Introduction
Cytochrome P450 (CYP) family 2 subfamily E member 1
(CYP2E1) is a member of the CYP family of heme proteins and is
expressed in the liver, brain and heart in humans and animals
(1–4). The CYP2E1 protein is highly
expressed on the mitochondria, Golgi apparatus, endoplasmic
reticulum and plasma membrane (5,6).
CYP2E1 is a key monooxygenase enzyme involved in the metabolism of
several endogenous substrates, as well as hundreds of xenobiotics,
obtained from diet and environmental contamination (2,4–6).
Compared with other CYP family members, CYP2E1 possesses remarkably
high NADPH oxidase activity. Thus, it can catalyze the reaction of
NADPH and O2 to generate reactive oxygen species (ROS)
(7,8). CYP2E1 is the major enzyme for the
production of cellular and mitochondrial ROS and reactive nitrogen
species (RNS), which induce tissue damage mainly by damaging
cellular and mitochondrial macromolecules, including mitochondrial
DNA (9–11).
The expression level of CYP2E1 is upregulated in a
wide variety of pathological states and diseases, including
obesity, diabetes mellitus, alcoholic liver disease, heart
diseases, inflammation, Parkinson's disease and cancer (12–17). In view of the expression
characteristic of CYP2E1, it has been used as a therapeutic target
in drug discovery and already has been applied in preliminarily
studies (18). Diallyl sulfide
(DAS) is a competitive inhibitor of CYP2E1, which is isolated from
garlic (18,19). It has been reported that DAS
treatment can recover ethanol-induced pathological changes in the
liver by inhibiting CYP2E1-mediated alcohol metabolism and
subsequent oxidative stress (20,21). Furthermore, DAS treatment can
prevent myocardial systolic dysfunction induced by chronic ethanol
exposure (22). Studies of the
effects of DAS suggested that CYP2E1 inhibition may have potential
effects on the development of diseases characterized with high
CYP2E1 expression.
Dilated cardiomyopathy (DCM), characterized by the
dilation of the heart chambers and impaired systolic functions, is
the most frequent cause of heart transplantation and it is the
third most common reason for heart failure (HF) (23). Accumulating evidence has indicated
the important role of oxidative stress in the pathophysiology of
cardiac remodeling and HF (24,25). Furthermore, it has been shown that
CYP2E1 is upregulated in multiple cardiovascular diseases,
including DCM, hypertrophic cardiomyopathy (HCM) and ischemia
(4,5,26–32). Our previous study revealed that
the phenotype of CYP2E1 overexpressing-transgenic mice was similar
to that of DCM, and knockdown of endogenous CYP2E1 expression
markedly prevented the development of DCM in mice (4,27).
In addition, since CYP2E1 is a major source of cellular and
mitochondrial ROS/RNS, inhibition of CYP2E1 may improve the
pathological development of DCM. The aforementioned studies suggest
that inhibition of CYP2E1 may improve the pathological development
of DCM and application of CYP2E1 inhibitors may become an effective
therapeutic approach; however, to the best of our knowledge, no
relevant studies have been reported so far.
Therefore, the present study aimed to investigate
whether DAS can be used to inhibit the development of the
pathological process of DCM and examined the possible underlying
mechanism. For this purpose, cTnTR141W transgenic mice,
a mouse model with typical DCM phenotypes, such as decreased
survival rate, dilated chambers, thin walls and cardiac
dysfunction, which were successfully established in our previous
studies (4,26,27), were used. Furthermore, the
possible inhibitory mechanism of DAS on development of DCM was also
investigated in the present study.
Materials and methods
Animals
The a-MHC-cTnTR141W transgenic mice were
generated in the Key Laboratory of Human Disease Comparative
Medicine and maintained in a C57BL/6J genetic background in the Key
Laboratory of Human Disease Comparative Medicine (Ministry of
Health, Peking Union Medical College, Beijing, China), and
exhibited DCM phenotypic characteristics consistent with those
reported previously (4). A total
of 58 4-month-old mice were used, including 40 cTnTR141W
transgenic mice and 18 wild-type mice. In total, there were 34
males weighing 28–32 g, and 26 females weighing 22–24 g. All mice
were bred in an American Association for Accreditation of
Laboratory Animal Care-accredited facility, housed at 21±2°C with
50±5% relative humidity under a 12-h day/night cycle and had free
access to food and drinking water. The procedures were approved by
the Animal Care and Use Committee at the Institute of Laboratory
Animal Science, Peking Union Medical College (approval no.
ZLF18004).
Groups and treatment
The 4-month-old male and female cTnTR141W
transgenic mice were randomly assigned to treatment groups. DAS
(cat. no. A35801; Sigma-Aldrich; Merck KGaA) was diluted in corn
oil to a final concentration of 80 mg/ml. In the treatment groups,
cTnTR141W transgenic mice were administered DAS at a
dose of 200 (n=12) or 400 mg/kg (n=10) via intraperitoneal
injection three times weekly for 6 weeks. A group of
cTnTR141W transgenic mice (n=9) and non-transgenic
littermates (NTG; n=8) were treated with corn oil (0.1 ml per
mouse) as the placebo control and wild-type normal control,
respectively. As a commercially available drug control, a group of
cTnTR141W transgenic mice were treated with enalaprilat
(cat. no. H20010498; Changzhou Pharmaceutical Factory) via
intraperitoneal injection three times weekly for 6 weeks, an
angiotensin-converting enzyme (ACE) inhibitor that has been widely
used in the clinical treatment of DCM and HF (33,34), at a dose of 0.76 mg/kg (n=9). The
dose of DAS was selected based on previous reports (35,36). Therefore, there were total five
groups in this study, including NTG normal control, placebo model
control, DAS treatment at high dose (400 mg/kg), DAS treatment at
low dose (200 mg/kg) and enalaprilat. After the last
echocardiograph examination, the animals were euthanasia via
cervical dislocation.
Echocardiography
Heart function and structure were analyzed via
echocardiography once every 2 weeks during treatment. Briefly, the
mice were lightly anesthetized via an intraperitoneal injection of
216 mg/kg tribromoethanol and then subjected to 2-D guided M-mode
echocardiography with a 30-MHz transducer for echocardiographic
examination (Vevo770; FUJIFILM VisualSonics Inc.). Measurements of
left ventricular (LV) fractional shortening (LVFS), LV ejection
fraction (LVEF), LV diameter at end systole (LVESD), LV diameter at
end diastole (LVEDD), LV posterior wall at end systole (LVPWS), LV
posterior wall at end diastole (LVPWD), LV anterior wall at end
systole (LVAWS) and LV anterior wall at end diastole (LVAWD) were
based on the analysis of ≥10 separate cardiac cycles.
Histological analysis
For light microscopy, heart tissues were first fixed
in 4% formaldehyde at room temperature for 24 h, embedded in
paraffin and cut into 4-µm thick sections (4). Heart tissue sections were imaged
after H&E and Masson staining. For H&E staining, the
sections were stained using a Hematoxylin and Eosin Staining kit
(cat. no. G1121; Beijing Solarbio Science & Technology Co.,
Ltd.). Briefly, paraffin-embedded sections were dewaxed in xylene
two times for 10 min each time at room temperature. The sections
were rehydrated in a series of ethanol (100, 95, 85 and 75%) for 3
min per gradient, and for 2 min in distilled water. Subsequently,
the sections were stained with hematoxylin stain for 3 min at room
temperature and washed with distilled water to remove floating
colors. The differentiation solution was differentiated for 3 min
and washed with distilled water twice, for 2 min each time. The
sections were put into eosin dye solution for 2 min, then washed
with distilled water for 2–3 sec at room temperature. Then, the
sections were proceeded to the dehydrated step through soaking in a
series of concentrations of alcohol at room temperature (for 2–3
sec at 75, 85 and 95%, for 1 min at 100%). The sections were then
incubated with xylene twice at room temperature (for 1 min each
time), followed by sealing in neutral gum and finally observed
under the microscope.
For Masson trichrome staining, the sections were
incubated in celestine blue solution (cat. no. G1345; Beijing
Solarbio Science & Technology Co., Ltd.) for 5 min and briefly
washed with H2O. Then, sections were incubated in
hemalun solution for 5 min and in fuchsine acid/ponceau xylidine
(0.5% fuchsine acid, 1.5% ponceau xylidine, 1,75% glacial acetic
acid) for 5 min. Subsequent, sections were briefly washed with
H2O and incubated in phosphomolybdic acid (1%) for 10
min and in aniline blue solution (2.5% anilin blue, 2.5% glacial
acetic acid) for 5 min. Next, they were briefly washed with
H2O and incubated in acetic acid (1%) for 1 min.
Finally, the sections were briefly incubated in an ascending
ethanol series followed by xylol, before they were embedded.
