Introduction
Strokes are the main cause of long-term
disabilities, the second leading cause of cardiovascular
disease-related deaths in the United States and the fifth leading
cause of death among all residents in the country according to a
report from the American Heart Association in 2019 (1). Ischemic stroke, a brain injury
caused by insufficient blood supply, accounted for 79.1 and 64.9%
of the global prevalence and incidence, respectively, of all
strokes in 2017 (2). Although a
previous investigation revealed that there were a maximum of 430
potentially useful stroke drug candidates between 1995 and 2015,
only 19 (4%), including aspirin, dipyridamole, atenolol, ramipril,
hydrochlorothiazide, Polycap and simvastatin, have been used
clinically worldwide (3).
Notably, a combination of dipyridamole and aspirin has been
reported to decrease ischemic stroke recurrence (4). The combination drug Polycap,
containing aspirin, hydrochlorothiazide, ramipril, atenolol and
simvastatin, might prevent stroke in high-risk subjects (5,6).
Therefore, combination therapy for ischemic stroke requires further
research.
DL-3-n-butylphthalide (NBP) is a natural product
extracted from celery, which acts as a neuroprotective agent
against ischemic brain damage (3). It is currently in clinical trials,
registered and approved by the China Food and Drug Administration
(3,7,8).
Furthermore, as a well-known free radical scavenger,
3-methyl-1-phenyl-2-pyrazolin-5-one (edaravone) also works as a
neuroprotective agent and is recommended for patients with acute
cerebral strokes in Japan, China, India and other countries, such
as those in Europe (9–12). Both NBP + edaravone and a hybrid
compound of a ring-opening derivative of NBP + edaravone (compound
10b) exhibit increased protective effects against brain damage
compared with NBP or edaravone alone in rats with
ischemia-reperfusion (13). These
drugs may be used for ischemic stroke treatment (13,14). The results of previous research
indicate that compound 10b exerts neuroprotective effects by
improving mitochondrial function (13). However, the mechanisms underlying
the neuroprotective effects of NBP combined with edaravone have not
yet been fully elucidated.
Recently, the neurovascular unit (NVU) has attracted
the attention of researchers as it emphasizes the importance of
communication among neurons, endothelial cells and astrocytes,
instead of only blood vessels or neurons, in ischemic injuries
(15). The NVU contributes to
disease development and responses; therefore, it could be a
therapeutic target (16,17). To the best of our knowledge,
whether NBP combined with edaravone targets the whole NVU and
protects against multiple cell death is unknown. Therefore, to
investigate the effects of NBP combined with edaravone on various
neurological aspects, the present study evaluated brain infarct
volume using 2,3,5-triphenyltetrazolium chloride (TTC) staining,
the neurological deficits in mice, and cell damage using
hematoxylin-eosin (HE) and Nissl staining. The dysfunction of the
major NVU components, including neurons, endothelial cells and
astrocytes, was also studied via immunofluorescence analysis in the
mice models with middle cerebral artery occlusion (MCAO) and
reperfusion. Furthermore, the effects of NBP combined with
edaravone on apoptosis-related proteins were determined using
western blotting.
Materials and methods
Ethics statement
Ethical permission was obtained from the Animal
Ethics Committee of North China University of the Science and
Technology (approval no. 2019068; Tangshan, China), which records
and regulates all research activities. The approval from the Animal
Ethics Committee included the permission to use mice under
anesthesia or euthanasia, and all experimental procedures were
conducted in strict accordance with recommendations in the Guide
for the Care and Use of Laboratory Animals of the National
Institutes of Health (18). Pre-
and post-surgery pain management was maintained via anesthesia with
3% isoflurane in all animals to ensure compliance with guidelines
set forth by the North China University of the Science and
Technology (Tangshan, China).
Mouse groups and drug
administration
Male C57BL/6 mice (weight, 20–25 g; age, 12 weeks)
were purchased from Shanghai Jiesijie Experimental Animal Co., Ltd.
(http://www.jsj-lab.com/; Grade II; certificate
no. 2020027). Mice (n=3 per cage) were raised under controlled
conditions with a 12-h light/dark cycle at room temperature
(21–23°C) and 40–60% humidity with free access to water and food. A
total of 39 mice were randomly divided into three groups (n=13 for
each group): i) Sham operation control; ii) MCAO and reperfusion;
and iii) NBP + edaravone MCAO (NBP + Edaravone). NBP (40 mg/kg;
batch no. 2019122001; CSPC Pharmaceutical Co. Ltd. (http://en.e-cspc.com/index.html) and edaravone (6
mg/kg; batch no. 2020010702; China National Medicines Guorui
Biomedical Technology Co., Ltd. (https://www.guorui.com.cn/wzsy) were administered by
intraperitoneal injection at 0 and 4 h after reperfusion. The
dosage was determined according to the manufacturers' instructions
and previous experiments (19,20). Briefly, from each of the three
groups (n=13 per group), 4 mice were used for neurological deficit
scores and infarct volume, while 3 mice each were used for HE/Nissl
staining, immunofluorescence analysis and protein extraction for
western blotting.
Anesthesia and euthanasia
Mice were anesthetized using box induction with 3%
isoflurane (cat. no. R510-22-8; batch no. 20191222; RWD Life
Science Co., Ltd. (https://www.rwdstco.com/) for ~3 min and then
maintained on 1.5% isoflurane in medical grade oxygen via a
facemask from a small animal anesthesia machine (cat. no. R500; RWD
Life Science Co., Ltd.). The body temperatures of the mice were
measured using a laser Doppler flowmeter (moorVMS-LDF2; Moor
Instruments, Ltd.) during the whole surgical procedure, and this
was required to be maintained at ~37±0.5°C with a heating blanket,
and the blood pressure was monitored with a small animal blood
pressure monitor (BP-2010A; RWD Life Science Co., Ltd.).
Before euthanasia, the mice were carefully examined
for the last health checkup and then exposed to 5% isoflurane for 5
min. Finally, the mice were sacrificed with dislocation of cervical
vertebra, and the criteria to judge the death of animals were
continuous absence of spontaneous breathing for 2–3 min, with no
blink reflex and no blood pressure.
Development of mice
ischemia-reperfusion model
The MCAO operation was performed by skilled
experimenters and lasted for ~10 min for each animal. After the
mice were anesthetized, the right common carotid artery was
inserted with a rounded tip 4-0 surgical monofilament nylon suture
(MSMC21B120PK50; RWD Life Science Co., Ltd.), and then the suture
was carefully advanced ~11 mm (from the bifurcation of the common
carotid artery) into the middle cerebral artery origin. A laser
Doppler flowmeter (moorVMS-LDF2) was adopted to confirm the
decrease of the middle cerebral artery blood flow immediately after
the occlusion to <70% of the basic cerebral blood flow. Those
mice with reduced blood flow to <70% of pre-ischemia levels were
used for further study. After a 1-h occlusion, the suture was
removed to obtain blood reperfusion, the wound was sutured and the
animal was kept breathing in pure medical oxygen for ~5 min to
recover from anesthesia and then finally transferred to the
individual home cage with the heating blanket. After the operation,
the animals were observed and recorded every 2 h, including
breathing, activity, diet and body temperature measured by a
portable infrared remote sensing thermometer (DKHIRT; Wuhan Dikai
Optoelectronics Technology Co., Ltd.). The sham mice were similarly
treated, although without focal cerebral ischemia-reperfusion.
There were no animal mortalities during the formal experiment,
while 2 of 10 mice died at ~24 h after the operation due to brain
edema in the pre-experiment. All results of the pre-experiment were
excluded from statistical analysis.
Neurological deficit scores
The neurological deficit scores were assessed for 24
h following reperfusion. The mice were scored according to the
method described by Bederson et al (21): Mice with no neurological symptoms
were scored as 0; when unable to flex the left forepaw fully, as 1;
when rotating while crawling and unable to move the contralateral
side, as 2; if unable to walk unaided, as 3; and if unconscious, as
4. The mice were euthanized as aforementioned at 24 h after
reperfusion.
Measurements of the infarct
volume
The mice were euthanized as aforementioned at 24 h
after reperfusion and the brains were extracted and stored at −20°C
for 10 min. Subsequently, the brain tissues were cut into
2-mm-thick slices and maintained in 0.25% TTC for 30 min at 37°C.
The slices were then fixed with 4% paraformaldehyde at 4°C for 24
h. The brain infarct volumes were measured by an experienced rater
who was blinded to the design of this experiment.
HE staining and Nissl staining
Parts of the brain tissues were used for HE
staining. The brains were fixed with 4% paraformaldehyde overnight
at 4°C, then embedded in paraffin, sliced at 5 µm and stained in
hematoxylin aqueous solution for 10 min and alcohol eosin staining
solution for 2 min at 25°C. Morphological changes and histological
observations were performed using an IX81 light microscope (Olympus
Corporation). Some brain tissue sections were placed in 60°C
incubator and dyed with 1% toluidine blue for 40 min for the Nissl
staining after the same preparatory process.
Immunofluorescence analysis
Although actual cell activity cannot be revealed
using immunofluorescence, markers for different protein activity
can be assessed. Therefore, an astrocyte marker glial fibrillary
acidic protein (GFAP), a neuronal marker neuronal nuclei (NeuN),
and an endothelial marker platelet and endothelial cell adhesion
molecule 1 (CD31) were analyzed using immunofluorescence
staining.
