Introduction
Intrauterine growth restriction (IUGR), defined as a
developing fetus weighing 10% or two standard deviations less than
the mean body weight of normal fetuses of the same gestational age
(1), has been proven to be an
independent risk factor of adult metabolic diseases, including
diabetes mellitus, obesity and coronary heart disease (2). Based on those findings, Barker
(2) introduced the theory of the
intrauterine programming of adult diseases, namely abnormal changes
in the morphology and function of the fetus caused by maternal
undernutrition, environmental pollutants and some other adverse
factors during pregnancy, which may persist until adulthood and
could be the origin of adult diseases (3).
Prenatal food restriction (PFR) causes IUGR of the
fetus. It was previously reported that IUGR of the offspring caused
by PFR resulted in an abnormal function of the
hypothalamic-pituitary-adrenal (HPA) axis (4). The altered intrauterine programming of
the HPA axis may be involved in the mechanism underlying the
intrauterine origin of adult diseases (4). Poore and Fowden (5) reported that stimulated serum cortisol
was detected in pigs with low birth weight in the first 3 months,
while an enhanced adrenal responsiveness to insulin-induced
hypoglycemia was observed at 12 months of age. Our previous study
found that the serum levels of adrenocorticotrophic hormone (ACTH)
and corticosterone (CORT) in the adult rat offspring with PFR were
lower compared with those in control rats before unpredictable
chronic stress (UCS), but were significantly increased after the
stress, which was accompanied by alterations in HPA axis-associated
neuroendocrine metabolism (6).
However, the underlying mechanism remains unknown.
Corticotrophin-releasing hormone (CRH) and arginine
vasopressin (AVP) are secreted by parvicellular neurons of the
paraventricular nucleus (PVN), a composite structure of the
hypothalamus, when the body is exposed to a stressor (7). Subsequently, CRH and AVP activate the
anterior pituitary, which releases ACTH. ACTH then promotes the
release of glucocorticoid (CORT in rodents or cortisol in humans)
by the adrenal gland (8).
Glucocorticoids activate the glucocorticoid receptor (GR) and
mineralocorticoid receptor (MR) in the hippocampus, which then
suppress the activity of the PVN in the hypothalamus via a feedback
loop (8). As PFR alters the serum
CORT levels and the function of the HPA axis in the offspring
(6), it was hypothesized that
disruption of the modulatory mechanism between the hippocampus and
the hypothalamus in the uterus may program the function of the HPA
axis to a low basic activity but a high susceptibility in the adult
offspring.
In the present study, an IUGR rat model of PFR was
constructed, and the male offspring were fed a high-fat diet to
magnify the susceptibility of the HPA axis. The adult male rat
offspring were then subjected to PFR. Behavioral changes,
histopathological changes of the hypothalamus, HPA axis function
and the modulatory mechanism between the hippocampus and the
hypothalamus were detected, which could help to identify the
underlying mechanism of the fetal programming of the HPA axis
function due to PFR.
Materials and methods
Materials
ACTH kits (cat. no. KIP0061) were purchased from the
North Institute of Biological Technology (Beijing, China). ELISA
kits for rat CORT were obtained from R&D Systems, Inc. (cat.
no. KGE009). TRIzol® reagent was obtained from Thermo
Fisher Scientific, Inc. (cat. no. 15596026). The First Strand cDNA
Synthesis kit and reverse transcription-quantitative PCR (RT-qPCR)
kits were purchased from Takara Biotechnology, Inc. (cat. nos.
6110A and RR039W). Oligonucleotide primers were synthesized by
Sangon Biotech Co., Ltd. Isoflurane was purchased from Baxter
International, Inc. Other chemicals and agents were of analytical
grade.
Animals and treatment
The animal experiments were performed in the Animal
Experiment Centre of Wuhan University, and were accredited by the
Association for Assessment and Accreditation of Laboratory Animal
Care International. All the procedures were in accordance with the
National Institutes of Health Guide for the Care and Use of
Laboratory Animals (8th edition) (9). All the protocols were approved by
Medical Ethics Committee of the Basic Medical School of Wuhan
University (approval no. 201719).
A total of 20 female Wistar rats (weight, 180–220 g)
and 10 male rats (weight, 260–300 g) were purchased from the
Experimental Centre of Hubei Medical Scientific Academy (animal
registration no. 2017-0004). All the rats were specific
pathogen-free and were aged 3 months. The animal feeding, grouping
and treatment were performed as described in our previous study
(6). The animals were maintained at
a constant temperature (18–22°C) and humidity (40–60%), under a
controlled 12-h light/dark cycle, with free access to water and
standard rodent chow. Two female rats were mated with one male rat
overnight after 1 week of acclimation. Mating was confirmed by the
appearance of sperm in a vaginal smear the next morning; this day
was designated as gestational day (GD) 0. Pregnant female rats were
then transferred to individual cages. Pregnant rats were fed ad
libitum or put on a restricted diet (50% of the daily food
intake of control rats, ~60 g/kg body weight) from GD11 until term
delivery. The offspring rats were kept in the same condition, but
fed a high-fat diet. Rodent chow was purchased from the
Experimental Centre of Hubei Medical Scientific Academy. The
standard rodent chow contained 21% kcal from protein, 68.5% kcal
from carbohydrate and 10.5% kcal from fat, while the high-fat diet
contained 88.0% corn flour, 11.5% lard and 0.5% cholesterol, which
provided 18.9% kcal from protein, 61.7% kcal from carbohydrate and
19.4% kcal from fat. In addition, behavioral changes of the rats
were examined using the open-field test, electronic maze experiment
and sucrose preference test following the standard protocols
(10,11). Animals were anesthetized using 5%
isoflurane at a flow of 0.4 l/min for the induction of inhalation
anesthesia. Then, the animals were sacrificed by decapitation after
being anesthetized. The whole brain was dissected for histological
examination of the hippocampus and hypothalamus. H&E staining
of the hypothalamus slices was applied to check whether the PVN was
included in the hypothalamus samples.
Assay for blood samples
Blood samples (250 µl) were collected though the
lateral tail vein from the same set of animals before (b)UCS and
after (a)UCS. Serum was collected from the whole blood via
centrifugation at 1,200 × g and 4°C for 10 min. The serum
concentrations of ACTH and CORT were determined following our
previously reported protocol (6).
The serum concentration of ACTH was determined using
radioimmunoassay kits, while the concentration of serum CORT was
determined with ELISA kits, following the manufacturer's protocol.
