Introduction
Emerging evidence suggests that cardiovascular
disease is the leading cause of death worldwide (1). Myocardial infarction, the most fatal
type of cardiovascular disease, has attracted widespread attention
because of its increasing incidence, complications, poor outcomes
and limited treatment. A complex issue is that clinical treatment
for myocardial infarction to restore cardiovascular perfusion can
cause secondary damage. There is a large body of work examining
various regulators in ischemia/reperfusion (I/R), however, the
molecular mechanisms underlying I/R injury remain unclear.
Therefore, there is a clear need to study the molecular etiology of
I/R injury in order to develop more effective treatment strategies
and improve prognosis.
Pyroptosis, also known as cellular inflammatory
necrosis, is a form of programmed cell death in which cells swell
until the plasma membrane ruptures, causing the release of cellular
contents to activate a strong inflammatory response (2). Pyroptosis is an important natural
immune response in the body and plays a significant role in
combating infection (3). It was
previously demonstrated that NOD-like receptor protein 3 (NLRP3)
inflammasome activation-dependent pyroptosis was involved in
cardiovascular I/R injury (3) and
that the BRCA1/BRCA2-containing complex subunit 3 (BRCC3), a
deubiquitinating enzyme, is a critical regulator of NLRP3 activity
(4). This indicated that the
regulation of BRCC3 expression, and its direct interaction with
NLRP3, could be a potential strategy for the treatment of
cardiovascular I/R injury.
Long non-coding RNAs (lncRNAs) are non-coding RNAs
that are >200 nucleotides in length (5). Studies have reported that lncRNAs play
an important role in numerous biological processes, such as dose
compensation effect, epigenetic regulation, cell cycle regulation
and cell differentiation regulation, and have become an important
area of interest in genetics (6).
It was found that the lncRNA nuclear enriched abundant transcript 1
(NEAT1) aggravated cardiovascular I/R injury (7), but the potential regulatory mechanisms
have not yet been elucidated. At present, no reports have examined
NEAT1 regulated NLRP3 inflammasome activation-dependent pyroptosis
in cardiovascular I/R injury.
MicroRNAs (miRNA/miRs) are a class of endogenous
regulatory non-coding RNAs found in eukaryotes that are 20–25
nucleotides in length (8). Emerging
evidence has suggested a functional role of lncRNAs by acting as
competing endogenous RNAs (ceRNAs) that regulate specific RNA
transcripts through competing for shared miRNAs (9). To the best of our knowledge, no
previous studies have explored miR-204 and NLRP3 inflammasome
activation-dependent pyroptosis in cardiovascular I/R injury.
In the present study, the regulatory effect of NEAT1
and related miR-204/BRCC3 axis in NLRP3 inflammasome
activation-dependent pyroptosis was investigated in human umbilical
vein endothelial cells (HUVECs) exposed to hypoxia/reoxygenation
(H/R).
Materials and methods
Cell culture
HUVECs (cat. no. aBFN6021424) were obtained from
Shanghai Institute of Biochemistry and Cell Biology and cultured in
DMEM containing 10% FBS and 1% penicillin/streptomycin (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C, 5% CO2.
Cell transfection
Short hairpin RNAs (shRNAs) against NEAT1
(NEAT1-shRNA, 5′-CCGTGGTGTGTGTTGTGGAATCTGT-3′)/BRCC3 (BRCC3-shRNA,
5′-GUACUGGGUUUGUUACAGAUU-3′) and corresponding negative control
(con-shRNA: NEAT1-con-shRNA, 5′-CCGTGTGTGTGGTGTAGTACGTTGT-3′;
BRCC3-con-shRNA, 5′-UCACUGCGCUCGAUGCAGUTT-3′), pcDNA-NEAT1 and
corresponding negative control (pcDNA-con), miR-204 mimics
(5′-TTCCCTTTGTCATCCTATGCCT-3′) and corresponding negative control
(miR-con, 5′-CGATCGCATCAGCATCGATTGC-3′), were constructed by
Shanghai GenePharma Co., Ltd. The constructs (50 nM) were
transfected into 1×106 HUVECs using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Transfection efficiency was observed, and cells were used for
subsequent experiments 48 h after transfection.
Establishment of H/R cell model
A cellular H/R model was constructed based on the
methods of Deng et al (10).
Briefly, medium containing 4.5 g/l glucose and 10% FBS was replaced
with a sugar-free, serum-free medium before exposure to hypoxic
conditions. HUVECs were cultured for 12 h at 1% oxygen, 5%
CO2 and 94% nitrogen to induce hypoxia. Following this,
cells were exposed to reoxygenation conditions for 4 h, at 37°C and
5% CO2, in medium containing 4.5 g/l sugar and 10%
FBS.
Cell Counting Kit (CCK)-8 cell
viability assay
CCK-8 was used according to the manufacturer's
protocol. Transfected or untransfected HUVECs were seeded in a
96-well plate at a density of 1×105 cells/100 µl. After
undergoing conventional H/R operations, 10 µl detection reagent
(Dojindo Molecular Technologies, Inc.) was added to HUVECs for 2–4
h at 37°C. The OD value at a wavelength of 450 nm was measured with
a microplate reader.
Lactate dehydrogenase (LDH) assay
Transfected or untransfected HUVECs were seeded in a
6-well plate at a density of 1×106 cells/2 ml. After
undergoing conventional H/R operations, LDH activity was determined
using commercial kits (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions.
Luciferase reporter assay
Wild-type (WT) or mutant (MUT) NEAT1 and BRCC3 were
inserted into a pGL3 promoter vector (Shanghai GenePharma Co.,
Ltd.). HUVECs were transfected with pGL3-NEAT1/BRCC3-WT or
pGL3-NEAT1/BRCC3-MUT and miR-mimics or mimics control (Shanghai
GenePharma Co., Ltd.) using Lipofectamine® 2000
transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
for 48 h. The luciferase activity was evaluated using a Luciferase
Reporter Assay System (Shanghai GenePharma Co., Ltd.).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from 1×106 HUVECs
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). The first strand cDNA was synthesized from 1 µg
total RNA using a Prime Script RT kit and gDNA Eraser (Takara Bio,
Inc.). The relative mRNA levels were analyzed using the
2−ΔΔCq method (11) and
normalized to the internal reference genes U6 and GAPDH (12).
