Introduction
Ischemic heart disease is the main cause of
cardiovascular disease, which could induce myocardial infarction
and lead to myocardial necrosis, impairing the quality of life of
patients (1,2). Blood flow recovery is the most
effective way to save ischemic cardiomyocytes and the lives of
patients (1). However, myocardial
ischemia-reperfusion (I/R) can lead to a series of adverse events,
including excessive reactive oxygen species, calcium ion overload
and endoplasmic reticulum stress, which could promote cardiomyocyte
apoptosis and aggravate the degree of myocardial injury (3). Thus, there is an urgent need to
explore the potential molecular mechanism of myocardial I/R injury
to improve the quality of life of patients.
Long non-coding RNAs (lncRNAs) are a type of RNA
transcripts without protein coding ability and are longer than 200
nucleotides (4). Numerous studies
have demonstrated that lncRNAs are involved in numerous important
processes in cell biology, including signal transduction
regulation, DNA modification, transcriptional activation and
protein function regulation (5). In
addition, lncRNAs serve an important role in the occurrence and
development of tumor cells (6).
Small nucleolar RNA host gene 1 (SNHG1) is a lncRNA located on
chromosome 11p2.3 (7). SNHG1 has a
protective effect on cardiomyocytes under various impairments. For
example, SNHG1 has been found to protect cardiomyocytes against
toxic damage induced by Adriamycin (8) and inhibit cardiomyocyte apoptosis
induced by H2O2 (9). A recent study revealed that SNHG1
could reduce vascular endothelial cell injury induced by
hypoxia/reoxygenation (H/R) (10).
In addition, SNHG1 acts as a competing endogenous RNA of microRNAs
(miRNAs/miRs) to suppress the regulation of target genes (11).
Therefore, it was hypothesized that SNHG1 may have a
protective effect on cardiomyocyte injury induced by H/R by
sponging miRNA. However, to the best of our knowledge, there are no
studies on the exact role and mechanism of SNHG1 in cardiomyocyte
injury induced by H/R. Thus, the purpose of the present study was
to explore the role and molecular mechanism of SNHG1 in ischemic
cardiomyocyte injury.
Materials and methods
Cell culture and treatment
H9c2 cells were obtained from BeNa Culture
Collection; Beijing Beina Chunglian Institute of Biotechnology
Research Institute. The cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; HyClone; Cytiva) containing 10% fetal bovine
serum (Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin. H9c2 cells were routinely cultured for 3
days and then subcultured. Cells in the logarithmic phase were used
for subsequent experiments.
H9c2 cells were treated with H/R to
establish the H/R model
Cells in the logarithmic phase were inoculated into
24-well plates at a density of 2×104 cells/well. When
the cell fusion degree reached 90%, the culture medium was replaced
with serum-free medium. Subsequently, cells were cultured in an
incubator containing 90% N2 and 5% CO2 for 6
h at 37°C. Next, cells were returned to the normal culture
conditions. Cells were then cultured in complete DMEM and
maintained in a 5% CO2 incubator for 6, 12 and 24 h at
37°C.
Cell transfection
Short hairpin RNA (shRNA) against GATA4
(shRNA-GATA4; 5′-CTGGATGTTGGGCAGGAC −3′) (100 nM), shRNA negative
control (shRNA-NC) (100 nM), miR-16-5p mimic
(5′-UAGCAGCACGUAAAUAUUGGCG-3′) (50 nM), mimic negative control
(miR-NC; 5′-UUCUCCGAACGUGUCACGUTT −3′, 100 nM), miR-16-5p inhibitor
(5′-CGCCAAUAUUUACGUGCUGCUA −3′, 50 nM), inhibitor negative control
(NC inhibitor; 5′-CAGUACUUUUGUGUAGUACAA −3′, 50 nM) pcDNA-GATA4
(100 nM) and empty control vector (pcDNA-NC; 100 nM) were
synthesized by Shanghai GenePharma Co., Ltd. The SNHG1
overexpression vector (pcDNA-SNHG1; 100 nM) and empty control
vector (pcDNA-NC; 100 nM) were constructed by Shanghai GenePharma
Co., Ltd. Cells were seeded into 6-well plates at a density of
3×105 cells/well and cultured for 24 h at 37°C.
In strict accordance with the instruction of
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) these aforementioned indicated plasmids were
transfected into the cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). Transfection
efficiency was detected via reverse transcription-quantitative PCR
(RT-qPCR) at 48 h after transfection.
RT-qPCR analysis
Total RNA was extracted from cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.). RNA
was synthesized into cDNA using a reverse transcription kit
(Promega Corporation) according to the manufacturer's instructions.
The PCR reaction was performed using SYBR Green Supermix (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and an ABI 7500 PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
PCR reaction conditions were as follows: 94°C for 30 sec, 55°C for
30 sec and 72°C for 30 sec (22 cycles). The relative expression
levels of target genes were calculated using the 2−ΔΔCq
method (12) and normalized to
those of the housekeeping gene GAPDH or U6. Primers used in this
study are listed in Table I.
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Sequences
(5′→3′) |
|---|
| SNHG1 | F:
AGGCTGAAGTTACAGGTC |
|
| R:
TTGGCTCCCAGTGTCTTA |
| miR-16-5p | F:
TAGCAGCACGTAAATATTGGCG |
|
| R:
TGCGTGTCGTGGAGTC |
| GATA4 | F:
GAGCTGGTACCTGGCCTTC |
|
| R:
GCTCTGCTGAAATCACTCTGA |
| GAPDH | F:
GTCAACGGATTTGGTCTGTATT |
|
| R:
AGTCTTCTGGGTGGCAGTGAT |
| U6 | F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
| GAPDH | F:
ATCTCCTTTGTTACCGCTTCC |
|
| R:
GAAGATGGTGATGGGATTTC |
Cell Counting Kit-8 (CCK-8) assay
Cell viability was detected using a CCK-8 assay.
Cells were incubated in 96-well plates at a density of
2×103 cells/well for 24, 48 and 72 h. A total of 10 µl
CCK-8 solution (cat. no. C0037; Beyotime Institute of
Biotechnology) was added to each well. Then, cells were incubated
for 1 h at 37°C. The culture was continued for an additional 2 h.
The absorbance of each well was measured at a wavelength of 450 nm
using a microplate reader (Synergy 2 Multi-Mode Microplate Reader;
BioTek Instruments, Inc.).
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labeling
(TUNEL) assay
Apoptosis was detected using the TUNEL Apoptosis
Assay kit (cat. no. C1088; Beyotime Institute of Biotechnology).
