Introduction
Liver cancer is a serious public health problem
worldwide and is the fourth leading cause of cancer-related deaths
worldwide (1). Among the
different subtypes of liver cancer, hepatocellular carcinoma (HCC)
is the main cause of primary liver cancer (2). HCC accounts for ~80% of all liver
cancer cases (3). Despite the
improved therapeutic strategies for patients with liver cancer, the
5-year survival rates are far from the expected result due to the
high rates of recurrence and metastasis of liver cancer (4). Thus, it is important to explore the
underlying molecular mechanisms of liver cancer to improve the
survival rate of patients.
Long non-coding RNAs (lncRNAs) are a type of RNA.
Although they do not encode proteins, lncRNAs still play
significant roles in gene expression and cellular biological
processes (5,6). In previous years, it has been
reported that lncRNAs are involved in the progression of human
diseases (7). For example, in
lymph node metastasis of bladder cancer, lncRNA ELNAT1 has been
shown to promote UBC9 expression to catalyze SUMOylation,
lymphangiogenesis and lymph node metastasis of breast cancer
(8). lncRNA lnc030 has also been
reported to be highly expressed in breast cancer stem cells in
vitro and in vivo, and to cooperate with
poly(rC)-binding protein 2 to stabilize squalene epoxidase mRNA
(9). Furthermore, lncRNA CRNDE
may bind to the splicing protein SRSF6 to inhibit its function and
affect alternative splicing by mediating the autophagic flux in
chemoresistant gastric cancer (GC) cells (10). lncRNA MIR4435-2HG has been found
to be highly expressed and directly involved in the tumorigenesis
of several types of cancer, including gastric cancer, non-small
cell lung cancer and colorectal cancer (11–13). Notably, the progression and
metastasis of GC have been shown to be regulated by the lncRNA
MIR4435-2HG via the Wnt/β-catenin signaling pathway (12). Knockdown of MIR4435-2HG may also
suppress colorectal cancer cell proliferation, invasion and
migration via the miR-206/YAP1 axis (11). In addition, MIR4435-2HG expression
has been reported to be increased in liver cancer (14,15). However, the functions of
MIR4435-2HG in liver cancer require further investigation.
The present study aimed to detect the expression
levels of MIR4435-2HG in liver cancer and to explore the molecular
mechanism underlying the regulatory effect of MIR4435-2HG on the
progress of liver cancer.
Materials and methods
Clinical specimens
Liver cancer tissues and adjacent normal tissues
were collected from patients with liver cancer (n=40) at the Second
Hospital of Tianjin Medical University (Tianjin, China) and were
stored in liquid nitrogen until further analysis. Tissue samples
were collected between January 2017 and December 2019. Of the 40
patients, 18 were women and 22 were men; 17 patients were <50
years old and 23 patients were ≥50 years old; the mean age of all
patients was 54 years old. None of the patients received
radiotherapy or chemotherapy before surgical resection. The
inclusion criteria were as follows: i) Patients diagnosed with
primary liver cancer; and ii) knew the purpose of the research and
provided written informed consent. The exclusion criteria were as
follows: i) Patients diagnosed with other tumors or liver
insufficiency; ii) patients had mental illness; and iii) patients
also had other types of cancer. The present study was approved by
the Ethics Committee of the Second Hospital of Tianjin Medical
University (approval no. KY2020K093). All patients provided written
informed consent prior to the study.
Cell culture and transfection
The normal liver cell line THLE-2, and human liver
cancer cell lines HCCLM3 and HepG2 were purchased from the American
Type Culture Collection. The cells were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin in an
incubator at 37°C with 5% CO2. All cell lines were
authenticated using short tandem repeat profiling.
For cell transfection assays, scrambled shRNA
negative control (shNC), two shMIR4435-2HG (sh2HG-1 and sh2HG-2),
empty pcDNA vector (pcDNA3), pcDNA MIR4435-2HG (MIR4435-2HG), pcDNA
B3GNT5 (B3GNT5), miR-NC mimics, miR-NC inhibitor, miR-136-5p mimics
and miR-136-5p inhibitor were constructed and purchased from
Shanghai GeneChem Co., Ltd. The sequences were as follows: miR-NC
mimics, 5′-UUCUCCGAACGUGUCACGUTT-3′; miR-136-5p mimics,
5′-ACUCCAUUUGUUUUGAUGAUGGA-3′; miR-NC inhibitor,
5′-CAGUACUUUUGUGUAGUACAA-3′; miR-136-5p inhibitor,
UCCAUCAUCAAAACAAAUGGAGU-3′. HCCLM3 and HepG2 cells at 70%
confluence were transfected with 1 µg shRNA and plasmids, or miRNA
mimics/inhibitor at 37°C using Lipofectamine® 3000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). After 48 h,
the cells were harvested for other assays.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA, including miRNA, was extracted from
tissues and cells using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). cDNA was then synthesized from
mRNA using the MMLV Reverse kit (Promega Corporation) according to
the manufacturer's instructions. RT reagents used for miRNA (cat.
no. D1801) were provided by HaiGene Co., Ltd and RT was performed
according to manufacturer's protocol. Gene expression was detected
by qPCR using SYBR® Premix EX Taq™ kit (Takara Bio,
Inc.). The thermocycling conditions were as follows: i) 95°C for 5
min; ii) 40 cycles at 95°C for 10 sec, 60°C for 30 sec and 72°C for
30 sec. The expression levels of MIR4435-2HG and B3GNT5 were
normalized to those of GAPDH, whereas U6 served as the internal
control for miR-136-5p. The relative expression levels of the genes
were calculated using the 2−ΔΔCq method (16). The primer sequences used in the
present study are as follows: MIR4435-2HG, forward,
5′-CTGGATGGTCGCTGCTTTTTA-3′ and reverse 5′-AGGGGGATGAGTCGTGATTT-3′;
miR-136-5p, forward 5′-ACTCCATTTGTTTTGATGATGGA-3′ and reverse
5′-TGGTGTCGTGGAGTCG-3′; B3GNT5, forward 5′-GGGCCTCGCTACCAATACTTG-3′
and reverse 5′-CGGAACGTCGATCATAGTTTTCA-3′; GAPDH, forward
5′-AATGGACAACTGGTCGTGGAC-3′ and reverse 5′-CCCTCCAGGGGATCTGTTTG-3′;
and U6, forward 5′-AGTAAGCCCTTGCTGTCAGTG-3′ and reverse
5′-CCTGGGTCTGATAATGCTGGG.
Western blotting
Proteins were extracted using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology). Protein concentrations were quantified using a BCA
Protein Assay Kit (Beyotime Institute of Biotechnology).
Subsequently, the proteins (30 µg) were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis on 10% gels and
transferred to a polyvinylidene difluoride membrane. The membranes
were blocked with 5% non-fat milk for 1 h at room temperature and
incubated with the corresponding primary antibodies: Anti-B3GNT5
(1:1,000; cat. no. PA5-26653; Thermo Fisher Scientific, Inc.) and
anti-GAPDH (1:5,000; cat. no. ab8245; Abcam) at 4°C overnight.
After washing the membrane with Tris-buffered saline with 0.1%
Tween-20, the membranes were incubated with the corresponding
horseradish peroxidase-conjugated secondary antibodies: Goat
anti-rabbit secondary antibody (1:3,000; cat. no. 7074; Cell
Signaling Technology, Inc.) and horse anti-mouse (1:3,000; cat. no.
7076; Cell Signaling Technology, Inc.) at room temperature for 1 h.
Protein bands were imaged using an Enhanced Chemiluminescence Kit
(Beyotime Institute of Biotechnology). The intensity of the protein
bands was analyzed using ImageJ 1.51 software (National Institutes
of Health).