At room temperature, 4% paraformaldehyde-fixed,
paraffin-embedded sections at 10-µm thickness were placed on
pre-cleaned and positively charged microscope slides and were
heated in a tissue-drying oven for 45 min at 60°C, and then
deparaffinization and rehydration procedures were conducted. The
slides were washed twice in xylene for 3 min each time and in
xylene 1:1 with 100% ethanol for 3 min, twice in 100% ethanol for 3
min each, and twice in 95% ethanol for 3 min each, in 70% ethanol
for 3 min and in 50% ethanol for 3 min. Slides were rinsed gently
with running distilled water for 5 min at room temperature, and
then antigen retrieval was conducted. Slides were boiled in 0.01 M
sodium citrate buffer (pH 6.0) at 100°C for 15–20 min, then the
slides were removed from the heat and allowed to stand at room
temperature in buffer for 20 min. Then, the slides were rinsed
twice with TBS-Tween-20 (TBST, with 20% Tween 20) for 5 min at room
temperature. Slices were blocked using 5% normal goat serum (cat.
no. ab7481; Abcam) for 2 h at room temperature, followed by
overnight incubation at 4°C with the following primary antibodies:
Mouse anti-GFAP (dilution, 1:200; cat. no. ab7260; Abcam), rabbit
anti-NeuN (dilution, 1:500; cat. no. ABN78; MilliporeSigma) and
mouse anti-CD31 (dilution, 1:500; cat. no. ab24590; Abcam).
Subsequently, after rinsing with PBS, the slices were incubated
with fluorescein isothiocyanate-labeled anti-rabbit IgG (dilution,
1:800; cat. no. ab7171; Abcam) and tetramethylrhodamine-conjugated
anti-mouse IgG (dilution, 1:800; cat. no. ab6668; Abcam) at room
temperature for 2 h. The nuclei were counterstained with 1 µg/ml
DAPI (MilliporeSigma) 10 min at room temperature before mounting,
and images were obtained with an Olympus light microscope and
analyzed using Image-Pro Plus 6.0 software (Media Cybernetics,
Inc.).
Western blotting
The animals were re-anesthetized as aforementioned
at 24 h after reperfusion, and brain tissues were obtained from the
ipsilateral hemisphere. The cortex tissues were cut into small
samples and mixed on ice. Subsequently, ~150 mg of the tissue mass
was collected for mitochondrial protein extraction, while the rest
was used for total protein extraction. The protein extraction of
both the total and the mitochondrial fractions was performed
according to the instructions of the Mitochondrial Protein
Extraction kit (cat. no. AR0156; Wuhan Boster Biological
Technology, Ltd.) and the Total Protein Extraction kit for Animal
Cultured Cells/Tissues (cat. no. BB-3101; BEST BIO Technical Co.,
Ltd.; http://www.bestbio.com.cn/search?q=BB-3101). The
protein concentration was quantified using the BCA method (BCA
Protein Assay kit; cat. no.P0010S; Beyotime Institute of
Biotechnology), and an equal amount of protein (20 µg) was loaded
for 10% SDS-PAGE. After electrophoresis, proteins were transferred
onto a nitrocellulose membrane, followed by blocking in 10% non-fat
milk at room temperature for 30 min. Subsequently, the membrane was
incubated overnight at 4°C with gentle shaking with the following
primary antibodies: rabbit anti-synaptophysin (SYP; dilution,
1:1,000; cat. no. 4329; Cell Signaling Technology, Inc.), rabbit
anti-post synaptic density protein 95 (PSD95; dilution, 1:1,000;
cat. no. 2507; Cell Signaling Technology, Inc.), anti-zonula
occludens-1 (ZO-1; dilution, 1:1,000; cat. no. ab190085; Abcam),
rabbit anti-Bcl-2 (dilution, 1:1,000; cat. no. 15071; Cell
Signaling Technology, Inc.), rabbit anti-Bax (dilution, 1:1,000;
cat. no. 2774; Cell Signaling Technology, Inc.), rabbit
anti-cleaved caspase-3 (dilution, 1:1,000; cat. no. 9664; Cell
Signaling Technology, Inc.), rabbit anti-cytochrome c
(Cyt-c; dilution, 1:1,000; cat. no. ab133504; Abcam) and anti-GAPDH
(dilution, 1:5,000; cat. no. ab8245; Abcam). The cleaved caspase-3
(Asp175) (5A1E) rabbit monoclonal antibody can detect endogenous
levels of the large fragment (17/19 kDa) of activated caspase-3,
resulting from cleavage adjacent to Asp175, and the antibody does
not recognize full length caspase-3 or other cleaved caspases.
After rinsing with TBST (with 20% Tween-20), the membrane was
incubated with horseradish peroxidase (HRP)-conjugated anti-rabbit
IgG (dilution, 1:2,000; cat. no. 7074; Cell Signaling Technology,
Inc.) and HRP-conjugated anti-mouse IgG (dilution, 1:2,000; cat.
no. 7076; Cell Signaling Technology, Inc.) for 1 h at room
temperature. After washing the slices with TBST, the protein levels
were determined with HRP-ECL method (BeyoECL Star Kit, cat. no.
P0018AS; Beyotime Institute of Biotechnology), and detected by the
ChemiDoc™ XRS + Imaging System (Bio-Rad Laboratories, Inc.). The
immunoblot density was analyzed using ImageJ software V1.49
(National Institutes of Health).
Statistical analysis
Each test was repeated at least three times, and all
experimental data are presented as the mean ± standard deviation.
Using SPSS version 20.0 (IBM Corp.), an unpaired Student's t-test
was performed to determine the statistical significance of the
differences between pairs of groups, while one-way ANOVA with post
hoc Bonferroni's multiple comparison tests was used to compare
multiple group means. P<0.05 was considered to indicate a
statistically significant difference.
Results
NBP combined with edaravone attenuates
infarct volumes and neurological deficit scores
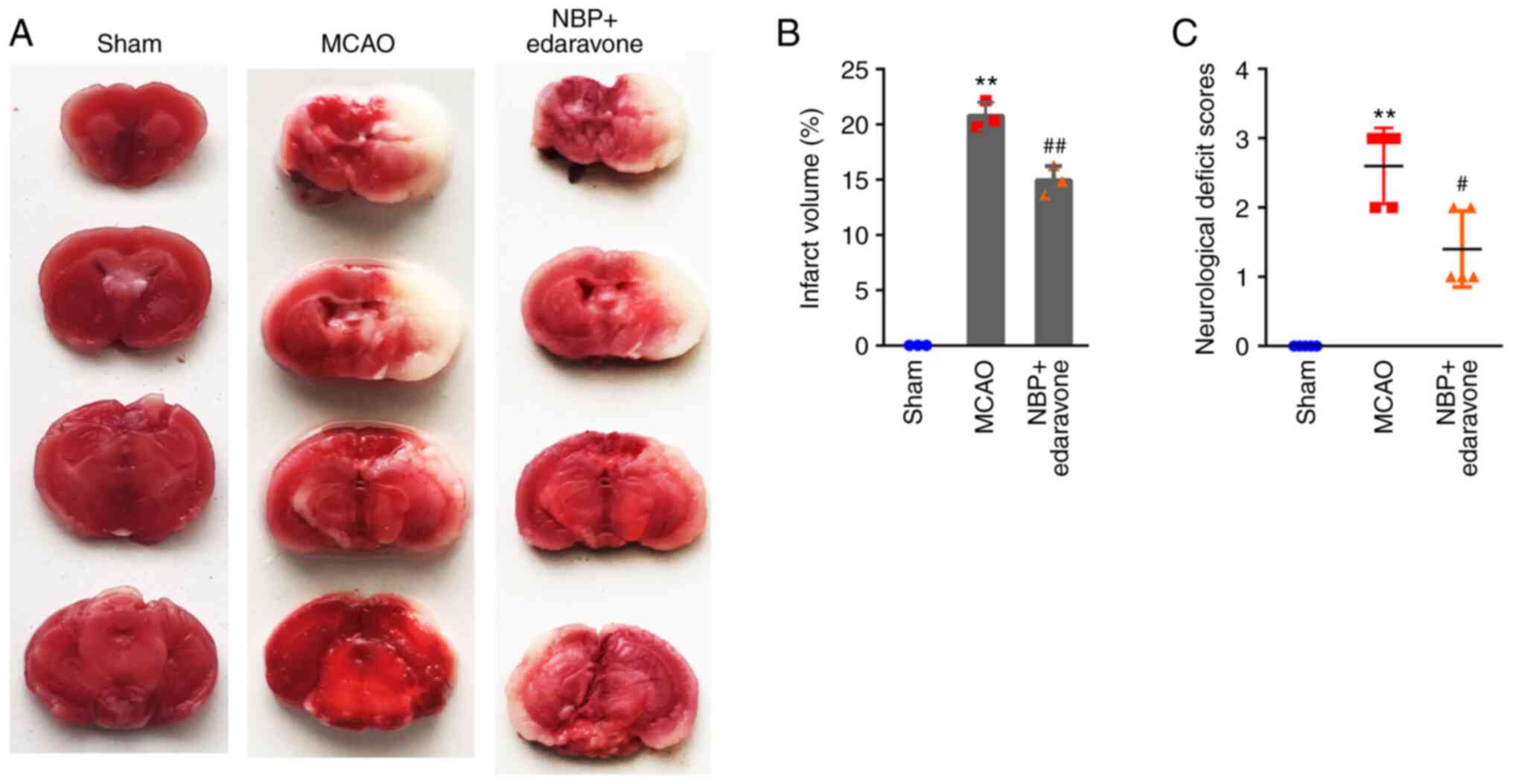
Infarct volumes were determined using TTC staining.