In addition, the rates of the increase in the serum concentrations
of serum ACTH and CORT were calculated as the following
formula:
RT-qPCR assay
Total RNA was collected from 30 mg hippocampal or
hypothalamic tissue using TRIzol reagent. Single-strand cDNA was
obtained using a First Strand cDNA Synthesis kit. The RT reaction
was performed at 50°C for 45 min to reduce the possibility of
non-specific amplification products. The primers were designed
using Primer Premier 5.0 (PREMIER Biosoft) and the NCBI BLAST
database (https://blast.ncbi.nlm.nih.gov/Blast.cgi; National
Center for Biotechnology Information) (12). The RT-qPCR assay was performed on a
StepOne thermal cycler (Thermo Fisher Scientific, Inc.) using the
Takara RT-qPCR kits under the following thermocycling conditions:
Initial denaturation at 95°C for 30 sec; 40 cycles of denaturation
at 95°C for 5 sec, annealing at 60°C for 30 sec and elongation at
72°C for 30 sec; final extension at 72°C for 10 min. Relative
standard curves were applied for relative quantification. Details
of primers and PCR conditions are listed in Table I. The relative expression of all the
target genes, including CRH, AVP, glutamic acid decarboxylase 65
(GAD65), vesicular glutamate transporter 2 (vGLUT2), GR and MR, was
standardized against β-actin expression levels. The Comparative Cq
Method (2−ΔΔCq Method) was applied to obtain the
relative expression level of the target genes (13).
 | Table I.Primers and conditions for reverse
transcription-quantitative PCR. |
Table I.
Primers and conditions for reverse
transcription-quantitative PCR.
| Genes | Forward
(5′-3′) | Reverse
(5′-3′) | Product (bp) | Annealing |
|---|
| β-actin |
GTTGCCAATAGTGATGACCT |
GGACCTGACAGACTACCTCA | 208 | 54°C, 20 sec |
| CRH |
AGAACAACAGTGCGGGCTCA |
GCTCCGGTTGCAAGAAATTCA | 196 | 60°C, 30 sec |
| AVP |
AAGAGGGCCACATCCGACA |
AGGGCAGGTAGTTCTCCTCCTG | 160 | 58°C, 20 sec |
| GAD65 |
TGCAGCCTTGGGGATCGGAA |
CCCCAAGCAGCATCCACATGCA | 237 | 60°C, 30 sec |
| VGluT2 |
TCCACCGGGGTGGCAAAGTT |
TGCGATGTATCCGCCCGGAA | 128 | 60°C, 30 sec |
| GR |
CACCCATGACCCTGTCAGTC |
AAAGCCTCCCTCTGCTAACC | 156 | 61°C, 30 sec |
| MR |
TGCATGATCTCGTGAGTGA |
AAGTTCTTCCTGGCCGGTAT | 190 | 62°C, 30 sec |
Histological examination
The samples were fixed at 4°C in a 4%
paraformaldehyde solution overnight and processed with the paraffin
section technique. Sections (thickness, 5 µm) were stained with
H&E using the following protocol: Immersing in xylene for 45
sec and then in propanol for another 45 sec, staining with
hematoxylin at 30°C for 45 sec in a thermostatic bath, rinsing with
warm distilled H2O for 45 sec, staining in eosin at 30°C
for 30 sec in a thermostatic bath, rinsing with cold distilled
H2O for 15 sec, dipping in propanol for 45 sec and then
45 sec in xylene. Then, the slices were observed under a light
microscope at original magnifications of ×100 and ×400.
A fetal rat hippocampal tissue sample was randomly
selected from each group and fixed at 4°C in 2.5% glutaral solution
for 2 h and at room temperature in 1% osmium acid solution for 1 h,
successively. Then, the samples were dehydrated in increasing ethyl
alcohol concentrations (in 50% ethyl alcohol for 15 min, 70% ethyl
alcohol for 15 min, 80% ethyl alcohol for 15 min, 95% ethyl alcohol
for 15 min and 100% ethyl alcohol for 15 min), embedded in Epon and
cut on an Ultrotome V ultramicrotome into 50-nm sections (LKB).
Subsequently, sections were stained with 2% (w/v) uranyl acetate
and 2% lead citrate at room temperature for 30 and 5 min,
respectively. The changes in ultrastructure were observed and
imaged using an H-600 transmission electron microscope (Hitachi,
Ltd.) at an original magnification of ×15,000.
Statistical analysis
Data are presented as the mean ± SEM, from n≥8
samples/group. All experiments were repeated at least once. A
Shapiro-Wilk test was applied to test the normality of the data.
Subsequently, unpaired Student's t-test was applied to analyze the
continuous data, including the mRNA expression levels and serum
concentration of hormones, as well as the ratios of vGLUT1/GAD65
and MR/GR, and the rise rate of serum ACTH, serum CORT and sucrose
preference of the rats. Two-way ANOVA and Bonferroni post hoc test
were applied to analyze the differences between the control and PFR
groups, as well as the differences between with and without UCS
groups. Data analysis was performed by using SPSS 17.0 (SPSS, Inc.)
and GraphPad Prism 5.0 software (GraphPad Software, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Adult male offspring rats with PFR and
high-fat diet
HPA axis activity in the rat offspring
with maternal PFR before and after PFR
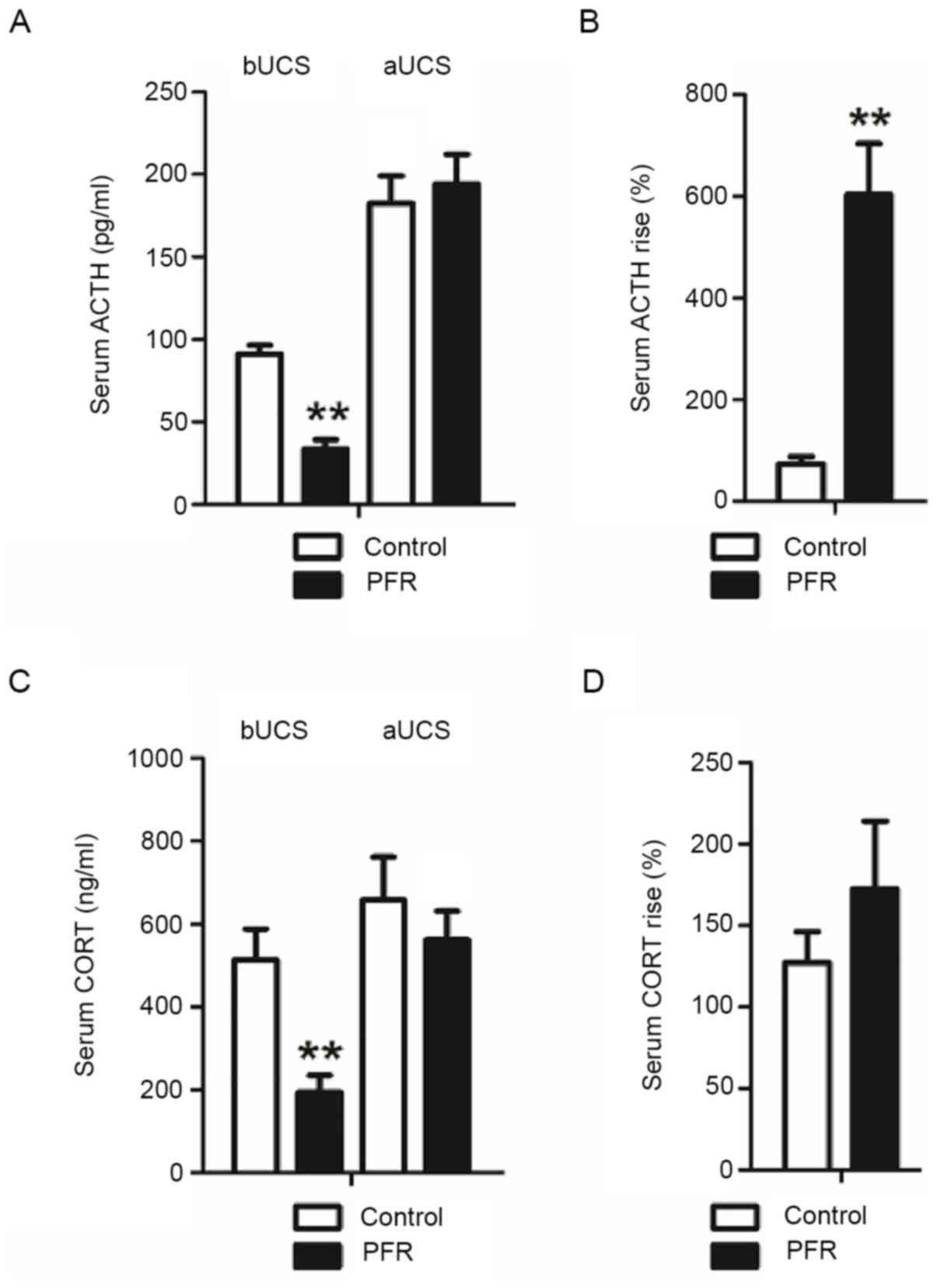
The serum ACTH and CORT concentrations in the adult
rat offspring with maternal PFR were lower compared with those in
the controls before UCS (P<0.01; Fig. 1A and C). However, no notable
differences were observed between the two groups after UCS. In
addition, the rate of the increase of serum ACTH in the PFR group
was significantly higher compared with that of the control
(P<0.01: Fig. 1B), while the
rate of the increase of serum CORT in the PFR group showed no
statistically significant changes after UCS (Fig. 1D).