Briefly, cells were washed with PBS and fixed with 4%
paraformaldehyde at room temperature for 20 min. Subsequently,
cells were treated with 0.1% Triton-X-100 for 10 min. Each sample
was supplemented with 50 µl TUNEL detection reagent for 40 min at
37°C in the dark. The cell nuclei were stained with 5 µg/ml DAPI at
37°C in the dark for 5 min. The morphological changes of apoptotic
cells were observed in three random fields under a fluorescence
microscope (Olympus FV500; Olympus Corporation; magnification
×200). Bright green fluorescence was considered to indicate
apoptotic cells.
Enzyme-linked immunosorbent assay
(ELISA)
Cell supernatant was collected and centrifuged at
700 × g at 4°C for 10 min. Then centrifugation was performed at
9,000 × g at 4°C for another 15 min and the supernatant collected.
According to the manufacturer's instructions, an ELISA kit (cat.
no. A020-1-2; Nanjing Jiancheng Bioengineering Institute) was used
to analyze the level of lactate dehydrogenase (LDH) release in the
supernatant. The samples of each group were analyzed using an
Automatic Microplate Reader (Syngene). All the experiments in this
study were repeated three times.
Luciferase reporter assay
Targets of SNHG1 were predicted using the
Encyclopedia of RNA Interactomes (ENCORI) database (https://starbase.sysu.edu.cn/). The cells were
inoculated into 96-well plates at a density of 2×104
cells per well. When the degree of cell fusion reached 60%, the
cells were co-transfected with SNHG1 3′-untranslated region
(3′-UTR) plasmid (wild-type; SNHG1 WT), SNHG11 3′-UTR plasmid
(mutant; SNHG1 MUT), GATA4 3′-UTR plasmid (wild-type; GATA4 WT),
GATA4 3′-UTR plasmid (mutant; GATA4 MUT) was cloned into psi-CHECK
(Promega Corporation) downstream of the firefly luciferase 3′-UTR.
These plasmids were amplified by Shanghai GenePharma Co., Ltd. The
psi-CHECK vector also provided Renilla luciferase as the
normalization signal and miR-16-5p mimics or miR-NC using
Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). At 48 h after transfection, the relative luciferase
intensity of each group was detected using a Dual Luciferase
Reporter Assay kit (Promega Corporation). Firefly luciferase
activities were normalized to Renilla luciferase
activities.
Chromatin immunoprecipitation (ChIP)
assay
The ChIP assay was performed using a ChIP assay kit
(Pierce™ Agarose ChIP Kit; Pierce; Thermo Fisher Scientific, Inc.).
Briefly, the chromatin fragments derived from H9c2 cells were
immunoprecipitated with 10 μg antibody against GATA4 (cat.
no. ab256782; Abcam) or 5 μg mouse IgG (cat. no. sc-2025;
Santa Cruz Biotechnology, Inc.) were used for the
immunoprecipitation. The cell lysate was incubated with a GATA
antibody (1:1000) or IgG antibody (0.2 µg/ml) and 40 μl
protein A/G magnetic beads (EMD Millipore) at 4°C overnight. On the
second day, products of immunoprecipitation were treated with ChIP
elution buffer provided by the kit. Immunoprecipitated complexes
were collected by centrifugation at 10,000 × g for 10 min at 4°C.
Subsequently, precipitated DNA samples were detected via
RT-qPCR.
Western blot analysis
Total RNA was extracted from cells using
radioimmunoprecipitation assay buffer (cat. no. P0013B; Beyotime
Institute of Biotechnology). The protein concentration was detected
using a bicinchoninic acid protein quantitative kit (cat. no.
P0012; Beyotime Institute of Biotechnology). A total of 20
µg protein was separated via 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, and subsequently
transferred to a polyvinylidene fluoride membrane (cat. no. FFP36;
Beyotime Institute of Biotechnology). These membranes were blocked
with 5% skimmed milk for 30 min at room temperature and incubated
with primary antibodies (1:1,000) against GATA4 (cat. no. ab256782;
Abcam), Bcl-2 (cat. no. ab182858; Abcam), Bax (cat. no. ab182733;
Abcam), caspase-3 (cat. no. ab32150; Abcam), Cleaved caspase-3
(cat. no. ab2302; Abcam), caspase-9 (cat. no. ab32539; Abcam),
Cleaved caspase-9 (cat. no. ab2324; Abcam) and GAPDH (cat. no.
ab9485; Abcam) overnight at 4°C. Subsequently, the membranes
were incubated with the corresponding horseradish
peroxidase-conjugated Goat Anti-Rabbit IgG H&L (HRP) secondary
antibody (1:10,000; cat. no. ab205718; Abcam) at 37°C for 2
h at the appropriate dilutions. An ECL Plus kit (cat. no. P0018;
Beyotime Institute of Biotechnology) was used to visualize the
protein bands. Densitometric analysis was performed using ImageJ
software (version 1.49v; National Institutes of Health).
Statistical analysis
All data were analyzed using GraphPad Prism 7
software (GraphPad Software, Inc.). The measurement data are
presented as the mean ± standard deviation. Student's t-test
(unpaired) was used to compare the differences between two groups.
Comparisons among multiple groups were performed using one-way
ANOVA followed by Tukey's post hoc test. P<0.05 was considered
to indicate a statistically significant difference. All experiments
were repeated at least three times.
Results
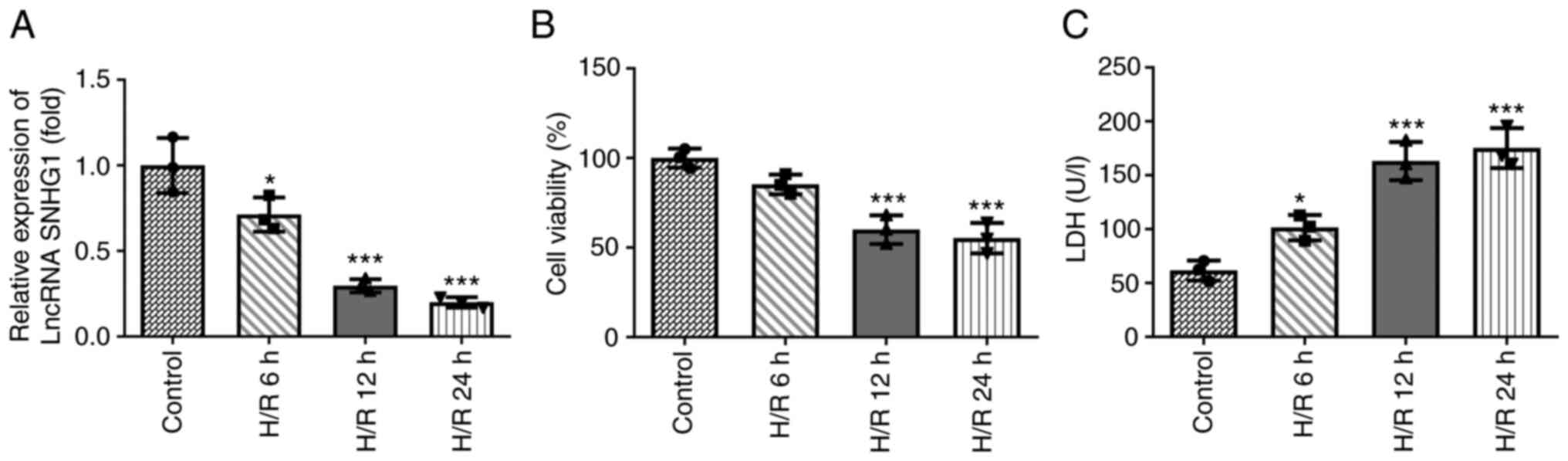
SNHG1 expression is downregulated in
H9c2 cells treated with H/R
The expression levels of SNHG1 in H9c2 cells treated
with H/R were assessed using RT-qPCR. As shown in Fig. 1A, the expression levels of SNHG1 in
H9c2 cells treated with H/R decreased gradually with increasing
treatment time. In addition, cell viability was detected using
CCK-8 and LDH assays. As shown in Fig.