The control mice in the sham group had no obvious infarcts, while
the average volumes of the NBP + edaravone and MCAO groups were
14.92±1.33 and 20.75±1.23%, respectively (Fig. 1A and B). The NBP + edaravone mice
exhibited significantly lower infarct volumes compared with the
MCAO group. Consistent with the TTC staining results, NBP combined
with edaravone significantly attenuated the neurological deficit
scores compared with the MCAO only group (Fig. 1C). The data revealed that
treatment of NBP combined with edaravone could ameliorate the
reduced neurological scores and the infarction volumes resulting
from cerebral ischemia.
NBP combined with edaravone attenuates
cerebral cell damage
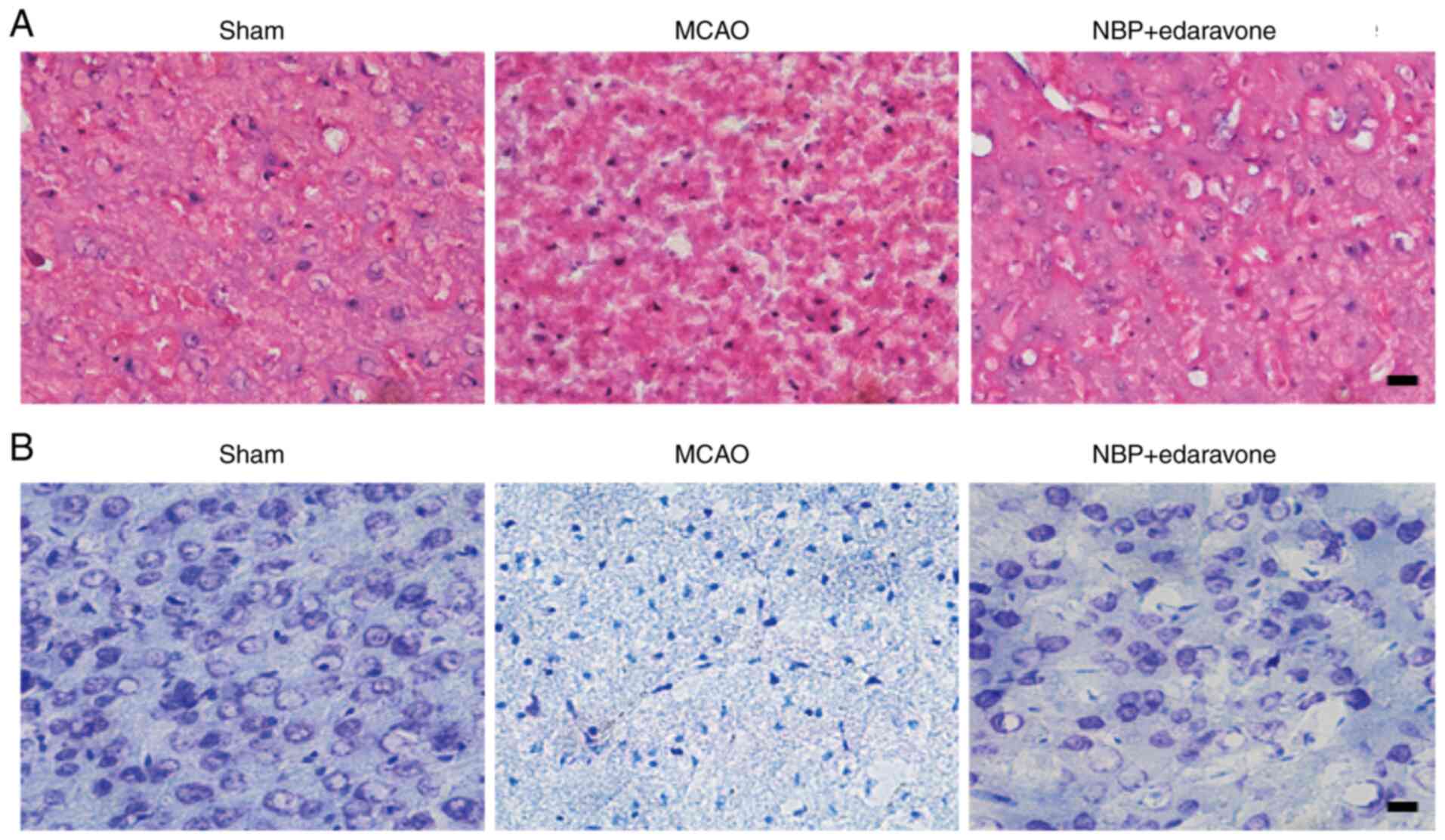
Cerebral cell damage was determined using Nissl and
HE staining. As presented in HE stained tissue sections, the
structures of cerebral cortical neurons were clear and complete,
the cells were arranged densely and orderly without edema, and the
cytoplasm of neurons was lightly stained in sham group (Fig. 2A, sham). In the MCAO group, the
structures of cortical neurons were disordered, including cell
swelling, nuclear pyknosis, fragmentation and dissolution (Fig. 2A, MCAO). The neuronal damage in
NBP combined with edaravone group was less than that in the MCAO
group, with the images displaying that most of the cell structures
were complete, but there were scattered necrotic and slight edema
between cells (Fig. 2A, NBP +
edaravone). For Nissl stained tissue sections, there were no marked
morphological changes in the sham group, the Nissl bodies were
uniformly stained; neurons were arranged orderly without necrosis
and edema (Fig. 2B, sham). By
contrast, significant morphological changes were detected in the
peri-infarct zone of the MCAO model mice, including neuronal loss,
nuclei shrinkage and dark staining of neurons (Fig. 2B, MCAO), while the neuronal loss,
nuclear shrinkage and morphological changes were significantly
reduced in the NBP combined with edaravone treated mice compared
with the MCAO group (Fig. 2B, NBP
+ edaravone). These results indicated that NBP combined with
edaravone could ameliorate neuronal damage caused by cerebral
ischemia-reperfusion injury.
Effects of NBP combined with edaravone
on the NVU cell immunoreactivity
Astrocytes, neurons and vascular endothelial cells
are the main components of the NVU (15). Therefore, the effects of NBP
combined with edaravone on the NVU were evaluated using the cell
immunoreactivity of astrocytes, neurons and vascular endothelial
cells, visualized using immunofluorescence staining.
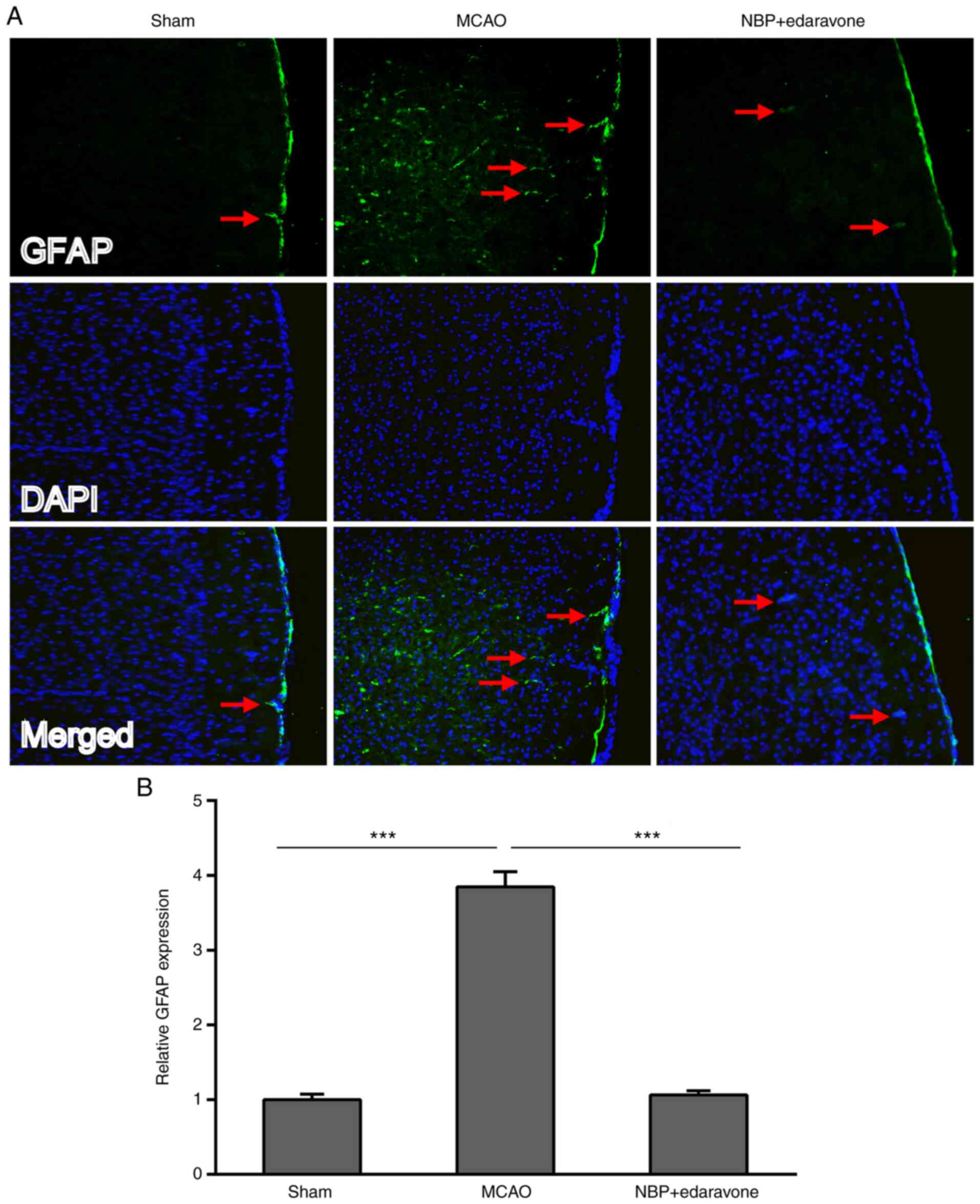
Astrocyte immunoreactivity was assessed using GFAP
immunofluorescence staining. GFAP marks the cytoskeleton of
astrocytes (22), and thus
indicates the astrocyte-positive and soma structure in the single
images. The GFAP expression of the MCAO group was significantly
increased compared with that of the sham group (Fig. 3). However, NBP combined with
edaravone significantly weakened the GFAP expression compared with
the MCAO group.