Western blot analysis
Total protein was extracted from 1×107
HUVECs using RIPA lysis buffer (BestBio). The proteins were
separated via sodium dodecyl sulfate polyacrylamide gel
electrophoresis on a 10% gel, and subsequently transferred to PVDF
membranes. The membranes were incubated with 5% fat-free milk at
room temperature for 1 h. Anti-BRCC3 antibodies (cat. no.
aPA5-20426, 1:1,000, Invitrogen; Thermo Fisher Scientific, Inc.),
anti-NLRP3 antibodies (cat. no. aab263899, 1:1,000, Abcam),
anti-cleaved caspase-1 (CASP1) antibodies (cat. no. aPA5-39882,
1:1,000, Invitrogen; Thermo Fisher Scientific, Inc.), anti-cleaved
gasdermin D (GSDMD) antibodies (cat. no. ab215203, 1:1,000, Abcam)
and anti-β actin antibodies (cat. no. ab8226, 1:1,000, Abcam) were
used as the primary antibodies at 4°C for 12 h. The secondary
antibodies (cat. nos. 31430 and G-21234, 1:10,000, Invitrogen;
Thermo Fisher Scientific, Inc.) was added for 2 h at room
temperature. Protein bands were detected using Pierce ECL Western
blot analysis substrate (Thermo Fisher Scientific, Inc.).
Statistical analyses
Data are presented as the mean ± SD. All statistical
analyses were conducted using SPSS 19.0 software (IBM Corp.). The
normal distribution and homogeneity of variance of data were tested
first. Differences between two groups were compared by an unpaired
Student's t-test, while multiple groups were compared by one-way
analysis of variance (ANOVA) when data were normally distributed
and had homogeneity of variance. Bonferroni correction was used to
assess multiple comparisons following one-way ANOVA. The rank-sum
test was used to test the data that was not normally distributed or
lacked homogeneity of variance. P<0.05 was considered to
indicate a statistically significant difference.
Results
H/R evokes damage and NLRP3
inflammasome activation-dependent pyroptosis in HUVECs
The HUVECs were subjected to an in vitro
model of hypoxia for 12 h and reoxygenation for 4 h. As a result,
H/R significantly decreased cell viability (Fig. 1A) and increased LDH activity
(Fig. 1B), protein expression of
BRCC3, NLRP3, ASC, cleaved CASP1 and cleaved GSDMD (Fig. 1F), mRNA expression of interleukin
(IL)-1β and IL-18 (Fig. 1G and H)
and secreted levels of IL-1β and IL-18 (Fig. 1I and J). Meanwhile, the expression
of NEAT1 (Fig. 1C) and BRCC3
(Fig. 1E) were significantly
increased, whereas miR-204 expression was significantly decreased
(Fig. 1D) in the H/R group as
measured by RT-qPCR.
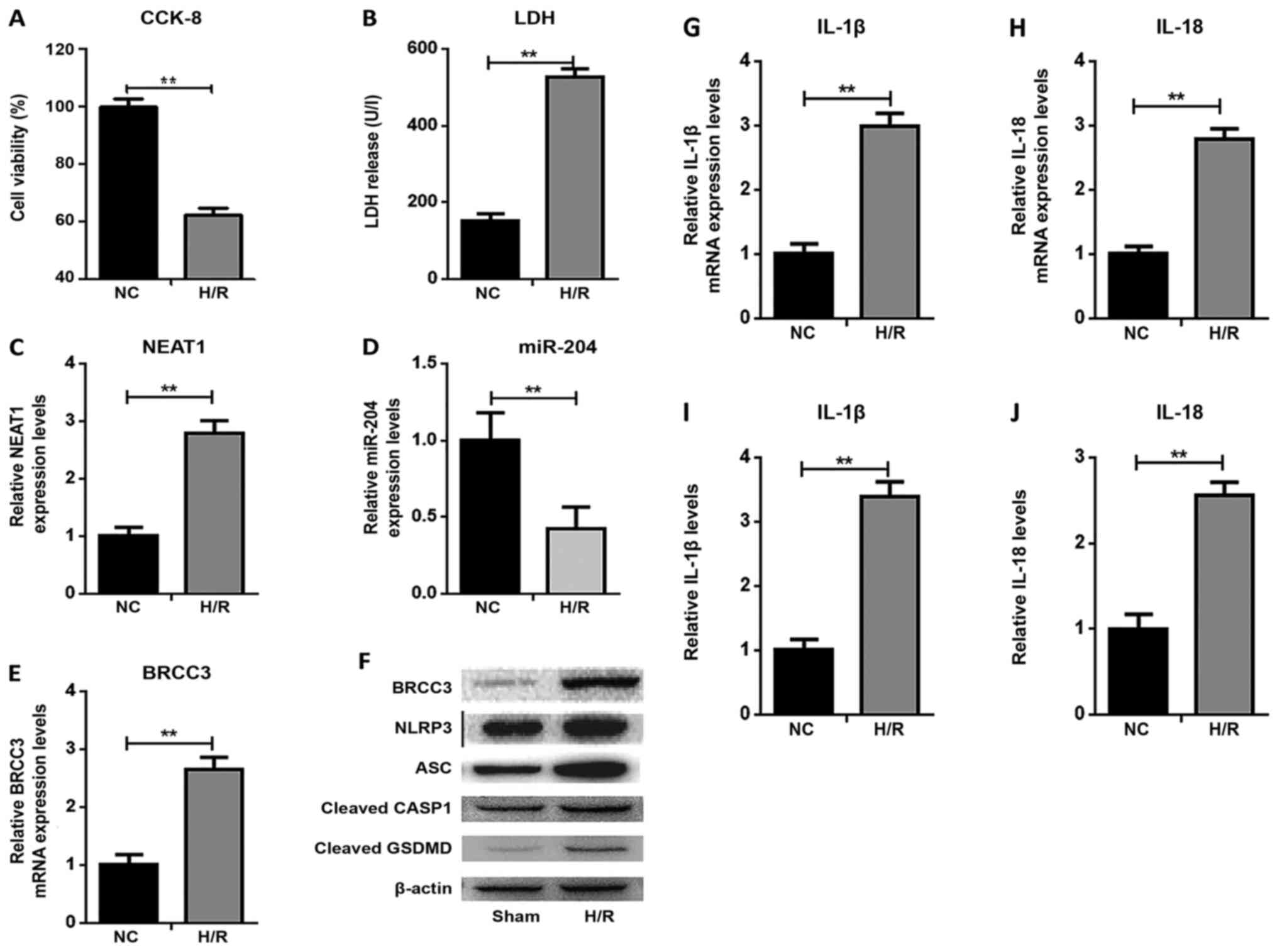 | Figure 1.H/R evokes damage and NLRP3
inflammasome activation-dependent pyroptosis in HUVECs. (A)
Detection of cell viability and (B) LDH activity in HUVECs after
H/R. RT-qPCR analysis of (C) NEAT1, (D) miR-204 and (E) BRCC3
expression. (F) The expression levels of BRCC3, NLRP3, ASC, cleaved
CASP1 and cleaved GSDMD protein were examined by western blotting.