Transmission electron microscopy
(TEM)
Heart tissues (~2 mm3) from the free wall
of the left ventricle were fixed in 2.5% glutaraldehyde
(preparation for 100 ml solution: 50 ml of 0.2 M phosphate buffer,
10 ml of 25% glutaraldehyde and 40 ml double distilled water) at
4°C for 12 h (4). After washing
three times with phosphate solution (pH=7.2), the samples were
fixed in 1% osmium acid at 4°C for 2 h. Dehydration was carried out
sequentially with four graded concentrations of ethanol between
50–100% at room temperature. After epichlorohydrin replacement and
epoxy embedding at room temperature, tissue sections were cut on an
ultramicrotome. The semi-thin sections (thickness, 900 nm) were
further stained with an equal mixture of 4% uranyl acetate and
acetone for 30 sec at room temperature, and then in lead citrate
for 2 min. The final ultra-thin sections (90 nm) were analyzed
under a JEM-1230 transmission electron microscope (JEOL Ltd.).
Measurement of ROS
After treatment, the mice were sacrificed and total
lysates were prepared as previously reported (4). H2O2,
malondialdehyde (MDA; lipid peroxidation) and glutathione (GSH) in
heart tissues were measured with relevant assay kits
(H2O2 Assay kit, cat. no. ab102500; MDA Assay
kit, cat. no. ab118970; GSH/GSSG Ratio Detection Assay kit, cat.
no. ab138881; all from Abcam) following the manufacturer's
procedures.
For the H2O2 assay, 10 mg
tissue was first washed in cold PBS and homogenized in 500 µl assay
buffer with a Dounce homogenizer sitting on ice. The sample was
centrifuged 13,000 × g for 5 min at 4°C at top speed using a
microcentrifuge to remove any insoluble material. Finally, the
content of H2O2 in the heart tissue was
measured with a microplate reader at excitation/emission
(Ex/Em)=535/587 nm.
For the lipid peroxidation assay, 10 mg tissue was
first washed in cold PBS and homogenized in 303 µl lysis solution
with a Dounce homogenizer sitting on ice. The sample was
centrifuged at 13,000 × g for 10 min at 4°C to remove insoluble
material. Then, thiobarbituric acid (TBA) reagent was added into
the supernatant to generate MDA-TBA adduct. Finally, the absorbance
was measured immediately at Ex/Em=532/553 nm.
For the glutathione assay, 10 mg tissue was first
washed in cold PBS, then was resuspended in 400 µl cold lysis
buffer. The samples were homogenized and centrifuged at 13,000 × g
for 15 min at 4°C to remove any insoluble material. Next, 50 µl GSH
assay mixture was added into each GSH standard or collected sample
supernatant. Finally, the absorbance was measured at Ex/Em=490/520
nm with a fluorescence microplate reader.
TUNEL assay
For apoptotic cell staining, paraffin sections of
heart tissues were stained using an ApopTag Plus Peroxidase In
Situ Apoptosis kit (cat. no. S7101; MilliporeSigma) following
the manufacturer's procedures. In briefly, heart tissues were first
fixed in 4% formaldehyde at room temperature for 24 h, embedded in
paraffin and cut into 4-µm thick sections. The tissue slides were
rehydrated and were treated with 25 µg/ml proteinase K at 37°C for
8 min at room temperature. After washing and incubation with
equilibration buffer for 5 min at room temperature, Tdt was diluted
at 1:3.9 with reaction buffer and added to heart sections for 1 h
at 37°C. After applying stop solution for 10 min at room
temperature, the samples were incubated with anti-digoxigenin
peroxidase conjugate at 37°C for 30 min. Slides were first treated
with a 1:20 dilution of diaminobenzidine (3,3′-diaminobenzidine)
substrate for 1 min at room temperature, then counterstained with
hematoxylin (cat. no. G1121; Beijing Solarbio Science &
Technology Co., Ltd.) for 30 sec at room temperature. The sections
were proceeded to the dehydrated step through soaking in a series
of concentrations of alcohol at room temperature (for 2–3 sec at
75, 85 and 95%, for 1 min at 100%). Then, the sections were
incubated with xylene twice at room temperature (for 1 min each
time), followed by sealing in neutral gum and finally observed
under the microscope. In total, nine visual fields were randomly
selected in each group to observe the apoptosis under a light
microscope (BX53; Olympus Corporation; magnification, ×400). The
results are expressed as the percentage of apoptotic cells among
the total cell population.
Reverse transcription (RT)-PCR
Total RNA was isolated from heart tissues using
TRIzol® reagent (cat. no. 15596018; Invitrogen; Thermo
Fisher Scientific, Inc.) and were used to synthesize cDNA with a RT
kit (cat. no. RR820A; Takara Bio, Inc.) following the
manufacturer's procedures. mRNA expression levels of procollagen
type III α1 (Col3α1) was detected via RT-PCR using GAPDH for
normalization under standard conditions. The primers were as
follows: Col3α1 forward, 5′-CTCAAGAGCGGAGAATACTGG-3′ and reverse,
5′-CAATGTCATAGGGTGCGATA-3′; and GAPDH forward,
5′-CAAGGTCATCCATGACAACTTTG-3′ and reverse,
5′-GTCCACCACCCTGTTGCTGTAG-3′.
Western blotting
Heart tissues were homogenized and extracted using
lysis buffer [tissue protein extraction reagent (cat. no. 78510);
protease inhibitor cocktail (cat. no. 87785); phosphatase inhibitor
cocktail (cat. no. 78420); PMSF (100 µM; cat. no. 36978; all from
Thermo Fisher Scientific, Inc.); 100:1:1:1 ratio for
configuration], followed by fractionation using a
Mitochondrial/Cytosol Fractionation kit (cat. no. ab65320; Abcam)
to obtain the cytosolic and mitochondrial fractions. An enhanced
BCA protein assay kit (cat. no. P0010; Beyotime Institute of
Biotechnology) was used to quantify the protein concentration. The
total lysates, cytosolic and mitochondrial fractions (50 µg) were
separated via 15% SDS-PAGE and transferred to a nitrocellulose
membrane (Immobilon NC; MilliporeSigma). Following blocking in 5%
fat-free milk in TBS-0.1% Tween-20 (TBST) for 1 h at room
temperature, the membranes were incubated at 4°C overnight with
primary antibodies targeted against: CYP2E1 (1:500; cat. no.
ab28146), cardiac troponin T (1:500; cTnT; cat. no. ab8295),
cytochrome c (1:500; cat. no. ab13575), procaspase 3
(1:1,000; cat. no. ab13847), procaspase 9 (1:1,000; cat. no.
ab47537; all from Abcam), cleaved (active) caspase 3 (1:500; cat.
no. 9507S) and cleaved caspase 9 (1:500; cat. no. 9664S; both from
Cell Signaling Technology, Inc.), anti-β-tubulin (1:1,000; cat. no.
ab21058) and anti-voltage dependent anion channel 1 (1:1,000; cat.
no. ab14734; both from Abcam) were used. Following washing with
TBST, the membranes were incubated with a HRP-conjugated secondary
antibody (1:10,000; anti-rabbit IgG, cat. no. ZB-2301 or anti-mouse
IgG, cat. no. ZB-2305; ZSGB-BIO, Inc.) at room temperature for 1 h.
Primary antibody binding was visualized using a chemiluminescent
detection system (Western Blotting Luminal Reagent; Santa Cruz
Biotechnology, Inc.) and analyzed using the densitometry function
of Quantity One software (version 3.0; Bio-Rad Laboratories,
Inc.).
Cell lines and culture
H9c2 cells (National Laboratory Cell Resource
Sharing Service Platform) were cultured in DMEM (Thermo Fisher
Scientific, Inc.), supplemented with 10% FBS (Thermo Fisher
Scientific, Inc.) and 100 U/ml penicillin-100 µg/ml streptomycin
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2. Cells were treated with DAS (12 h; 100 µmol/l; cat.
no. A35801; Sigma-Aldrich; Merck KGaA) or isoprenaline
hydrochloride (ISO; 12 h; 50 µmol/l; cat. no. I5627; Sigma-Aldrich;
Merck KGaA) at 37°C with 5% CO2 as required.
Calcein-AM/PI double staining
Cell viability was evaluated using calcein-AM/PI
double staining following the manufacturer's instruction (cat. no.
C542; Dojindo Laboratories, Inc.). Briefly, H9c2 cells were seeded
in 96-well plates in triplicate at a density of 1×104
cells/100 µl/well with 100 µl culture medium and cultured for 24 h.
The cells were treated with ISO (50 µmol/l) or DAS (100 µmol/l) for
12 h at 37°C with 5% CO2 and divided into four groups,
including control, ISO, control + DAS and ISO + DAS groups. The
cells were then washed twice with PBS and the working solution
(PBS: calcein-AM: PI=1,000:1:1) and incubated for 15 min at 37°C.
Live cells with yellow-green fluorescence and dead cells with red
fluorescence (Leica Microsystems GmbH) were observed using a
fluorescence microscope (magnification, ×100) at 490±10 nm and 545
nm excitation wavelengths. In total, three images of each well were
captured for a total of three replicate wells for counting.
Statistical analysis
All experiments were performed ≥3 times. Statistical
analyses were performed using SPSS software (version 19.0; IBM
Corp.). The data were analyzed using two-tailed unpaired t-tests
and one-way ANOVA followed by Tukey's post hoc analysis. Data are
presented as the mean ± SD. P<0.05 was considered to indicate a
statistically significant difference.