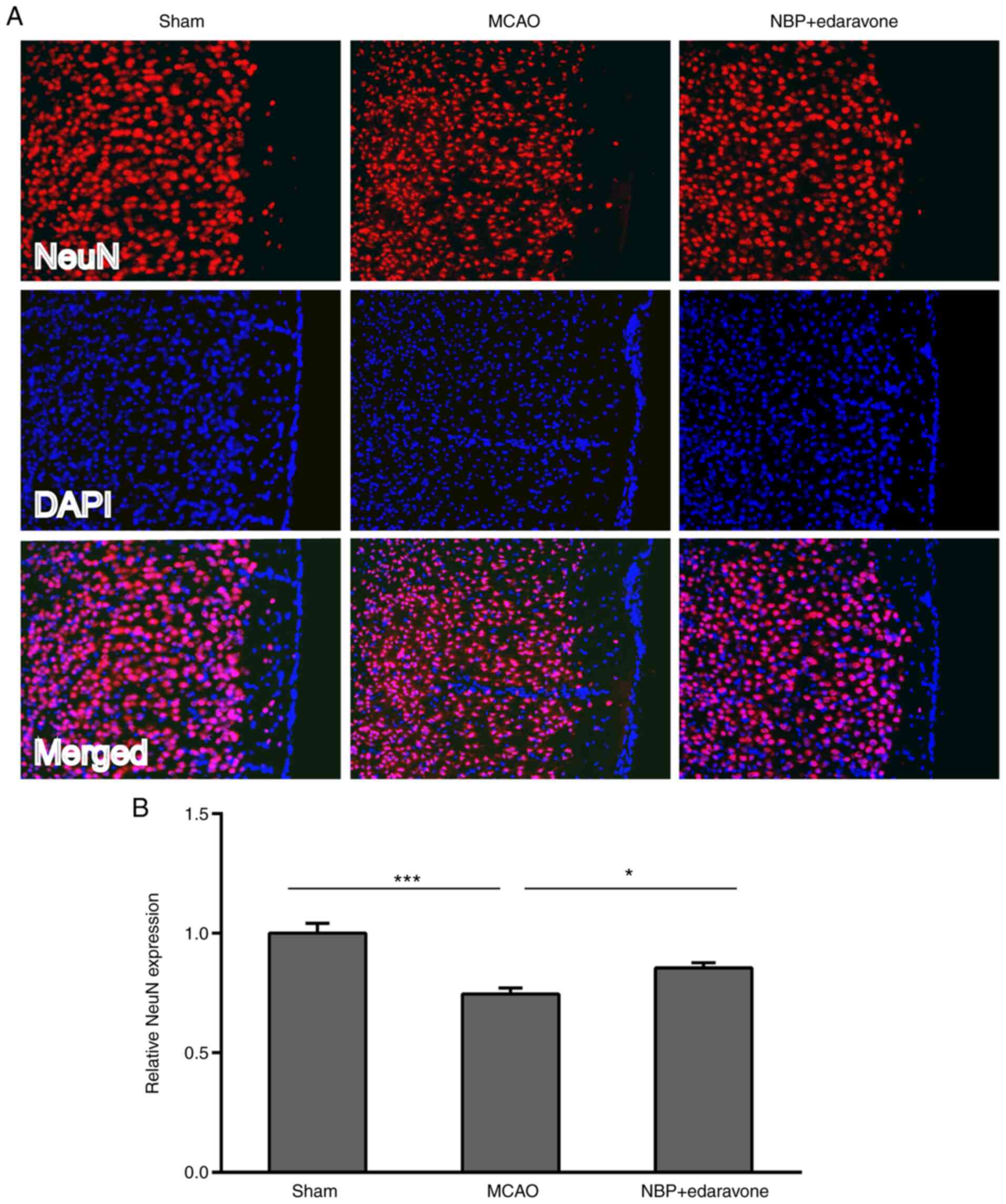
Neuron immunoreactivity was assessed by NeuN
immunofluorescence staining. As presented in Fig. 4, the MCAO group exhibited
significantly decreased NeuN expression compared with the sham
operation group. By contrast, NBP combined with edaravone
significantly increased the NeuN expression compared with the MCAO
group.
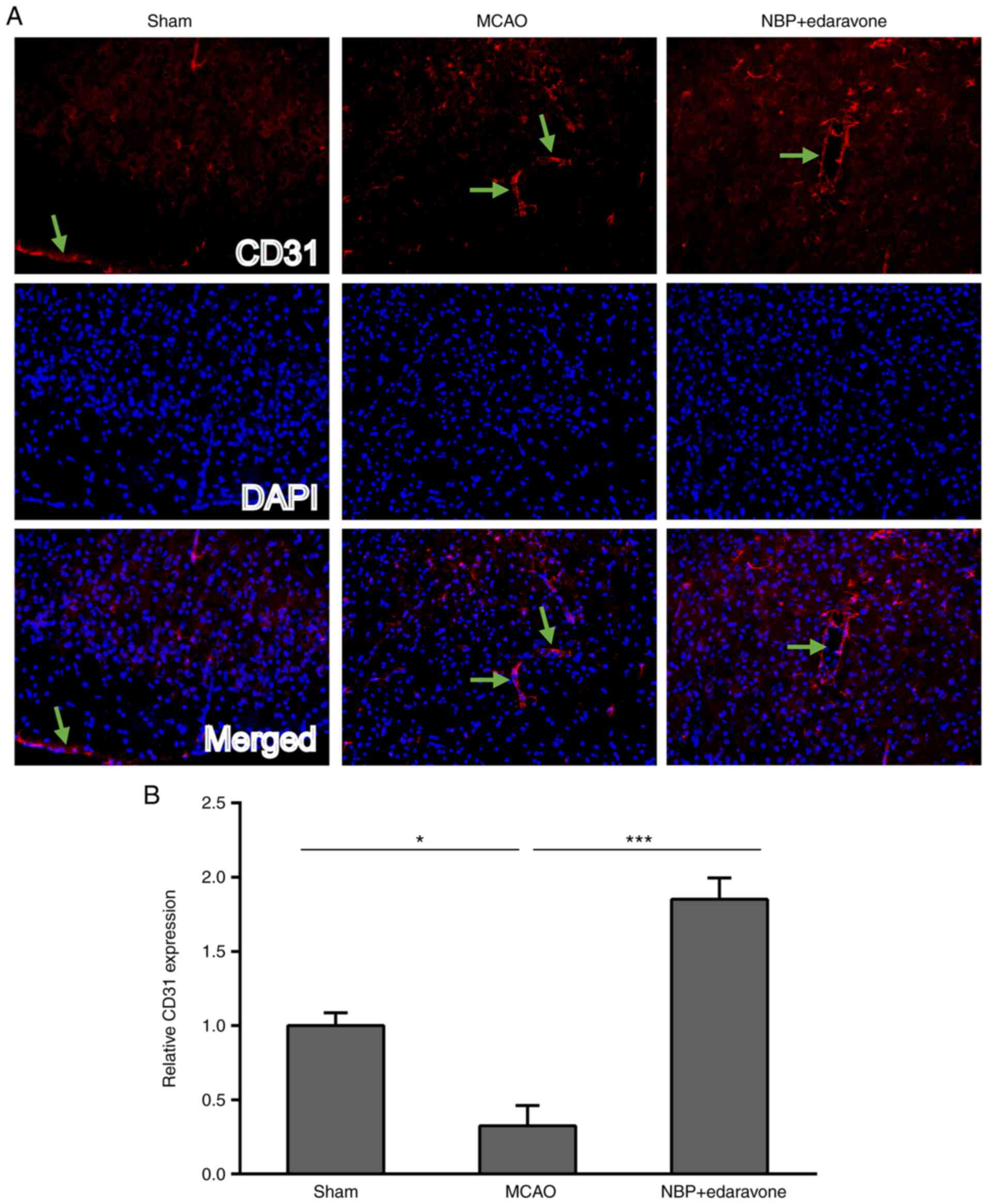
CD31 is a marker of vascular endothelial cells.