Expression levels of the key
regulators of the HPA axis in the hypothalamus and hippocampus of
the rat offspring with maternal PFR with or without PFR
vGluT2 and GAD65 are often considered as the markers
of the glutamatergic neurons and γ-aminobutyric acid (GABA)ergic
neuron, respectively, as vGluT2 and GAD65 determine the
accumulation of glutamic acid (Glu) and GABA at the synapses,
respectively, and also are similarly located with the glutamatergic
neurons and GABAergic neurons in the central nervous system
(14). Therefore, the ratio of the
GAD65/vGlut2 was determined to examine the balance of Glu and GABA
accumulation at the synapses. CRH mRNA expression level in the
hypothalamus was significantly lower in the PFR without UCS group
compared with that in controls without UCS (P<0.05; Fig. 2A), while no significant difference
was observed in the AVP, GAD65, and vGLUT2 mRNA expression levels,
as well as the vGLUT2/GAD65 ratio (Fig.
2B-E). However, the AVP, GAD65 and vGLUT2 mRNA expression
levels, as well as the vGLUT2/GAD65 ratio, were all significantly
increased in the PFR group with UCS when compared with those of the
control with UCS (P<0.01; Fig.
2B-E).
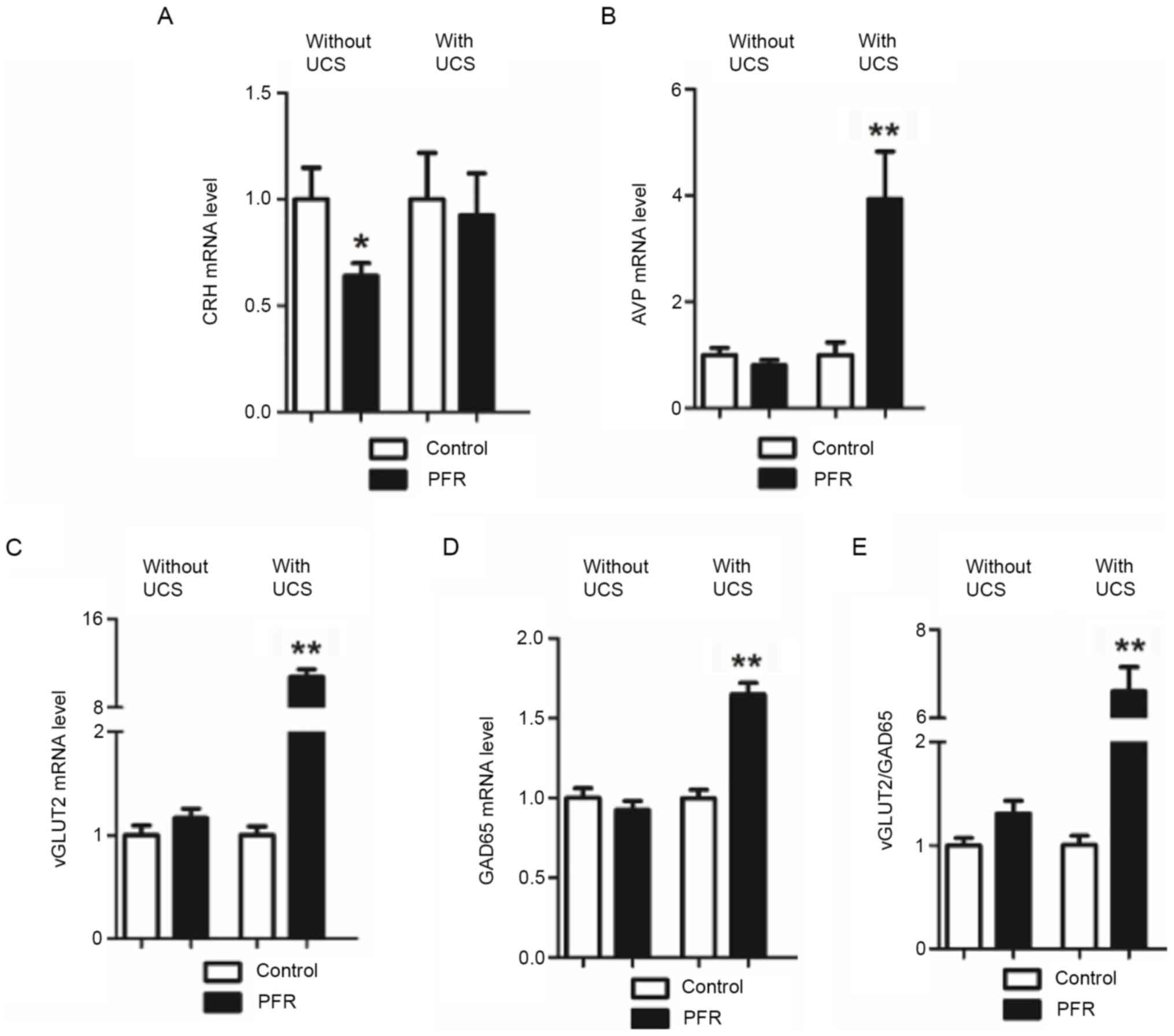 | Figure 2.Effects of PFR on the
hypothalamic-pituitary-adrenal-axis-related gene expression in the
hypothalamus of the adult male rat offspring. Brain samples were
collected from the different set of animals with UCS and without
UCS. Gene expression levels of (A) CRH and (B) AVP in the
hypothalamus of the adult rat offspring bUCS and aUCS. Gene
expression levels of (C) vGLUT2 and (D) GAD65, (E) as well as the
ratio of the vGLUT2 and GAD65 expression levels in the hypothalamus
of the adult rat offspring bUCS and aUCS. Data are presented as the
mean ± SEM, n≥8. *P<0.05, **P<0.01 vs. control. PFR, prenatal
food restriction; CRH, corticotrophin-releasing hormone; AVP,
arginine vasopressin; UCS, unpredictable chronic stress; GAD65,
glutamic acid decarboxylase 65; vGLUT2, vesicular glutamate
transporter 2; bUCS, before UCS; aUCS, after UCS. |
It was found that the MR mRNA expression level in
the hippocampus of the rat offspring with maternal PFR was lower
compared with the control, while the GR mRNA expression level was
higher than the control, when the rats received no UCS (P<0.05;
Fig. 3A and B). However, both the
MR and GR mRNA expression levels in the PFR group were upregulated
when compared with those of the control, when the rats were exposed
to UCS (P<0.05; Fig. 3A and B).