1B and C, compared with the control group, the viability of
H9c2 cells was decreased, while the levels of LDH were increased
with increasing treatment time. When the treatment time was 12 h,
the change in cell viability tended to be stable. Therefore, in the
follow-up experiments, the treatment time of H/R was set as 12
h.
Overexpression of SNHG1 attenuates the
injury of H9c2 cells induced by H/R
The expression levels of SNHG1 in H9c2 cells treated
with H/R were increased by transfection with pcDNA-SNHG1. As shown
in Fig. 2A, the expression levels
of SNHG1 in the pcDNA-SNHG1 group were markedly higher than those
in the control group. As shown in Fig.
2B and C, overexpression of SNHG1 partly alleviated the
inhibition of H9c2 cell viability by H/R. In addition, a TUNEL
assay and western blotting were used to detect cell apoptosis. As
shown in Fig. 2D and E, compared
with the control group, H/R markedly increased the number of
positive apoptotic cells, decreased the expression levels of
anti-apoptotic protein Bcl-2, and increased the expression levels
of pro-apoptotic proteins Bax, Cleaved caspase-3 and Cleaved
caspase-9. These aforementioned changes could be partially reversed
by overexpression of SNHG1.
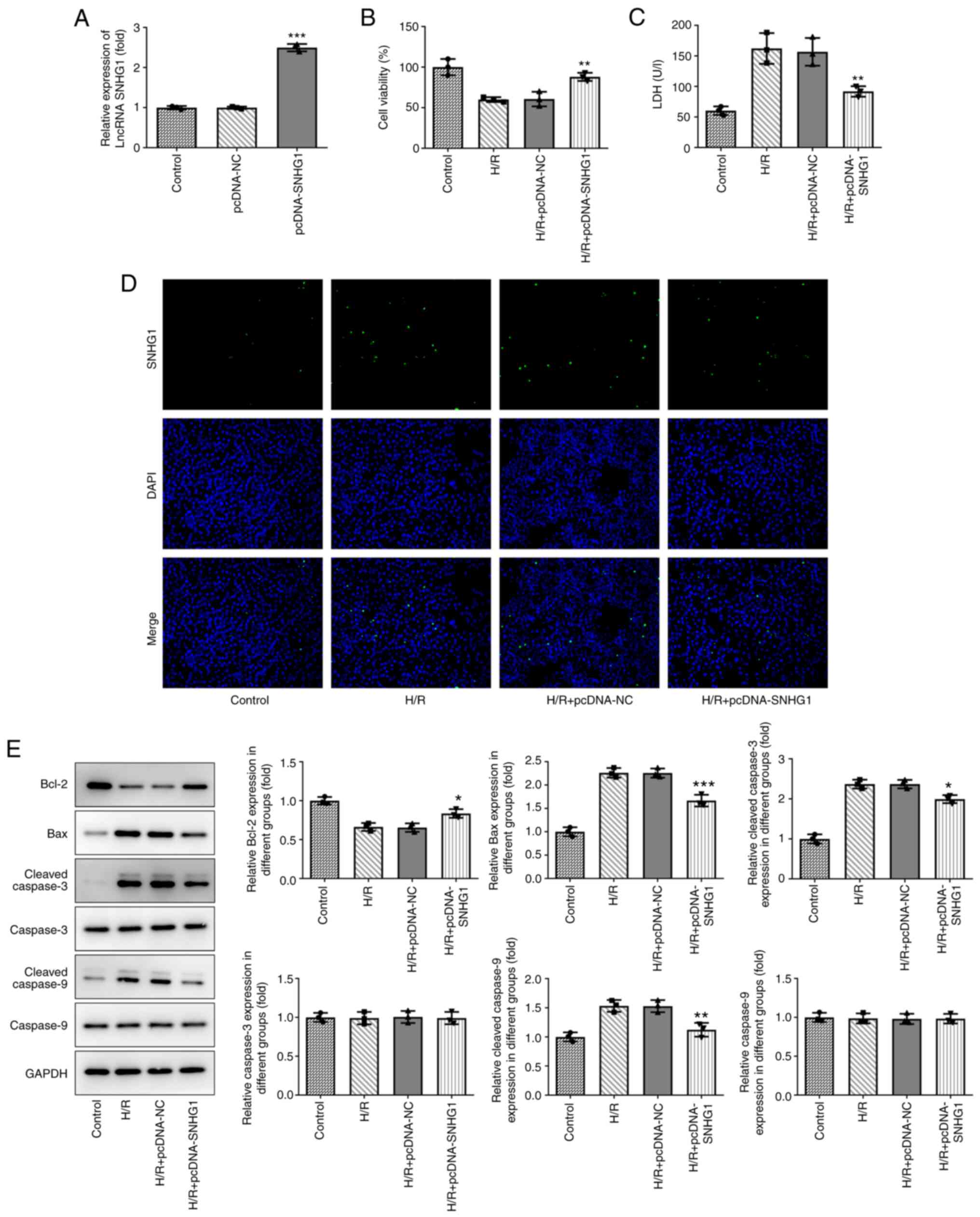 | Figure 2.Overexpression of SNHG1 attenuates
H/R-induced H9c2 cell injury. (A) The efficiency of SNHG1
overexpression was examined via reverse transcription-quantitative
PCR in transfected H9c2 cell lines. ***P<0.001 vs. pcDNA-NC. (B)
Cell proliferation was determined using a Cell Counting Kit-8
assay. **P<0.01 vs. H/R + pcDNA-NC. (C) Cytotoxicity to the