Vascular endothelial cell immunoreactivity was determined using
CD31 immunofluorescence staining. CD31 protein expression indicated
that the two markers are closely associated with the basic
structure of cells in the merged image (23). CD31 expression was significantly
decreased in the MCAO group compared with the sham operation group
(Fig. 5), whereas NBP combined
with edaravone significantly increased the CD31 expression compared
with the MCAO group.
These results indicated that GFAP, NeuN and CD31
were expressed in neurons in mice, and also suggested that cerebral
ischemia can increase the expression of GFAP, but decrease the
expression of both NeuN and CD31, while treatment of NBP combined
with edaravone could decrease the expression of GFAP and increase
the expression of both NeuN and CD31 in MCAO model mice.
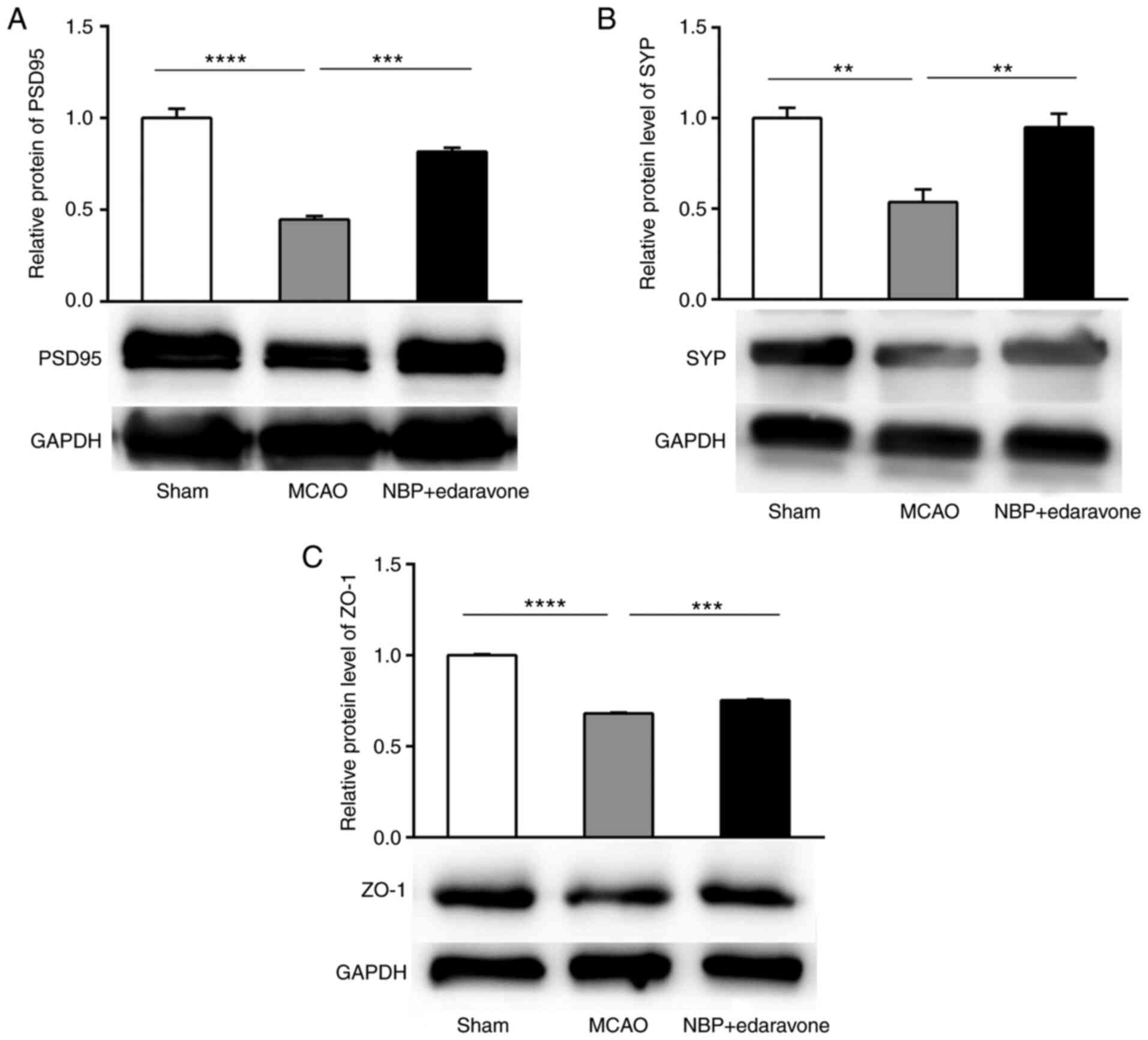
Effects of NBP combined with edaravone
on the blood-brain barrier (BBB)
To explore the effects of NBP combined with
edaravone on the BBB, western blotting was performed to measure the
expression levels of PSD95, the synaptic protein SYP and the tight
junction protein ZO-1. The expression levels of PSD95 and SYP were
significantly reduced in the MCAO model mice, while NBP + edaravone
significantly increased their expression levels compared with the
MCAO group (Fig. 6A and B). ZO-1
expression was significantly decreased in the MCAO group compared
with the sham operation group, while NBP + edaravone significantly
increased ZO-1 protein expression compared with the MCAO group
(Fig. 6C). These results
suggested that cerebral ischemia could decrease the protein
expression levels of PSD95, SYP and ZO-1 in the MCAO groups at 24
h, but administration of NBP + edaravone significantly increased
the expression levels of these three proteins compared with the
MCAO group at the same time.
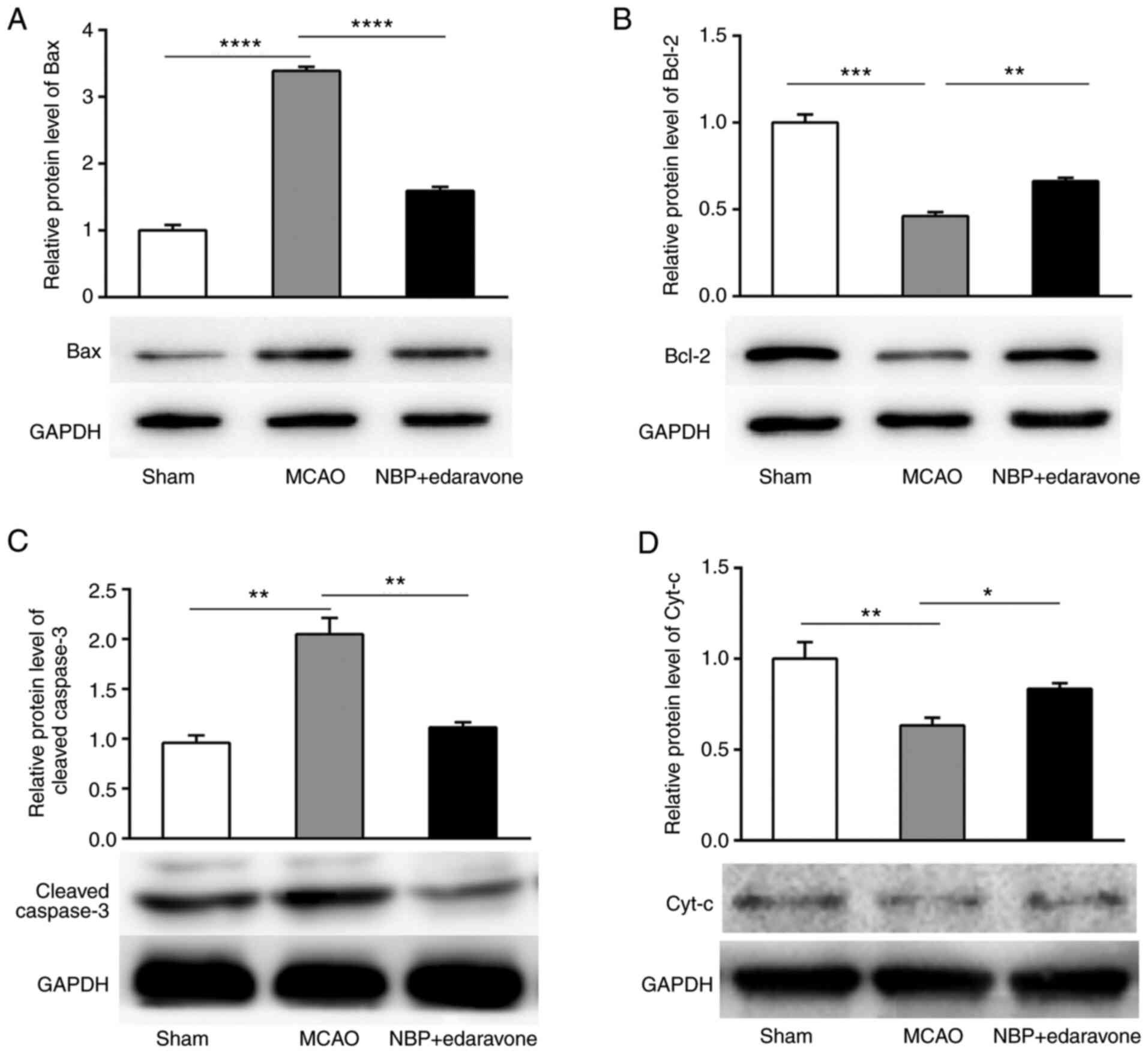
Effects of NBP combined with edaravone
on the expression levels of apoptosis-related proteins
The expression levels of apoptosis-related proteins
were detected using western blotting. The expression levels of
cleaved caspase-3 and pro-apoptotic protein Bax were significantly
increased, while anti-apoptotic protein Bcl-2 expression was
significantly decreased in the MCAO model mice compared with the
sham group. NBP + edaravone significantly decreased cleaved
caspase-3 and pro-apoptotic protein Bax expression, and
significantly increased anti-apoptotic protein Bcl-2 expression
compared with the MCAO mice (Fig.