The MR/GR mRNA expression ratio in the PFR group presented a
decreasing trend when compared with the control either with or
without UCS, while an increasing trend was observed in the ratio
with UCS when compared with that without UCS, whereas all these
changes showed no statistical difference (P<0.05; Fig. 3C).
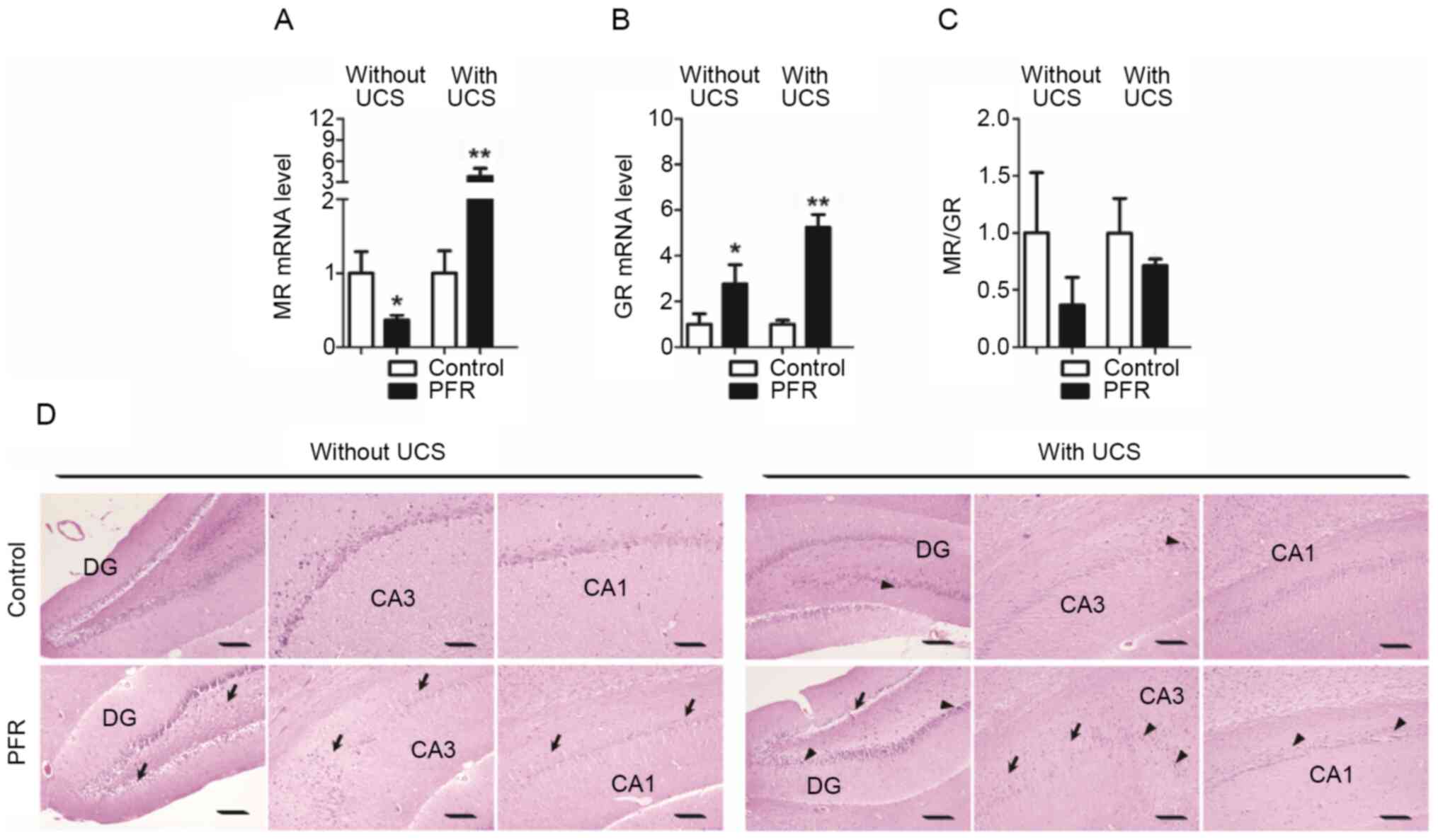 | Figure 3.Effects of PFR on the functional gene
expression and morphology of the hippocampus in the adult male rat
offspring. Brain samples were collected from the different set of
animals with UCS and without UCS. Gene expression levels of (A) MR
and (B) GR, (C) as well as the ratio of the MR/GR bUCS and aUCS.
(D) Histology of the hippocampus was also observed. Arrows indicate
disordered neurons, whereas arrowheads indicate hyperchromatic
nuclei of the neurons. Data are presented as the mean ± SEM, n=8.
*P<0.05, **P<0.01 vs. control. Scale bar, 100 µm. DG, dentate
gyrus; CA, cornu ammonis; PFR, prenatal food restriction; UCS,
unpredictable chronic stress; bUCS, before UCS; aUCS, after
UCS. |
With regards to histology, orderly distributed
neurons in the dentate gyrus (DG), cornu ammonis (CA) 1 area and
CA3 area were observed in the control rats without UCS. However,
the neurons in the DG, CA1 and CA3 areas of the rat offspring with
PFR were found to be disorderly distributed, with hyperchromatic
nuclei in a proportion of the neurons (Fig. 3D). Neurons in the DG, CA1 and CA3
areas were also orderly distributed, with only a few hyperchromatic
nuclei observed in the control rats with UCS (Fig. 3D). Moreover, neurons in the DG, CA1
and CA3 areas of the rat offspring with maternal PFR were further
disorderly distributed, with hyperchromatic nuclei observed in
several neurons, in the PFR rats with UCS (Fig. 3D). Behavioral changes were also
observed in the rat offspring with maternal PFR, and included fewer
crossing times and rearing times before UCS and fewer rearing times
after the stress in the open field test, as well as decreased total
time needed for escape and increased times of correct reaction
after UCS (Fig. S1).
Male rat fetuses with maternal
PFR
Ultrastructure and the key regulators of the HPA
axis in the hypothalamus and hippocampus
To identify the possible origins of the changes in
the adult male rat offspring, the fetuses were further
investigated, including the ultrastructural, the histopathological
and the expressional changes of the key regulators of the HPA axis
in the hypothalamus and hippocampus. Swelling of the mitochondria
and endoplasmic reticulum, as well as the cytoplasmic vacuolation,
were observed in the neurons located in the hypothalamus of the
fetuses with maternal PFR (Fig.