Cell Counting Kit-8 (CCK-8) assay
The CCK-8 kit (Dojindo Molecular Technologies, Inc.)
was used to measure the prolferation ability of transfected liver
cancer cells. Transfected cells (5×103/well) were seeded
into 96-well plates and incubated for the indicated time periods
(24, 48 and 72 h) at 37°C. After incubation, 10 µl CCK-8 solution
was added to the corresponding wells for 2 h at 37°C. The optical
density values at 450 nm were measured using a microplate reader.
Cell proliferation ability was calculated according to the
manufacturer's instructions.
Wound healing assay
HCCLM3 and HepG2 cells were seeded in six-well
plates. The wound was made when the cells reached >90%
confluence and the scratched cells were cultured in serum-free
medium. The width of the scratch was imaged at 0 and 24 h using an
inverted light microscope. The relative mobility of the cells was
then calculated using ImageJ 1.51 software.
Transwell assay
Transwell assays were conducted to assess cell
migration and invasion. For the migration assay, the upper chambers
of the Transwell inserts were not coated with Matrigel, whereas in
the invasion assay the upper chambers were coated with Matrigel at
37°C for 7 h. HCCLM3 and HepG2 cells (1×105) were added
into the upper chambers of Transwell inserts in 200 µl medium. The
bottom chambers of the Transwell inserts were filled with 600 µl
medium supplemented with 10% FBS. After 48 h, the cells in the
upper chambers were fixed with 4% paraformaldehyde for 10 min at
room temperature and stained with 0.4% crystal violet for 10 min at
room temperature. Images of the migrated or invaded cells were
captured and cells were counted in five random fields using an
inverted light microscope.
Dual-luciferase reporter assay
The binding sites between genes were identified
using bioinformatics software starBase3.0 (http://starbase.sysu.edu.cn). The wild-type (WT) or
mutant (MUT) 3′-UTRs of MIR4435-2HG or B3GNT5 were cloned into the
pGL3-luciferase vector (Promega Corporation). Briefly, to
synthesize MUT sequences, the nucleotide sequences of MIR4435-2HG
or B3GNT5, which exhibit complementary binding sequences to
miR-136-5p, were changed by replacing the original base sequences
so they were no longer complementary to miR-136-5p. Subsequently,
miR-NC, miR-136-5p mimic, WT-MIR4435-2HG, MUT-MIR4435-2HG,
WT-B3GNT5 or MUT-B3GNT5 were co-transfected into the indicated
groups of 293T cells (American Type Culture Collection) with the
indicated groups using Lipofectamine 3000. The groups were as
follows: WT-MIR4435-2HG + miR-NC mimics, WT-MIR4435-2HG +
miR-136-5p mimics, MUT-MIR4435-2HG + miR-NC mimics, MUT-MIR4435-2HG
+ miR-136-5p mimics, WT-B3GNT5 + miR-NC mimics, WT-B3GNT5 +
miR-136-5p mimics, MUT-B3GNT5 + miR-NC mimics and MUT-B3GNT5 +
miR-136-5p mimics). After 48 h, luciferase activity was measured
using a dual-luciferase reporter assay system (Promega Corporation)
according to the manufacturer's instructions. Luciferase activity
was normalized to Renilla luciferase activity.
Bioinformatics analysis
MIR4435-2HG expression in The Cancer Genome Atlas
(TCGA) was analyzed using Gene Set Cancer Analysis (GSCA;
http://bioinfo.life.hust.edu.cn/GSCA/#/) in all cancer
datasets, Gene Expression Profiling Interactive Analysis 2 (GEPIA2;
http://gepia.cancer-pku.cn/) in the
liver hepatocellular carcinoma (LIHC) dataset and DeepBase 3.0
(http://rna.sysu.edu.cn/deepbase3/index.html) in the
LIHC dataset. The relationship between MIR4435-2HG expression and
cancer metastasis was studied using the Human Cancer Metastasis
Database (http://hcmdb.i-sanger.com/index). The relationship
between MIR4435-2HG expression and prognosis of patients with liver
cancer was analyzed using GEPIA2. Potential target miRNAs of
MIR4435-2HG were predicted using the starBase 3.0 website
(http://starbase.sysu.edu.cn). Target
genes of miR-136-5p were predicted by starBase 3.0, miRWalk 3.0
(http://mirwalk.umm.uni-heidelberg.de/), TargetScan 7.1
(http://www.targetscan.org/vert_71/),
miRDB 6.0 (http://mirdb.org/) and ONCOMIR
(http://www.oncomir.org/oncomir/index.html) databases.
The expression of B3GNT5 and its effects on the prognosis of
patients with liver cancer were analyzed using the GEPIA2 database.
Heat map analysis of the expression of targets of miR-136-5p was
performed using the UALCAN database (http://ualcan.path.uab.edu/). The variation in copy
number, including single nucleotide variation (SNV) and copy number
variation (CNV), and methylation of targets of miR-136-5p were
analyzed using the GSCA database; SNV and CNV of genes were
obtained by selecting different commands in the GSCA database. The
correlation between MIR4435-2HG and B3GNT5 expression was
determined using the GEPIA database.
Statistical analysis
Data are shown as the mean ± standard deviation and
were analyzed using SPSS 19.0 (IBM Corp.) and GraphPad Prism 8.0
(GraphPad Software, Inc.). Differences between normal and tumor
tissues were analyzed using a paired t-test. Other differences
between two independent groups were analyzed using an unpaired
t-test. Differences between multiple groups were analyzed using
one-way ANOVA followed by Tukey's post hoc test. The survival
analysis was performed using the Kaplan-Meier method followed by a
log-rank test in the GEPIA database. Linear regression analysis was
used to detect the association between MIR4435-2HG and miR-136-5p.
Pearson's correlation analysis was used to detect the correlation
between MIR4435-2HG and B3GNT5 gene expression levels. Spearman's
correlation analysis was used to analyze the correlation between
B3GNT5 methylation and expression at the mRNA level. P<0.05 was
considered to indicate a statistically significant difference.