Results
DAS improves cardiac morphology
breakage and dysfunction in cTnTR141W DCM model
mice
Compared with the NTG group, the
cTnTR141W transgenic mice showed typical DCM phenotypes,
such as dilated chambers, thin walls and cardiac dysfunction. The
DCM phenotypes were evidenced by increases in LVESD and LVEDD, and
decreases in LVPWS, LVPWD, LVAWS, LVAWD, LVEF and LVFS (Fig. 1A-I; Table SI,Table SII,Table SIII).
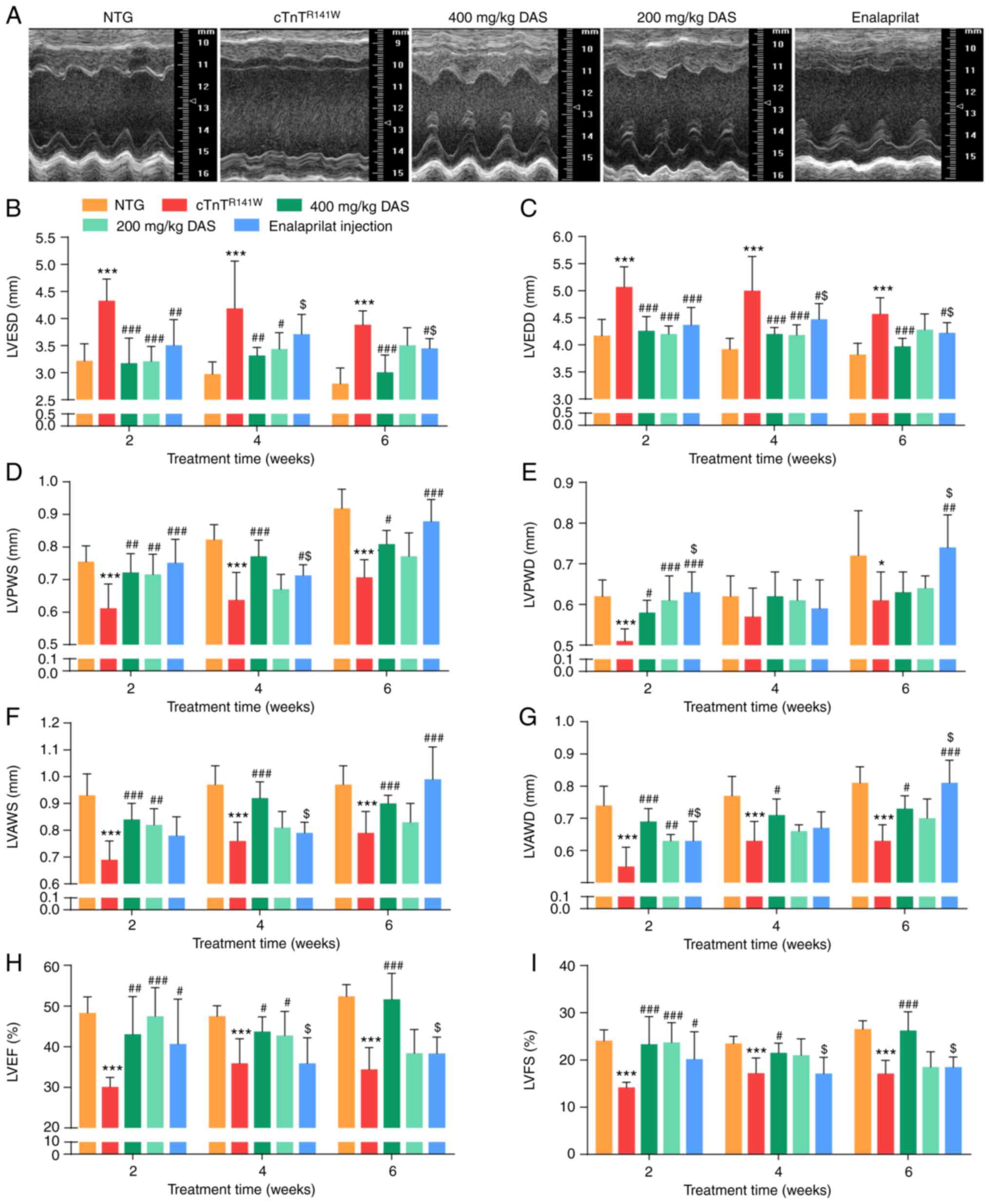 | Figure 1.Echocardiographic analysis of cardiac
morphology and function. Mice in the five groups were analyzed
after 6 weeks of treatment: NTG (wild-type control),
cTnTR141W (placebo control), treatment with 400 mg/kg
DAS, treatment with 200 mg/kg DAS and treatment with enalaprilat
(commercially available drug control). (A) Representative M-mode
echocardiographic images of the LV long axis. (B) LVESD. (C) LVEDD.
(D) LVPWS. (E) LVPWD. (F) LVAWS. (G) LVAWD. (H) LVEF. (I) LVFS.
n=7. *P<0.05, ***P<0.001 vs. NTG group;
#P<0.05, ##P<0.01,
###P<0.001 vs. cTnTR141W group;
$P<0.05 vs. 400 mg/kg DAS group. NTG, non-transgenic;
LV, left ventricular; LVESD, left ventricular end-systole diameter;
LVEDD, left ventricular end-diastole diameter; LVPWS, left
ventricular ventricle posterior wall at end systole; LVPWD, left
ventricular posterior wall at end diastole; LVAWS, left ventricular
anterior wall at end systole; LVAWD, left ventricular anterior wall
at end diastole; LVEF, left ventricular ejection fraction; LVFS,
left ventricular fractional shortening; DAS, diallyl sulfide. |
Firstly, after 2 weeks of treatment, the early stage
of drug intervention, it was found that improvement was already
observed in both DAS groups, and echocardiographic parameters in
both DAS groups showed an improved inhibition trend compared with
that of the enalaprilat group. For example, LVESD decreased by 26.5
and 25.9% in the DAS high and low dose group, respectively,
compared with the placebo model control group (P<0.001; Fig. 1B), while LVESD decreased by 18.9%
in the enalaprilat group compared with the placebo model control
group (P<0.01; Fig. 1B).
Moreover, LVEF increased by 57.8 and 43.1% in the DAS high and low
dose group, respectively, compared with the placebo model control
group (P<0.001; Fig. 1H),
while LVEF increased by 35.2% in the enalaprilat group compared
with the placebo model control group (P<0.05; Fig. 1H).
After 6 weeks of treatment, LVESD decreased by 22.4%
in the DAS high dose group (P<0.001; Fig. 1B), while it decreased by 11.1% in
the enalaprilat group (P<0.05; Fig. 1B) compared with the placebo model
control group. There was a significant increase in LVEF in the DAS
high dose group compared with the placebo model control group
(P<0.001; Fig. 1H), while
there was no difference between the enalaprilat group and the
placebo model control group (P>0.05; Fig. 1H). Furthermore, there was a
significant increase in LVAWD in the enalaprilat group compared
with the DAS high dose group (P<0.001; Fig. 1G). LVPWS increased in both the DAS
high dose group and the enalaprilat group compared with the placebo
model group (P<0.05; Fig. 1D),
while it showed no significant difference between these two groups
(P>0.05).
In summary, both DAS groups and the enalaprilat
group exhibited a significant improvement on DCM phenotypes, as
evidenced by changes in cardiac function, chamber size and wall
thickness. While each factor has its advantages, DAS exhibited a
stronger effect on control of chamber dilation and dysfunction, and
the enalaprilat group was more effective at increasing wall
thickness. Furthermore, this improvement on morphology and function
of model mice occurred earlier in both DAS groups compared with
that of the enalaprilat group.
DAS inhibits cardiac pathological
development in cTnTR141W DCM model mice
After drug treatment, hearts from all five groups
were sampled for gross morphology and pathological
examinations.
First of all, the protein expression level of CYP2E1
was determined via western blotting after treatment with different
doses of DAS. Compared with the placebo model control group, the
protein expression level of CYP2E1 in 200 and 400 mg/kg DAS groups
decreased by 35.5% (n=3; P<0.01) and 51.8% (n=3; P<0.001;
Fig. 2A and B), respectively.
Moreover, CYP2E1 protein expression decreased by 35.9% in the
enalaprilat group (n=3; P<0.01; Fig. 2A and B). The expression level of
cTnT protein was also analyzed. The mutant form of cTnT is the
human mutant form that was introduced by the transgenic method,
which is the cTnT band at the higher molecular weight in Fig. 2A, and the location of endogenous
cTnT proteins in mouse myocardium was slightly lower than that of
exogenous mutants. The expression level of the cTnT showed no
difference between the three therapeutic groups and the placebo
model group (Fig. 2A and C).