cells was measured via ELISA. **P<0.01 vs. H/R + pcDNA-NC. (D)
Cell apoptosis was detected via a TUNEL assay. Bright green
fluorescence was considered to indicate apoptotic cells
(magnification ×200). (E) Expression of apoptosis-related proteins,
cell proliferation-related proteins (Bcl-2, Bax, Cleaved caspase-3,
caspase-3, Cleaved caspase-9 and caspase-9) were detected by
western blotting. *P<0.05, **P<0.01 and ***P<0.001 vs. H/R
+ pcDNA-NC. SNHG1, small nucleolar RNA host gene 1; H/R,
hypoxia/reoxygenation; LDH, lactate dehydrogenase; lncRNA, long
non-coding RNA; NC, negative control. |
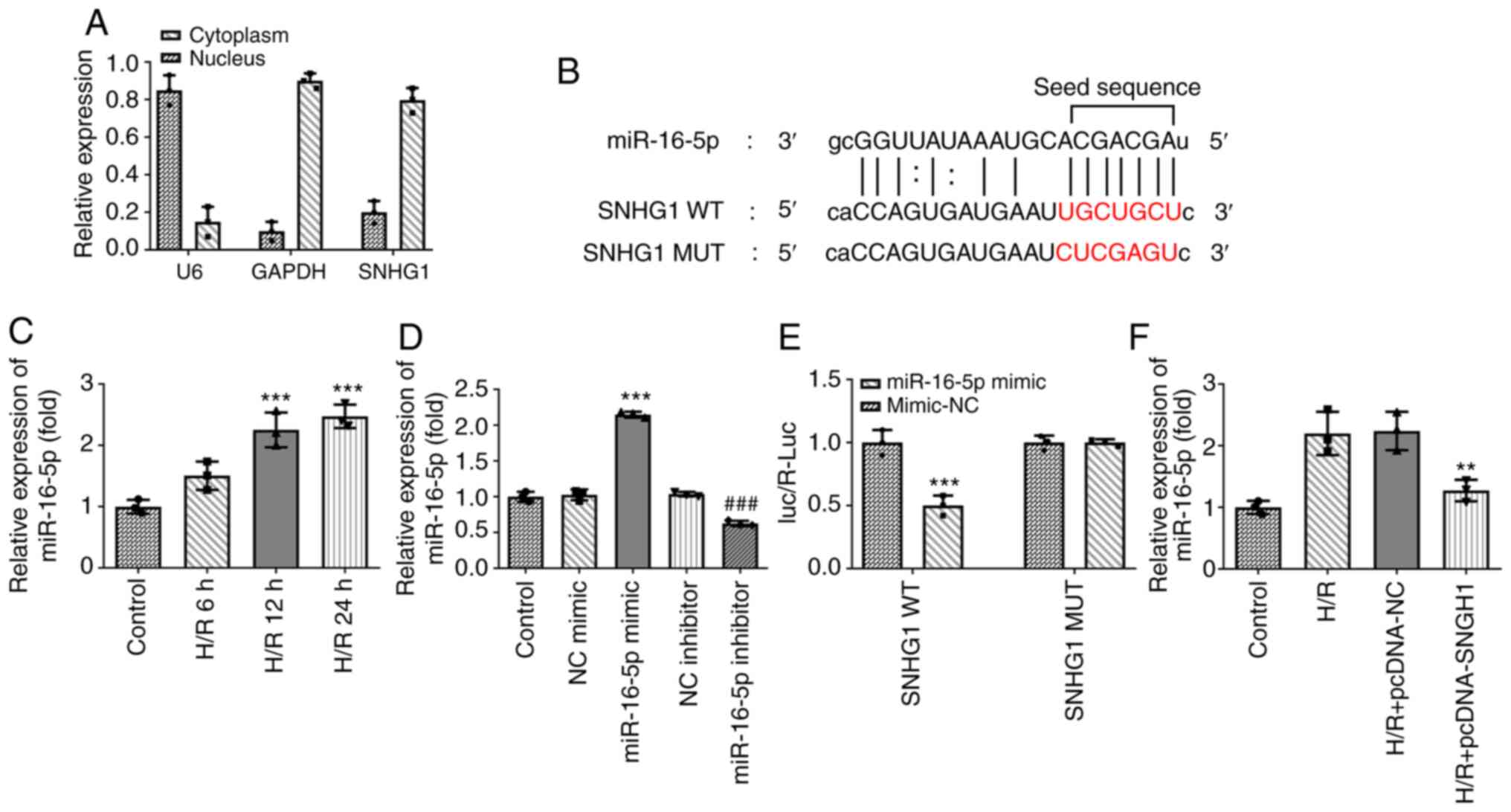
SNHG1 directly targets miR-16-5p
The present study further examined the underlying
mechanism of SNHG1 in myocardial ischemic injury. miR-16-5p was
predicted to be a target gene of SNHG1 using The Encyclopedia of
RNA Interactomes (ENCORI) database (Fig. 3B). In addition, the results of
RT-qPCR revealed that SNHG1 was localized in the cytoplasm and
miR-16-5p was highly expressed in H9c2 cells treated with H/R
(Fig. 3A and C). Subsequently,
miR-16-5p mimic and miR-16-5p inhibitor were successfully
transfected to modify the expression levels of miR-16-5p in cells
(Fig. 3D). The results of the
luciferase reporter assays demonstrated that compared with the
miR-NC group, miR-16-5p mimic significantly decreased the
luciferase activity of SNHG1 WT, while miR-116-5p mimic had no
effect on the luciferase activity of SNHG1 MUT (Fig. 3E). As shown in Fig. 3F, overexpression of SNHG1 markedly
inhibited miR-16-5p expression in H9c2 cells treated with H/R. In
general, SNHG1 could negatively regulate miR-16-5p.