4A). Furthermore, the mRNA expression levels of AVP, GAD65 and
vGLUT2 in the hypothalamus were all lower compared with those in
the controls, while the vGLUT2/GAD65 ratio was significantly higher
in the hypothalamus (P<0.05; Fig.
4B-D).
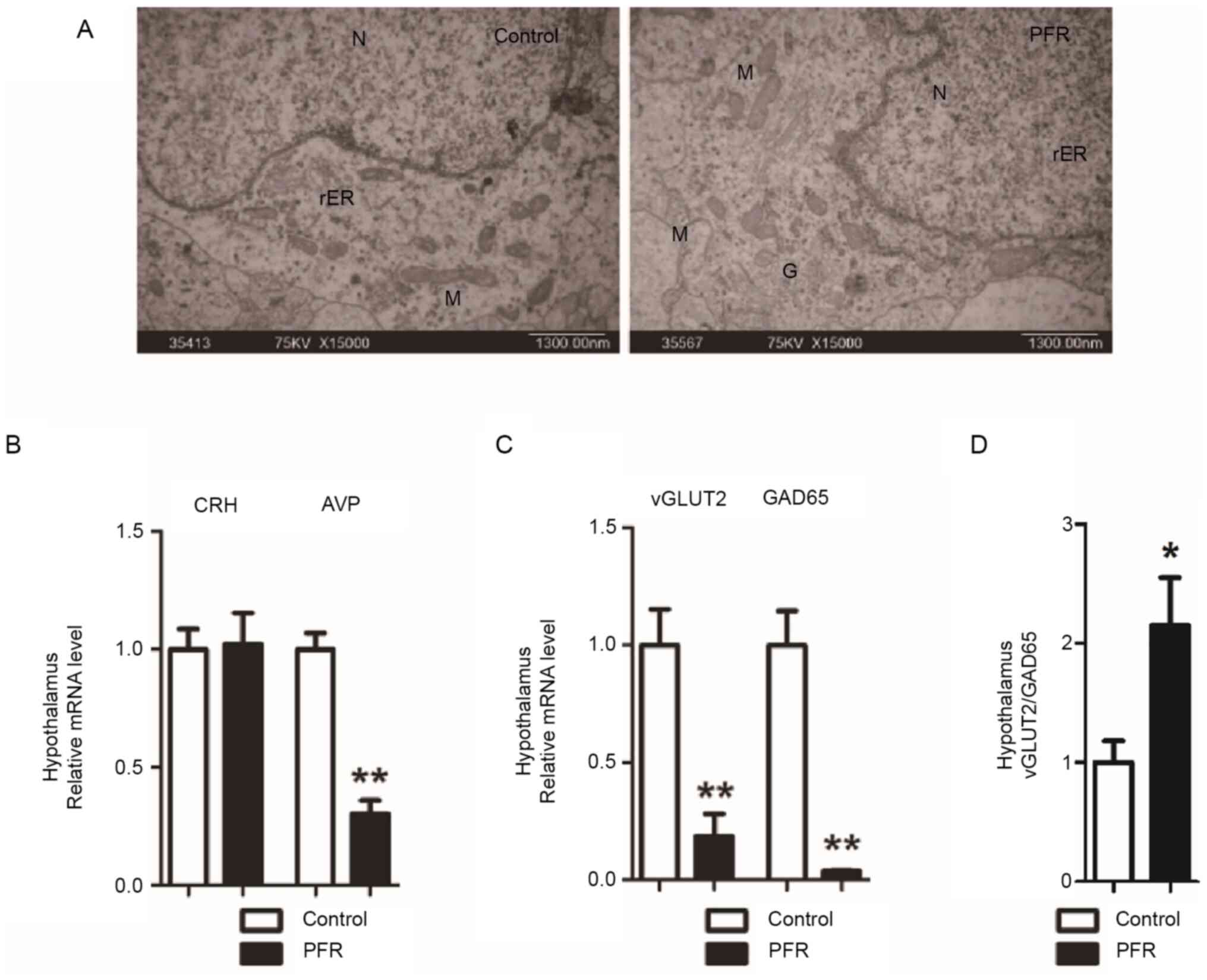 | Figure 4.Effects of PFR on the ultrastructure
of hypothalamus and hypothalamic-pituitary-adrenal-axis-related
gene expression in the hypothalamus of the fetal male rat fetuses.
(A) Ultrastructure changes of the fetal hypothalamus. Scale bar,
1.3 µm. Gene expression levels of (B) CRH and AVP, (C) GAD65 and
vGLUT2, (D) as well as the ratio of the vGLUT2/GAD65 expression
level in the fetal hypothalamus. Data are presented as the mean ±
SEM, n≥8. *P<0.05, **P<0.01 vs. control. N, nucleus; M,
mitochondrion; rER, rough surfaced endoplasmic reticulum; G, Golgi
apparatus; V, vacuole; PFR, prenatal food restriction; GAD65,
glutamic acid decarboxylase 65; vGLUT2, vesicular glutamate
transporter 2; CRH, corticotrophin-releasing hormone; AVP, arginine
vasopressin. |
Changes in the hippocampus were also observed. The
serum CORT concentration was significantly higher in the male PFR
fetuses compared with that in the controls (P<0.05; Fig. 5A). A notable decreasing trend in the
MR expression level was also observed in the male fetuses with
maternal PFR (Fig. 5B). However, no
statistical significance was observed in the mRNA expression of MR
and GR, or the ratio of MR/GR (Fig. 5B
and C). Swelling of the mitochondria and endoplasmic reticulum.
Swelling of the mitochondria and endoplasmic reticulum, cytoplasmic
vacuolation, as well as the aggregation of heterochromatin were
observed in the neurons of the hippocampus from the fetuses with
maternal PCR, which was rarely observed in the controls (Fig. 5D). Furthermore, the cortex of the
hippocampus in the control rats was more mature compared with that
in the PFR rats. Neurons in the DG, CA1 and CA3 areas of the
control rats were orderly distributed, while neurons in the DG, CA1
and CA3 areas of the rats with maternal PFR were more disordered
(Fig. 5E).