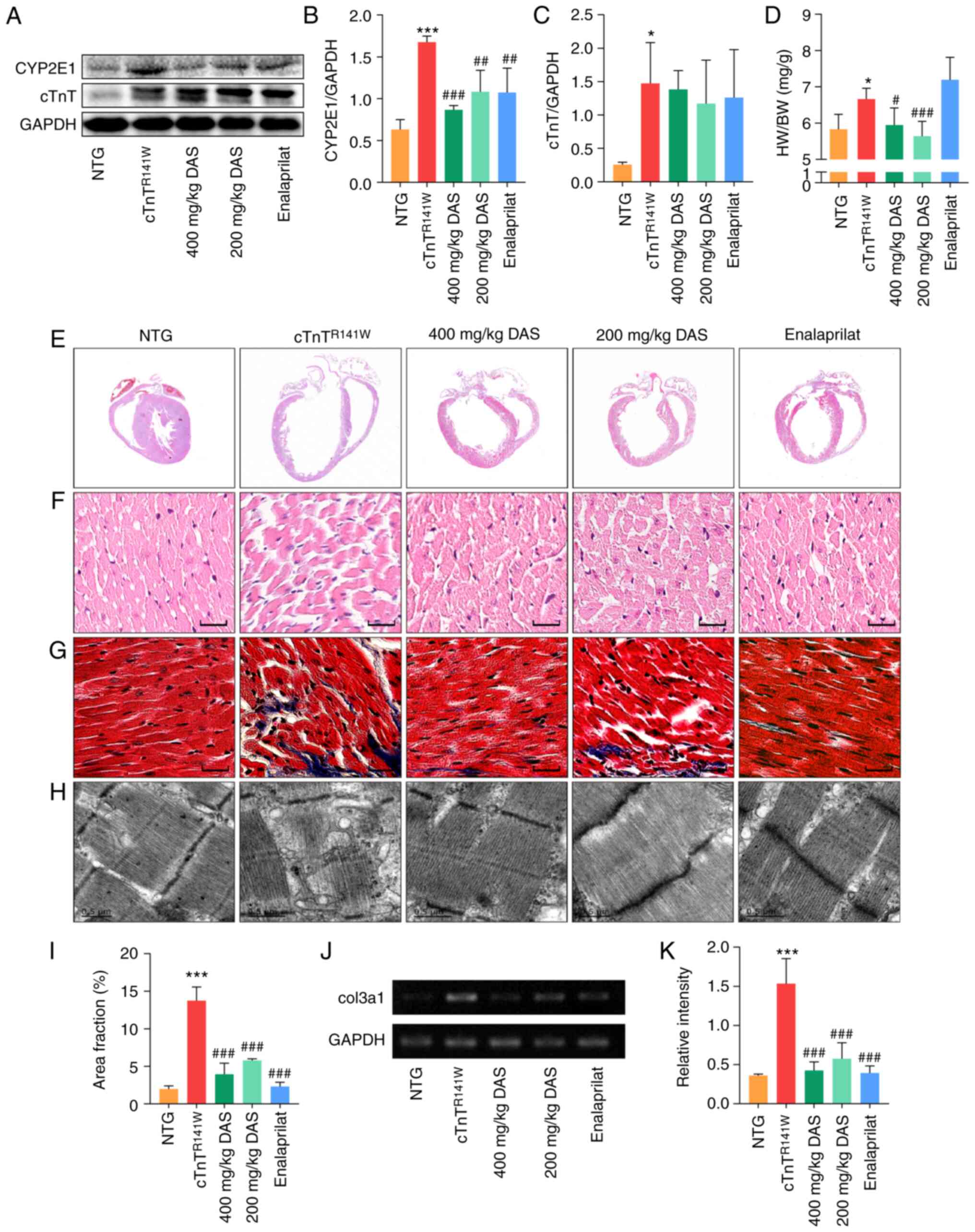 | Figure 2.Pathological histology observation.
(A) CYP2E1 and cTnT expression in the heart tissues of mice in the
NTG, cTnTR141W, 400, 200 mg/kg DAS and enalaprilat
groups was detected via western blotting. (B) CYP2E1 and (C) cTnT
were semi-quantitatively analyzed, using GAPDH for normalization
(n=3). (D) Ratio of HW to BW (n=6). (E) H&E staining patterns
of whole-heart longitudinal sections. (F) Magnification of
H&E-stained sections of the left ventricle (magnification,
×400; scale bar, 20 µm). (G) Magnification of Masson's
trichrome-stained left ventricle sections (magnification, ×400,
scale bar, 20 µm). Myocytes are stained red; collagenous tissue is
stained blue. (H) Ultrastructure observation via transmission
electron microscopy (scale bar, 0.5 µm). (I) Quantitative analysis
of Masson staining (n=3). (J) Col3α1 expression was detected and
(K) analyzed (n=3). *P<0.05, ***P<0.001 vs. NTG group;
#P<0.05, ##P<0.01,
###P<0.001 vs. cTnTR141W mice. cTnT,
cardiac troponin T; HW, heart weight; BW, body weight; Col3α1,
procollagen type III α1; NTG, non-transgenic; DAS, diallyl sulfide;
CYP2E1, cytochrome P450 family 2 subfamily E member 1. |
The increased heart to body weight ratio of
cTnTR141W mice was reversed by DAS treatment to almost
the normal level both in the 400 and 200 mg/kg group (P<0.05;
Fig. 2D). DAS treatment
significantly improved the chamber dilation, wall thinning and
myocyte disarray in cTnTR141W DCM model mice, as
determined via H&E staining (Fig.
2E and F). Through ultrastructure observation using TEM, it was
found that DAS treatment improved the poor myofibril organization,
including diffusion, damage and lysis, in cTnTR141W mice
(Fig. 2H). Quantitative analysis
of the Masson staining and the mRNA expression level of Col3α1
showed that collagen deposition in the interstitial space of
cTnTR141W mice was significantly reduced in both DAS and
enalaprilat groups compared with the placebo model control group
(Fig. 2G and I-K).
Both DAS and enalaprilat treatment exhibited a
favorable therapeutic effect on improving of the destroyed
microstructure and ultrastructure of the myocardium in DCM model
mice. Moreover, the DAS group exhibited a greater advantage in
reducing index of heart to body weight ratio.
To verify the ameliorative effect of DAS on the
pathological phenotype of cardiomyopathy in vitro. Cell
experiments were performed, and it was found that DAS could
significantly inhibit the increased expression of CYP2E1 induced by
ISO treatment in H9c2 cells, as well as significantly improve the
cell death induced by ISO (Fig.
S1), which were all consistent with the results in
vivo.
DAS reduces oxidative stress level in
cTnTR141W DCM model mice
CYP2E1 catalyzes the production of ROS even in the
absence of substrate, leading to oxidative stress (37). Therefore,
H2O2, MDA and GSH levels were measured as
indicators of oxidative stress in all five groups.
H2O2 and MDA levels were
significantly decreased (P<0.001; Fig. 3A and B), while GSH levels were
significantly increased (P<0.01 and P<0.001; Fig. 3C) in both 400 and 200 mg/kg DAS
treatment groups compared with those of the placebo model group.
Inhibition of the expression of CYP2E1 by DAS resulted in a
reversion of the levels of H2O2, MDA and GSH
in the myocardium to almost the normal levels in both the 400 and
200 mg/kg DAS groups. By contrast, no significant differences in
those three indicators were observed in the enalaprilat group
compared with the placebo model group (P<0.05; Fig. 3A-C).