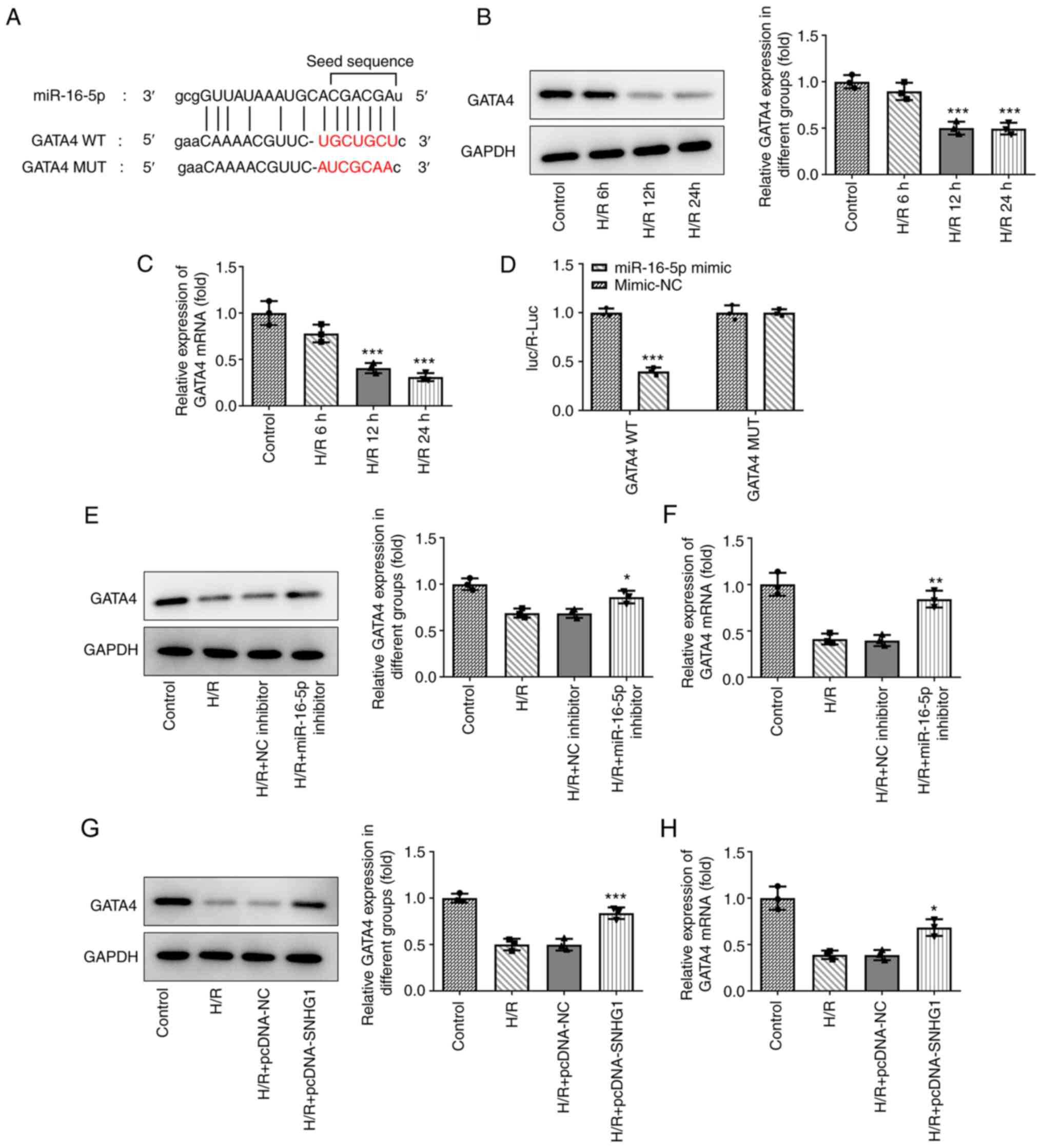
miR-16-5p directly targets GATA4
The downstream target genes of miR-16-5p were
predicted using the ENCORI database. As shown in Fig. 4A, GATA4 was predicted to be a target
gene of miR-16-5p. Compared with those in the control group, the
expression levels of GATA4 were downregulated in H9c2 cells treated
with H/R (Fig. 4B and C). The
results of the luciferase reporter assays demonstrated that
compared with the miR-NC group, miR-16-5p mimic markedly decreased
the luciferase activity of GATA4 WT, while miR-16-5p mimic had no
effect on the luciferase activity of GATA4 MUT (Fig. 4D). In addition, miR-16-5p inhibitor
(Fig. 4E and F) and overexpression
of SNHG1 (Fig. 4G and H) and
markedly promoted GATA4 expression in H9c2 cells treated with
H/R.
GATA4 and SNHG1 promoters combine to
form the positive feedback loop
The present study aimed to examine the associations
among SNHG1, miR-16-5p and GATA4. The expression levels of GATA4 in
cells were altered by transfection with shRNA-GATA4 and
pcDNA-GATA4. As shown in Fig. 5A,
compared with those in the control group, the expression levels of
GATA4 in the shRNA-GATA-2 group were the lowest and those in the
pcDNA-GATA4 group were the highest. In addition, overexpression of
GATA4 markedly promoted GATA4 expression (Fig. 5B). Of note, it was predicted by the
JASPAR database that there are two binding sites, S1 and S2,
between GATA4 and SNHG1 (Fig. 5C).
The results of the luciferase reporter assay revealed that
overexpression of GATA4 markedly promoted the luciferase activity
of SNHG1 WT compared with the control group, while overexpression
of GATA4 had no effect on the luciferase activity of SNHG1 MUT
(Fig. 5D and E). The results of
ChIP assay showed the enrichment of GATA4 on the promoter of SNHG1
(Fig. 5F).