The expression levels of (G) IL-1β and (H) IL-18 mRNA were examined
by RT-qPCR. The levels of secreted (I) IL-1β and (J) IL-18 were
evaluated by ELISA. Data are shown as the mean ± SD from five
different experiments. **P<0.01. H/R, hypoxia/reoxygenation;
LDH, lactate dehydrogenase; IL, interleukin; NLRP3, NOD-like
receptor protein 3; ASC, apoptosis speck-like protein; CASP1,
caspase-1; GSDMD, gasdermin D; NEAT1, nuclear enriched abundant
transcript 1; BRCC3, BRCA1/BRCA2-containing complex subunit 3; miR,
microRNA; RT-qPCR, reverse transcription-quantitative PCR; NC,
negative control; HUVEC, human umbilical vein endothelial cell. |
Knockdown of BRCC3 represses NLRP3
inflammasome activation-dependent pyroptosis and cell damage in
HUVECs
To investigate the expression of BRCC3 and the
interaction between NLRP3 and BRCC3 in H/R-induced pyroptosis and
cell damage, BRCC3 expression was depleted by the transfection of
BRCC3-shRNA into cells prior to induction of hypoxia (Fig. 2A). As a result, BRCC3-shRNA
treatment significantly reversed the H/R-induced reduction in cell
viability (Fig. 2B) and H/R-induced
increase in LDH activity (Fig. 2C).
Protein expression of BRCC3, NLRP3, ASC, cleaved CASP1 and cleaved
GSDMD (Fig. 2D) were all reduced
following transfection with BRCC3-shRNA but not in con-shRNA.
Similarly, mRNA expression and secreted protein levels of IL-1β and
IL-18 (Fig. 2E-H) were reduced by
depletion of BRCC3 prior to H/R.
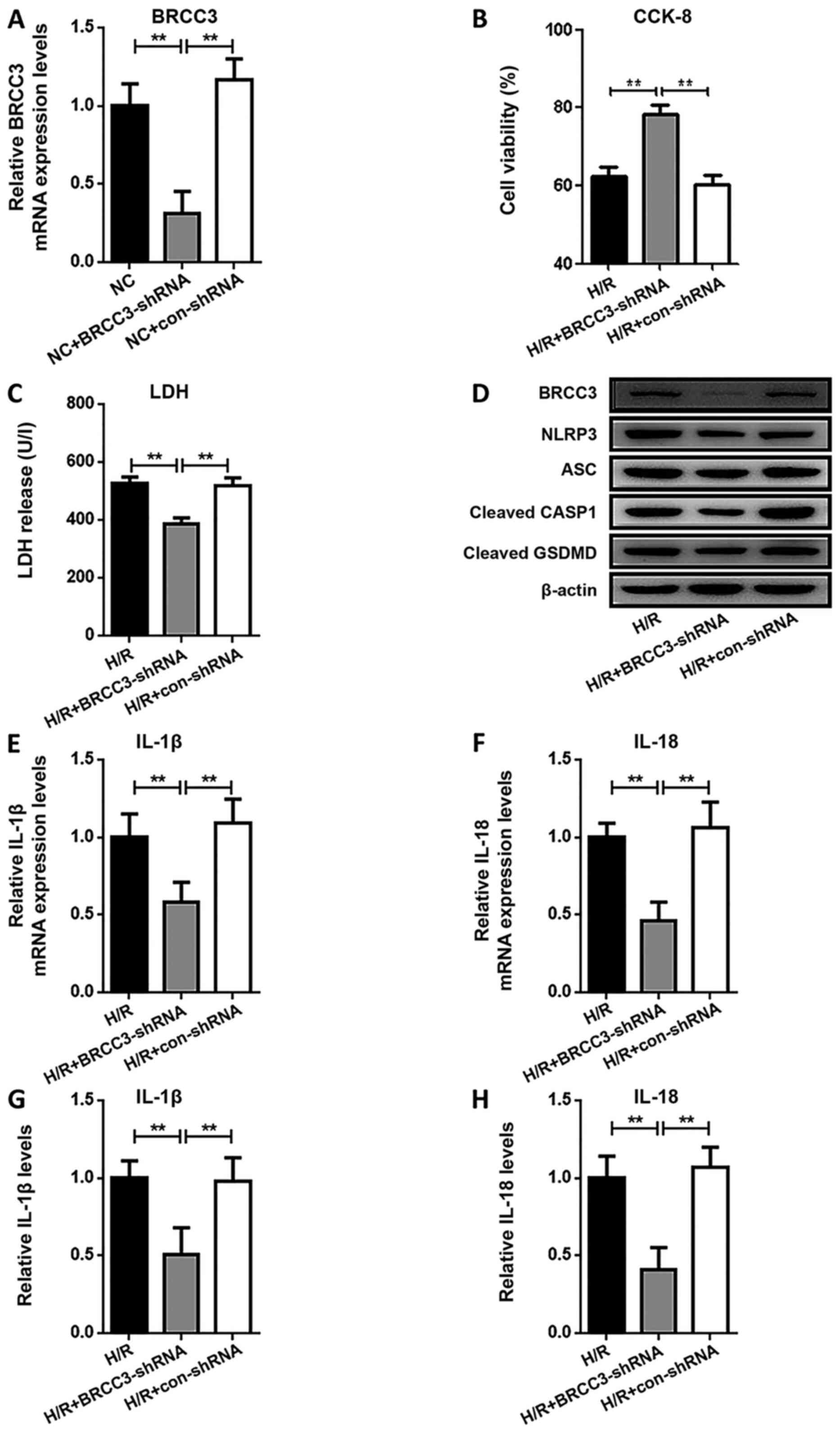 | Figure 2.Knockdown of BRCC3 represses NLRP3
inflammasome activation-dependent pyroptosis and cell damage in
HUVECs. (A) The expression of BRCC3 mRNA was examined by RT-qPCR.
Detection of (B) cell viability and (C) LDH activity in HUVECs
following H/R. (D) The expression levels of BRCC3, NLRP3, ASC,
cleaved CASP1 and cleaved GSDMD protein were examined by western
blotting. The expression levels of (E) IL-1β and (F) IL-18 mRNA
were examined by RT-qPCR. The levels of secreted (G) IL-1β and (H)
IL-18 were evaluated by ELISA. Data are shown as the mean ± SD from
five different experiments. **P<0.01. H/R,
hypoxia/reoxygenation; LDH, lactate dehydrogenase; IL, interleukin;
NLRP3, NOD-like receptor protein 3; ASC, apoptosis speck-like
protein; CASP1, caspase-1; GSDMD, gasdermin D; BRCC3,
BRCA1/BRCA2-containing complex subunit 3; RT-qPCR, reverse
transcription-quantitative PCR; NC, negative control; CCK-8, Cell
Counting Kit-8; shRNA, short hairpin RNA; HUVEC, human umbilical
vein endothelial cell. |
Knockdown of NEAT1 reverses
H/R-induced cell damage and NLRP3 inflammasome activation-dependent
pyroptosis
HUVECs were transfected with either NEAT1-shRNA or
shRNA-con before the induction of hypoxia to observe the function
of NEAT1 (Fig. 3A) in H/R-induced
cell damage and NLRP3 inflammasome activation-dependent pyroptosis.
