasmic Nrf2 in JME/CF15 cells treated with IL-13, IL-13 +
formononetin or IL-13 + formononetin + EX527 were detected using
western blotting. Relative mRNA and protein expression levels of
MUC5AC in JME/CF15 cells treated with IL-13, IL-13 + formononetin
or IL-13 + formononetin + EX527 were detected using (F) reverse
transcription-quantitative PCR and (G) western blotting,
respectively. *P<0.05, **P<0.01, ***P<0.001 vs. control;
#P<0.05, ##P<0.01,
###P<0.001 vs. IL-13; @P<0.05,
@@P<0.01, @@@P<0.001 vs. IL-13 +
formononetin. GM-CSF, granulocyte-macrophage colony-stimulating
factor; Cox-2, cyclooxygenase 2; p-, phosphorylated; Nrf2, nuclear
factor erythroid 2-related factor 2; MUC5AC, mucin 5AC oligomeric
mucus/gel-forming. |
Discussion
Allergic rhinitis, which currently affects ~40% of
the world population, is a global health concern that is associated
with a notable socioeconomic burden and may lead to serious
complications, including asthma and otitis media (29). Allergic rhinitis is an atopic
disorder of the nasal mucosa that is mainly characterized by
IgE-mediated histamine release and involves a variety of
immunocompetent cells and cytokines, such as pathogenic memory T
helper 2 cells, IL-4, IL-5 and IL-13, which are triggered following
exposure of the atopic individual to allergens (30,31).
Risk factors for allergic rhinitis are present at all ages
(32), and include genetic factors
(33,34), environmental pollution (35) and environmental allergen exposure
(36). The symptoms of the disease
may be effectively controlled through standardized treatment
(examples include treatment with intranasal corticosteroids,
H1-antihistamine and leukotriene receptor antagonist) (5) and care (37); however, long-term treatment poses a
heavy economic burden (38).
Formononetin, a type of phytoestrogen derived from
the medicinal plant Trifolium pretense L., has been proven
to be an active component with promising anticancer properties, as
evidenced by previous research (39,40).
In addition to its anticancer properties, formononetin has also
been found to exert antioxidant and anti-inflammatory effects in
various types of disease. For example, previous studies have
reported that formononetin played a role in preventing allergic
diseases and that it could effectively attenuate atopic dermatitis
through regulation of G-protein-coupled estrogen receptor and A20
expression (14,41). It was also shown that formononetin
regulated NF-κB signaling to inhibit the inflammatory response and
promote the angiogenesis of the gastric mucosa in mice with gastric
ulcers (42). However, to the best
of our knowledge, whether formononetin has a beneficial effect on
allergic rhinitis has yet to be reported. Excessive release of
histamine induced by allergen exposure is commonly known to be an
active contributor to the occurrence of allergic rhinitis (43). IgE is an important mediator of the
endogenous release of histamine, the level of which is also closely
associated with asthma, food allergies and allergic rhinitis
(44–47). It was previously demonstrated that
formononetin decreased the levels of histamine and proinflammatory
cytokines to alleviate allergic inflammation (21). In the present study, a cellular
model of allergic rhinitis was established using IL-13 stimulation,
in which high levels of histamine, IgE and proinflammatory
cytokines, TNF-α, IL-1β and IL-6, were detected. However, treatment
with formononetin markedly decreased these levels in a
dose-dependent manner. Previous studies have reported high
concentrations of GM-CSF in patients with nasal allergy, which was
also found to be associated with aggravated rhinorrhea (48,49). A
more recent study also demonstrated the value of GM-CSF in
assessing chronic rhinitis due to its positive correlation with
eosinophilic inflammation (50).
The results of the present study revealed high levels of both
GM-CSF and eotaxin in IL-13-stimulated JME/CF15 cells, while
treatment with formononetin was demonstrated to effectively
suppress GM-CSF and eotaxin levels. Upregulated Cox-2 expression
has been proven to be a cancer-related biomarker and is also
closely associated with inflammation (51,52).