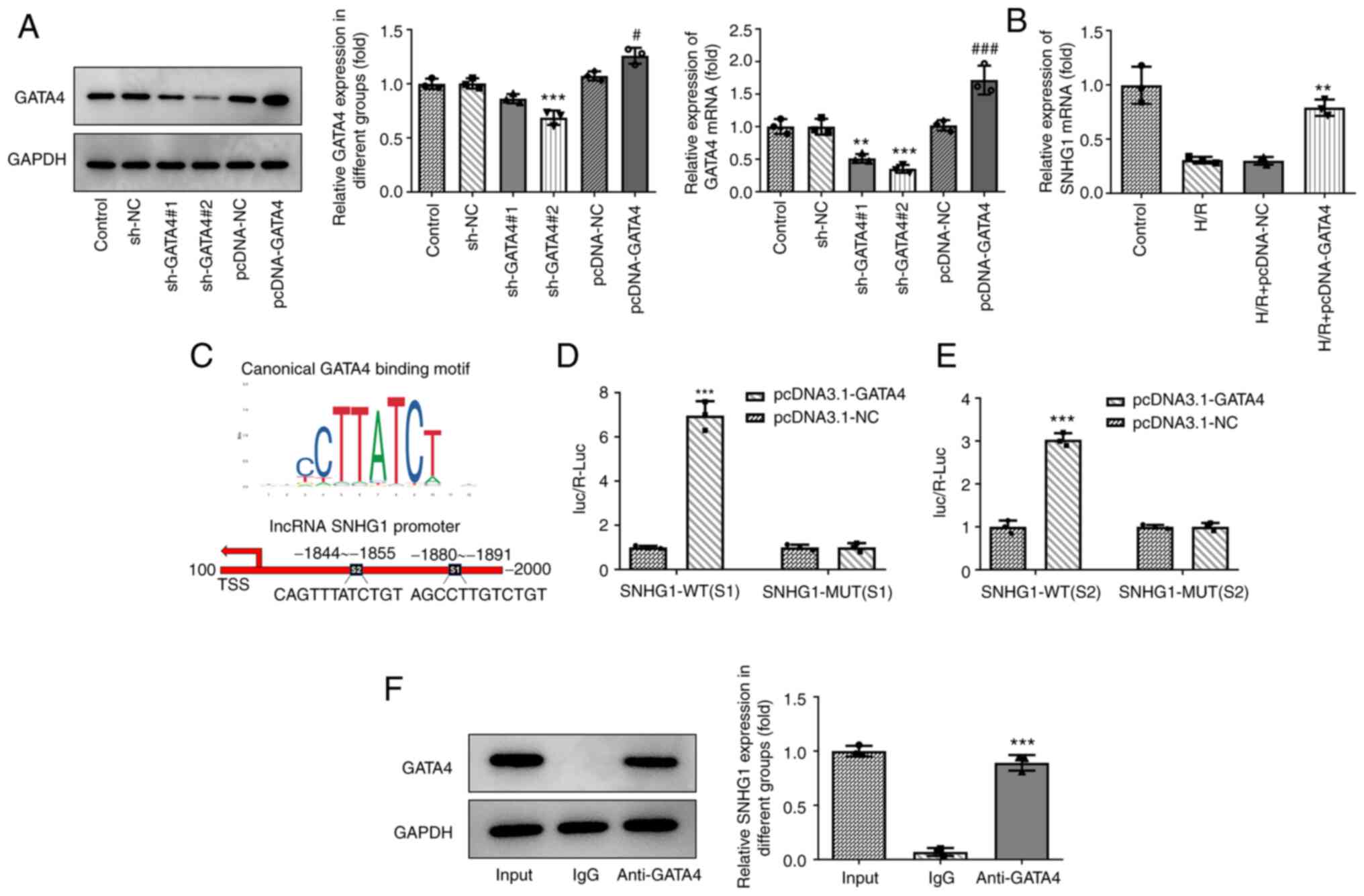 | Figure 5.GATA4 and SNHG1 promoters combine to
form the positive loop feedback. (A) The expression of GATA4 after
transfection of sh-GATA4 and pcDNA-GATA4 was measured via RT-qPCR
and western blotting. **P<0.01 and ***P<0.001 vs. sh-NC;
#P<0.05 and ###P<0.001 vs. pcDNA-NC.
(B) Effect of GATA4 overexpression on the expression of SNHG1 in
H9c2 cells was measured via RT-qPCR. **P<0.01 vs. H/R +
pcDNA-NC. (C) JASPAR analysis predicted two GATA4 binding sites, S1
and S2, on the SNHG1 promoter. (D and E) The relationship between
GATA4 and SNHG1 was detected using a luciferase reporter assay.
***P<0.001 vs. pcDNA-NC. (F) Chromatin immunoprecipitation
assay-coupled to RT-qPCR to verify GATA4 binding to the promoter of
SNHG1. ***P<0.001 vs. IgG. GATA4, GATA binding protein 4; SNHG1,
small nucleolar RNA host gene 1; sh-, short hairpin RNA; RT-qPCR,
reverse transcription-quantitative PCR; NC, negative control; H/R,
hypoxia/reoxygenation; WT, wild-type; MUT, mutant. |
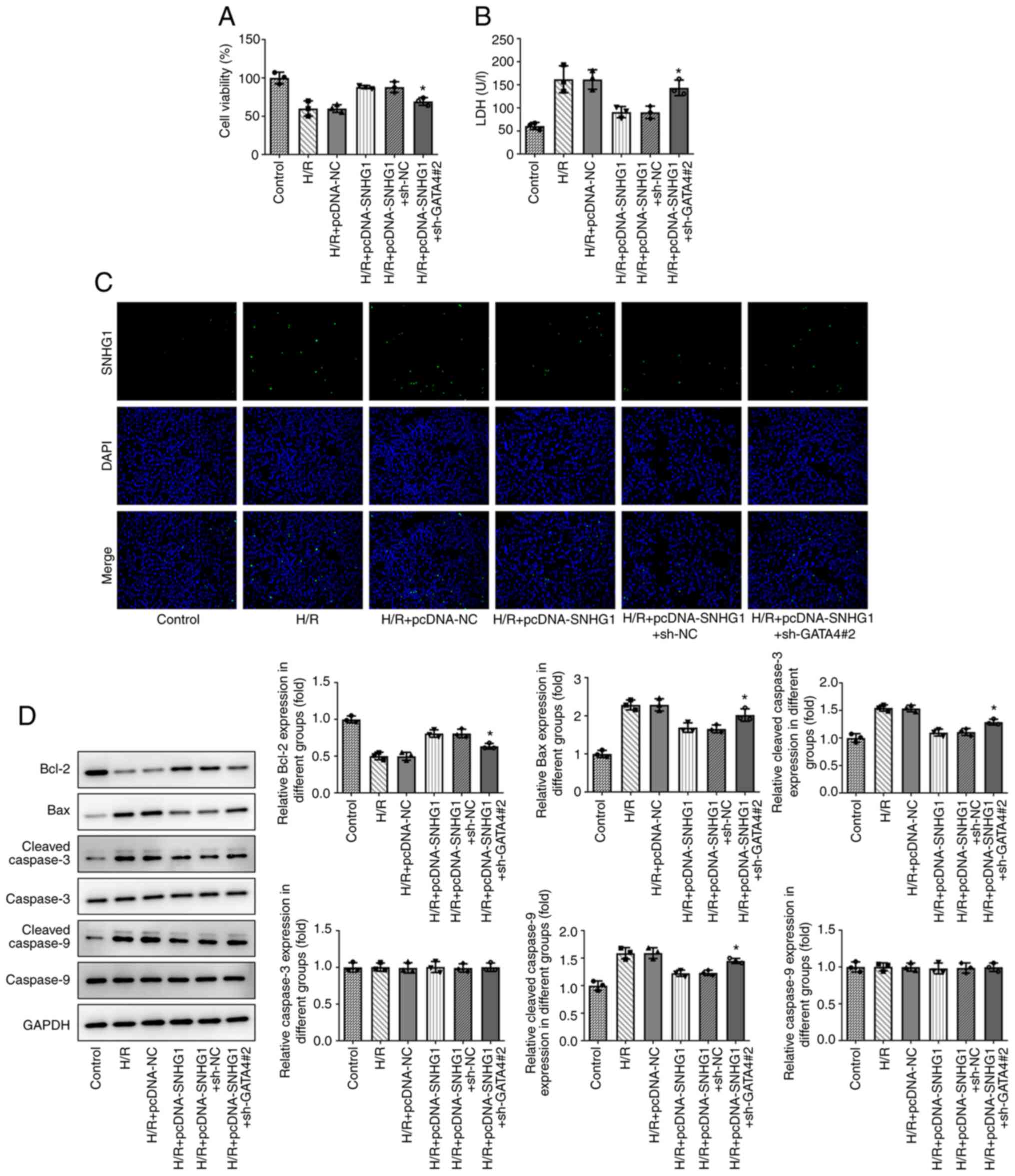
Overexpression of SNHG1 attenuates the
injury of H9c2 cells treated with H/R by upregulating GATA4
As shown in Fig. 6A,
CCK-8 revealed that GATA4 knockdown reduced the inhibitory effect
of SNHG1 overexpression on the viability of H9c2 cells treated with
H/R. Furthermore, the results of LDH showed that GATA4 knockdown
reduced the inhibitory effect of SNHG1 overexpression on the
expression of LDH in H9c2 cells treated with H/R (Fig. 6B). As shown in Fig. 6C and D, TUNEL analysis and western
blotting demonstrated that GATA4 knockdown partially reversed the
effect of SNHG1 overexpression on the apoptosis of H9c2 cells
treated with H/R.