As shown in Fig. 3, NEAT1-shRNA
significantly reversed the H/R-induced reduction in cell viability
(Fig. 3B) and H/R-induced increase
in LDH activity (Fig. 3C).
Additionally, NEAT1-shRNA transfection reduced the H/R-induced
increase in protein expression of BRCC3, NLRP3, ASC, cleaved CASP1
and cleaved GSDMD (Fig. 3G).
Similar to BRCC3 depletion, mRNA expression and secreted protein
levels of IL-1β and IL-18 (Fig.
3H-K) were significantly decreased by depletion of NEAT1 before
H/R. Of note, the expression of miR-204 was significantly increased
(Fig. 3E), whereas the expression
of BRCC3 was significantly reduced (Fig. 3F), following NEAT1-shRNA
transfection.
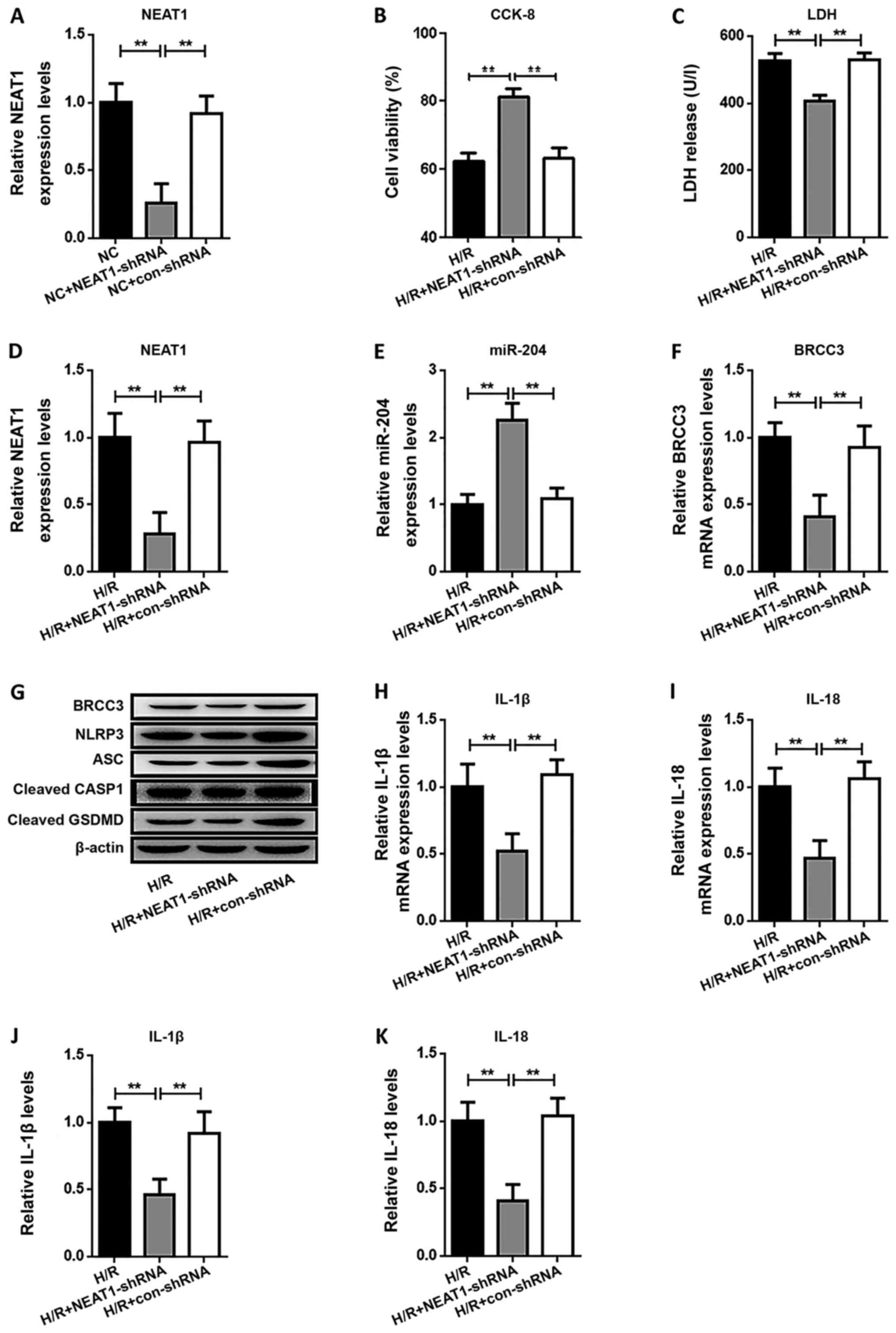 | Figure 3.Knockdown of NEAT1 reverses
H/R-induced cell damage by reducing NLRP3 inflammasome
activation-dependent pyroptosis. (A) The expression of NEAT1 mRNA
was examined by RT-qPCR. Detection of (B) cell viability and (C)
LDH activity in human umbilical vein endothelial cells following
H/R. RT-qPCR analysis of (D) NEAT1, (E) miR-204 and (F) BRCC3
expression. (G) The expression levels of BRCC3, NLRP3, ASC, cleaved
CASP1 and cleaved GSDMD protein were examined by western blotting.