Results
MIR4435-2HG is upregulated in liver
cancer tissue in TCGA database
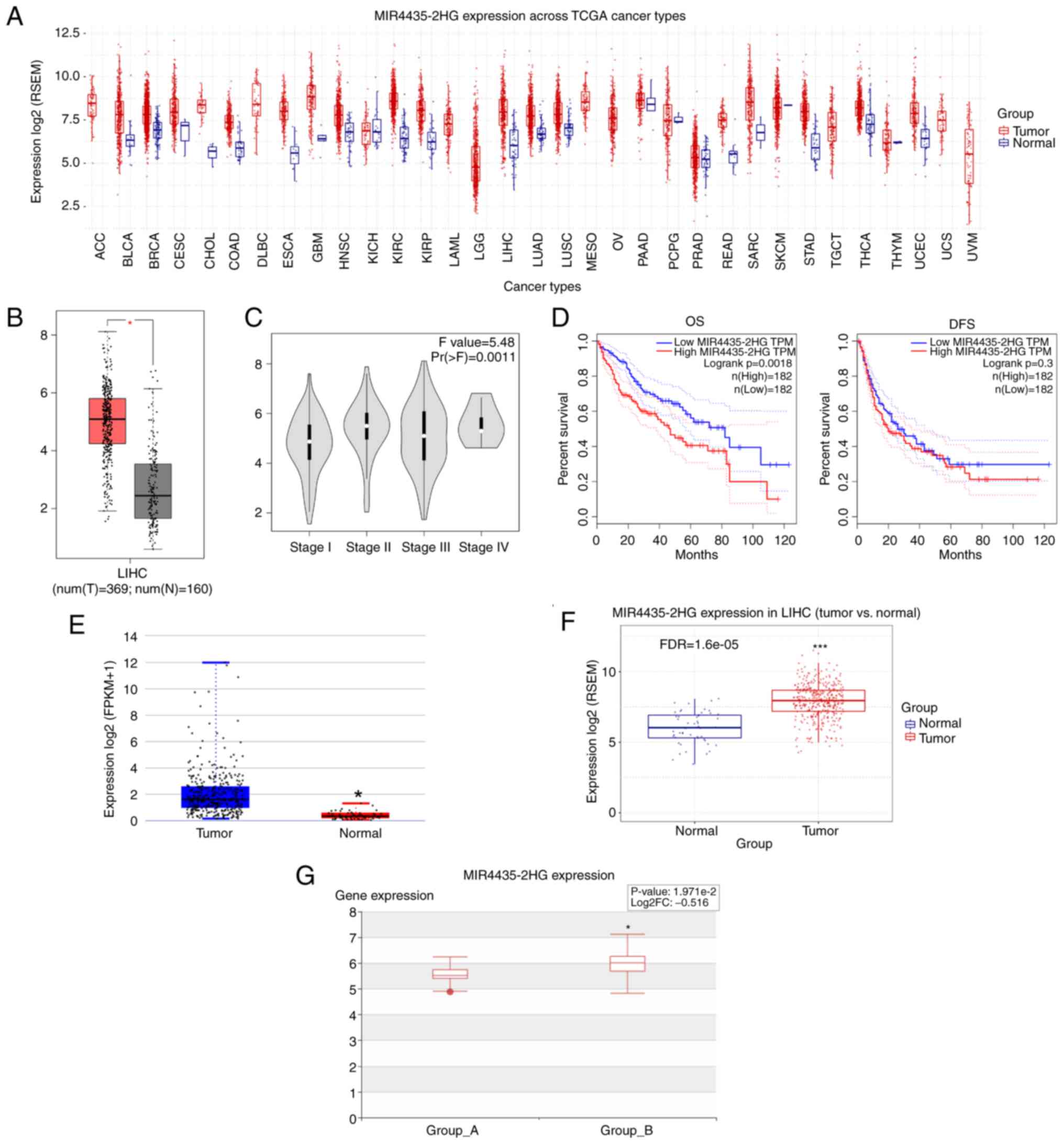
First, MIR4435-2HG expression across TCGA cancer
types was analyzed using the GSCA database. The results indicated
that the expression levels of MIR4435-2HG were significantly
upregulated in a various types of cancer, including liver cancer
(Fig. 1A). TCGA data in the GEPIA
database further demonstrated that the expression levels of
MIR4435-2HG were increased in liver cancer tissues compared with
those in adjacent normal tissues (Fig. 1B). Moreover, increased MIR4435-2HG
expression was associated with liver cancer progression (Fig. 1C). Kaplan-Meier analysis showed
that patients with liver cancer with higher MIR4435-2HG expression
had poor overall survival (OS), but it had no obvious effect on
disease-free survival (DFS) (Fig.
1D). Furthermore, MIR4435-2HG expression was found to be
increased in patients with liver cancer in the DeepBase and GSCA
databases, respectively (Fig. 1E and
F). Of note, in patients with liver cancer, the expression of
MIR4435-2HG was further increased with the occurrence of metastasis
(Fig. 1G). These data suggested
that MIR4435-2HG may be a pivotal oncogene in liver cancer.
MIR4435-2HG knockdown inhibits the
malignant behavior of liver cancer cells
In the present study, the expression levels of
MIR4435-2HG in liver cancer tissues and cell lines were measured by
RT-qPCR. The results revealed that MIR4435-2HG expression levels
were significantly upregulated in liver cancer tissues and cell
lines compared with those in normal tissues and cell lines
(Fig. 2A and B). Subsequently,
the role of MIR4435-2HG was investigated in liver cancer cells.
Liver cancer cells were transfected with shNC, shMIR4435-2HG
(sh2HG-1 and sh2HG-2), pcDNA3 or MIR4435-2HG. It was revealed that
the expression levels of MIR4435-2HG were significantly affected by
sh2HG-1, sh2HG-2 and MIR4435-2HG (Fig. 2C and D). Since sh2HG-1 produced a
more obvious inhibitory effect on MIR4435-2HG, sh2HG-1 was used in
subsequent experiments. In addition, proliferation was measured
using the CCK-8 assay. The results revealed that sh2HG-1 resulted
in decreased proliferation, whereas overexpression of MIR4435-2HG
in cancer cells exerted the opposite effect (Fig. 2E). Wound healing and Transwell
assays were performed to assess cell migration and invasion
capacities, and revealed that sh2HG-1 reduced the migration and
invasion abilities of liver cancer cells (Fig. 2F-H). However, in liver cancer
cells transfected with MIR4435-2HG, migration and invasion were
significantly potentiated (Fig.
2F-H). These results suggested that the silencing of
MIR4435-2HG may reduce the malignant biological behavior of liver
cancer cells by influencing cell proliferation, migration and
invasion.
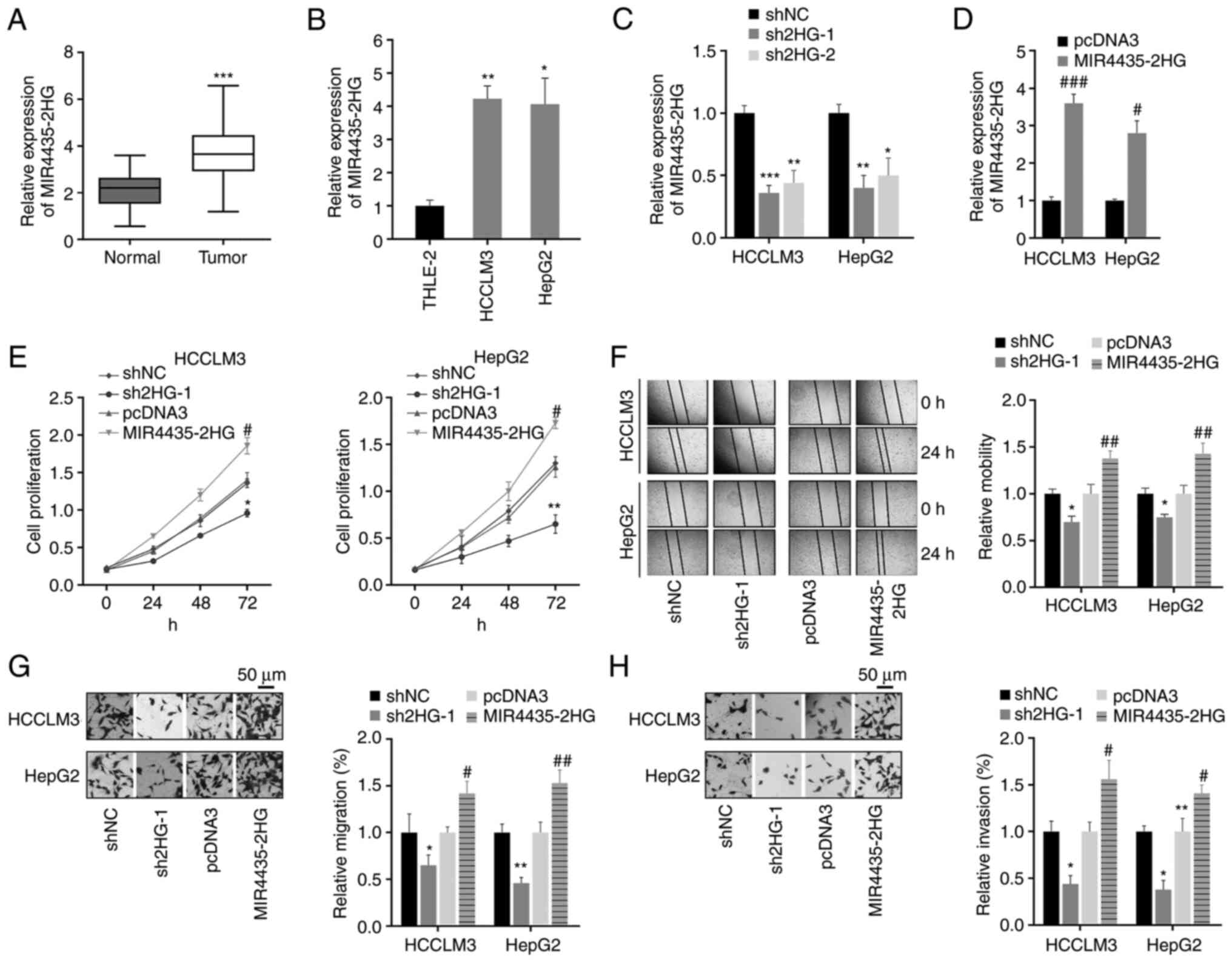 | Figure 2.Knockdown of MIR4435-2HG inhibits the
proliferation, migration and invasion of liver cancer cells. (A)
MIR4435-2HG expression was detected in liver cancer and normal
liver tissues via RT-qPCR. ***P<0.001 vs. normal tissues. (B)
MIR4435-2HG expression in normal liver cells and liver cancer cell
lines. *P<0.05, **P<0.01 vs. THLE-2 cells. (C and D) After
transfection with sh2HG-1, sh2HG-2, MIR4435-2HG or corresponding