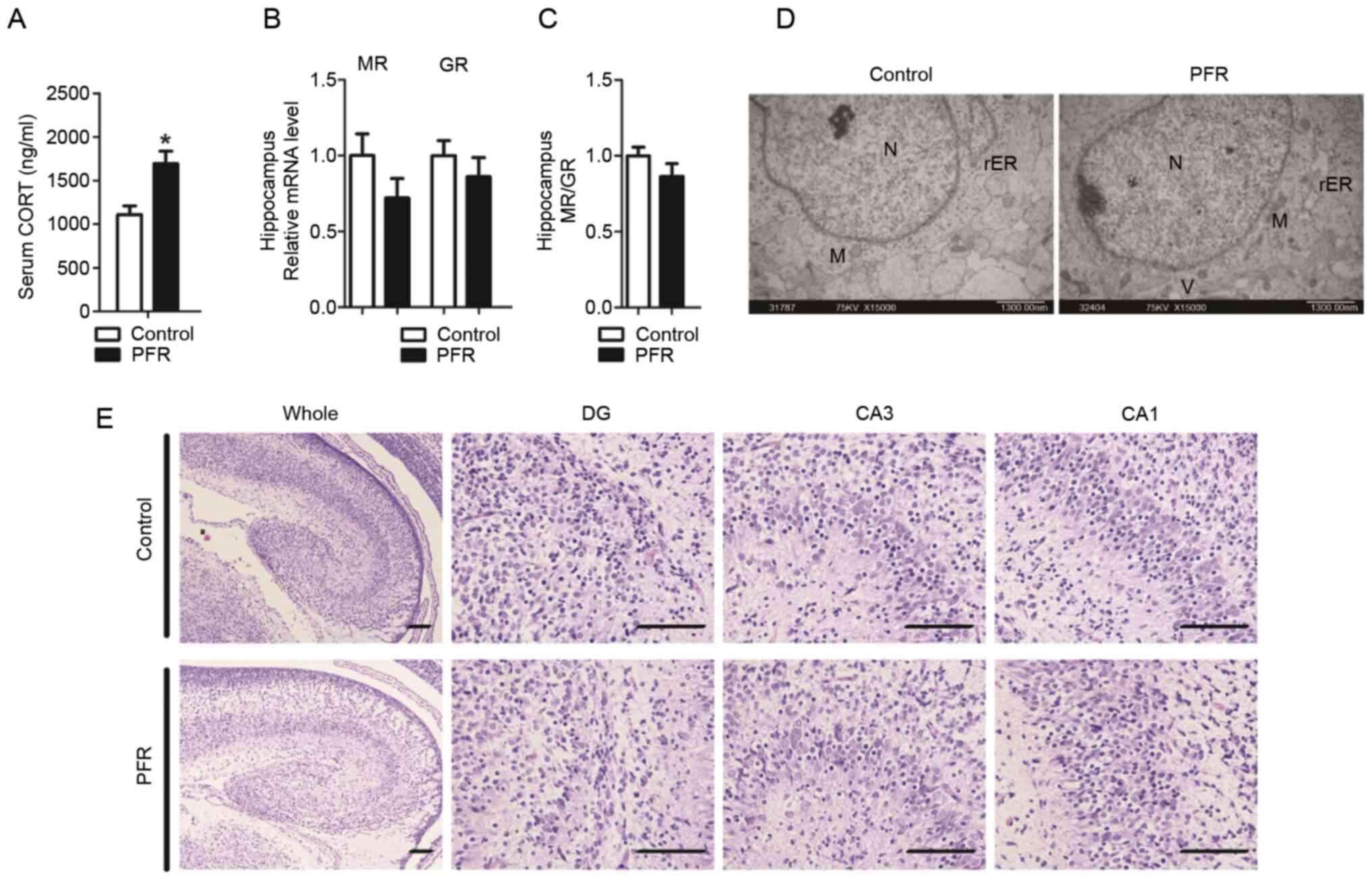 | Figure 5.Effects of PFR on the morphology of
the hippocampus and the expressions of MR and GR in the hippocampus
of the male rat fetuses. (A) Serum CORT concentration, (B) gene
expression levels of MR and GR, (C) as well as the ratio of the
MR/GR in fetal hippocampus. (D) Ultrastructural changes of the
fetal hippocampus. Scale bar, 1.3 µm. (E) Morphological changes of
the hippocampus. Scale bar, 100 µm. Data are presented as the mean
± SEM, n≥8. *P<0.05 vs. control. N, nucleus; M, mitochondrion;
rER, rough surfaced endoplasmic reticulum; G, Golgi apparatus; V,
vacuole; PFR, prenatal food restriction; MR, mineralocorticoid
receptor; GR, glucocorticoid receptor; CORT, corticosterone; DG,
dentate gyrus; CA, cornu ammonis. |
Discussion
Over the past decades, an increasing number of
studies have reported that the programmed alteration of the HPA
axis serves a key role in the development origin of adult diseases,
such as stress and hypertension (2,15–17).
Our previous study revealed that the serum levels of ACTH and CORT
in adult rat offspring with PFR were lower compared with those in
the controls, but were significantly increased after UCS, which was
accompanied by alterations in HPA axis-associated neuroendocrinal
metabolism (6). Such changes
indicated an intrauterine programming of the HPA axis to a low
basic activity but a high susceptibility in the adult offspring.
Furthermore, a suppressed basic function of the HPA axis in the
childhood, but a stimulated sensitivity to stressors during
adulthood in the offspring with maternal PFR, were also reported by
other studies (18–21). In the present study, the programmed
alteration of HPA axis was also identified in the male adult
offspring with maternal PFR. Namely, lower serum concentrations of
ACTH and CORT were observed in the PFR rat without UCS, while
higher serum concentrations of ACTH and CORT were detected in the
PFR rat with UCS, as well as a higher increasing rate of serum
ACTH. Moreover, lower expression levels of CRH mRNA in the
hypothalamus were found in the PFR rats without UCS, when compared
with the controls. However, increases in the serum concentration of
ACTH and the mRNA expression level of AVP were detected in the PFR
rat with UCS, when compared with the controls. These finding
indicated that a low basic activity but a high susceptibility of
the HPA axis was induced in the male adult offspring by maternal
PFR.
Glu and GABA are the key neurotransmitters in the
central nervous system, and the balance of which in the
hypothalamus helps to maintain the normal activity of the HPA axis
(22). vGLUT2 is the key
transporter of Glu at chemical synapses, while GAD65 converts the
transformation of Glu to GABA (23). Thus, vGLUT2 and GAD65 determine the
accumulation of Glu and GABA, respectively, at the synapses, and
are also synchronically located with glutamatergic neurons and
GABAergic neurons in the central nervous system. Furthermore, the
gene expression levels of both vGLUT2 and GAD65 are relatively
stable in the brain. Thus, vGLUT2 and GAD65 are considered as the
markers of the glutamatergic neurons and GABAergic neurons,
respectively, and the ratio of vGLUT2/GAD65 is used to determine
the balance of Glu and GABA accumulation at the synapses (14). The present study demonstrated that
although no significant difference was observed in the vGLUT2 mRNA
expression level, the GAD65 mRNA expression level and the
expression ratio of vGLUT2/GAD65 in the hypothalamus of the PFR
rats without UCS compared with the control, markedly higher vGLUT2
and GAD65 mRNA expression levels, as well as the expression ratio
of vGLUT2/GAD65 were observed in the PFR rats with UCS, when
compared with the control groups. These findings suggested that a
local stimulation of the activity of the hypothalamus may be
induced in the male adult offspring by maternal PFR.