The expression levels of (H) IL-1β and (I) IL-18 mRNA were examined
by RT-qPCR. The levels of secreted (J) IL-1β and (K) IL-18 were
evaluated by ELISA. Data are shown as the mean ± SD from five
different experiments. **P<0.01. H/R, hypoxia/reoxygenation;
LDH, lactate dehydrogenase; IL, interleukin; NLRP3, NOD-like
receptor protein 3; ASC, apoptosis speck-like protein; CASP1,
caspase-1; GSDMD, gasdermin D; NEAT1, nuclear enriched abundant
transcript 1; BRCC3, BRCA1/BRCA2-containing complex subunit 3; miR,
micro RNA; RT-qPCR, reverse transcription-quantitative PCR; NC,
negative control; CCK-8, Cell Counting Kit-8; shRNA, short hairpin
RNA. |
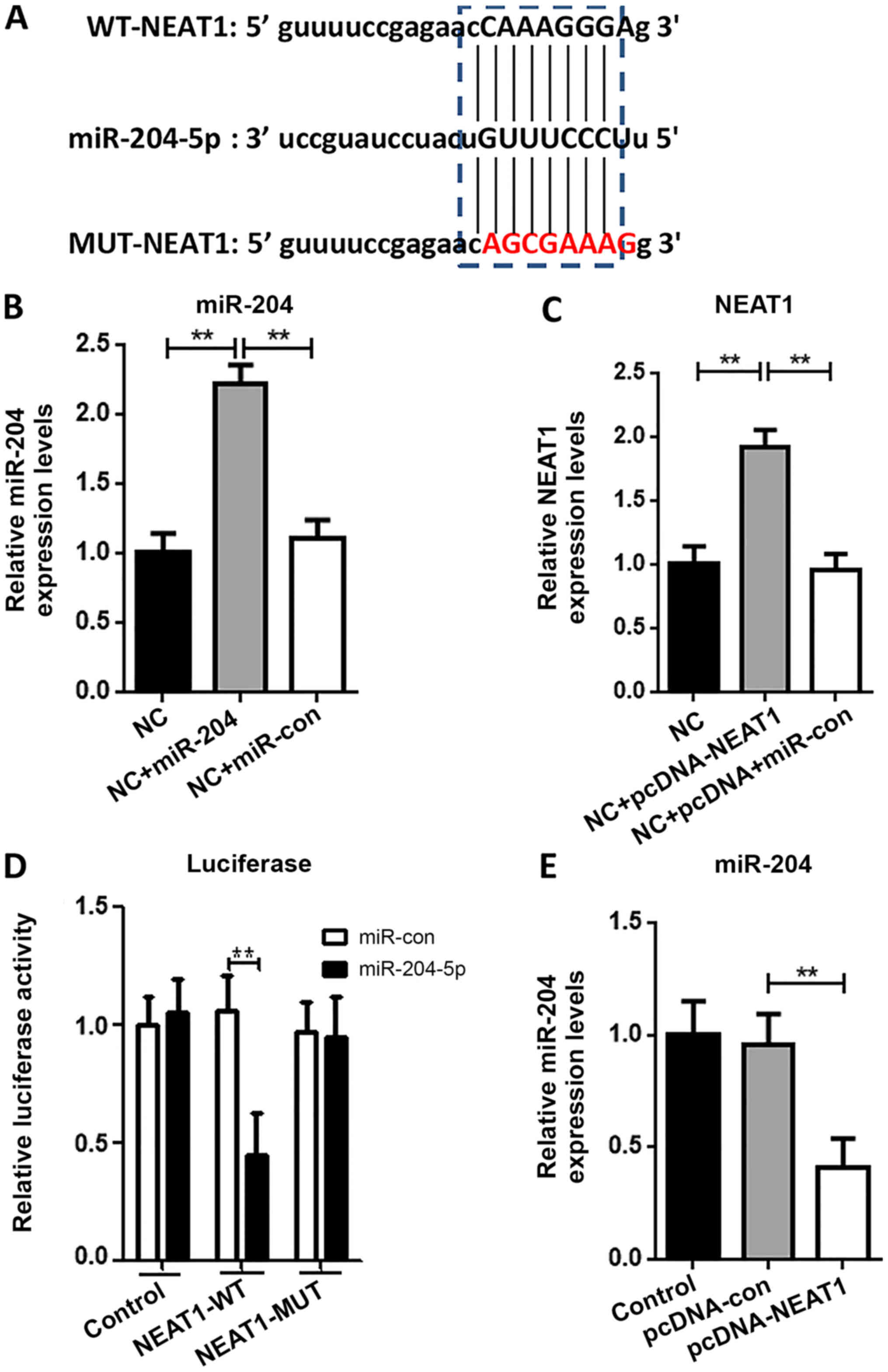
NEAT1 functions as a sponge of
miR-204
Prior research has indicated that lncRNAs act as
miRNA sponges in multiple diseases including I/R injury (7). In the present study it was found that
miR-204 was a target of NEAT1 (Fig.
4A). To further explore the association between NEAT1 and
miR-204, luciferase reporter assays were performed (Fig. 4B-C). This demonstrated that
NEAT1-WT, but not NEAT1-MUT, could be specifically reduced by
miR-204 mimics (Fig. 4D). In
addition, miR-204 expression was significantly decreased by
transient transfection of pcDNA-NEAT1, but not pcDNA-con (Fig. 4E).
NEAT1 aggravates H/R-induced cell
damage and NLRP3 inflammasome activation-dependent pyroptosis and
can be rescued by miR-204 overexpression
To further confirm whether NEAT1 regulates
H/R-induced injury by sponging miR-204, pcDNA-NEAT1 or pcDNA-con
was co-transfected into HUVECs with miR-204 mimics. It was found
that miR-204 mimics significantly increased cell viability
(Fig. 5A) and decreased LDH
activity (Fig. 5B) following H/R
compared with the control group. The relative mRNA levels of NEAT1,
and BRCC3 were reduced by overexpression of miR-204, while the
relative mRNA levels of miR-204 were increased (Fig. 5C-E). Protein expression of NLRP3,
ASC, cleaved CASP1 and cleaved GSDMD (Fig. 5F) were all reduced by overexpression
of miR-204. Both mRNA expression and secreted protein levels of
IL-1β and IL-18 (Fig. 5G-J) were
significantly reduced in cells transfected with miR-204 mimic.
However, overexpression of NEAT1 by transient transfection of
pcDNA-NEAT1 reversed the aforementioned changes induced by miR-204
expression, further demonstrating the role of NEAT1 as a miR-204
sponge.