7A-C). In the sham group, the expression levels of Cyt-c in the
mitochondria were significantly increased compared with those in
the MCAO mice. In the NBP + edaravone group, Cyt-c expression was
significantly increased compared with that in the MCAO group
(Fig. 7D). These data indicated
that NBP + edaravone treatment may play a key role in preventing
apoptosis during cerebral ischemia-reperfusion injury.
Discussion
The neurons, BBB, microglial cells and extracellular
matrix that maintain the integrity of brain tissue are the
structural basis of the NVU (15,16). The experimental results indicated
that NBP + edaravone protected the brain against ischemic stroke
damage by targeting the whole NVU in the mouse MCAO model. NBP +
edaravone alleviated brain injury in MCAO mice, as demonstrated by
the reduced infarct volumes and neurological deficit scores. These
neuroprotective effects might be due to these drugs attenuating the
cerebral cell damage, positively affecting the expression of some
associated neuroproteins, such as PSD95, SYP, ZO-1, claudin-5 and
CD31, for cell activity in the NVU, improving the BBB function and
inhibiting apoptosis by altering the expression levels of
apoptosis-related proteins.
NBP reportedly attenuates neuronal injuries
following both in vitro and in vivo ischemic strokes
(24,25). NeuN immunosignals are decreased in
the neocortex and striatum of mice following 24-h ischemia
(26). Additionally, the NBP
derivative, (S)-ZIM-289, reportedly protects neurons in the brain
from ischemic damage by decreasing the brain infarct area,
improving the neurological function and preventing neuronal loss
and apoptosis in a rat MCAO model (27). In the present study, NBP +
edaravone increased NeuN expression compared with that in MCAO
model mice, demonstrating their neuronal protective effect against
ischemic brain injury.
GFAP is a major mature astrocyte filament and is
considered a promising serum biomarker to differentiate between
intracerebral hemorrhage and acute ischemic stroke (28). GFAP expression is low in the
healthy mouse cortex (29,30).
However, GFAP expression levels are high in various neurological
dysfunction diseases, including ischemic stroke (31). High GFAP expression is considered
a marker of reactive astrogliosis, a process in which astrocytes
respond to various neurological dysfunction diseases (31–33). High serum GFAP levels are
associated with poor outcomes in patients with acute ischemic
stroke (34). Several studies
have demonstrated that the expression levels of GFAP may increase
at different time intervals after unilateral MCAO (35,36); however, some neuroprotective
agents can reduce the expression levels of GFAP (37,38). Similarly, in the present study,
the MCAO mice exhibited significantly increased GFAP expression
compared with the sham mice. NBP + edaravone significantly
decreased GFAP expression, indicating their suppressive effects on
superabundant gliogenesis. These results suggested that the
combination of butylphthalide and edaravone would not cause damage
to the NVU, and the combination of the two drugs should have
protective effects on the NVU due to the decrease of GFAP
expression at 24 h after reperfusion in mice.
NBP has been reported to promote angiogenesis and
improve the BBB function following ischemic injury in a rat MCAO
model (39–41). NBP markedly increases the levels
of circulating endothelial progenitor cells in patients with acute
ischemic stroke, improving the clinical prognosis (42). CD31 is associated with
angiogenesis (43,44). Physical exercise reduces the brain
infarct area, improves neurological function and promotes
angiogenesis by increasing CD31 expression and protein expression
in a rat MCAO model (45). In the
present study, CD31 expression was decreased in mice following
ischemic stroke injury, while NBP + edaravone significantly
increased CD31 expression compared with the MCAO model mice. The
positive effects of NBP + edaravone in individuals with ischemic
stroke may be partly due to the promotion of angiogenesis.
Zhang et al (46) reported an improvement in
neurological function and survival rate in a mouse MCAO model with
non-erythropoietic mutant erythropoietin injected
intraperitoneally, which may be due to erythropoietin promoting
angiogenesis and neurogenesis, while suppressing superabundant
gliogenesis. Based on the GFAP, NeuN and CD31 immunofluorescence
staining findings of the present study, it was hypothesized that
NBP + edaravone might have a similar mechanism by inhibiting
superabundant gliogenesis and promoting neurogenesis and
angiogenesis.
PSD95, SYN and other proteins, such as
microtubule-associated protein 2, which are closely associated with
synaptic formation and neurotransmission, can be considered as
markers of presynaptic and postsynaptic components. PSD95 and SYP
are also markers of synaptic plasticity (47). The decrease of PSD95 and SYP
expression indicates a loss of synapses (47,48). Tight junctions are a hallmark of
polarized epithelial cells, and ZO-1 is a known key regulator of
tight junction formation. ZO-1 is the first confirmed tight
junction cytoplasmic protein, and it not only provides a scaffold
for connecting adhesion and transmembrane proteins, but is also
associated with the increase of BBB permeability when its loss and
degradation occur (49).
Therefore, ZO-1 serves a notable role in maintaining the continuity
and integrity of tight junctions.
An ischemic stroke will first damage the BBB,
altering its structure and function and increasing permeability. In
a rat ischemic stroke model, the BBB integrity was disrupted
following ischemia (50). The
deterioration is accompanied by reduced expression levels of the
synaptic proteins PSD95 and SYP and the tight junction protein ZO-1
(34). The acute inflammatory
disorder drug, ulinastatin, protects the brain from ischemic damage
by increasing ZO-1 expression (51). Alogliptin attenuates brain
disruption and restores ZO-1 expression in a mouse MCAO model
(52). Consistent with the
aforementioned reports, the present study revealed that the MCAO
mice had significantly increased brain infarct volumes and
neurological deficit scores, and significantly reduced expression
levels of PSD95, SYP and ZO-1 compared with the sham mice.
The protective effects of NBP or edaravone on
ischemia-induced apoptosis have been reported previously (7,8,53).
NBP (40 mg/kg; i.p. immediately after ischemia/reperfusion)
markedly increases the ratio of Bcl-2/Bax in the hippocampus of
Mongolian gerbils after global cerebral ischemia and reperfusion
damage (54). Data has
illustrated that a single NBP or edaravone treatment may inhibit
the mitochondria-dependent apoptotic cascade after ischemia and
reperfusion (7,55); however, to the best of our
knowledge, few animal experiments have examined the
apoptosis-related effects of both drugs combined. Therefore, it was
important to investigate the effects of NBP + edaravone on the
release of Cyt-c, the ratio of Bcl-2/Bax and the activation of
cleaved caspase-3 after transient focal cerebral ischemia.
A previous study has investigated the effects of NBP
on apoptosis induced by transient focal cerebral ischemia in rats
and compared the expression levels of Cyt-c in cytosolic and
mitochondrial fractions (24).
The release of Cyt-c was maximal at 24 h after reperfusion, and NBP
markedly inhibited the distributional change of Cyt-c. The levels
of Cyt-c in cytosolic and mitochondrial fractions were different at
24 h after reperfusion. There was minimal Cyt-c expression in the
cytoplasm of the sham group. By contrast, Cyt-c expression was high
in the mitochondria of the same group, and low in both the
cytoplasm and mitochondria of the vehicle group. The expression
levels of Cyt-c in the cytoplasm and mitochondria of the NBP group
were markedly higher compared with those in the vehicle group
(24). The results of the present
study regarding Cyt-c in mitochondrial protein were consistent with
these previous findings. These observations can preliminarily
explain how the combination of these two drugs may serve a
protective role via Cyt-c regulation. However, the present study
only detected Cyt-c levels in the mitochondrial proteins without
synchronous data of the cytoplasmic fraction. Therefore, the
differences between the two fractions could not be compared, and
this absence is a limitation of the present study.
Numerous studies have demonstrated that NBP can
improve post-stroke symptoms through several mechanisms and
multi-target effects (7,8), including reducing the inflammatory
response (56), improving
collateral circulation (40–42), protecting mitochondrial function
(24,27), inhibiting apoptosis (24,54) and reducing oxidative stress
(25). The aim of the present
study was to explore whether the co-administration of NBP and
edaravone had protective multi-target effects on the NVU of mice
with ischemic strokes, and focused on the changes of vascular
endothelium in MCAO mice. Although these associated factors were
investigated, there are still some deficiencies, and some
associated fields require further research.
Numerous studies investigating the effect of
butylphthalide on inflammation have been carried out, and the
anti-inflammatory effects of butylphthalide have been confirmed
(7,8,56).
However, not investigating inflammatory targets and proteins is a
limitation of the present study, and the anti-inflammatory effect
of butylphthalide combined with edaravone remains unclear.
Therefore, subsequent research should further clarify the
multi-target effects of butylphthalide combined with edaravone
involved in anti-inflammatory and antioxidant effects. However,
there are still some technical limitations in the present study.
Firstly, the lack of low magnification brain slices is a limitation
of the pathological study due to some unexpected interference, and
future studies should provide these. Secondly, the present study
only focused on cleaved caspase-3, and if all types of caspase-3,
such as pro-caspase-3 and cleaved caspase-3, had been detected at
the same time, the evidence would have been more reliable. Thirdly,
the current study would have been improved if the mice groups had
been arranged for administration in different dosage (low, middle
and high dosage) groups for the combined drugs. It is expected that
future studies will be more precise and perfect in experimental
design, methods and technologies, in order to make the results more
credible and reliable.