NCs, the expression levels of MIR4435-2HG were examined using
RT-qPCR. *P<0.05, **P<0.01, ***P<0.001 vs. shNC,
#P<0.05, ###P<0.001 vs. pcDNA3. (E)
Proliferation was measured using the CCK-8 assay in liver cancer
cells with indicated transfections. (F) Wound healing assay was
used to assess cell migration (magnification, ×50). (G and H) The
effects of MIR4435-2HG on cell migration and invasion were measured
by Transwell assays (magnification, ×400). *P<0.05, **P<0.01
vs. shNC, #P<0.05, ##P<0.01 vs. pcDNA3.
RT-qPCR, reverse transcription-quantitative PCR; sh, short hairpin
RNA; sh2HG, shMIR4435-2HG; NC, negative control. |
miR-136-5p is a target of
MIR4435-2HG
To explore the potential molecular mechanisms of
MIR4435-2HG in the regulation of liver cancer progression, its
target genes were predicted using bioinformatic tools. By analyzing
previous research reports (17–20), it was found that miR-136-5p
(identified as a target of MIR4435-2HG using bioinformatics
analysis) expression was significantly decreased in various types
of cancer, indicating that miR-136-5p may act as a tumor
suppressor. However, the role of miR-136-5p in the liver has rarely
been studied. Therefore, miR-136-5p was chose as a potential target
of MIR4435-2HG for further analysis; the potential binding site of
miR-136-5p on MIR4435-2HG is shown in Fig. 3A. First, the expression levels of
miR-136-5p in liver cancer tissues were detected; it was revealed
that miR-136-5p expression was significantly downregulated in liver
cancer tissues (Fig. 3B). To
confirm the direct binding between miR-136-5p and MIR4435-2HG, 293T
cells were co-transfected with miR-136-5p mimics and
WT/MUT-MIR4435-2HG luciferase reporter plasmids. The luciferase
reporter assay showed that miR-136-5p significantly suppressed the
luciferase activity of cells transfected with WT-MIR4435-2HG, but
not MUT-MIR4435-2HG (Fig. 3C).
These findings indicated that MIR4435-2HG can target and sponge
miR-136-5p. Cancer cells were transfected with miR-136-5p mimics or
inhibitor before functional analysis of miR-136-5p (Fig. S1A and B). It was found that
transfection of cells with miR-136-5p mimics significantly reduced
the expression of MIR4435-2HG, whereas transfection with a
miR-136-5p inhibitor significantly upregulated MIR4435-2HG
expression (Fig. 3D). Similarly,
the expression of miR-136-5p in liver cancer cells was affected by
the overexpression of MIR4435-2HG (Fig. 3E); this may be due to the fact
that MIR4435-2HG can competitively bind to miR-136-5p, which leads
to the downregulation of miR-136-5p. Furthermore, linear regression
analysis validated the moderate negative association between
MIR4435-2HG and miR-136-5p expression in liver cancer tissues
(Fig. 3F). Functionally, the
inhibition of cell proliferation, migration and invasion via
MIR4435-2HG knockdown were partially restored by the miR-136-5p
inhibitor (Fig. 3G-J). In
general, these data indicated that MIR4435-2HG affected the
proliferation, migration and invasion of liver cancer cells by
negatively regulating miR-136-5p expression.
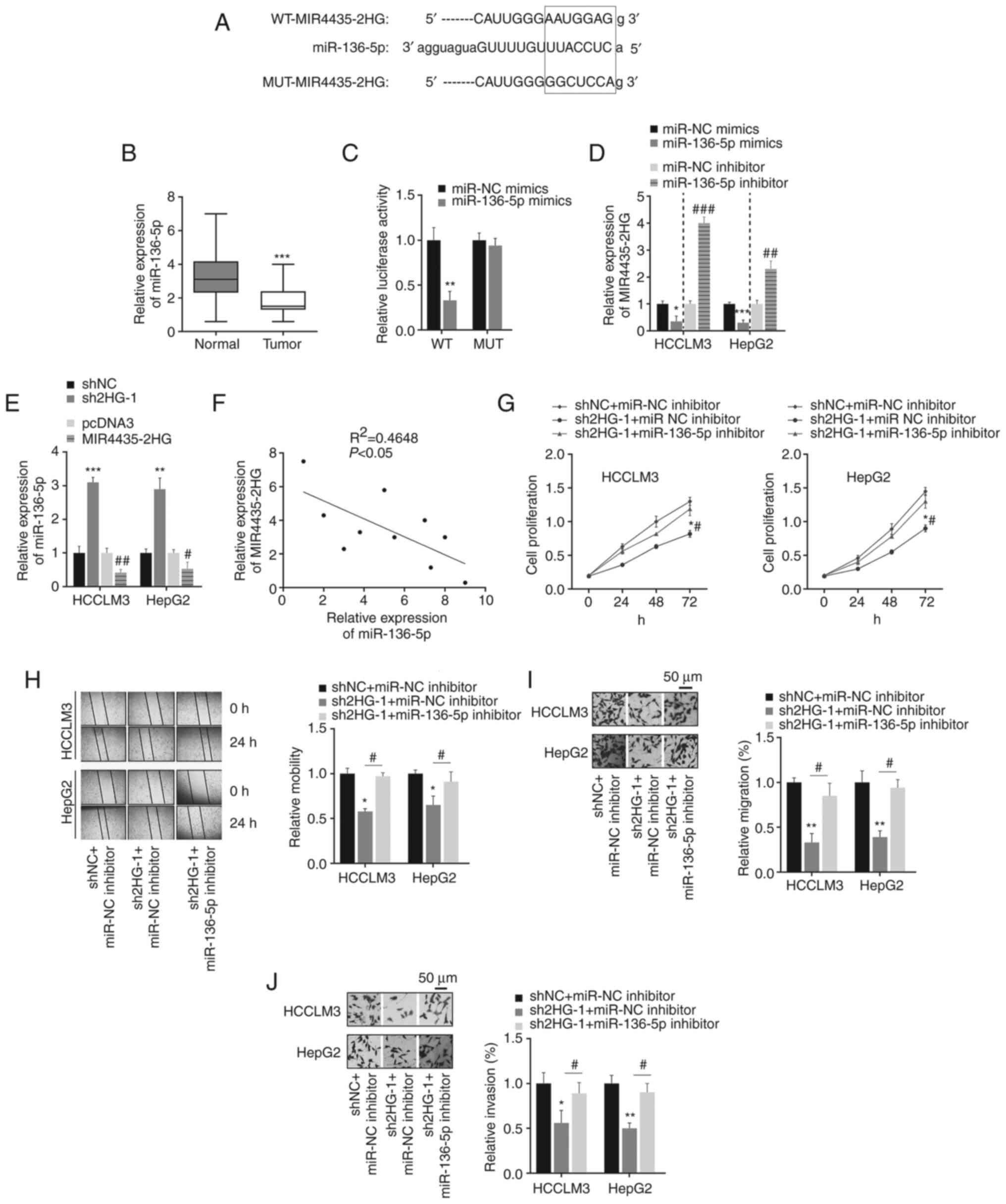 | Figure 3.MIR4435-2HG targets miR-136-5p. (A)
The WT or MUT binding sites of MIR4435-2HG on miR-136-5p. (B)
miR-136-5p expression in liver cancer and normal tissues.