Some of the anti-inflammatory effects of formononetin were
discovered to be mediated through its suppression of Cox-2
expression in neurological lesions and kidney injury, among others
(53,54). NF-κB p65 is a member of the NF-κB
family that acts as a master regulator of inflammation and is
widely found in animal cells (55).
Formononetin was reported to inactivate NF-κB p65 to obstruct
osteoclastogenesis and protect against liver injury (56,57).
Consistent with previous findings, downregulated expression levels
of Cox-2 and p-NF-κB p65 were also found in the present study
following formononetin treatment in IL-13-stimulated JME/CF15
cells. Taken together, these changes suggested that formononetin
may inhibit IL-13-induced proinflammatory cytokine secretion in
JME/CF15 cells.
MUC5AC is a gel-forming mucin that is normally
present in human airways, the high expression of which leads to the
oversecretion of mucus and was recently identified in the airway
mucus of patients with severe coronavirus-19 disease (58). In the present study, IL-13
significantly upregulated the protein expression levels of MUC5AC
in JME/CF15 cells, which were markedly suppressed following
formononetin treatment, as evidenced by the results of the RT-qPCR,
western blotting and IF experiments. These findings indicated that
formononetin may inhibit mucus formation in allergic rhinitis.
In addition to the effects of formononetin on
allergic rhinitis, the mechanism underlying its effects was also
investigated. SIRT1 is a well-known regulator of chronic
inflammatory responses, which has been investigated in the context
of the alleviation of asthma as well as allergic rhinitis (15,59).
Wu et al (60) found that
SIRT1 activation upregulated Nrf2 expression and inhibited the
NF-κB signaling pathway to relieve sepsis-induced inflammation and
apoptosis. Furthermore, it was demonstrated in a previous study
that formononetin served as a SIRT1 activator and improved diabetic
neuropathy (61). In addition,
formononetin was found to upregulate SIRT1 expression and inhibit
lipopolysaccharide-induced high mobility group box 1 release,
thereby inhibiting the cellular inflammatory response (62). SIRT1 activated by resveratrol could
also alleviate ovalbumin-induced allergic rhinitis in mice
(63). SIRT1 knockdown alleviated
the symptoms and inflammatory indicators of allergic rhinitis in
HIF1αCD11c−/− mice (64). Activation of the Nrf2/HO-1 signaling
pathway could suppress allergic rhinitis activity (65). In addition, mangiferin attenuated
allergic rhinitis by enhancing the Nrf2/HO-1 signaling pathways
(16). The present study revealed
that formononetin may activate the SIRT1/Nrf2 signaling pathway.
Normally, Nrf2 and Kelch like ECH associated protein (Keap1) bind
as a dimer in the cytoplasm, and when stimulated, Nrf2 dissociates
from Keap1 and translocates into the nucleus, thereby inducing the
expression of encoded related enzymes, such as HO-1 (66). In the present study, formononetin
was shown to upregulate the expression of Nrf2 in the nucleus and
promote the nuclear translocation of Nrf2, and thus exert an
effect. Moreover, inhibiting SIRT1 with EX527 largely reversed the
effects of formononetin on IL-13-induced inflammation and mucus
secretion in JME/CF15 cells, and reduced the nuclear translocation
of Nrf2, further verifying the role of SIRT1/Nrf2 signaling in the
mechanism of action of formononetin. However, the present
conclusion is based solely on cell experiments and further animal
experiments should be conducted to validate the current study
findings.