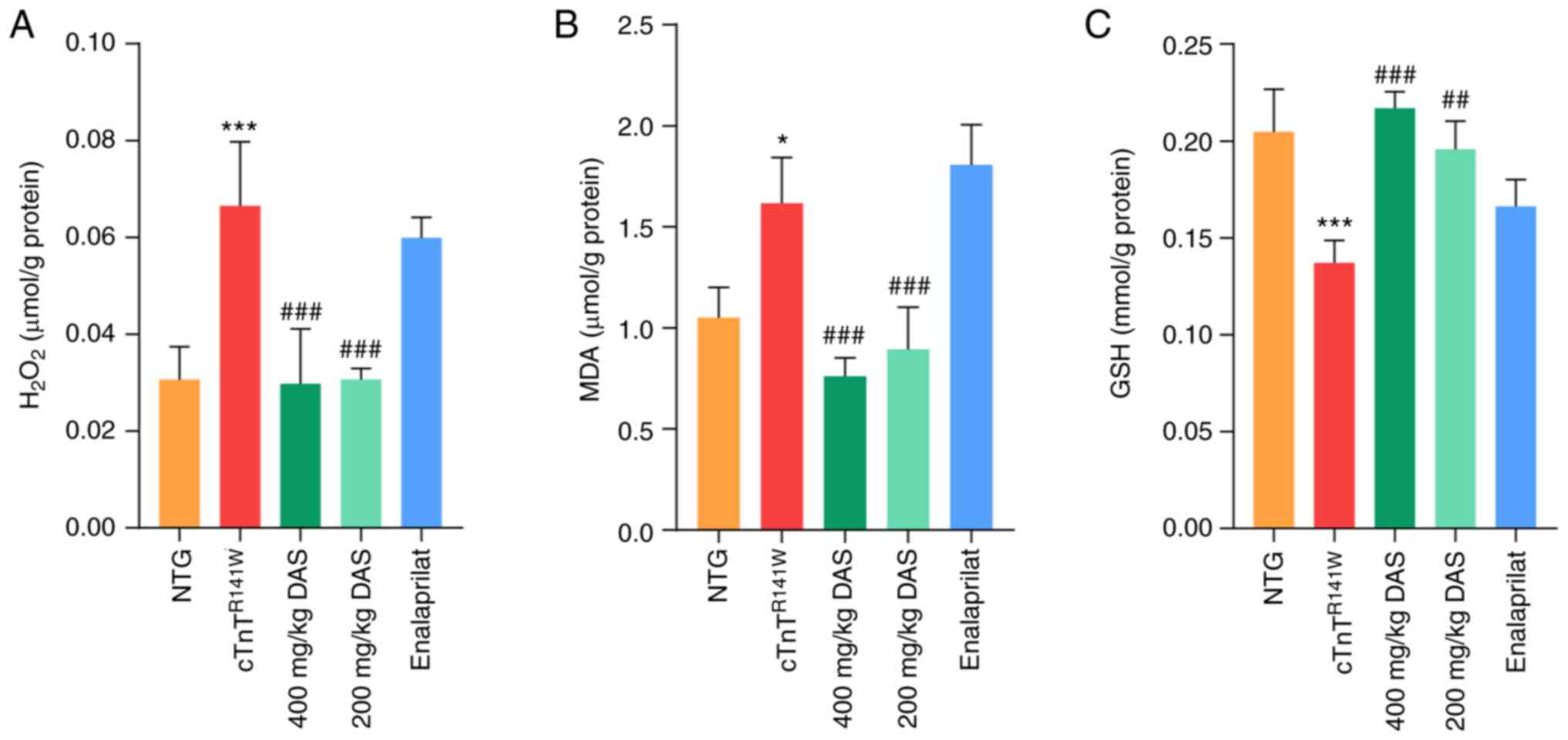 | Figure 3.Determination of oxidative stress
levels. Levels of (A) H2O2, (B) MDA and (C)
GSH in the heart tissues of mice in the NTG, cTnTR141W,
400, 200 mg/kg DAS and enalaprilat groups were determined using
colorimetric assays (n=3). *P<0.05, ***P<0.001 vs. NTG group;
##P<0.01, ###P<0.001 vs.
cTnTR141W mice. MDA, malondialdehyde; GSH, glutathione;
NTG, non-transgenic; DAS, diallyl sulfide. |
Therefore, inhibition of oxidative stress level is
one of the possible underlying mechanisms of DAS, rather than
enalaprilat, that is involved in the protection against the
pathological process of DCM.
DAS inhibits mitochondrial pathways of
apoptosis in cTnTR141W DCM model mice
The induction of CYP2E1 causes cytochrome c
release and activation of the mitochondrial apoptosis pathway in
the heart in cTnTR141W DCM model mice (4).
In both DAS groups, the increased release of
cytochrome c in cTnTR141W DCM model mice almost
reached those of normal levels (P<0.05; Fig. 4A and B). The activation of caspase
9 was decreased by 66.6% (P<0.001) and 66.9% (P<0.001;
Fig. 4A and C) in the 400 and 200
mg/kg DAS groups, respectively, compared with the placebo model
group. The activation of caspase 3 was decreased by 65.1%
(P<0.001) and 62.7% (P<0.001; Fig. 4A and D) in the 400 and 200 mg/kg
DAS groups, respectively, compared with the placebo model
group.
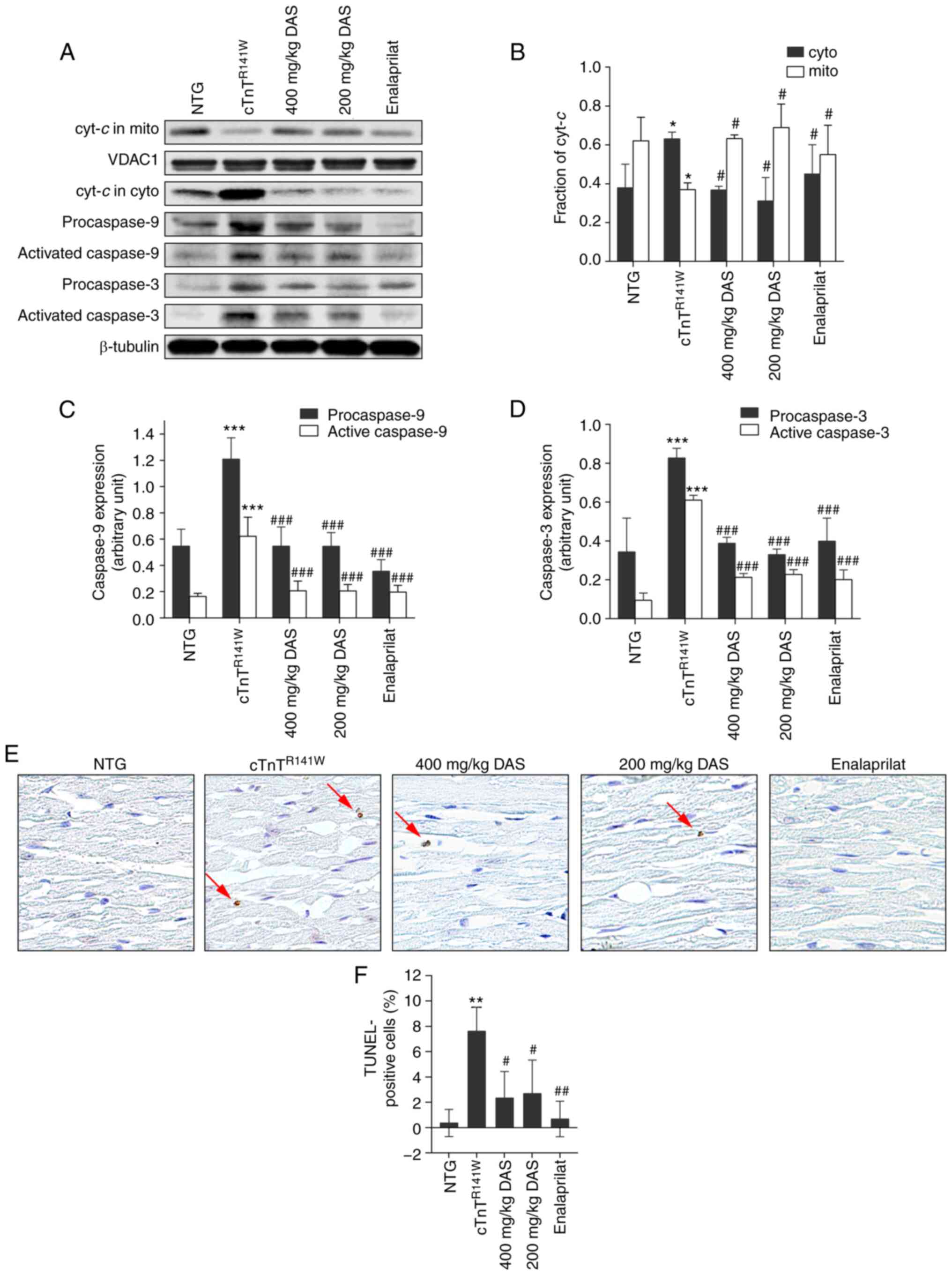 | Figure 4.Analysis of the
mitochondria-dependent apoptosis pathways. (A) Cyt-c release
and activation of caspase 9 and caspase 3 in the heart tissues of
mice in the NTG, cTnTR141W, 400, 200 mg/kg DAS and
enalaprilat groups were detected via western blotting. (B)
Cyt-c in the cyto and mito was semi-quantitatively analyzed
using β-tubulin or VDAC1 for normalization. (C) Procaspase 9 and
(D) procaspase 3 and active caspases 3 and 9 were
semi-quantitatively analyzed using β-tubulin for normalization
(n=3). (E) Cardiac myocyte apoptosis was detected using a TUNEL
assay, and the arrows indicate TUNEL-positive cells (magnification,
×400). (F) Number of positive cells was counted, and the proportion
of apoptotic cells among the total cells in each image was
calculated (n=3). *P<0.05, **P<0.01, ***P<0.001 vs. NTG
group; #P<0.05, ##P<0.01,
###P <0.001 vs. cTnTR141W mice.
Cyt-c, Cytochrome c; cyto, cytoplasm; mito,
mitochondria; VDAC1, voltage dependent anion channel 1; NTG,
non-transgenic; DAS, diallyl sulfide. |
The release of cytochrome c from the
mitochondria to the cytoplasm triggers the apoptosis of cardiac
myocytes in cTnTR141W model mice (38,39). It was determined that there were
69.3 and 64.7% decreases in proportion of apoptotic cells in the
400 (P<0.05) and 200 mg/kg groups (P<0.05; Fig. 4E and F), respectively, compared
with the placebo model group. Furthermore, DAS and enalaprilat
exhibited no difference in apoptosis inhibition.
Therefore, inhibition of the mitochondrial apoptosis
pathway is one of the possible mechanisms of DAS that is involved
in the protection against the pathological process of DCM.