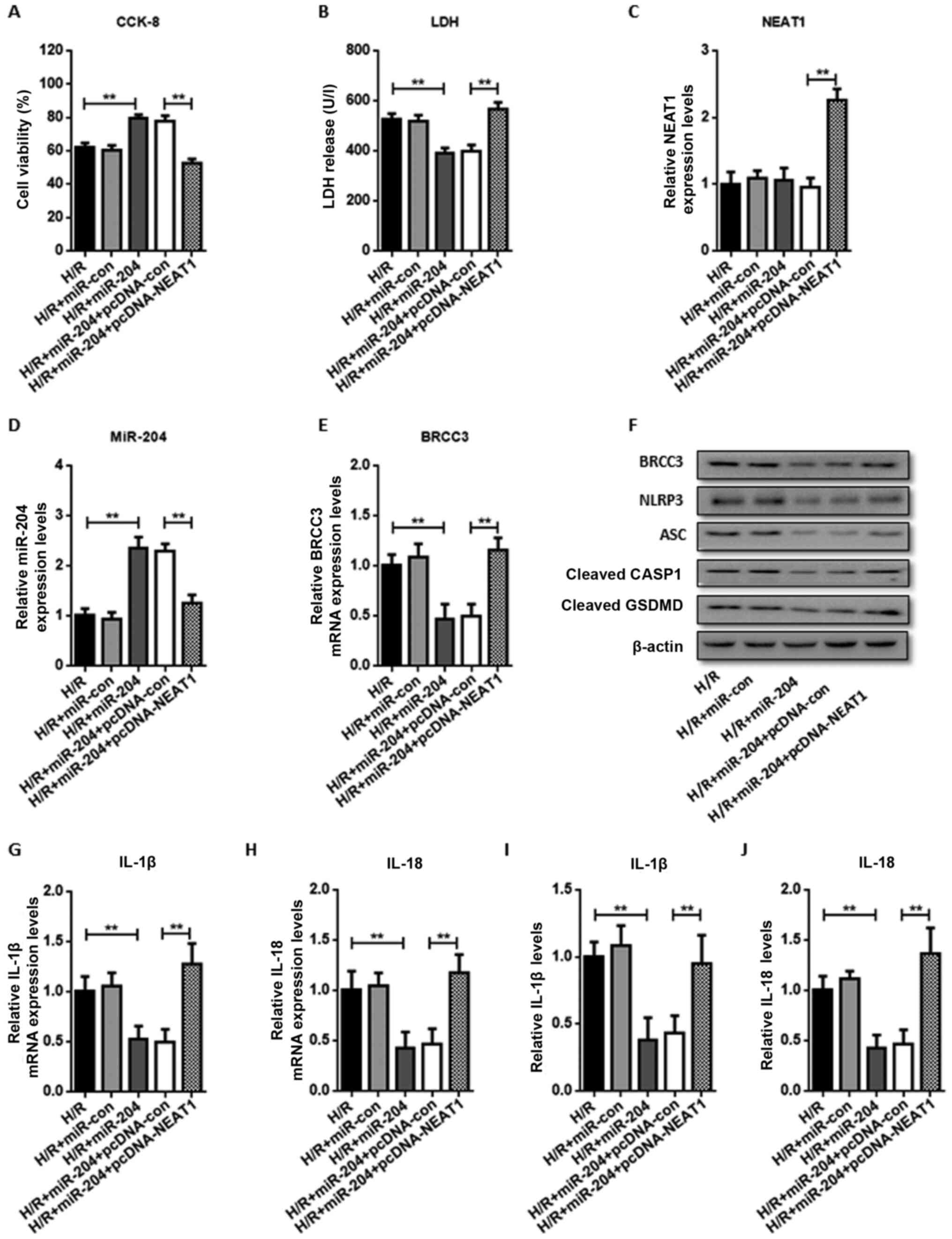 | Figure 5.NEAT1 aggravates H/R-induced cell
damage and NLRP3 inflammasome activation-dependent pyroptosis and
is rescued by miR-204 overexpression. HUVECs were co-transfected
with miR-204 or miR-con and pcDNA-NEAT1 or pcDNA-con before H/R.
Detection of (A) cell viability and (B) LDH activity in HUVECs
following H/R. RT-qPCR analysis of (C) NEAT1, (D) miR-204 and (E)
BRCC3 expression. (F) The expression levels of BRCC3, NLRP3, ASC,
cleaved CASP1 and cleaved GSDMD protein were examined by western
blotting. The expression levels of (G) IL-1β and (H) IL-18 mRNA
were examined by RT-qPCR. The levels of secreted (I) IL-1β and (J)
IL-18 were evaluated by ELISA. Data are shown as the mean ± SD from
five different experiments. **P<0.01. H/R,
hypoxia/reoxygenation; LDH, lactate dehydrogenase; IL, interleukin;
NLRP3, NOD-like receptor protein 3; ASC, apoptosis speck-like
protein; CASP1, caspase-1; GSDMD, gasdermin D; NEAT1, nuclear
enriched abundant transcript 1; BRCC3, BRCA1/BRCA2-containing
complex subunit 3; miR, microRNA; RT-qPCR, reverse
transcription-quantitative PCR; HUVEC, human umbilical vein
endothelial cell; CCK-8, Cell Counting Kit-8. |
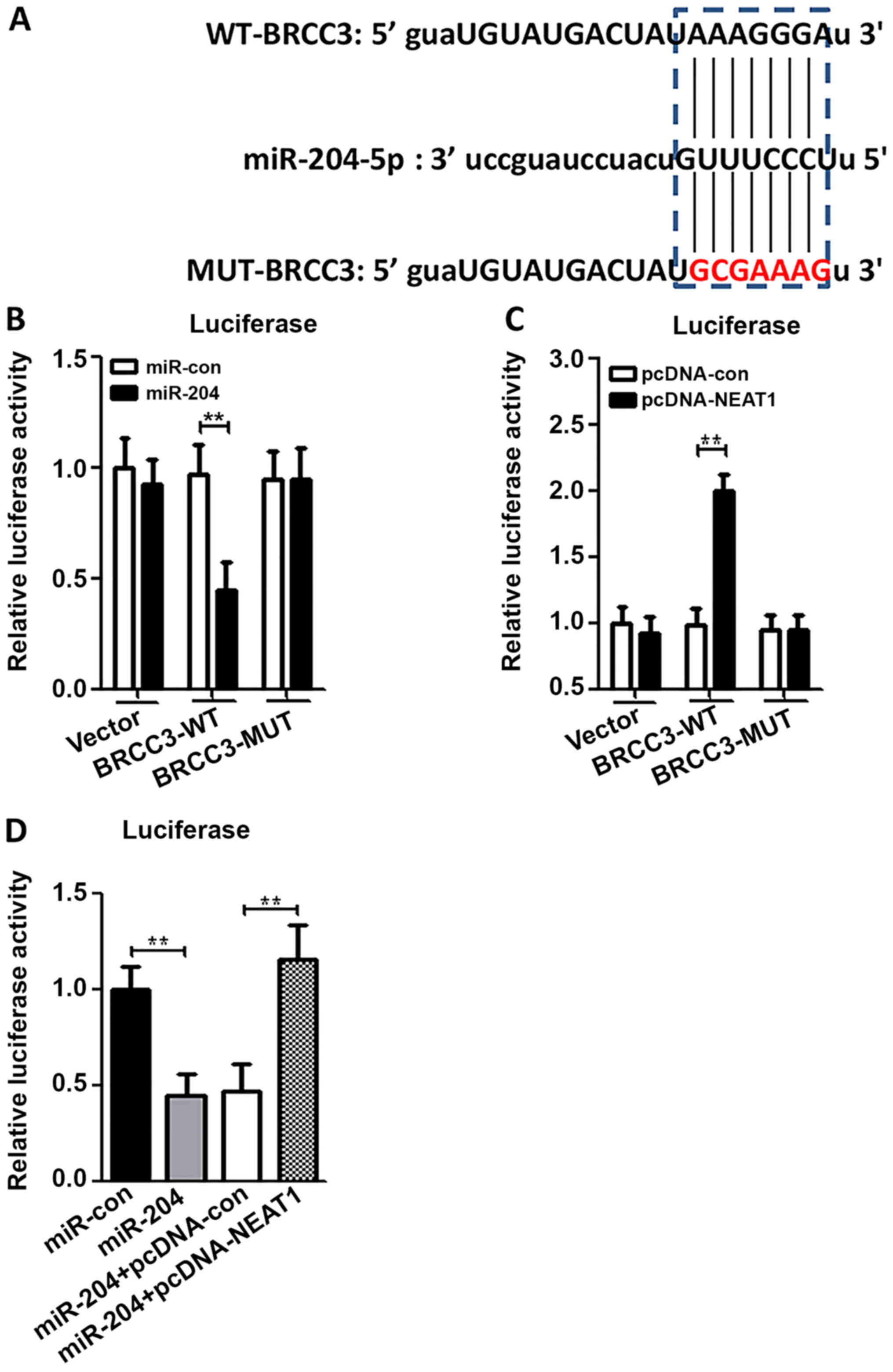
NEAT1 serves as a ceRNA for miR-204 to
target BRCC3
To ensure whether NEAT1 functioned as a ceRNA for
miR-204 to target BRCC3, luciferase reporter systems driven by
BRCC3-WT and BRCC3-MUT were constructed to verify predicted miR-204
binding sites (Fig. 6A). It was
found that miR-204 mimics significantly decreased the luciferase
activity of BRCC3-WT but not BRCC3-MUT in HUVECs. This effect was
reversed by co-transfection of pcDNA-NEAT1 and miR-204 mimics, but
not co-transfection of pcDNA-con and miR-204 mimics (Fig. 6B and C). In addition, the expression
of BRCC3 was also confirmed this tendency (Fig. 6D).