When the body is under stress, the secretion of CRH
and AVP from the hypothalamus is stimulated, which then promotes
the secretion of ACTH from the anterior pituitary. Subsequently,
the ACTH reaches the adrenal gland via the blood circulation, and
stimulates the synthesis and secretion of CORT from the adrenal
cortex (18). However, when the
serum CORT reaches a certain level, the hippocampal GR and MR are
activated, participating in the feedback regulation of the HPA
axis, which helps to maintain the normal function of the HPA axis
(8). As the center regulators of
the HPA axis, GR and MR are abundantly expressed in the
hippocampus. However, the affinity of MR to CORT is much stronger
compared with GR. Thus, almost all CORT binds to MR when the serum
CORT level is normal (24). When
the serum CORT level increases, particularly under stress, the MR
in the hippocampus gets overloaded, and then the GR is activated
(25). The activation of GR in the
hippocampus induces the secretion of Glu, and successively
activates GABAergic neurons via the Glu-GABA interaction, thereby
suppressing the function of the hypothalamic CRH neurons and
avoiding the overactivation of the HPA axis (26–28).
Thus, the balance of MR and GR in the hippocampus is critical for
maintain the function of the HPA axis, which may also determine the
local activity of the hypothalamus (29). In addition, the overactivation of GR
may induce the influx of Ca2+, which further induces the
degeneration and apoptosis of hippocampal neurons, and primarily
the cone neurons in the CA3 area and the granular cells in the DG
area are the most common victims (30–33).
In the present study, increased GR expression,
suppressed MR expression and decreased MR/GR ratio, as well as
histopathological changes including decreased number and disordered
distribution of neurons in the DG, CA1 and CA3 areas, were observed
in the hippocampus of adult male offspring with maternal PFR
without UCS. However, immunostaining of the bio-markers of neurons
is required to further quantify the cell numbers of the neurons and
to evaluate the effect of PFR on the structure of the hippocampus,
which is one of the limitations of the current work. The present
study demonstrated the mRNA expression levels of both MR and GR
were stimulated in the PFR rats with UCS, when compared with the
control. Moreover, altered behaviors were observed, including
reduced activity in the open field test and poor performance in the
maze experiment before and after UCS. All these changes indicated
that maternal PFR may programmed a stimulated feedback regulation
of hippocampus on the HPA axis at the baseline by activating GR
instead of MR in the hypothalamus, which resulted in low basic
activity of HPA axis. However, when the animal was subjected to
UCS, the programmed feedback regulation from the hippocampus cannot
compensate for the overactivation of the HPA axis due to the
stimulated local activity of the hypothalamus, leading to the high
sensitivity of the HPA axis in the male adult offspring.
Glucocorticoids are key hormones that modulate the
growth and maturation of the fetus, and also participate in the
occurrence of IUGR (34,35). It has been reported that multiple
adverse factors during pregnancy, including maternal prenatal food
restriction, smoking and alcohol consumption, may induce IUGR, and
increased serum glucocorticoids levels are observed in those
fetuses with adverse factors during pregnancy (36–40).
Our previous studies also revealed that the increased serum
glucocorticoid level originated from the maternal blood, which
could suppress the development of the HPA axis, thereby causing
IUGR in the fetuses (39,41). In the present study, increased serum
CORT concentration was observed in the IUGR fetuses induced by PFR,
along with decreased AVP, GAD65 and vGLUT2 mRNA expression levels,
and an increased vGLUT2/GAD65 ratio. Such changes were similar to
the data from the adult male offspring, which indicated that the
stimulated potential excitability of the hypothalamus may originate
from the fetus. Furthermore, histopathological changes of the
hippocampus and hypothalamus were identified in the fetus, which
may form the functional changes of the HPA axis. Combined with the
evidence from the adult rats, it was suggested that the excessive
CORT induced by PFR suppressed the development of both the
hippocampus and hypothalamus, but created a stimulated potential
excitability of the hypothalamus and a poor feedback regulation
from the hippocampus in the fetus, which persisted to the adulthood
and manifested as a low basic activity but a high susceptibility in
the male adult offspring.
In summary, PFR induced an intrauterine programming
of the HPA axis with a low basic activity but a high susceptibility
in the male adult offspring under UCS, which appears to have
originated from the stimulated potential excitability of the
hypothalamus and disturbed feedback regulation from the hippocampus
in the fetus.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant nos. 81430089, 81371483 and
81603214).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
Conceptualization: YXW, SYC, DX and HW. Methodology:
YXW, SYC, CZ, JL, XH and ZXJ. Software: YXW and SYC. Validation:
YXW, SYC, DX and HW. Formal Analysis: YXW, SYC, DX and HW.
Investigation: YXW, SYC, CZ, JL, XH and ZXJ. Data curation: YXW and
SYC. Original draft preparation: YXW. Review and editing: DX and
HW. Visualization: YXW. Supervision: DX and HW. Project
administration: YXW, DX and HW. Funding acquisition: YXW, DX and
HW. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All the protocols were approved by Medical Ethics
Committee of the Basic Medical School of Wuhan University (approval
no. 201719).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang C, Xu D, Luo H, Lu J, Liu L, Ping J
and Wang H: Prenatal xenobiotic exposure and intrauterine
hypothalamus-pituitary-adrenal axis programming alteration.
Toxicology. 325:74–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barker DJ: The developmental origins of
chronic adult disease. Acta Paediatr Suppl. 93:26–33. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Clark PM: Programming of the
hypothalamo-pituitary-adrenal axis and the fetal origins of adult
disease hypothesis. Eur J Pediatr. 157 (Suppl 1):S7–S10. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lesage J, Sebaai N, Leonhardt M,
Dutriez-Casteloot I, Breton C, Deloof S and Vieau D: Perinatal
maternal undernutrition programs the offspring
hypothalamo-pituitary-adrenal (HPA) axis. Stress. 9:183–198. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Poore KR and Fowden AL: The effect of
birth weight on hypothalamo-pituitary-adrenal axis function in
juvenile and adult pigs. J Physiol. 547((Pt 1)): 107–116. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang L, Xu D, Zhang B, Liu Y, Chu F, Guo
Y, Gong J, Zheng X, Chen L and Wang H: Prenatal food restriction
induces a hypothalamic-pituitary-adrenocortical axis-associated
neuroendocrine metabolic programmed alteration in adult offspring
rats. Arch Med Res. 44:335–345. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Holsboer F and Ising M: Stress hormone
regulation: Biological role and translation into therapy. Annu Rev
Psychol. 61:81–109. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tasker JG and Herman JP: Mechanisms of
rapid glucocorticoid feedback inhibition of the
hypothalamic-pituitary-adrenal axis. Stress. 14:398–406. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
National Research Council (US), .
Committee for the Update of the Guide for the Care Use of
Laboratory Animals: Guide for the Care and Use of Laboratory
Animals. (8th edition). National Academies Press. (Washington, DC).
2011.
|
|
10
|
Brenes Sáenz JC, Villagra OR and
Fornaguera Trías J: Factor analysis of forced swimming test,
sucrose preference test and open field test on enriched, social and
isolated reared rats. Behav Brain Res. 169:57–65. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deacon RM and Rawlins JN: T-maze
alternation in the rodent. Nat Protoc. 1:7–12. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ye J, McGinnis S and Madden TL: BLAST:
Improvements for better sequence analysis. Nucleic Acids Res 34
(Web Server Issue). W6–W9. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Flak JN, Ostrander MM, Tasker JG and
Herman JP: Chronic stress-induced neurotransmitter plasticity in
the PVN. J Comp Neurol. 517:156–165. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smith SM and Vale WW: The role of the
hypothalamic-pituitary-adrenal axis in neuroendocrine responses to
stress. Dialogues Clin Neurosci. 8:383–395. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sebaai N, Lesage J, Breton C, Vieau D and
Deloof S: Perinatal food deprivation induces marked alterations of
the hypothalamo-pituitary-adrenal axis in 8-month-old male rats
both under basal conditions and after a dehydration period.