In conclusion, the findings of the present study
provided evidence to suggest that formononetin may ameliorate
IL-13-induced proinflammatory cytokine secretion and mucus
formation in JME/CF15 cells by activating the SIRT1/Nrf2 signaling
pathway. These findings are significant as they indicate the
curative potential of formononetin in the treatment of allergic
rhinitis, and it may be considered as a novel therapeutic strategy
for this disease.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JH conceived and designed the experiments in the
present study. JH, XC and AX performed the experiments. JH and XC
analyzed the data. JH and AX wrote the manuscript. All authors read
and approved the final manuscript. JH and XC confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hoyte FCL and Nelson HS: Recent advances
in allergic rhinitis. F1000Res. 7:F1000Faculty Rev-1333. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dávila I, Mullol J, Ferrer M, Bartra J,
del Cuvillo A, Montoro J, Jáuregui I, Sastre J and Valero A:
Genetic aspects of allergic rhinitis. J Investig Allergol Clin
Immunol. 19 (Suppl 1):S25–S31. 2009.PubMed/NCBI
|
|
3
|
Incorvaia C, Cavaliere C, Frati F and
Masieri S: Allergic rhinitis. J Biol Regul Homeost Agents. 32
(Suppl 1):S61–S66. 2018.PubMed/NCBI
|
|
4
|
Numminen J: Allergic rhinitis. Duodecim.
133:473–478. 2017.PubMed/NCBI
|
|
5
|
Brożek JL, Bousquet J, Agache I, Agarwal
A, Bachert C, Bosnic-Anticevich S, Brignardello-Petersen R,
Canonica GW, Casale T, Chavannes NH, et al: Allergic rhinitis and
its impact on asthma (ARIA) guidelines-2016 revision. J Allergy
Clin Immunol. 140:950–958. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bao H, Si D, Gao L, Sun H, Shi Q, Yan Y,
Damchaaperenlei D, Li C, Yu M and Li Y: Acupuncture for the
treatment of allergic rhinitis: A systematic review protocol.
Medicine (Baltimore). 97:e137722018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Meltzer EO: Allergic rhinitis: Burden of
illness, quality of life, comorbidities, and control. Immunol
Allergy Clin North Am. 36:235–248. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gampe N, Darcsi A, Lohner S, Béni S and
Kursinszki L: Characterization and identification of isoflavonoid
glycosides in the root of Spiny restharrow (Ononis spinosa L.) by
HPLC-QTOF-MS, HPLC-MS/MS and NMR. J Pharm Biomed Anal. 123:74–81.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ong SKL, Shanmugam MK, Fan L, Fraser SE,
Arfuso F, Ahn KS, Sethi G and Bishayee A: Focus on formononetin:
Anticancer potential and molecular targets. Cancers (Basel).
11:6112019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yao JN, Zhang XX, Zhang YZ, Li JH, Zhao
DY, Gao B, Zhou HN, Gao SL and Zhang LF: Discovery and anticancer
evaluation of a formononetin derivative against gastric cancer
SGC7901 cells. Invest New Drugs. 37:1300–1308. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang J, Liu L, Wang J, Ren B, Zhang L and
Li W: Formononetin, an isoflavone from Astragalus membranaceus
inhibits proliferation and metastasis of ovarian cancer cells. J
Ethnopharmacol. 221:91–99. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang L, Gong Y, Wang S and Gao F:
Anti-colorectal cancer mechanisms of formononetin identified by
network pharmacological approach. Med Sci Monit. 25:7709–7714.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yi L, Cui J, Wang W, Tang W, Teng F, Zhu
X, Qin J, Wuniqiemu T, Sun J, Wei Y and Dong J: Formononetin
attenuates airway inflammation and oxidative stress in murine
allergic asthma. Front Pharmacol. 11:5338412020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yuan W, Chen Y, Zhou Y, Bao K, Yu X, Xu Y,
Zhang Y, Zheng J, Jiang G and Hong M: Formononetin attenuates
atopic dermatitis by upregulating A20 expression via activation of
G protein-coupled estrogen receptor. J Ethnopharmacol.