Discussion
Emerging evidence has suggested that lncRNAs can
function as ceRNAs that regulate specific RNA transcripts by
competing for shared miRNAs. In the present study, it was found
that NEAT1 and BRCC3 expression was significantly increased in
H/R-induced injury, whereas miR-204 expression was decreased.
Furthermore, it was demonstrated that NEAT1 positively regulated
BRCC3 expression and NLRP3 inflammasome activation-dependent
pyroptosis by competitively binding to miR-204. To the best of our
knowledge, the present study was the first to demonstrate that
NEAT1 and miR-204 regulated NLRP3 inflammasome activation-dependent
pyroptosis in H/R injury, and provides a potential mechanism and
novel therapeutic targets for cardiovascular I/R damage.
Recent studies have revealed that lncRNAs acts as a
ceRNA for miRNA to target mRNA to regulate cardiovascular disease
(12). Several lncRNAs, including
H19 (13), metastasis-associated
lung adenocarcinoma transcript 1 (14) and taurine-upregulated gene 1
(15) have been confirmed to
regulate cardiovascular I/R injury through the ceRNA network.
Moreover, previous studies partially support the results of the
present study. It was previously revealed that NEAT1 was
overexpressed in myocardial I/R injury cells compared with normal
myocardial cells, and downregulation of NEAT1 enhanced cell
proliferation while inhibiting cell apoptosis via targeting
miR-193a in I/R injury H9C2 cells (16). Luo et al (9) revealed that NEAT1 contributed to I/R
injury via the modulation of miR-495-3p and MAPK6 in H9C2 cells. In
addition, it was previously reported that NEAT1 contributed to
myocardial I/R injury by activating the MAPK pathway (7). All of these studies confirmed that
NEAT1 may deteriorate myocardium in I/R damage; however, the role
of NEAT1 in endothelial I/R injury and cardiovascular pyroptosis
has not been reported at present. In the present study, it was
reported that the regulation and potential mechanism of NEAT1 in
cardiovascular I/R injury was via the modulation of NLRP3
inflammasome activation-dependent pyroptosis, and could provide a
theoretical basis for the clinical treatment of I/R injury.
miRNA links upstream lncRNA with downstream mRNA
with a shared competitive binding site for lncRNA and mRNA. It was
found that upregulation of miR-204 alleviated ventricular
remodeling and improved cardiac function in mice following
myocardial I/R injury (12).
Furthermore, the function of miR-204 to protect against I/R injury
has also been confirmed to be applicable to the kidney (17) and diabetic myocardial I/R (18). However, there is currently no
research demonstrating a relationship between NEAT1 and miR-204,
and no studies have reported that miR-204 provides a shared
competitive binding site for NEAT1 and mRNA. Therefore, the present
study was the first to show that NEAT1 served as a ceRNA for
miR-204 to target BRCC3. In addition, it was reported that miR-204
played a key role in endothelial cell I/R injury and pyroptosis,
and elucidated its ability to regulate BRCC3 expression and
activity.
Conversely, other studies have demonstrated contrary
evidence to those presented in the present study. Yan et al
(19) found that NEAT1 functioned
as a miRNA sponge to inhibit cardiomyocyte apoptosis and may be a
novel therapeutic target for cardiomyocyte apoptosis associated
heart diseases. These findings are consistent with other studies
reporting that NEAT1 knockdown improves LPS-induced myocardial
injury in mice by inhibiting the toll-like receptor 2/NF-κB
signaling pathway (20). It was
previously reported that inhibition of miR-204 exerts spinal cord
protection against I/R injury, possibly via promotion of autophagy
and anti-apoptotic effects (21).
The reason for these differences is likely to be due to differences
in experimental models and differences in the ceRNA network
studied. This also suggests that NEAT1 is likely to play multiple
roles in cardiovascular I/R injury and can act as a double-edged
sword.
However, it should be noted that there are some
limitations to the research in the present study. Although the data
in the present study validated the key role and novel mechanism of
the NEAT1/miR-204/BRCC3 axis in the regulation of NLRP3
inflammasome activation dependent-pyroptosis in vitro, the
absence of in vivo animal experiments or human tissue
samples reduces the clinical application potential of the
conclusions. In addition, some other important forms of regulatory
cell death, including ferroptosis, apoptosis, autophagy and
necroptosis, have also been indicated to serve an important role in
myocardial I/R injury (22–26). However, the present study did not
focus on the effects of NEAT1/miR-204/BRCC3 signals on these forms
of regulatory cell death.
The present study focused on NLRP3 inflammasome
activation-dependent pyroptosis in cardiovascular I/R injury and
demonstrated that NEAT1 positively regulated BRCC3 expression and
NLRP3 inflammasome activation-dependent pyroptosis by competitively
binding with miR-204. Taken together, these findings demonstrated
that inhibition of lncRNA NEAT1 may protect HUVECs against
H/R-induced NLRP3 inflammasome activation by targeting the
miR-204/BRCC3 axis.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the Social Welfare
Project in Bao'an District, Shenzhen, China (grant no. 2020JD360 to
TY; grant no. 2021JD070 to SL).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TY and XY conceived and designed the project. TY
performed the experiments with the help of YS, SL and JG. YS
analyzed the data. TY wrote the manuscript. TY and XT confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen J, Jiang Z, Zhou X, Sun X, Cao J, Liu
Y and Wang X: Dexmedetomidine Preconditioning Protects
Cardiomyocytes Against Hypoxia/Reoxygenation-Induced Necroptosis by
Inhibiting HMGB1-Mediated Inflammation. Cardiovasc Drugs Ther.