***P<0.001 vs. normal tissues. (C) Dual-luciferase reporter
assay indicated that the luciferase activities were decreased after
co-transfection of WT-MIR4435-2HG and miR-136-5p mimics.
**P<0.01 vs. miRNA NC mimics. (D) The effect of miR-136-5p on
MIR4435-2HG expression. *P<0.05, ***P<0.001 vs. miRNA NC
mimics; ##P<0.05, ###P<0.001 vs. miRNA
NC inhibitor. (E) MIR4435-2HG regulated miR-136-5p expression in
liver cancer cells. **P<0.01, ***P<0.001 vs. shNC;
#P<0.05, ##P<0.01 vs. pcDNA3. (F)
Negative association between MIR4435-2HG and miR-136-5p expression
in liver cancer tissues. (G) Transfection with sh2HG inhibited cell
proliferation, which was rescued by miR-136-5p inhibitor. (H-J)
Transfection with sh2HG affected cell migration and invasion by
targeting miR-136-5p. Magnification, ×50. *P<0.05, **P<0.01
vs. shNC + miR-NC inhibitor; #P<0.05 vs. sh2HG-1 +
miR-136-5p inhibitor. miR/miRNA, microRNA; WT, wild-type; MUT,
mutant; NC, negative control; sh, short hairpin RNA sh2HG,
shMIR4435-2HG. |
Expression analysis of the potential
miR-136-5p target B3GNT5 in liver cancer
Next, the molecular mechanism of miR-136-5p in liver
cancer was further revealed by predicting its downstream targets
using bioinformatics tools (starBase, miRWalk, TargetScan, ONCOMIR
and miRDB; Fig. 4A). Among these
targets, gene sets that existed in different databases and had no
more than 10 target genes were selected for heatmap analysis of
expression using the UALCAN database (data not shown). It was found
that seven genes (PBX3, B3GNT5, FZD4, DRAM1, PTPDC1, SFXN3, MLLT3)
had the highest expression in liver cancer; thus, the seven genes
were selected for further analysis. Next, the higher expression
levels of seven target genes were found in liver cancer tissues
compared with those in normal tissues from patients with liver
cancer using the UALCAN database (Fig. 4B), which indicated that the higher
expression levels of the seven target genes may be related to liver
cancer. To the best of our knowledge, the function of B3GNT5 has
not yet been reported in liver cancer; therefore, the present study
focused on B3GNT5 and explored its function in liver cancer. First,
the expression of B3GNT5 was determined using the GEPIA database
and it was revealed to be highly expressed in liver cancer tissues
compared with in adjacent normal tissues (Fig. 4C). Patients with higher B3GNT5
expression levels had shorter OS and DFS (Fig. 4D). Furthermore, the SNV of six of
the seven targets was determined; notably, the SNV of DRAM1 is not
currently included in the GSCA database. The results revealed that
four of the six genes (PTPDC1, MLLT3, FZD4 and B3GNT5) exhibited
SNV in liver cancer (Fig. 4E). A
missense mutation of B3GNT5 was observed in liver cancer (Fig. 4F). In addition, CNV was analyzed;
the results showed that B3GNT5 also exhibited CNV in liver cancer
(Fig. 4G). In most cases, the
cancer progression is accompanied by changes in gene methylation
levels (21–24). By analyzing the GSCA database, the
B3GNT5 methylation levels across TCGA cancer types are presented in
Fig. 4H. Methylation levels of
the five of the seven targets were decreased and negatively
associated with mRNA expression; methylation differences of FZD4
and SFXN3 in LIHC are not currently included in the GSCA database
(Fig. 4I and J). Our previous
results revealed that the expression levels of these target genes
were elevated in liver cancer, and the decrease in methylation
levels may be conducive to the increase in their expression levels.
These findings indicated that these target genes may be mutated in
liver cancer and their expression levels may also be increased in
liver cancer. Furthermore, Spearman's correlation analysis
indicated a negative correlation between B3GNT5 methylation and
expression at the mRNA level (Fig.
4K), which may indicate that the decrease in B3GNT5 methylation
contributes to the expression of B3GNT5 in liver cancer. These
results showed that B3GNT5 expression was increased in liver
cancer. Therefore, B3GNT5 may act as an oncogene in liver
cancer.