Neuroendocrinology. 79:163–173. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hawkins P, Hanson MA and Matthews SG:
Maternal undernutrition in early gestation alters molecular
regulation of the hypothalamic-pituitary-adrenal axis in the ovine
fetus. J Neuroendocrinology. 13:855–861. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vieau D, Sebaai N, Leonhardt M,
Dutriez-Casteloot I, Molendi-Coste O, Laborie C, Breton C, Deloof S
and Lesage J: HPA axis programming by maternal undernutrition in
the male rat offspring. Psychoneuroendocrinology. 32 (Suppl
1):S16–S20. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Leonhardt M, Lesage J, Croix D,
Dutriez-Casteloot I, Beauvillain JC and Dupouy JP: Effects of
perinatal maternal food restriction on pituitary-gonadal axis and
plasma leptin level in rat pup at birth and weaning and on timing
of puberty. Boil Reprod. 68:390–400. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hawkins P, Steyn C, McGarrigle HH, Saito
T, Ozaki T, Stratford LL, Noakes DE and Hanson MA: Effect of
maternal nutrient restriction in early gestation on responses of
the hypothalamic-pituitary-adrenal axis to acute isocapnic
hypoxaemia in late gestation fetal sheep. Exp Physiol. 85:85–96.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tegethoff M, Pryce C and Meinlschmidt G:
Effects of intrauterine exposure to synthetic glucocorticoids on
fetal, newborn, and infant hypothalamic-pituitary-adrenal axis
function in humans: A systematic review. Endocr Rev. 30:753–789.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gohlke JM, Griffith WC and Faustman EM: A
systems-based computational model for dose-response comparisons of
two mode of action hypotheses for ethanol-induced
neurodevelopmental toxicity. Toxicol Sci. 86:470–484. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Herman JP, Mueller NK and Figueiredo H:
Role of GABA and glutamate circuitry in
hypothalamo-pituitary-adrenocortical stress integration. Ann N Y
Acad Sci. 1018:35–45. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
de Kloet ER, Meijer OC, de Nicola AF, de
Rijk RH and Joëls M: Importance of the brain corticosteroid
receptor balance in metaplasticity, cognitive performance and
neuro-inflammation. Front Neuroendocrinol. 49:124–145. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kozlovsky N, Matar MA, Kaplan Z, Zohar J
and Cohen H: A distinct pattern of intracellular
glucocorticoid-related responses is associated with extreme
behavioral response to stress in an animal model of post-traumatic
stress disorder. Eur Neuropsychopharmacol. 19:759–771. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Treccani G, Musazzi L, Perego C, Milanese
M, Nava N, Bonifacino T, Lamanna J, Malgaroli A, Drago F, Racagni
G, et al: Stress and corticosterone increase the readily releasable
pool of glutamate vesicles in synaptic terminals of prefrontal and
frontal cortex. Mol Psychiatry. 19:433–443. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Matthews SG: Early programming of the
hypothalamo-pituitary-adrenal axis. Trends Endocrinol Metab.
13:373–380. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sapolsky RM and Meaney MJ: Maturation of
the adrenocortical stress response: Neuroendocrine control
mechanisms and the stress hyporesponsive period. Brain Res.
396:64–76. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Krishnamurthy S, Garabadu D and Joy KP:
Risperidone ameliorates post-traumatic stress disorder-like
symptoms in modified stress re-stress model. Neuroendocrinology.
75:62–77. 2013.
|
|
30
|
Liao XM, Yang XD, Jia J, Li JT, Xie XM, Su
YA, Schmidt MV, Si TM and Wang XD: Blockade of
corticotropin-releasing hormone receptor 1 attenuates early-life
stress-induced synaptic abnormalities in the neonatal hippocampus.
Hippocampus. 24:528–540. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
de Quervain DJ, Aerni A, Schelling G and
Roozendaal B: Glucocorticoids and the regulation of memory in
health and disease. Front Neuroendocrinol. 30:358–370. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Arbel I, Kadar T, Silbermann M and Levy A:
The effects of long-term corticosterone administration on
hippocampal morphology and cognitive performance of middle-aged
rats. Brain Res. 657:227–235. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
De Kloet ER, Vreugdenhil E, Oitzl MS and
Joëls M: Brain corticosteroid receptor balance in health and
disease. Endocr Rev. 19:269–301. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Arends N, Johnston L, Hokken-Koelega A,
van Duijn C, de Ridder M, Savage M and Clark A: Polymorphism in the
IGF-I gene: Clinical relevance for short children born small for
gestational age (SGA). J Clin Endocrinol Metab. 87:27202002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qiu XS, Huang TT, Deng HY, Shen ZY, Ke ZY,
Mei KY and Lai F: Effects of early nutrition intervention on IGF1,
IGFBP3, intestinal development, and catch-up growth of intrauterine
growth retardation rats. Chin Med Sci J. 19:189–192.
2004.PubMed/NCBI
|
|
36
|
Thamotharan M, Shin BC, Suddirikku DT,
Thamotharan S, Garg M and Devaskar SU: GLUT4 expression and
subcellular localization in the intrauterine growth-restricted
adult rat female offspring. Am J Physiol Endocrinol Metab.
288:E935–E947. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sloboda DM, Moss TJ, Li S, Matthews SG,
Challis JR and Newnham JP: Expression of glucocorticoid receptor,
mineralocorticoid receptor, and 11beta-hydroxysteroid dehydrogenase
1 and 2 in the fetal and postnatal ovine hippocampus: Ontogeny and
effects of prenatal glucocorticoid exposure. J Endocrinol.
197:213–220. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tan Y, Liu J, Deng Y, Cao H, Xu D, Cu F,
Lei Y, Magdalou J, Wu M, Chen L and Wang H: Caffeine-induced fetal
rat over-exposure to maternal glucocorticoid and histone
methylation of liver IGF-1 might cause skeletal growth retardation.
Toxicol Lett. 214:279–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu D, Chen M, Pan XL, Xia LP and Wang H:
Dexamethasone induces fetal developmental toxicity through
affecting the placental glucocorticoid barrier and depressing fetal
adrenal function. Environ Toxicol Pharmacol. 32:356–363. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen M, Wang T, Liao ZX, Pan XL, Feng YH
and Wang H: Nicotine-induced prenatal overexposure to maternal
glucocorticoid and intrauterine growth retardation in rat. Exp
Toxicol Pathol. 59:245–251. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu D, Zhang B, Liang G, Ping J, Kou H, Li
X, Xiong J, Hu D, Chen L, Magdalou J and Wang H: Caffeine-induced
activated glucocorticoid metabolism in the hippocampus causes
hypothalamic-pituitary-adrenal axis inhibition in fetal rats. PLoS
One. 7:e444972012. View Article : Google Scholar : PubMed/NCBI
|