266:1133972021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan Y, Liu Q, Zhao J, Tang H and Sun J:
SIRT1 attenuates murine allergic rhinitis by downregulated HMGB
1/TLR4 pathway. Scand J Immunol. 87:e126672018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Piao CH, Fan YJ, Nguyen TV, Song CH and
Chai OH: Mangiferin alleviates ovalbumin-induced allergic rhinitis
via Nrf2/HO-1/NF-κB signaling pathways. Int J Mol Sci. 21:34152020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang S, Wei L, Xia R, Liu L, Chen Y, Zhang
W, Li Q, Feng K, Yu M, Zhang W, et al: Formononetin ameliorates
cholestasis by regulating hepatic SIRT1 and PPARα. Biochem Biophys
Res Commun. 512:770–778. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wills-Karp M: Interleukin-13 in asthma
pathogenesis. Immunol Rev. 202:175–190. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Matsukura S, Stellato C, Georas SN,
Casolaro V, Plitt JR, Miura K, Kurosawa S, Schindler U and
Schleimer RP: Interleukin-13 upregulates eotaxin expression in
airway epithelial cells by a STAT6-dependent mechanism. Am J Respir
Cell Mol Biol. 24:755–761. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang B, Gao Y, Zheng G, Ren X, Sun B, Zhu
K, Luo H, Wang Z and Xu M: Platycodin D inhibits
interleukin-13-induced the expression of inflammatory cytokines and
mucus in nasal epithelial cells. Biomed Pharmacother. 84:1108–1112.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu N and An J: Formononetin ameliorates
mast cell-mediated allergic inflammation via inhibition of
histamine release and production of pro-inflammatory cytokines. Exp
Ther Med. 14:6201–6206. 2017.PubMed/NCBI
|
|
22
|
Wang Y, Che J, Zhao H, Tang J and Shi G:
Paeoniflorin attenuates oxidized low-density lipoprotein-induced
apoptosis and adhesion molecule expression by autophagy enhancement
in human umbilical vein endothelial cells. J Cell Biochem.
120:9291–9299. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia M, Chen X, Liu J and Chen J: PTEN
promotes apoptosis of H2O2-injured rat nasal epithelial cells
through PI3K/Akt and other pathways. Mol Med Rep. 17:571–579.
2018.PubMed/NCBI
|
|
24
|
Sun B, Wang B and Xu M: Esculetin inhibits
histamine-induced expression of inflammatory cytokines and mucin in
nasal epithelial cells. Clin Exp Pharmacol Physiol. 46:821–827.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kwak S, Choi YS, Na HG, Bae CH, Song SY
and Kim YD: Fipronil upregulates inflammatory cytokines and MUC5AC
expression in human nasal epithelial cells. Rhinology. 58:66–73.
2020.PubMed/NCBI
|
|
27
|
Shah SA, Ishinaga H, Hou B, Okano M and
Takeuchi K: Effects of interleukin-31 on MUC5AC gene expression in
nasal allergic inflammation. Pharmacology. 91:158–164. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang X, Li Y, Luo D, Wang X, Zhang Y, Liu
Z, Zhong N, Wu M and Li G: Lyn regulates mucus secretion and MUC5AC
via the STAT6 signaling pathway during allergic airway
inflammation. Sci Rep. 7:426752017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng L, Chen J, Fu Q, He S, Li H, Liu Z,
Tan G, Tao Z, Wang D, Wen W, et al: Chinese society of allergy
guidelines for diagnosis and treatment of allergic rhinitis.
Allergy Asthma Immunol Res. 10:300–353. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kakli HA and Riley TD: Allergic rhinitis.
Prim Care. 43:465–475. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Meng Y, Wang C and Zhang L: Recent
developments and highlights in allergic rhinitis. Allergy.
74:2320–2328. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schuler Iv CF and Montejo JM: Allergic
rhinitis in children and adolescents. Pediatr Clin North Am.
66:981–993. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Noguchi E, Shibasaki M, Arinami T, Takeda
K, Maki T, Miyamoto T, Kawashima T, Kobayashi K and Hamaguchi H:
Evidence for linkage between asthma/atopy in childhood and
chromosome 5q31-q33 in a Japanese population. Am J Respir Crit Care
Med. 156:1390–1393. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Andiappan AK, Wang de Y, Anantharaman R,
Suri BK, Lee BT, Rotzschke O, Liu J and Chew FT: Replication of
genome-wide association study loci for allergic rhinitis and house
dust mite sensitization in an Asian population of ethnic Chinese in
Singapore. J Allergy Clin Immunol. 131:1431–1433.e8. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Teng B, Zhang X, Yi C, Zhang Y, Ye S, Wang
Y, Tong DQ and Lu B: The association between ambient air pollution
and allergic rhinitis: Further epidemiological evidence from
Changchun, Northeastern China. Int J Environ Res Public Health.