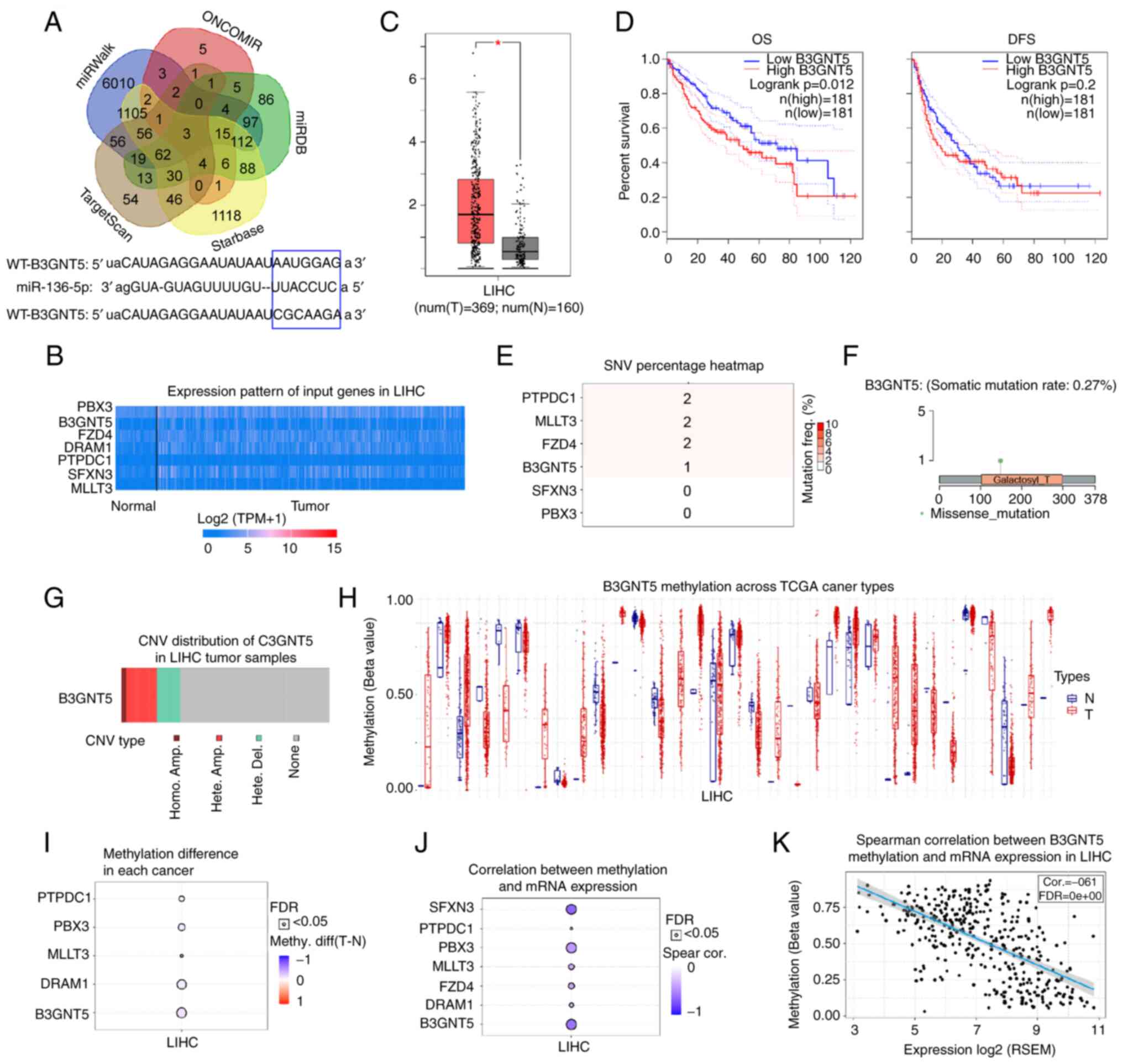 | Figure 4.Prediction and analysis of potential
target genes of miR-136-5p. (A) Target genes of miR-136-5p were
predicted using bioinformatics tools (starBase, miRWalk,
TargetScan, ONCOMIR and miRDB). (B) Expression profile of seven
target genes of miR-136-5p was analyzed using the UALCAN database.
(C) Increased B3GNT5 expression was detected in LIHC in the GEPIA
database. (D) OS and DFS of patients with liver cancer with
different expression profiles of B3GNT5. GSCA database was used to
study the SNV, copy number alteration and methylation levels of
target genes. (E) SNV percentage of target genes in liver cancer.
(F) Missense mutation of B3GNT5 in liver cancer. (G) CNV types of
B3GNT5 in liver cancer. (H) B3GNT5 methylation levels across TCGA
cancer types. (I) Methylation difference of target genes in liver
cancer. (J) Correlation between methylation and mRNA expression
levels of target genes of miR-136-5p. (K) Negative correlation
between B3GNT5 methylation level and mRNA expression. *P<0.05.
miR, microRNA; GEPIA, Gene Expression Profiling Interactive
Analysis; OS, overall survival; DFS, disease-free survival; SNV,
single nucleotide variation; CNV, copy number variation; TCGA, The
Cancer Genome Atlas; WT, wild-type; MUT, mutant; FDR, false
discovery rate; LIHC, liver hepatocellular carcinoma. |
MIR4435-2HG promotes B3GNT5 expression
by regulating miR-136-5p
In general, miRNAs exercise their role by targeting
mRNA and inhibiting protein activity (25,26). Thus, the association between
B3GNT5 and miR-136-5p was confirmed using a dual-luciferase
reporter assay (Fig. 5A). The
mRNA and protein expression levels of B3GNT5 were detected using
RT-qPCR and western blotting, respectively. It was found that
B3GNT5 expression was abnormally increased in liver cancer tissues
and cells (Figs. 5B-E and
S1C and D). Furthermore, western
blot analysis suggested that MIR4435-2HG positively regulated
B3GNT5 expression by inhibiting miR-136-5p in liver cancer cells
(Fig. 5F and G). To further
verify that MIR4435-2HG promotes liver cancer progression by
mediating miR-136-5p and B3GNT5 expression, cancer cells were
transfected with sh2HG-1 and B3GNT5. B3GNT5 overexpression was
confirmed, as presented in Fig.
5H. It was found that MIR4435-2HG knockdown significantly
inhibited cell proliferation; however, the overexpression of B3GNT5
partially rescued the effect of MIR4435-2HG knockdown (Fig. 5I). The decreased cell migration
and invasion abilities caused by sh2HG-1 were also reversed by the
overexpression of B3GNT5 (Fig.
5J-L). Consistent with the aforementioned results, it was found
that the expression of MIR4435-2HG was slightly positively
correlated with that of B3GNT5 in liver cancer specimens (Fig. 5M). Thus, these results suggested a
critical role of the MIR4435-2HG/miR-136-5p/B3GNT5
axis in liver cancer.
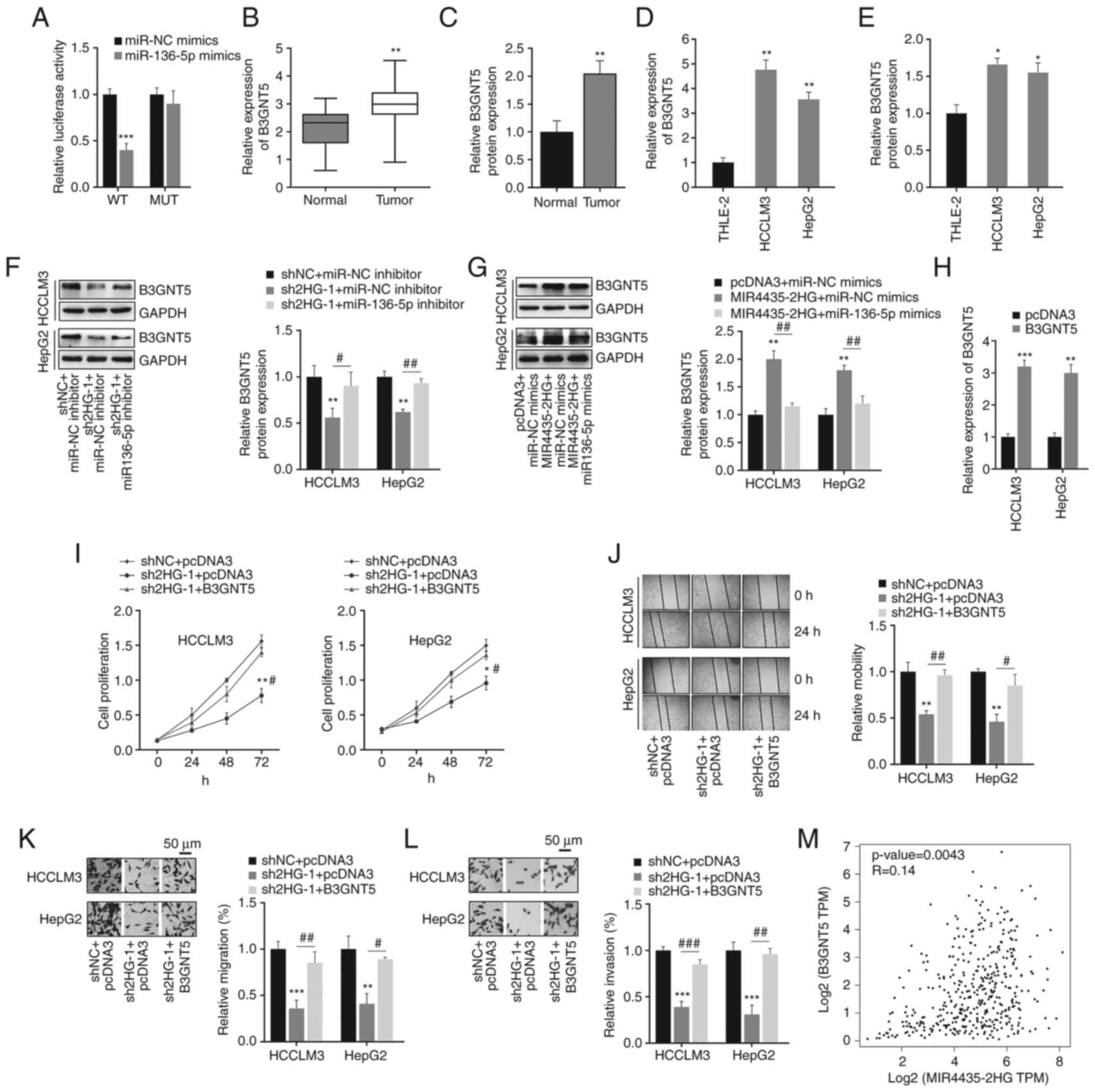 | Figure 5.MIR4435-2HG promotes B3GNT5
expression in liver cancer cells. (A) Dual-luciferase reporter
assay was used to detect the binding between B3GNT5 and miR-136-5p.