Discussion
CYP2E1 is an inducible gene that is upregulated
under multiple conditions, such as fasting and nutrition intake, as
well as in a wide variety of pathophysiological states (2,12–17). In addition to metabolizing
endogenous substrates and xenobiotics, CYP2E1 is the main source of
cellular ROS, and its NADPH oxidase activity is higher than that of
other CYP family members (9–11).
Our previous studies revealed that the expression
level of CYP2E1 was increased in multiple mouse models of heart
disease, including those of DCM and HCM, and the possible mechanism
of upregulation of CYP2E1 expression were examined, in which it was
found that Myc was upregulated under pathological stimulus and
binds to the CYP2E1 promoter to activate its transcription
(26,27). Furthermore, it was determined that
the increase of CYP2E1 expression induced cardiac myocyte apoptosis
via mitochondrial pathways, and knockdown of endogenous of CYP2E1
expression using small interfering RNA could attenuate the
pathological development of DCM in cTnTR141W mice
(4).
Therefore, in the present study, DAS, a selective
inhibitor of CYP2E1, was used to treat cTnTR141W DCM
model mice, and to observe whether DAS can be used to inhibit the
development of the pathological process of DCM and its possible
mechanism.
DAS is an organosulfur compound derived from the
metabolism of allicin and has anti-cancer properties (40). DAS inhibits the activity of CYP2E1
and thus has attracted attention as a potential therapeutic or
prophylactic agent (41,42). DAS treatment has been applied in
several diseases. For instance, DAS treatment can attenuate the
pathogenesis of diseases associated with CYP2E1 upregulation in
animal models, such as alcoholic liver disease, nonalcoholic
steatohepatitis, diabetes and alcoholic cardiomyopathy (18,43–46). At present, to the best of our
knowledge, the use of DAS in treating DCM animal models to observe
its therapeutic effects had not been previously reported.
DAS, as an active ingredient in garlic, it is less
toxic than other garlic sulfides, such as diallyl disulfide and
diallyl trisulfide. DAS can be rapidly metabolized into diallyl
sulfoxide (DASO), diallyl sulfone (DASO2) and allyl mercaptan.
Allyl mercaptan is the decomposition product of DAS, and DASO and
DASO2 are sulfur oxidation products of DAS. DASO and DASO2 are the
main metabolites among DAS metabolites, and their formation is
mediated by the CYP2E1 enzyme. DAS is both a selective inhibitor
and a substrate of CYP2E1. Since sulfur is more nucleophilic than
DAS terminal carbon, sulfur strongly binds to heme of CYP2E1,
leading to competitive inhibition of CYP2E1 by DAS or metabolism
into DASO (18,47,48). DAS inhibits the metabolism of
P-450 2E1 substrates via competitive inhibition mechanisms and by
inactivating P-450 2E1 via a suicide-inhibitory action of DASO2
(18,47,48).
cTnTR141W mice develop typical DCM
phenotypes from 4 months of age, with death from HF after 8 months
of age (27,49). cTnTR141W transgenic
mice displayed typical familial dilated cardiomyopathy phenotypes
with dilated chambers, thin walls and cardiac dysfunction, as well
as pathological phenotypes with myocytes disarray and fibrosis
(4,26,27).
In the present study, it was identified that DAS
treatment improved the DCM phenotypes of chamber dilation, wall
thinning, myocyte disarray, fibrosis, poor myofibril organization
and decreased ventricular blood ejection in cTnTR141W
DCM model mice. Furthermore, DAS treatment inhibited ROS production
and decreased cytochrome c release, caspase 9-dependent
caspase 3 activation, and thus, apoptosis of myocytes in
cTnTR141W DCM model mice.
The current results are in line with others. DAS
analogues reduce the cytotoxicity associated with metabolism of
alcohol, analgesics and xenobiotic by inhibiting CYP2E1 activity.
In pathological conditions associated with CYP2E1 upregulation or
CYP2E1-mediated adverse reactions (including
diabetes/hyperlipidemia, Parkinson's disease, AIDS and cancer), DAS
can reduce the damage caused by the pathological stimulus (18). DAS has been reported as a
potentially effective intervention. The experimental results from
previous studies have shown that DAS could not only inhibit ethanol
and drug-mediated cytotoxicity, but also inhibit HIV protein- and
diabetes-mediated toxicities by selectively inhibiting CYP2E1 in
various cell types (18,50–53). In addition, the
antioxidant/anti-inflammatory effects of DAS further support its
use as dietary supplements (18).
The current research, along with that of others, indicated that DAS
could improve the symptoms of related diseases by targeting CYP2E1.
In addition, the present study examined a NTG normal group with DAS
treatment, and the heart structure and function of NTG group mice
treated with DAS (400 mg/kg) did not shown abnormality in the
echocardiography analysis (Table
SIV). However, DAS, as a result of its rapid metabolism to DASO
and DASO2, also causes cellular toxicity (18,54). Therefore, there is a need to
modify the parent DAS into an analogue, which is a stronger
inhibitor, but a weaker substrate of CYP2E1 (18,54).
Enalaprilat was selected in the present study to
compare it with the therapeutic effect and characteristics of DAS.
Enalaprilat is an ACE inhibitor that has been widely used in the
clinical treatment of HF, including in patients with DCM (33,34,55–57). Enalaprilat is characterized by
difficulties in direct absorption in the gastrointestinal tract and
low oral absorption, and in terms of renal clearance, there is a
barrier to entry of enalaprilat into the kidneys (58–60). DAS, as an active ingredient of
garlic, is less toxic than other sulfides in garlic. The
characteristic of DAS includes its action specificity, rapid
metabolic clearance and cytotoxicity; therefore, DAS can be
chemically modified to adjust its cytotoxicity and metabolic
clearance rate (18,47,48).
In the present study, both of DAS and enalaprilat
exhibited satisfactory inhibition on the pathological development
of DCM in cTnTR141W mice; however, each has its own
characteristics. DAS treatment showed advantages in improving
cardiac function and chamber dilation in vivo, as evidenced
by the changes in echocardiographic parameters of LVFS, LVEF, LVESD
and LVEDD. Furthermore, this improvement was more pronounced in the
early stage of the treatment in both DAS groups than in the
enalaprilat group, according to the significance findings after 2,
4 and 6 weeks of treatment. However, enalaprilat treatment was more
effective in improving the microstructure and ultrastructure of
myocardium. In terms of the mechanism of action, DAS, rather than
enalaprilat, exhibited an improved inhibitory effect on controlling
oxidative stress levels, using H2O2, MDA and
GSH as indicators. Moreover, both of DAS and enalaprilat showed an
excellent control effect on the mitochondrial apoptosis
pathway.
The results of enalapril in improving oxidative
stress are not satisfactory, and the exact mechanism remains
unknown. However, it has been reported that MDA levels were not
reduced in the enalapril-treated endothelial cells, while this
treatment improved those of glutathione peroxidase and superoxide
dismutase (61). This suggested
that the specific indicators of enalaprilat in controlling
oxidative stress are different from those detected in the present
study, which may partly explain why the enalaprilat does not show a
favorable control ability on improving MDA and other indicators of
oxidative stress.
In addition, the current study selected a widely
used myocardial cell line, H9c2, to verify the ameliorative effect
of DAS on the pathological phenotype of cardiomyopathy in
vitro. DAS could significantly inhibit the increased expression
of CYP2E1 induced by ISO in H9c2 cells, and it also significantly
improve the cell death induced by ISO, which were all consistent
with the results in vivo.
According to previous research, underlying cellular
processes, including inflammation and hypertrophy, myocyte
apoptosis and necrosis, and deposition of extracellular matrix,
have a direct association with the oxidative state of cardiac cells
(54,62). Garlic and its active ingredients
(such as phenols and saponins) have antioxidant effect (63). DAS can improve the oxidative
stress induced by thallium acetate (TI), reduce the increase of MDA
and nitric oxide levels in serum and liver, the decrease of GSH and
catalase activities in liver and the decrease of total antioxidant
capacity caused by TI (64). DAS
can oxidize and glycosylate low-density lipoprotein to prevent
additional oxidation or deterioration of glycosylation, which may
benefit patients with diabetes-related vascular disease (65). Garlic oil is a source of DAS, and
studies reported that garlic oil could be recommended as an option
for treating hypertrophic cardiovascular disease (66). Cardiovascular diseases involve
serum total cholesterol, low-density lipoprotein oxidation,
platelet aggregation and hypertension (66). In vitro studies have shown
garlic-induced inhibition of lipid synthesis, platelet aggregation,
erythrocyte lipid peroxidation and low-density lipoprotein
oxidation (66).
A limitation of the current study was that the
improvement of DAS occurred in the early stage of the treatment,
while the effect did not last a sufficient duration in the DAS low
dose group. Therefore, future work will examine the range of
diseases that DAS could be applied, as well as develop structural
analogues with a lower cytotoxicity and metabolic rate.