Discussion
To the best of our knowledge, the present study was
the first to demonstrate the protective effect of SNHG1 on ischemic
cardiomyocyte injury. Specifically, SNHG1 inhibited ischemic
cardiomyocyte injury via regulation of the miR-16-5p/GATA4 axis.
Notably, GATA4 could bind to the promoter region of SNHG1. These
results demonstrated the existence of the SNHG1/miR-16-5p/GATA4
positive feedback loop and provided a novel therapeutic target for
the treatment of ischemic cardiomyocyte injury.
It has been demonstrated that lncRNAs serve an
important role in the occurrence and development of tumors, and
abnormal lncRNA expression has been used as a prognostic marker for
numerous tumors, such as hepatocellular carcinoma, colorectal
cancer and breast cancer (4–6).
lncRNA SNHG1, located on chromosome 11, is abnormally highly
expressed in a variety of tumor tissues and can promote the
proliferation of lung, liver, gastric and prostate cancer, and is
associated with poor prognosis in tumors (13–16).
Beyond this tumor-suppressive role, SNHG1 has a protective effect
on cardiomyocyte injury induced by various factors. For example,
SNHG1 protects cardiomyocytes from toxic damage induced by
doxorubicin by inhibiting miR-195 (8). SNHG1 has also been found to inhibit
cardiomyocyte injury induced by H2O2 by
regulating the miR-195/Bcl-2 axis (9). In addition, a recent study revealed
that SNHG1 attenuates vascular endothelial cell injury induced by
sponging miR-140-3p (10).
Therefore, it was suggested that SNHG1 may have a protective effect
on cardiomyocyte injury induced by H/R. In the present study, SNHG1
expression was downregulated in H9c2 cells induced by H/R.
Overexpression of SNHG1 markedly promoted the viability of H9c2
cells treated with H/R and inhibited apoptosis. These results
suggested that SNHG1 serves an important protective role in
cardiomyocyte injury induced by H/R.
A recent study revealed that lncRNAs contain miRNA
response elements, which can bind to their target miRNAs to inhibit
their biological functions (17).
This mechanism has been repeatedly demonstrated in multiple tumor
models (18–20). In addition, miRNAs serve an
important role in the occurrence and development of various
cardiovascular diseases (21). In
the present study, bioinformatics analysis revealed that miR-16-5p
was a potential target gene of SNHG1. The results of the luciferase
reporter gene assay revealed that SNHG1 could directly target
miR-16-5p. In addition, miR-16-5p expression is downregulated in a
variety of tumor tissues and exerts an antitumor effect via
induction of apoptosis (22,23).
It is worth noting that miR-16-5p has been found to be highly
expressed in AC16 cardiomyocytes treated with I/R, and miR-16-5p
knockdown could markedly promote cell viability and angiogenesis
via inhibition of apoptosis (24).
In the present study, miR-16-5p expression was upregulated in H9c2
cells treated with H/R, and the overexpression of SNHG1 markedly
decreased the expression levels of miR-16-5p.
The present study further explored the mechanism of
SNHG1 in cardiomyocyte injury induced by H/R. GATA4 was predicted
as a potential target gene of miR-16-5p using bioinformatics
analysis. GATA4 is an important regulator in the early stage of
development and serves an important role in the development of the
heart and intestines (25). A
previous study revealed that overexpression of GATA4 promoted P19
cells to differentiate into cardiomyocytes (26). It is worth noting that
downregulation of miR-122 has been shown to reduce cardiomyocyte
apoptosis induced by H/R by regulating GATA4 (27). In addition, Astragaloside IV can
enhance the activity of H9c2 cells treated with H/R by upregulating
GATA4 (28). Importantly, using the
JASPAR database in the present study, it was identified that there
may be two binding sites between GATA4 and SNHG1, indicating the
existence of the SNHG1/miR-16-5p/GATA4 positive feedback loop. In
the present study, miR-16-5p could negatively target GATA4. The
results of the luciferase reporter gene and ChIP assays
demonstrated that GATA4 could bind to the SNHG1 promoter. SNHG1
knockdown partially reversed the effects of SNHG1 overexpression on
cardiomyocyte viability and apoptosis induced by H/R.
In conclusion, the present results demonstrated that
the positive feedback loop of SNHG1/miR-16-5p/GATA4 could
potentially serve an important role in cardiomyocyte injury induced
by H/R. SNHG1 could improve cardiomyocyte injury via upregulation
of GATA4 by targeting miR-16-5p. However, due to the limitation of
time and funds, only H9c2 cells were used to explore the regulatory
effects of SNHG1 in H/R-induced cardiomyocyte injury via
upregulation of GATA4 by targeting miR-16-5p. Further studies are
needed to focus on nuclear factor E2-associated factor 2/heme
oxygenase 1 signaling pathways, and future experiments should
include animal models to further support the findings of the
present study.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JG and LD carried out the data collection and data
analysis. JG and YZ conceived and designed the study, and wrote and
revised the manuscript. JG and YZ confirm the authenticity of all
the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hall TM, Gordon C, Roy R and Schwenke DO:
Delayed coronary reperfusion is ineffective at impeding the dynamic
increase in cardiac efferent sympathetic nerve activity following
myocardial ischemia. Basic Res Cardiol. 111:352016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hao PP, Jiang F, Chen YG, Yang J, Zhang K,
Zhang MX, Zhang C, Zhao YX and Zhang Y: Traditional Chinese
medication for cardiovascular disease. Nat Rev Cardiol. 12:115–122.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Writing Group Members, . Mozaffarian D,
Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, Das SR, de
Ferranti S, Després JP, et al: Heart disease and stroke
statistics-2016 update: A report from the American heart
association. Circulation. 133:e38–e360. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ou F, Su K, Sun J, Liao W, Yao Y, Zheng Y
and Zhang Z: The LncRNA ZBED3-AS1 induces chondrogenesis of human
synovial fluid mesenchymal stem cells. Biochem Biophys Res Commun.