***P<0.001 vs. miRNA NC. (B and C) mRNA and protein expression
levels of B3GNT5 in liver cancer and adjacent normal tissues.
**P<0.01 vs. normal tissues. (D and E) mRNA and protein
expression levels of B3GNT5 in normal liver cell line and liver
cancer cell lines. *P<0.05, **P<0.01 vs. THLE-2 cells. (F)
MIR4435-2HG regulated B3GNT5 protein expression by targeting
miR-136-5p, as shown following transfection with sh2HG-1 and
miR-136-5p inhibitor. **P<0.01 vs. shNC + miR-NC inhibitor;
#P<0.05, ##P<0.01 vs. sh2HG + miR-NC
inhibitor. (G) MIR4435-2HG regulated B3GNT5 protein expression by
targeting miR-136-5p, as shown following transfection with
MIR4435-2HG and miR-136-5p mimics. **P<0.01 vs. pcDNA3 + miR-NC
mimics; ##P<0.01 vs. MIR4435-2HG + miR-NC mimics. (H)
Overexpression efficiency of B3GNT5. **P<0.01, ***P<0.001 vs.
pcDNA3. (I-L) Knockdown of MIR4435-2HG significantly suppressed the
proliferation, migration and invasion of liver cancer cells, these
effects were reversed by the overexpression of B3GNT5.
Magnification, ×50. *P<0.05, **P<0.01, ***P<0.001 vs. shNC
+ pcDNA3, #P<0.05, ##P<0.01,
###P<0.001 vs. sh2HG-1 + pcDNA3. (M) Data in the Gene
Expression Profiling Interactive Analysis database demonstrated the
positive relationship between B3GNT5 and MIR4435-2HG in liver
cancer. miR/miRNA, microRNA; WT, wild-type; MUT, mutant; NC,
negative control; sh, short hairpin RNA; sh2HG, shMIR4435-2HG. |
Discussion
Increasing evidence has indicated that abnormally
expressed lncRNAs play key roles in various types of cancer
(27). In the present study, the
expression levels of MIR4435-2HG were evaluated using TCGA database
and RT-qPCR assays. It was found that the expression levels of
MIR4435-2HG were significantly upregulated in liver cancer tissues
and cell lines, which was consistent with previous reports
(14,15,28). Further analysis indicated that
MIR4435-2HG overexpression was associated with cancer metastasis
and poor prognosis in patients with liver cancer. Additionally, to
explore the function of MIR4435-2HG in liver cancer progression,
silencing or overexpression of MIR4435-2HG in liver cancer cells
was performed. It was observed that silencing MIR4435-2HG reduced
the proliferation, migration and invasion of liver cancer cells,
whereas its overexpression resulted in the opposite effect on the
malignant phenotype of liver cancer cells. Regarding the mechanism
of action, the results showed that MIR4435-2HG promoted liver
cancer tumorigenesis by binding miR-136-5p to activate the
expression of B3GNT5. In previous studies, MIR4435-2HG expression
has been found to be elevated in cancer tissues compared with in
adjacent normal tissues in patients with liver cancer, and its
overexpression has been reported to promote cell proliferation by
upregulating miRNA-487a expression (15). In addition, MIR4435-2HG has been
observed to be highly expressed in liver cancer tissues, and the
malignant progression of liver cancer may be promoted by the
MIR4435-2HG/miR-22-3p/YWHAZ axis (14).
It is commonly known that lncRNAs partially mediate
cancer progression by acting as ceRNAs. For example, knockdown of
lncRNA SOX2OT has been reported to induce cell apoptosis, and
promote the proliferation, migration and invasion of glioblastoma
stem cells via the SOX2OT-miR-194-5p/miR-122-SOX3-TDGF-1 signaling
pathway (29). Furthermore, the
increased expression of lncRNA RP11-436H11.5 affected cell
proliferation and invasion in vitro and in vivo by
regulating miR-335-5p expression in renal cell carcinoma (30). In addition, HOXD-AS1 has been
demonstrated to facilitate cancer metastasis and inhibit apoptosis
via the miR19a/ARHGAP11A signaling axis in HCC (31). In the present study, the potential
target miRNAs of MIR4435-2HG were predicted using bioinformatics
tools. It has been revealed that miR-136-5p expression may be
markedly decreased in various types of cancer. For example,
miR-136-5p may inhibit HCC cell proliferation, migration and
invasion by targeting the WNK1 pathway (32). It may also inhibit lung squamous
cell carcinoma progression by deactivating ROCK1 (33). Furthermore, miR-136-5p has been
reported to regulate the expression of IRX5 to suppress cell
proliferation in HCC (34). The
present study reported that MIR4435-2HG could function as a ceRNA
of miR-136-5p. The mutual negative regulatory association between
MIR4435-2HG and miR-136-5p was elucidated. Furthermore, it was
demonstrated that the MIR4435-2HG knockdown-mediated inhibition of
cell proliferation, migration and invasion was reversed by
miR-136-5p silencing. Bioinformatics tools were used in the current
study to search for candidate targets of miR-136-5p, and B3GNT5 was
used for expression analysis. To the best of our knowledge, the
function of B3GNT5 in liver cancer development has not been
previously reported. In the present study, mutations in B3GNT5 in
liver cancer were identified through online database analysis. The
expression of B3GNT5 in liver cancer was observed to be
upregulated, but its methylation level was decreased, indicating
that its increased expression in liver cancer may be beneficial for
cancer progression. Therefore, the investigation of B3GNT5
methylation and its role in liver cancer may be a potential future
research direction. From the perspective of gene expression
regulation, the epigenetic results indeed found that the expression
of B3GNT5 was altered in liver cancer, which indicated that B3GNT5
may play an important role in the occurrence and development of
liver cancer and is worthy of further study. Therefore, B3GNT5 may
be a candidate target for the treatment of liver cancer.