In conclusion, the present study demonstrated that
DAS had advantages in terms of improved chamber dilation and
increased ventricular blood ejection. Furthermore, the regulatory
mechanisms underlying DAS involvement in the DCM pathological
development, included inhibition of both the oxidative stress
levels and the mitochondria-dependent apoptosis pathways.
Supplementary Material
Supporting Data
Acknowledgements
The authors thank Dr Ya-jun Yang (Institute of
Material Medical, Chinese Academy of Medical Sciences and Peking
Union Medical College) for detection and analysis in the
pharmacokinetic study.
Funding
The present work was supported by Chinese Academy of
Medical Sciences Innovation Fund for Medical Sciences (grant no.
2016-I2M-1-015), National Natural Science Foundation of China
(grant no. 31872314) and Beijing Natural Science Foundation (grant
no. 5212017).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ conceived the experiments and wrote the paper. DL
analyzed the data and wrote the paper. SP performed most of the
experiments and analyzed the data, including echocardiography
parameters analysis, pathological analysis, western blotting,
biochemical analysis and immunohistochemical analysis. WD
contributed to the echocardiography procedure. NL and SG
contributed to the genotyping and animal breeding and management.
JL contributed to the TEM analysis. XZ contributed to animal
administration procedures and RT-PCR analysis. LZ and DL confirmed
the authenticity of all the raw data. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
All animal procedures were approved by the Animal
Care and Use Committee at the Institute of Laboratory Animal
Science, Peking Union Medical College (approval no. ZLF18004). All
mice were bred in an AAALAC-accredited facility and animal
experiments were carried out in accordance with animal welfare
requirements and the Guidelines for the Care and Use of Laboratory
Animals published by the Chinese Ministry of Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Leung T, Rajendran R, Singh S, Garva R,
Krstic-Demonacos M and Demonacos C: Cytochrome P450 2E1 (CYP2E1)
regulates the response to oxidative stress and migration of breast
cancer cells. Breast Cancer Res. 15:R1072013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sheng Y, Yang H, Wu T, Zhu L, Liu L and
Liu X: Alterations of cytochrome P450s and
UDP-glucuronosyltransferases in brain under diseases and their
clinical significances. Front Pharmacol. 12:6500272021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cederbaum AI: Alcohol metabolism. Clin
Liver Dis. 16:667–685. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu D, Ma Y, Zhang W, Bao D, Dong W, Lian
H, Huang L and Zhang L: Knockdown of cytochrome P450 2E1 inhibits
oxidative stress and apoptosis in the cTnT(R141W) dilated
cardiomyopathy transgenic mice. Hypertension. 60:81–89. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Murray B, Peng H, Barbier-Torres L,
Robinson AE, Li TWH, Fan W, Tomasi ML, Gottlieb RA, Van Eyk J, Lu
Z, et al: Methionine adenosyltransferase α1 is targeted to the
mitochondrial matrix and interacts with cytochrome P450 2E1 to
lower its expression. Hepatology. 70:2018–2034. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Harjumäki R, Pridgeon CS and
Ingelman-Sundberg M: CYP2E1 in alcoholic and non-alcoholic liver
injury. Roles of ROS, reactive intermediates and lipid overload.
Int J Mol Sci. 22:82212021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tomasi ML, Ramani K, Ryoo M, Cossu C,
Floris A, Murray BJ, Iglesias-Ara A, Spissu Y and Mavila N:
SUMOylation regulates cytochrome P450 2E1 expression and activity
in alcoholic liver disease. FASEB J. 32:3278–3288. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leung TM and Nieto N: CYP2E1 and oxidant
stress in alcoholic and non-alcoholic fatty liver disease. J
Hepatol. 58:395–398. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schattenberg JM and Czaja MJ: Regulation
of the effects of CYP2E1-induced oxidative stress by JNK signaling.
Redox Biol. 3:7–15. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Santra S, Bishnu D, Dhali GK, Santra A and
Chowdhury A: Expression of type I collagen in response to isoniazid
exposure is indirect and is facilitated by collateral induction of
cytochrome P450 2E1: An in-vitro study. PLoS One. 15:e02369922020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuzgun G, Başaran R, Arıoğlu İnan E and
Can Eke B: Effects of insulin treatment on hepatic CYP1A1 and
CYP2E1 activities and lipid peroxidation levels in
streptozotocin-induced diabetic rats. J Diabetes Metab Disord.
19:1157–1164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Paradies G, Paradies V, Ruggiero FM and
Petrosillo G: Oxidative stress, cardiolipin and mitochondrial
dysfunction in nonalcoholic fatty liver disease. World J
Gastroenterol. 20:14205–14218. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Navarro-Mabarak C, Camacho-Carranza R and
Espinosa-Aguirre JJ: Cytochrome P450 in the central nervous system
as a therapeutic target in neurodegenerative diseases. Drug Metab
Rev. 50:95–108. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baltazar MT, Dinis-Oliveira RJ, de Lourdes
Bastos M, Tsatsakis AM, Duarte JA and Carvalho F: Pesticides
exposure as etiological factors of Parkinson's disease and other
neurodegenerative diseases-a mechanistic approach. Toxicol Lett.
230:85–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Abdelmegeed MA, Ha SK, Choi Y, Akbar M and
Song BJ: Role of CYP2E1 in mitochondrial dysfunction and hepatic
injury by alcohol and non-alcoholic substances. Curr Mol Pharmacol.
10:207–225. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin X, Xiong W, Wang Y, Tang W, Xi W, Qian
S and Guo Y: Association of CYP2E1 gene polymorphisms with bladder
cancer risk: A systematic review and meta-analysis. Medicine
(Baltimore). 97:e119102018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Martinez-Gil N, Vidal-Gil L,
Flores-Bellver M, Maisto R, Sancho-Pelluz J, Diaz-Llopis M, M
Barcia J and Romero FJ: Ethanol-induced oxidative stress modifies
inflammation and angiogenesis biomarkers in retinal pigment
epithelial cells (ARPE-19): Role of CYP2E1 and its inhibition by
antioxidants. Antioxidants (Basel). 9:7762020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rao PS, Midde NM, Miller DD, Chauhan S,
Kumar A and Kumar S: Diallyl sulfide: Potential use in novel
therapeutic interventions in alcohol, drugs, and disease mediated
cellular toxicity by targeting cytochrome P450 2E1. Curr Drug
Metab. 16:486–503. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hong JY, Wang ZY, Smith TJ, Zhou S, Shi S,
Pan J and Yang CS: Inhibitory effects of diallyl sulfide on the
metabolism and tumorigenicity of the tobacco-specific carcinogen
4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) in A/J mouse
lung. Carcinogenesis. 13:901–904. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jin M, Ande A, Kumar A and Kumar S:
Regulation of cytochrome P450 2e1 expression by ethanol: Role of
oxidative stress-mediated pkc/jnk/sp1 pathway. Cell Death Dis.
4:e5542013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Morimoto M, Hagbjörk AL, Wan YJ, Fu PC,
Clot P, Albano E, Ingelman-Sundberg M and French SW: Modulation of
experimental alcohol-induced liver disease by cytochrome P450 2E1
inhibitors. Hepatology. 21:1610–1617. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang RH, Gao JY, Guo HT, Scott GI, Eason
AR, Wang XM and Ren J: Inhibition of CYP2E1 attenuates chronic
alcohol intake-induced myocardial contractile dysfunction and
apoptosis. Biochim Biophys Acta. 1832:128–141. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
van Riet EE, Hoes AW, Wagenaar KP, Limburg
A, Landman MA and Rutten FH: Epidemiology of heart failure: The
prevalence of heart failure and ventricular dysfunction in older
adults over time. A systematic review. Eur J Heart Fail.
18:242–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shakib S and Clark RA: Heart failure
pharmacotherapy and supports in the elderly-a short review. Curr
Cardiol Rev. 12:180–185. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
van der Pol A, van Gilst WH, Voors AA and
van der Meer P: Treating oxidative stress in heart failure: Past,
present and future. Eur J Heart Fail. 21:425–435. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang W, Lu D, Dong W and Zhang L, Zhang
X, Quan X, Ma C, Lian H and Zhang L: Expression of CYP2E1 increases
oxidative stress and induces apoptosis of cardiomyocytes in
transgenic mice. FEBS J. 278:1484–1492. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guan F, Yang X, Li J, Dong W, Zhang X, Liu
N, Gao S, Wang J, Zhang L and Lu D: New molecular mechanism
underlying Myc-mediated cytochrome P450 2E1 upregulation in
apoptosis and energy metabolism in the myocardium. J Am Heart
Assoc. 8:e0098712019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Westphal C, Konkel A and Schunck WH:
Cytochrome p450 enzymes in the bioactivation of polyunsaturated
Fatty acids and their role in cardiovascular disease. Adv Exp Med
Biol. 851:151–187. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Thum T and Borlak J: Cytochrome P450
mono-oxygenase gene expression and protein activity in cultures of
adult cardiomyocytes of the rat. Br J Pharmacol. 130:1745–1752.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aberle NS II and Ren J: Short-term
acetaldehyde exposure depresses ventricular myocyte contraction:
Role of cytochrome P450 oxidase, xanthine oxidase, and lipid
peroxidation. Alcohol Clin Exp Res. 27:577–583. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sidorik L, Kyyamova R, Bobyk V, Kapustian
L, Rozhko O, Vigontina O, Ryabenko D, Danko I, Maksymchuk O,
Kovalenko VN, et al: Molecular chaperone, HSP60, and cytochrome
P450 2E1 co-expression in dilated cardiomyopathy. Cell Biol Int.