Numerous studies have focused on different
multi-targets and have revealed some mechanisms associated with the
neuroprotective effects of butylphthalide or edaravone; however,
few studies specifically involving MCAO models have been carried
out to investigate the combined administration of the two
drugs.
In conclusion, the present study preliminarily
suggested that NBP + edaravone could exert neuroprotective effects
in a mouse model by targeting multiple NVU components. These
neuroprotective effects mainly included reducing cell damage and
apoptosis, increasing the number of neurons and vascular
endothelial cells and decreasing astrocyte gliogenesis, thus
reducing the brain infarct volume and neurological deficits.
Therefore, NBP + edaravone might be a promising future therapy for
patients with ischemic stroke.
Acknowledgements
Not applicable.
Funding
This work was supported by the Hebei Key Research
and Development Program: Health Care and Biomedical Special Project
(grant no. 18277787D).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YC and YG conceived and designed the research. YG
and PL conducted all experiments. YL, LG and QW helped conduct
experiments. YC and YG confirmed the authenticity of all the raw
data. YG and YC wrote the manuscript. All authors revised the
manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Ethical permission was obtained from the Animal
Ethics Committee of North China University of the Science and
Technology (approval no. 2019068; Tangshan, China), which records
and regulates all research activities. The approval from the Animal
Ethics Committee included the permission of using mice under
euthanasia, and all experimental procedures were conducted in
strict accordance with recommendations in the Guide for the Care
and Use of Laboratory Animals of the National Institutes of
Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BBB
|
blood-brain barrier
|
|
CD31
|
platelet and endothelial cell adhesion
molecule 1
|
|
HE
|
hematoxylin-eosin
|
|
HRP
|
horseradish peroxidase
|
|
MCAO
|
middle cerebral artery occlusion
|
|
NBP
|
DL-3-n-butylphthalide
|
|
NeuN
|
neuronal nuclei
|
|
NVU
|
neurovascular unit
|
|
PSD95
|
post synaptic density protein 95
|
|
SYP
|
synaptophysin
|
|
TTC
|
2,3,5-triphenyltetrazolium
chloride
|
References
|
1
|
Benjamin EJ, Muntner P, Alonso A,
Bittencourt MS, Callaway CW, Carson AP, Chamberlain AM, Chang AR,
Cheng S, Das SR, et al: Heart disease and stroke statistics-2019
update: A report from the American heart association. Circulation.
139:e56–e528. 2019. View Article : Google Scholar
|
|
2
|
GBD 2017 Disease, Injury Incidence and
Prevalence Collaborators: Global, regional, and national incidence,
prevalence, and years lived with disability for 354 diseases and
injuries for 195 countries and territories, 1990–2017: A systematic
analysis for the global burden of disease study 2017. Lancet.
392:1789–1858. 2018. View Article : Google Scholar
|
|
3
|
Chen X and Wang K: The fate of medications
evaluated for ischemic stroke pharmacotherapy over the period
1995–2015. Acta Pharm Sin B. 6:522–530. 2016. View Article : Google Scholar
|
|
4
|
Zhang JJ and Liu X: Aspirin plus
dipyridamole has the highest surface under the cumulative ranking
curves (SUCRA) values in terms of mortality, intracranial
hemorrhage, and adverse event rate among 7 drug therapies in the
treatment of cerebral infarction. Medicine (Baltimore).
97:e01232018. View Article : Google Scholar
|
|
5
|
Yusuf S, Joseph P, Dans A, Gao P, Teo K,
Xavier D, López-Jaramillo P, Yusoff K, Santoso A, Gamra H, et al:
Polypill with or without aspirin in persons without cardiovascular
disease. N Engl J Med. 384:216–228. 2021. View Article : Google Scholar
|
|
6
|
Ibraheem M and Goldstein LB: Polypill
trials for stroke prevention-main results, critical appraisal, and
implications for US population. Curr Neurol Neurosci Rep.
20:102020. View Article : Google Scholar
|
|
7
|
Abdoulaye IA and Guo YJ: A Review of
recent advances in neuroprotective potential of 3-N-butylphthalide
and its derivatives. Biomed Res Int. 2016:50123412016. View Article : Google Scholar
|
|
8
|
Wang S, Ma F, Huang L, Zhang Y and Peng Y,
Xing C, Feng Y, Wang X and Peng Y: Dl-3-n-butylphthalide (NBP): A
promising therapeutic agent for ischemic stroke. CNS Neurol Disord
Drug Targets. 17:338–347. 2018. View Article : Google Scholar
|
|
9
|
Edaravone Acute Infarction Study Group, :
Effect of a novel free radical scavenger, edaravone (MCI-186), on
acute brain infarction. Randomized, placebo-controlled,
double-blind study at multicenters. Cerebrovasc Dis. 15:222–229.
2003. View Article : Google Scholar
|
|
10
|
Enomoto M, Endo A, Yatsushige H, Fushimi K
and Otomo Y: Clinical effects of early edaravone use in acute
ischemic stroke patients treated by endovascular reperfusion
therapy. Stroke. 50:652–658. 2019. View Article : Google Scholar
|
|
11
|
Kern R, Nagayama M, Toyoda K, Steiner T,
Hennerici MG and Shinohara Y: Comparison of the European and
Japanese guidelines for the management of ischemic stroke.
Cerebrovasc Dis. 35:402–418. 2013. View Article : Google Scholar
|
|
12
|
Chen C, Li M, Lin L, Chen S, Chen Y and
Hong L: Clinical effects and safety of edaravone in treatment of
acute ischaemic stroke: A meta-analysis of randomized controlled
trials. J Clin Pharm Ther. 46:907–917. 2021. View Article : Google Scholar
|
|
13
|
Hua K, Sheng X, Li TT, Wang LN, Zhang YH,
Huang ZJ and Ji H: The edaravone and 3-n-butylphthalide
ring-opening derivative 10b effectively attenuates cerebral
ischemia injury in rats. Acta Pharmacol Sin. 36:917–927. 2015.
View Article : Google Scholar
|
|
14
|
Sheng X, Hua K, Yang C, Wang X, Ji H, Xu
J, Huang Z and Zhang Y: Novel hybrids of 3-n-butylphthalide and
edaravone: Design, synthesis and evaluations as potential
anti-ischemic stroke agents. Bioorg Med Chem Lett. 25:3535–3540.
2015. View Article : Google Scholar
|
|
15
|
Zhao Y, Yang J, Li C, Zhou G, Wan H, Ding
Z, Wan H and Zhou H: Role of the neurovascular unit in the process
of cerebral ischemic injury. Pharmacol Res. 160:1051032020.
View Article : Google Scholar
|
|
16
|
Wang L, Xiong X, Zhang L and Shen J:
Neurovascular unit: A critical role in ischemic stroke. CNS
Neurosci Ther. 27:7–16. 2021. View Article : Google Scholar
|
|
17
|
Boltze J, Aronowski JA, Badaut J,
Buckwalter MS, Caleo M, Chopp M, Dave KR, Didwischus N, Dijkhuizen
RM, Doeppner TR, et al: New mechanistic insights, novel treatment
paradigms, and clinical progress in cerebrovascular diseases. Front
Aging Neurosci. 13:6237512021. View Article : Google Scholar
|
|
18
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press; Washington, DC: 2011
|
|
19
|
Feng L, Sharma A, Niu F, Huang Y, Lafuente
JV, Muresanu DF, Ozkizilcik A, Tian ZR and Sharma HS:
TiO2-nanowired delivery of DL-3-n-butylphthalide
(DL-NBP) attenuates blood-brain barrier disruption, brain edema
formation, and neuronal damages following concussive head injury.
Mol Neurobiol. 55:350–358. 2018. View Article : Google Scholar
|
|
20
|
Alzoubi KH, Shatnawi A, Al-Qudah MA and
Alfaqih MA: Edaravone prevents memory impairment in an animal model
of post-traumatic distress. Behav Pharmacol. 30:201–207. 2019.
View Article : Google Scholar
|
|
21
|
Bederson JB, Pitts LH, Tsuji M, Nishimura
MC, Davis RL and Bartkowski H: Rat middle cerebral artery
occlusion: Evaluation of the model and development of a neurologic
examination. Stroke. 17:472–476. 1986. View Article : Google Scholar
|
|
22
|
Middeldorp J and Hol EM: GFAP in health
and disease. Prog Neurobiol. 93:421–443. 2011. View Article : Google Scholar
|
|
23
|
Lertkiatmongkol P, Liao D, Mei H, Hu Y and
Newman PJ: Endothelial functions of platelet/endothelial cell
adhesion molecule-1 (CD31). Curr Opin Hematol. 23:253–259. 2016.
View Article : Google Scholar
|
|
24
|
Chang Q and Wang XL: Effects of chiral
3-n-butylphthalide on apoptosis induced by transient focal cerebral
ischemia in rats. Acta Pharmacol Sin. 24:796–804. 2003.
|
|
25
|
Li J, Li Y, Ogle M, Zhou X, Song M, Yu SP
and Wei L: DL-3-n-butylphthalide prevents neuronal cell death after
focal cerebral ischemia in mice via the JNK pathway. Brain Res.