14:2262017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li CW, Chen DH DH, Zhong JT, Lin ZB, Peng
H, Lu HG, Yang Y, Yin J and Li TY: Epidemiological characterization
and risk factors of allergic rhinitis in the general population in
Guangzhou City in China. PLoS One. 9:e1149502014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Akhouri S, House SA and Doerr C: Allergic
rhinitis (nursing). StatPearls; Treasure Island (FL): 2021
|
|
38
|
Bernstein DI, Schwartz G and Bernstein JA:
Allergic rhinitis: Mechanisms and treatment. Immunol Allergy Clin
North Am. 36:261–278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang XS, Guan SY, Liu A, Yue J, Hu LN,
Zhang K, Yang LK, Lu L, Tian Z, Zhao MG and Liu SB: Anxiolytic
effects of formononetin in an inflammatory pain mouse model. Mol
Brain. 12:362019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tay KC, Tan LT, Chan CK, Hong SL, Chan KG,
Yap WH, Pusparajah P, Lee LH and Goh BH: Formononetin: A review of
its anticancer potentials and mechanisms. Front Pharmacol.
10:8202019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li L, Wang Y, Wang X, Tao Y, Bao K, Hua Y,
Jiang G and Hong M: Formononetin attenuated allergic diseases
through inhibition of epithelial-derived cytokines by regulating
E-cadherin. Clin Immunol. 195:67–76. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yi L, Lu Y, Yu S, Cheng Q and Yi L:
Formononetin inhibits inflammation and promotes gastric mucosal
angiogenesis in gastric ulcer rats through regulating NF-κB
signaling pathway. J Recept Signal Transduct Res. Oct 26–2020.(Epub
ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Taylor-Clark T: Histamine in allergic
rhinitis. Adv Exp Med Biol. 709:33–41. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hanzlikova J, Sedlacek D, Liska M,
Gorcikova J, Vlas T, Amiramini S, Panzner P and Maly M: Histamine
increases the level of IFNγ produced by HIV-1 specific CTLs and
this production depends on total IgE level. J Immunol Methods.
375:1–6. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gould HJ and Sutton BJ: IgE in allergy and
asthma today. Nat Rev Immunol. 8:205–217. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Barni S, Liccioli G, Sarti L, Giovannini
M, Novembre E and Mori F: Immunoglobulin E (IgE)-mediated food
allergy in children: Epidemiology, pathogenesis, diagnosis,
prevention, and management. Medicina (Kaunas). 56:1112020.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bayar Muluk N, Bafaqeeh SA and Cingi C:
Anti-IgE treatment in allergic rhinitis. Int J Pediatr
Otorhinolaryngol. 127:1096742019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fireman P: Cytokines and allergic
rhinitis. Allergy Asthma Proc. 17:175–178. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ohkubo K, Ikeda M, Pawankar R, Gotoh M,
Yagi T and Okuda M: Mechanisms of IL-6, IL-8, and GM-CSF release in
nasal secretions of allergic patients after nasal challenge.
Rhinology. 36:156–161. 1998.PubMed/NCBI
|
|
50
|
Peric A, Spadijer-Mirkovic C,
Matkovic-Jozin S, Jovancevic L and Vojvodic D:
Granulocyte-macrophage colony-stimulating factor production and
tissue eosinophilia in chronic rhinitis. Int Arch Otorhinolaryngol.
20:364–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
López DE and Ballaz SJ: The role of brain
cyclooxygenase-2 (Cox-2) beyond neuroinflammation: Neuronal
homeostasis in memory and anxiety. Mol Neurobiol. 57:5167–5176.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cui J and Jia J: Natural COX-2 inhibitors
as promising anti-inflammatory agents: An update. Curr Med Chem.