29:51–55. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jing L, Jin CM, Li SS, Zhang FM, Yuan L,
Li WM, Sang Y, Li S and Zhou LJ: Chronic alcohol intake-induced
oxidative stress and apoptosis: Role of CYP2E1 and calpain-1 in
alcoholic cardiomyopathy. Mol Cell Biochem. 359:283–292. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kobaladze N, Tsibadze T, Iakobashvili M
and Tabidze G: Angiotensin-converting enzyme inhibitor treatment of
heart failure due to dilated cardiomyopathy. Georgian Med News.
41–44. 2005.(In Russian). PubMed/NCBI
|
|
34
|
Cernecka H, Ochodnicka-Mackovicova K,
Kucerova D, Kmecova J, Nemcekova V, Doka G, Kyselovic J, Krenek P,
Ochodnicky P and Klimas J: Enalaprilat increases PPARβ/δ
expression, without influence on PPARα and PPARγ, and modulate
cardiac function in sub-acute model of daunorubicin-induced
cardiomyopathy. Eur J Pharmacol. 714:472–477. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Salminen WF Jr, Roberts SM, Pumford NR and
Hinson JA: Immunochemical comparison of 3′-hydroxyacetanilide and
acetaminophen binding in mouse liver. Drug Metab Dispos.
26:267–271. 1998.PubMed/NCBI
|
|
36
|
Saldaña-Ruíz S, Boadas-Vaello P,
Sedó-Cabezón L and Llorens J: Reduced systemic toxicity and
preserved vestibular toxicity following co-treatment with nitriles
and CYP2E1 inhibitors: A mouse model for hair cell loss. J Assoc
Res Otolaryngol. 14:661–671. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cederbaum AI, Wu D, Mari M and Bai J:
CYP2E1-dependent toxicity and oxidative stress in HepG2 cells. Free
Radic Biol Med. 31:1539–1543. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wollert KC, Heineke J, Westermann J, Lüdde
M, Fiedler B, Zierhut W, Laurent D, Bauer MK, Schulze-Osthoff K and
Drexler H: The cardiac Fas (APO-1/CD95) receptor/Fas ligand system:
Relation to diastolic wall stress in volume-overload hypertrophy in
vivo and activation of the transcription factor AP-1 in cardiac
myocytes. Circulation. 101:1172–1178. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yaniv G, Shilkrut M, Lotan R, Berke G,
Larisch S and Binah O: Hypoxia predisposes neonatal rat ventricular
myocytes to apoptosis induced by activation of the Fas (CD95/Apo-1)
receptor: Fas activation and apoptosis in hypoxic myocytes.
Cardiovasc Res. 54:611–623. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Thomson M and Ali M: Garlic (Allium
sativum): A review of its potential use as an anti-cancer agent.
Curr Cancer Drug Targets. 3:67–81. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nicastro HL, Ross SA and Milner JA: Garlic
and onions: Their cancer prevention properties. Cancer Prev Res
(Phila). 8:181–189. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
McCaskill ML, Rogan E and Thomas RD:
Diallyl sulfide inhibits diethylstilbestrol induced DNA damage in
human breast epithelial cells (MCF-10A). Steroids. 92:96–100. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Begriche K, Knockaert L, Massart J, Robin
MA and Fromenty B: Mitochondrial dysfunction in nonalcoholic
steatohepatitis (NASH): Are there drugs able to improve it? Drug
Discov Today Dis Mech. 6:e11–e23. 2009. View Article : Google Scholar
|
|
44
|
Louvet A and Mathurin P: Alcoholic liver
disease: Mechanisms of injury and targeted treatment. Nat Rev
Gastroenterol Hepatol. 12:231–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Starley BQ, Calcagno CJ and Harrison SA:
Nonalcoholic fatty liver disease and hepatocellular carcinoma: A
weighty connection. Hepatology. 51:1820–1832. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wieckowska A, McCullough AJ and Feldstein
AE: Noninvasive diagnosis and monitoring of nonalcoholic
steatohepatitis: Present and future. Hepatology. 46:582–589. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Brady JF, Ishizaki H, Fukuto JM, Lin MC,
Fadel A, Gapac JM and Yang CS: Inhibition of cytochrome P-450 2E1
by diallyl sulfide and its metabolites. Chem Res Toxicol.
4:642–647. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Brady JF, Wang MH, Hong JY, Xiao F, Li Y,
Yoo JS, Ning SM, Lee MJ, Fukuto JM, Gapac JM, et al: Modulation of
rat hepatic microsomal monooxygenase enzymes and cytotoxicity by
diallyl sulfide. Toxicol Appl Pharmacol. 108:342–354. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tayal U, Prasad S and Cook SA: Genetics
and genomics of dilated cardiomyopathy and systolic heart failure.
Genome Med. 9:202017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wu D and Cederbaum AI: Cyclosporine A
protects against arachidonic acid toxicity in rat hepatocytes: Role
of CYP2E1 and mitochondria. Hepatology. 35:1420–1430. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Dai Y and Cederbaum AI: Cytotoxicity of
acetaminophen in human cytochrome P4502E1-transfected HepG2 cells.
J Pharmacol Exp Ther. 273:1497–1505. 1995.PubMed/NCBI
|
|
52
|
Isaguliants M, Smirnova O, Ivanov AV,
Kilpelainen A, Kuzmenko Y, Petkov S, Latanova A, Krotova O,
Engström G, Karpov V, et al: Oxidative stress induced by HIV-1
reverse transcriptase modulates the enzyme's performance in gene
immunization. Hum Vaccin Immunother. 9:2111–2119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Swaminathan K, Kumar SM, Clemens DL and
Dey A: Inhibition of CYP2E1 leads to decreased advanced glycated
end product formation in high glucose treated ADH and CYP2E1
over-expressing VL-17A cells. Biochim Biophys Acta. 1830:4407–4416.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jin L and Baillie TA: Metabolism of the
chemoprotective agent diallyl sulfide to glutathione conjugates in
rats. Chem Res Toxicol. 10:318–327. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Travessa AM and Menezes Falcão L:
Vasodilators in acute heart failure-evidence based on new studies.
Eur J Intern Med. 51:1–10. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Momma K: ACE inhibitors in pediatric
patients with heart failure. Paediatr Drugs. 8:55–69. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Smeets NJL, Schreuder MF, Dalinghaus M,
Male C, Lagler FB, Walsh J, Laer S and de Wildt SN: Pharmacology of
enalapril in children: A review. Drug Discov Today. Aug
21–2020.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
58
|
de Lannoy IA, Nespeca R and Pang KS: Renal
handling of enalapril and enalaprilat: Studies in the isolated red
blood cell-perfused rat kidney. J Pharmacol Exp Ther.
251:1211–1222. 1989.PubMed/NCBI
|
|
59
|
Tocco DJ, deLuna FA, Duncan AE, Vassil TC
and Ulm EH: The physiological disposition and metabolism of
enalapril maleate in laboratory animals. Drug Metab Dispos.
10:15–19. 1982.PubMed/NCBI
|
|
60
|
Gross DM, Sweet CS, Ulm EH, Backlund EP,
Morris AA, Weitz D, Bohn DL, Wenger HC, Vassil TC and Stone CA:
Effect of N-[(S)-1-carboxy-3-phenylpropyl]-L-Ala-L-Pro and its
ethyl ester (MK-421) on angiotensin converting enzyme in vitro and
angiotensin I pressor responses in vivo. J Pharmacol Exp Ther.
216:552–557. 1981.PubMed/NCBI
|
|
61
|
Mailloux A, Deslandes B, Vaubourdolle M
and Baudin B: Captopril and enalaprilat decrease antioxidant
defences in human endothelial cells and are unable to protect
against apoptosis. Cell Biol Int. 27:825–830. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hantson P: Mechanisms of toxic
cardiomyopathy. Clin Toxicol (Phila). 57:1–9. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shang A, Cao SY, Xu XY, Gan RY, Tang GY,
Corke H, Mavumengwana V and Li HB: Bioactive compounds and
biological functions of garlic (Allium sativum L.). Foods.
8:2462019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Abdel-Daim MM and Abdou RH: Protective
effects of diallyl sulfide and curcumin separately against
thallium-induced toxicity in rats. Cell J. 17:379–388.
2015.PubMed/NCBI
|
|
65
|
Huang CN, Horng JS and Yin MC:
Antioxidative and antiglycative effects of six organosulfur
compounds in low-density lipoprotein and plasma. J Agric Food Chem.
52:3674–3678. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Suman S and Shukla Y: Diallyl sulfide and
its role in chronic diseases prevention. Adv Exp Med Biol.
929:127–144. 2016. View Article : Google Scholar : PubMed/NCBI
|