1359:216–226. 2010. View Article : Google Scholar
|
|
26
|
Zhao Y, Liu D, Li J, Zhang X and Wang X:
L-NBP, a multiple growth factor activator, attenuates ischemic
neuronal impairments possibly through promoting neuritogenesis.
Neurochem Int. 124:94–105. 2019. View Article : Google Scholar
|
|
27
|
Zhao Q, Zhang C, Wang X, Chen L, Ji H and
Zhang Y: (S)-ZJM-289, a nitric oxide-releasing derivative of
3-n-butylphthalide, protects against ischemic neuronal injury by
attenuating mitochondrial dysfunction and associated cell death.
Neurochem Int. 60:134–144. 2012. View Article : Google Scholar
|
|
28
|
Cabezas JA, Bustamante A, Giannini N,
Pecharroman E, Katsanos AH, Tsivgoulis G, Rozanski M, Audebert H,
Mondello S, Llombart V and Montaner J: Discriminative value of
glial fibrillar acidic protein (GFAP) as a diagnostic tool in acute
stroke. Individual patient data meta-analysis. J Investig Med.
68:1379–1385. 2020. View Article : Google Scholar
|
|
29
|
Li H, Zhang N, Sun G and Ding S:
Inhibition of the group I mGluRs reduces acute brain damage and
improves long-term histological outcomes after
photothrombosis-induced ischaemia. ASN Neuro. 5:195–207. 2013.
View Article : Google Scholar
|
|
30
|
Li H, Zhang N, Lin HY, Yu Y, Cai QY, Ma L
and Ding S: Histological, cellular and behavioral assessments of
stroke outcomes after photothrombosis-induced ischemia in adult
mice. BMC Neurosci. 15:582014. View Article : Google Scholar
|
|
31
|
Choudhury GR and Ding S: Reactive
astrocytes and therapeutic potential in focal ischemic stroke.
Neurobiol Dis. 85:234–244. 2016. View Article : Google Scholar
|
|
32
|
Mestriner RG, Saur L, Bagatini PB,
Baptista PP, Vaz SP, Ferreira K, Machado SA, Xavier LL and Netto
CA: Astrocyte morphology after ischemic and hemorrhagic
experimental stroke has no influence on the different recovery
patterns. Behav Brain Res. 278:257–261. 2015. View Article : Google Scholar
|
|
33
|
Liu P, Zhang R, Liu D, Wang J, Yuan C,
Zhao X, Li Y, Ji X, Chi T and Zou L: Time-course investigation of
blood-brain barrier permeability and tight junction protein changes
in a rat model of permanent focal ischemia. J Physiol Sci.
68:121–127. 2018. View Article : Google Scholar
|
|
34
|
Liu G and Geng J: Glial fibrillary acidic
protein as a prognostic marker of acute ischemic stroke. Hum Exp
Toxicol. 37:1048–1053. 2018. View Article : Google Scholar
|
|
35
|
Fahrig T: Changes in the solubility of
glial fibrillary acidic protein after ischemic brain damage in the
mouse. J Neurochem. 63:1796–1801. 1994. View Article : Google Scholar
|
|
36
|
Cheung WM, Wang CK, Kuo JS and Lin TN:
Changes in the level of glial fibrillary acidic protein (GFAP)
after mild and severe focal cerebral ischemia. Chin J Physiol.
42:227–235. 1999.
|
|
37
|
He F, Dai R, Zhou X, Li X, Song X, Yan H,
Meng Q, Yang C and Lin Q: Protective effect of 4-methoxy benzyl
alcohol on the neurovascular unit after cerebral ischemia
reperfusion injury. Biomed Pharmacother. 118:1092602019. View Article : Google Scholar
|
|
38
|
Pang XB, Xie XM, Wang HY and Wang BQ:
Protective effect of mailuoning injection on cerebral
ischemia/reperfusion injury in rats and its mechanism. Zhongguo
Zhong Yao Za Zhi. 39:721–725. 2014.(In Chinese).
|
|
39
|
Zhou PT, Wang LP, Qu MJ, Shen H, Zheng HR,
Deng LD, Ma YY, Wang YY, Wang YT, Tang YH, et al:
Dl-3-N-butylphthalide promotes angiogenesis and upregulates sonic
hedgehog expression after cerebral ischemia in rats. CNS Neurosci
Ther. 25:748–758. 2019. View Article : Google Scholar
|
|
40
|
Ye ZY, Xing HY, Wang B, Liu M and Lv PY:
DL-3-n-butylphthalide protects the blood-brain barrier against
ischemia/hypoxia injury via upregulation of tight junction
proteins. Chin Med J (Engl). 132:1344–1353. 2019. View Article : Google Scholar
|
|
41
|
Chong ZZ and Feng YP:
dl-3-n-butylphthalide attenuates reperfusion-induced blood-brain
barrier damage after focal cerebral ischemia in rats. Zhongguo Yao
Li Xue Bao. 20:696–700. 1999.
|
|
42
|
Zhao H, Yun W, Zhang Q, Cai X, Li X, Hui
G, Zhou X and Ni J: Mobilization of circulating endothelial
progenitor cells by dl-3-n-butylphthalide in acute ischemic stroke
patients. J Stroke Cerebrovasc Dis. 25:752–760. 2016. View Article : Google Scholar
|
|
43
|
DeLisser HM, Newman PJ and Albelda SM:
Molecular and functional aspects of PECAM-1/CD31. Immunol Today.
15:490–495. 1994. View Article : Google Scholar
|
|
44
|
Abbott NJ, Rönnbäck L and Hansson E:
Astrocyte-endothelial interactions at the blood-brain barrier. Nat
Rev Neurosci. 7:41–53. 2006. View Article : Google Scholar
|
|
45
|
Hu X, Zheng H, Yan T, Pan S, Fang J, Jiang
R and Ma S: Physical exercise induces expression of CD31 and
facilitates neural function recovery in rats with focal cerebral
infarction. Neurol Res. 32:397–402. 2010. View Article : Google Scholar
|
|
46
|
Zhang SJ, Wang RL, Zhao HP, Tao Z, Li JC,
Ju F, Han ZP, Ma QF, Liu P, Ma SB, et al: MEPO promotes
neurogenesis and angiogenesis but suppresses gliogenesis in mice
with acute ischemic stroke. Eur J Pharmacol. 849:1–10. 2019.
View Article : Google Scholar
|
|
47
|
Lin Y, Dong J, Yan T, He X, Zheng X, Liang
H and Sui M: Involuntary, forced and voluntary exercises are
equally capable of inducing hippocampal plasticity and the recovery
of cognitive function after stroke. Neurol Res. 37:893–901. 2015.
View Article : Google Scholar
|
|
48
|
Sell GL, Barrow SL and McAllister AK:
Chapter 1-molecular composition of developing glutamatergic
synapses. Synapse Development and Maturation. Rubenstein J, Rakic
P, Chen B, Kwan KY, Cline HT and Cardin J: Academic Press; London:
pp. 3–32. 2020, View Article : Google Scholar
|
|
49
|
Fanning AS and Anderson JM: Zonula
occludens-1 and −2 are cytosolic scaffolds that regulate the
assembly of cellular junctions. Ann N Y Acad Sci. 1165:113–120.
2009. View Article : Google Scholar
|
|
50
|
Abdullahi W, Tripathi D and Ronaldson PT:
Blood-brain barrier dysfunction in ischemic stroke: Targeting tight
junctions and transporters for vascular protection. Am J Physiol
Cell Physiol. 315:C343–C356. 2018. View Article : Google Scholar
|
|
51
|
Li XF, Zhang XJ, Zhang C, Wang LN, Li YR,
Zhang Y, He TT, Zhu XY, Cui LL and Gao BL: Ulinastatin protects
brain against cerebral ischemia/reperfusion injury through
inhibiting MMP-9 and alleviating loss of ZO-1 and occludin proteins
in mice. Exp Neurol. 302:68–74. 2018. View Article : Google Scholar
|
|
52
|
Hao FL, Han XF, Wang XL, Zhao ZR, Guo AH,
Lu XJ and Zhao XF: The neurovascular protective effect of
alogliptin in murine MCAO model and brain endothelial cells. Biomed
Pharmacother. 109:181–187. 2019. View Article : Google Scholar
|
|
53
|
Kikuchi K, Uchikado H, Miyagi N, Morimoto
Y, Ito T, Tancharoen S, Miura N, Miyata K, Sakamoto R, Kikuchi C,
et al: Beyond neurological disease: New targets for edaravone
(Review). Int J Mol Med. 28:899–906. 2011.
|
|
54
|
Hai W, Yang Y, Wang YL and Nie YX: The
effect of dl-3n-butylphthalide on the neurons in the hippocampus of
mongolian gerbil and the expression of p-ERK, Bcl-2 and Bax after
global cerebral ischemia and reperfusion damage. Chin J Clinicians
(Electronic Edition). 2015:1157–1162. 2015.(In Chinese).
|
|
55
|
Li C, Mo Z, Lei J, Li H, Fu R, Huang Y,
Luo S and Zhang L: Edaravone attenuates neuronal apoptosis in
hypoxic-ischemic brain damage rat model via suppression of TRAIL
signaling pathway. Int J Biochem Cell Biol. 99:169–177. 2018.
View Article : Google Scholar
|
|
56
|
Zeng Z, Gong X and Hu Z:
L-3-n-butylphthalide attenuates inflammation response and brain
edema in rat intracerebral hemorrhage model. Aging (Albany NY).
12:11768–11780. 2020. View Article : Google Scholar
|