28:3622–3646. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li Z, Dong X, Zhang J, Zeng G, Zhao H, Liu
Y, Qiu R, Mo L and Ye Y: Formononetin protects TBI rats against
neurological lesions and the underlying mechanism. J Neurol Sci.
338:112–117. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Aladaileh SH, Hussein OE, Abukhalil MH,
Saghir SAM, Bin-Jumah M, Alfwuaires MA, Germoush MO, Almaiman AA
and Mahmoud AM: Formononetin upregulates Nrf2/HO-1 signaling and
prevents oxidative stress, inflammation, and kidney injury in
methotrexate-induced rats. Antioxidants (Basel). 8:4302019.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pradère JP, Hernandez C, Koppe C, Friedman
RA, Luedde T and Schwabe RF: Negative regulation of NF-κB p65
activity by serine 536 phosphorylation. Sci Signal. 9:ra852016.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Huh JE, Lee WI, Kang JW, Nam D, Choi DY,
Park DS, Lee SH and Lee JD: Formononetin attenuates
osteoclastogenesis via suppressing the RANKL-induced activation of
NF-κB, c-Fos, and nuclear factor of activated T-cells cytoplasmic 1
signaling pathway. J Nat Prod. 77:2423–2431. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liao L, Huang L, Wei X, Yin L, Wei X and
Li T: Bioinformatic and biochemical studies of formononetin against
liver injure. Life Sci. 272:1192292021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Okuda K, Chen G, Subramani DB, Wolf M,
Gilmore RC, Kato T, Radicioni G, Kesimer M, Chua M, Dang H, et al:
Localization of secretory mucins MUC5AC and MUC5B in normal/healthy
human airways. Am J Respir Crit Care Med. 199:715–727. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang YZ, Wu QJ, Yang X, Xing XX, Chen YY
and Wang H: Effects of SIRT1/Akt pathway on chronic inflammatory
response and lung function in patients with asthma. Eur Rev Med
Pharmacol Sci. 23:4948–4953. 2019.PubMed/NCBI
|
|
60
|
Wu Z, Chen J, Zhao W, Zhuo CH and Chen Q:
Inhibition of miR-181a attenuates sepsis-induced inflammation and
apoptosis by activating Nrf2 and inhibiting NF-κB pathways via
targeting SIRT1. Kaohsiung J Med Sci. 37:200–207. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Oza MJ and Kulkarni YA: Formononetin
ameliorates diabetic neuropathy by increasing expression of SIRT1
and NGF. Chem Biodivers. 17:e20001622020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hwang JS, Kang ES, Han SG, Lim DS, Paek
KS, Lee CH and Seo HG: Formononetin inhibits
lipopolysaccharide-induced release of high mobility group box 1 by
upregulating SIRT1 in a PPARδ-dependent manner. PeerJ. 6:e42082018.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li J, Wang B, Luo Y, Zhang Q, Bian Y and
Wang R: Resveratrol-mediated SIRT1 activation attenuates
ovalbumin-induced allergic rhinitis in mice. Mol Immunol.
122:156–162. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Niu Y, Wang J, Li Z, Yao K, Wang L and
Song J: HIF1α deficiency in dendritic cells attenuates symptoms and
inflammatory indicators of allergic rhinitis in a SIRT1-dependent
manner. Int Arch Allergy Immunol. 181:585–593. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Van Nguyen T, Piao CH, Fan YJ, Shin DU,
Kim SY, Song HJ, Song CH, Shin HS and Chai OH: Anti-allergic
rhinitis activity of α-lipoic acid via balancing Th17/Treg
expression and enhancing Nrf2/HO-1 pathway signaling. Sci Rep.
10:125282020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Jung KA and Kwak MK: The Nrf2 system as a
potential target for the development of indirect antioxidants.
Molecules. 15:7266–7291. 2010. View Article : Google Scholar : PubMed/NCBI
|