33:45–54. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Burzynski LC and Clarke MCH: Death Is
Coming and the Clot Thickens, as Pyroptosis Feeds the Fire.
Immunity. 50:1339–1341. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
An N, Gao Y, Si Z, Zhang H, Wang L, Tian
C, Yuan M, Yang X, Li X, Shang H, et al: Regulatory Mechanisms of
the NLRP3 Inflammasome, a Novel Immune-Inflammatory Marker in
Cardiovascular Diseases. Front Immunol. 10:15922019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Py BF, Kim MS, Vakifahmetoglu-Norberg H
and Yuan J: Deubiquitination of NLRP3 by BRCC3 critically regulates
inflammasome activity. Mol Cell. 49:331–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Su Q, Liu Y, Lv XW, Ye ZL, Sun YH, Kong BH
and Qin ZB: Inhibition of lncRNA TUG1 upregulates miR-142-3p to
ameliorate myocardial injury during ischemia and reperfusion via
targeting HMGB1- and Rac1-induced autophagy. J Mol Cell Cardiol.
133:12–25. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu H, Xu D, Zhong X, Xu D, Chen G, Ge J
and Li H: lncRNA-mRNA competing endogenous RNA network depicts
transcriptional regulation in ischaemia reperfusion injury. J Cell
Mol Med. 23:2272–2276. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Du XJ, Wei J, Tian D, Yan C, Hu P, Wu X,
Yang W and Hu X: NEAT1 promotes myocardial ischemia-reperfusion
injury via activating the MAPK signaling pathway. J Cell Physiol.
234:18773–18780. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wei Z, Qiao S, Zhao J, Liu Y, Li Q, Wei Z,
Dai Q, Kang L and Xu B: miRNA-181a over-expression in mesenchymal
stem cell-derived exosomes influenced inflammatory response after
myocardial ischemia-reperfusion injury. Life Sci. 232:1166322019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo M, Sun Q, Zhao H, Tao J and Yan D:
Long noncoding RNA NEAT1 sponges miR-495-3p to enhance myocardial
ischemia-reperfusion injury via MAPK6 activation. J Cell Physiol.
235:105–113. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Deng F, Wang S, Cai S, Hu Z, Xu R, Wang J,
Feng D and Zhang L: Inhibition of Caveolae Contributes to Propofol
Preconditioning-Suppressed Microvesicles Release and Cell Injury by
Hypoxia-Reoxygenation. Oxid Med Cell Longev. 2017:35421492017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xue X and Luo L: lncRNA HIF1A-AS1
contributes to ventricular remodeling after myocardial
ischemia/reperfusion injury by adsorption of microRNA-204 to
regulating SOCS2 expression. Cell Cycle. 18:2465–2480. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo H, Wang J, Liu D, Zang S, Ma N, Zhao
L, Zhang L, Zhang X and Qiao C: The lncRNA H19/miR-675 axis
regulates myocardial ischemic and reperfusion injury by targeting
PPARα. Mol Immunol. 105:46–54. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu P, Yang M, Ren H, Shen G, Chen J,
Zhang J, Liu J and Sun C: Long noncoding RNA MALAT1 downregulates
cardiac transient outward potassium current by regulating
miR-200c/HMGB1 pathway. J Cell Biochem. 119:10239–10249. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang N, Xia J, Jiang B, Xu Y and Li Y:
TUG1 alleviates hypoxia injury by targeting miR-124 in H9c2 cells.
Biomed Pharmacother. 103:1669–1677. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ren L, Chen S, Liu W, Hou P, Sun W and Yan
H: Downregulation of long non-coding RNA nuclear enriched abundant
transcript 1 promotes cell proliferation and inhibits cell
apoptosis by targeting miR-193a in myocardial ischemia/reperfusion
injury. BMC Cardiovasc Disord. 19:1922019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu Y, Yin X, Li J and Zhang L:
Overexpression of microRNA-204-5p alleviates renal
ischemia-reperfusion injury in mice through blockage of Fas/FasL
pathway. Exp Cell Res. 381:208–214. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu SY, Dong B, Fang ZF, Hu XQ, Tang L and
Zhou SH: Knockdown of lncRNA AK139328 alleviates myocardial
ischaemia/reperfusion injury in diabetic mice via modulating
miR-204-3p and inhibiting autophagy. J Cell Mol Med. 22:4886–4898.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan H, Liang H, Liu L, Chen D and Zhang Q:
Long noncoding RNA NEAT1 sponges miR-125a-5p to suppress
cardiomyocyte apoptosis via BCL2L12. Mol Med Rep. 19:4468–4474.
2019.PubMed/NCBI
|
|
20
|
Wang SM, Liu GQ, Xian HB, Si JL, Qi SX and
Yu YP: lncRNA NEAT1 alleviates sepsis-induced myocardial injury by
regulating the TLR2/NF-κB signaling pathway. Eur Rev Med Pharmacol
Sci. 23:4898–4907. 2019.PubMed/NCBI
|
|
21
|
Yan L, Shi E, Jiang X, Shi J, Gao S and
Liu H: Inhibition of MicroRNA-204 Conducts Neuroprotection Against
Spinal Cord Ischemia. Ann Thorac Surg. 107:76–83. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li JY, Yao YM and Tian YP: Ferroptosis: A
Trigger of Proinflammatory State Progression to Immunogenicity in
Necroinflammatory Disease. Front Immunol. 12:7011632021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Deng F, Zhao BC, Yang X, Lin ZB, Sun QS,
Wang YF, Yan ZZ, Liu WF, Li C, et al: The gut microbiota metabolite
capsiate promotes Gpx4 expression by activating TRPV1 to inhibit
intestinal ischemia reperfusion-induced ferroptosis. Gut Microbes.
13:1–21. 2021. View Article : Google Scholar
|
|
24
|
Ye L, He S, Mao X, Zhang Y, Cai Y and Li
S: Effect of Hepatic Macrophage Polarization and Apoptosis on Liver
Ischemia and Reperfusion Injury During Liver Transplantation. Front
Immunol. 11:11932020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Martin PK and Cadwell K: Regulation of
interferon signaling in response to gut microbes by autophagy. Gut
Microbes. 11:126–134. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Deng Y, Wu S, Yang Y, Meng M, Chen X, Chen
S, Li L, Gao Y, Cai Y, Imani S, Chen B, Li S, Deng Y and Li X:
Unique Phenotypes of Heart Resident Type 2 Innate Lymphoid Cells.
Front Immunol. 11:8022020. View Article : Google Scholar : PubMed/NCBI
|