487:457–463. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu Y, Zheng Y, Liu H and Li T: Modulation
of IGF2BP1 by long non-coding RNA HCG11 suppresses apoptosis of
hepatocellular carcinoma cells via MAPK signaling transduction. Int
J Oncol. 51:791–800. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tycowski KT, Shu MD and Steitz JA:
Requirement for intron-encoded U22 small nucleolar RNA in 18S
ribosomal RNA maturation. Science. 266:1558–1561. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen S, Wang J and Zhou Y: Long non-coding
RNA SNHG1 protects human AC16 cardiomyocytes from doxorubicin
toxicity by regulating miR-195/Bcl-2 axis. Biosci Rep.
39:BSR201910502019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang N, Meng X, Mei L, Hu J, Zhao C and
Chen W: The long non-coding RNA SNHG1 attenuates cell apoptosis by
regulating miR-195 and BCL2-like protein 2 in human cardiomyocytes.
Cell Physiol Biochem. 50:1029–1040. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang S, Ren K, Li B, Li F, Liang Z, Hu J,
Xu B and Zhang A: LncRNA SNHG1 alleviates
hypoxia-reoxygenation-induced vascular endothelial cell injury as a
competing endogenous RNA through the HIF-1α/VEGF signal pathway.
Mol Cell Biochem. 465:1–11. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang L, Jiang X, Wang Z, Zhong X, Tai S
and Cui Y: Small nucleolar RNA host gene 1: A new biomarker and
therapeutic target for cancers. Pathol Res Pract. 214:1247–1252.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative Pcr and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cui Y, Zhang F, Zhu C, Geng L, Tian T and
Liu H: Upregulated lncRNA SNHG1 contributes to progression of
non-small cell lung cancer through inhibition of miR-101-3p and
activation of Wnt/β-catenin signaling pathway. Oncotarget.
8:17785–17794. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Zhou D, Ying M, Chen M, Chen P,
Chen Z and Zhang F: Expression of long non-coding RNA (lncRNA)
small nucleolar RNA host gene 1 (SNHG1) exacerbates hepatocellular
carcinoma through suppressing miR-195. Med Sci Monit. 22:4820–4829.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu Y, Ma Z, He Y, Liu W, Su Y and Tang Z:
LncRNA-SNHG1 contributes to gastric cancer cell proliferation by
regulating DNMT1. Biochem Biophys Res Commun. 491:926–931. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Zhang Z, Xiong L, Guo C, Jiang T,
Zeng L, Li G and Wang J: SNHG1 lncRNA negatively regulates
miR-199a-3p to enhance CDK7 expression and promote cell
proliferation in prostate cancer. Biochem Biophys Res Commun.
487:146–152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden. RNA
language? Cell. 146:353–358. 2011.PubMed/NCBI
|
|
18
|
Lu Q, Shan S, Li Y, Zhu D, Jin W and Ren
T: Long noncoding RNA SNHG1 promotes non-small cell lung cancer
progression by up-regulating MTDH via sponging miR-145-5p. FASEB J.
32:3957–3967. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao L, Kong H, Sun H, Chen Z, Chen B and
Zhou M: LncRNA-PVT1 promotes pancreatic cancer cells proliferation
and migration through acting as a molecular sponge to regulate
miR-448. J Cell Physiol. 233:4044–4055. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mou K, Liu B, Ding M, Mu X, Han D, Zhou Y
and Wang LJ: lncRNA-ATB functions as a competing endogenous RNA to
promote YAP1 by sponging miR-590-5p in malignant melanoma. Int J
Oncol. 53:1094–1104. 2018.PubMed/NCBI
|
|
21
|
Meng Q, Liu Y, Huo X, Sun H, Wang Y and Bu
F: MicroRNA-221-3p contributes to cardiomyocyte injury in
H2O2-treated H9c2 cells and a rat model of
myocardial ischemia-reperfusion by targeting p57. Int J Mol Med.
42:589–596. 2018.PubMed/NCBI
|
|
22
|
Xu F, Zhang X, Lei Y, Liu X, Liu Z, Tong T
and Wang W: Loss of repression of HuR translation by miR-16 may be
responsible for the elevation of HuR in human breast carcinoma. J
Cell Biochem. 111:727–734. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jang JY, Lee JK, Jeon YK and Kim CW:
Exosome derived from epigallocatechin gallate treated breast cancer
cells suppresses tumor growth by inhibiting tumor-associated
macrophage infiltration and M2 polarization. BMC Cancer.
13:4212013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang X, Shang Y, Dai S, Wu W, Yi F and
Cheng L: MicroRNA-16-5p aggravates myocardial infarction injury by
targeting the expression of insulin receptor substrates 1 and
mediating myocardial apoptosis and angiogenesis. Curr Neurovasc
Res. 17:11–17. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Molkentin JD: The zinc finger-containing
transcription factors GATA-4,-5, and-6 ubiquitously expressed
regulators of tissue-specific gene expression. J Biol Chem.
275:38949–38952. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ang YS, Rivas RN, Ribeiro AJS, Srivas R,
Rivera J, Stone NR, Pratt K, Mohamed TMA, Fu JD, Spencer CI, et al:
Disease model of GATA4 mutation reveals transcription factor
cooperativity in human cardiogenesis. Cell. 167:1734–1749. e222016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang W, Guo J, Li J, Bai C and Dong Y:
Downregulation of miR-122 attenuates hypoxia/reoxygenation
(H/R)-induced myocardial cell apoptosis by upregulating GATA-4.
Biochem Biophys Res Commun. 478:1416–1422. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang JJ, Zhang XH, Ma XH, Duan WJ, Xu NG,
Chen YJ and Liang L: Astragaloside IV enhances GATA-4 mediated
myocardial protection effect in hypoxia/reoxygenation injured H9c2
cells. Nutr Metab Cardiovasc Dis. 30:829–842. 2020. View Article : Google Scholar : PubMed/NCBI
|