Additionally, it was found that B3GNT5 was highly expressed in
liver cancer tissues and cell lines, and was related to poor
prognosis of patients. Moreover, B3GNT5 expression was regulated by
the MIR4435-2HG/miR-136-5p axis. Functionally, the
overexpression of B3GNT5 relieved the sh-2HG-1-induced inhibition
of cell proliferation, migration and invasion. These results were
consistent with the previous study in human glioma in which B3GNT5
was revealed to act as a tumor promoter (35). However, a major limitation of the
current study is that B3GNT5-related signaling pathways and the
effect of MIR4435-2HG in liver cancer in vivo were not
comprehensively studied and thus will be explored further in the
future.
In summary, it was found that MIR4435-2HG was highly
expressed in liver cancer tissues and cells. These data indicated
the oncogenic role of MIR4435-2HG in liver cancer by competitively
sponging miR-136-5p to mediate B3GNT5 expression. To the best of
our knowledge, this was the first study to reveal the potential
mechanism of the MIR4435-2HG/miR-136-5p/B3GNT5 axis
in liver cancer. The present study may provide a potential
biomarker for the diagnosis and treatment of liver cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GXi, YZ and BL designed and conceived the study. YZ,
BL, GXu and CH conducted the experiments. All authors analyzed the
data. GXi, YZ and BL drafted the manuscript. All authors have read
and approved the final manuscript. GXi and YZ confirmed the
authenticity of all the raw data.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
the Second Hospital of Tianjin Medical University (approval no.
KY2020K093; Tianjin, China). All patients provided written informed
consent prior to the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Satriano L, Lewinska M, Rodrigues PM,
Banales JM and Andersen JB: Metabolic rearrangements in primary
liver cancers: Cause and consequences. Nat Rev Gastroenterol
Hepatol. 16:748–766. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Anwanwan D, Singh SK, Singh S, Saikam V
and Singh R: Challenges in liver cancer and possible treatment
approaches. Biochim Biophys Acta Rev Cancer. 1873:1883142020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harris PS, Hansen RM, Gray ME, Massoud OI,
McGuire BM and Shoreibah MG: Hepatocellular carcinoma surveillance:
An evidence-based approach. World J Gastroenterol. 25:1550–1559.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aldrighetti L, Pulitano C, Catena M, Arru
M, Guzzetti E, Halliday J and Ferla G: Liver resection with portal
vein thrombectomy for hepatocellular carcinoma with vascular
invasion. Ann Surg Oncol. 16:12542009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li J, Li Z, Zheng W, Li X, Wang Z, Cui Y
and Jiang X: LncRNA-ATB: An indispensable cancer-related long
noncoding RNA. Cell Prolif. 50:e123812017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ulitsky I and Bartel DP: lincRNAs:
Genomics, evolution, and mechanisms. Cell. 154:26–46. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: A new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen C, Zheng H, Luo Y, Kong Y, An M, Li
Y, He W, Gao B, Zhao Y, Huang H, et al: SUMOylation promotes
extracellular vesicle-mediated transmission of lncRNA ELNAT1 and
lymph node metastasis in bladder cancer. J Clin Invest.
131:e1464312021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qin Y, Hou Y, Liu S, Zhu P, Wan X, Zhao M,
Peng M, Zeng H, Li Q, Jin T, et al: A novel long non-coding RNA
lnc030 maintains breast cancer stem cell stemness by stabilizing
SQLE mRNA and increasing cholesterol synthesis. Adv Sci (Weinh).
8:20022322020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang F, Wang H, Yu J, Yao X, Yang S, Li
W, Xu L and Zhao L: LncRNA CRNDE attenuates chemoresistance in
gastric cancer via SRSF6-regulated alternative splicing of PICALM.
Mol Cancer. 20:62021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dong X, Yang Z, Yang H, Li D and Qiu X:
Long non-coding RNA MIR4435-2HG promotes colorectal cancer
proliferation and metastasis through miR-206/YAP1 axis. Front
Oncol. 10:1602020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Wu M, Lu Y, He K, Cai X, Yu X, Lu
J and Teng L: LncRNA MIR4435-2HG targets desmoplakin and promotes
growth and metastasis of gastric cancer by activating Wnt/β-catenin
signaling. Aging (Albany NY). 11:6657–6673. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang M, He X, Huang X, Wang J, He Y and
Wei L: LncRNA MIR4435-2HG-mediated upregulation of TGF-β1 promotes
migration and proliferation of nonsmall cell lung cancer cells.
Environ Toxicol. 35:582–590. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shen X, Ding Y, Lu F, Yuan H and Luan W:
Long noncoding RNA MIR4435-2HG promotes hepatocellular carcinoma
proliferation and metastasis through the miR-22-3p/YWHAZ axis. Am J
Transl Res. 12:6381–6394. 2020.PubMed/NCBI
|
|
15
|
Kong Q, Liang C, Jin Y, Pan Y, Tong D,
Kong Q and Zhou J: The lncRNA MIR4435-2HG is upregulated in
hepatocellular carcinoma and promotes cancer cell proliferation by
upregulating miRNA-487a. Cell Mol Biol Lett. 24:262019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang S, Zhang X, Li Z, Wang W, Li B, Huang
X, Sun G, Xu J, Li Q, Xu Z, et al: Circular RNA profile identifies
circOSBPL10 as an oncogenic factor and prognostic marker in gastric
cancer. Oncogene. 38:6985–7001. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Han C, Fu Y, Zeng N, Yin J and Li Q:
LncRNA FAM83H-AS1 promotes triple-negative breast cancer
progression by regulating the miR-136-5p/metadherin axis. Aging
(Albany NY). 12:3594–3616. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kang W, Wang Q, Dai Y, Wang H, Wang M,
Wang J, Zhang D, Sun P, Qi T, Jin X and Cui Z: Hypomethylation of
PlncRNA-1 promoter enhances bladder cancer progression through the
miR-136-5p/Smad3 axis. Cell Death Dis. 11:10382020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Z, Huang C, Zhang A, Lu C and Liu L:
Overexpression of circRNA_100290 promotes the progression of
laryngeal squamous cell carcinoma through the miR-136-5p/RAP2C
axis. Biomed Pharmacother. 125:1098742020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu N, Wu YP, Ke ZB, Liang YC, Cai H, Su
WT, Tao X, Chen SH, Zheng QS, Wei Y and Xue XY: Identification of
key DNA methylation-driven genes in prostate adenocarcinoma: An
integrative analysis of TCGA methylation data. J Transl Med.
17:3112019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kresovich JK, Joyce BT, Gao T, Zheng Y,
Zhang Z, Achenbach CJ, Murphy RL, Just AC, Shen J, Yang H, et al:
Promoter methylation of PGC1A and PGC1B predicts cancer incidence
in a veteran cohort. Epigenomics. 10:733–743. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Weisenberger DJ, Liang G and Lenz HJ: DNA
methylation aberrancies delineate clinically distinct subsets of
colorectal cancer and provide novel targets for epigenetic
therapies. Oncogene. 37:566–577. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kwok CT, Marshall AD, Rasko JE and Wong
JJ: Genetic alterations of m 6 A regulators predict
poorer survival in acute myeloid leukemia. J Hematol Oncol.
10:392017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fernandes T, Barretti DL, Phillips MI and
Menezes Oliveira E: Exercise training prevents obesity-associated
disorders: Role of miRNA-208a and MED13. Mol Cell Endocrinol.
476:148–154. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Correia de Sousa M, Gjorgjieva M, Dolicka
D, Sobolewski C and Foti M: Deciphering miRNAs' action through
miRNA editing. Int J Mol Sci. 20:62492019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Klingenberg M, Matsuda A, Diederichs S and
Patel T: Non-coding RNA in hepatocellular carcinoma: Mechanisms,
biomarkers and therapeutic targets. J Hepatol. 67:603–618. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang B, Tang D, Zhang Z and Wang Z:
Identification of aberrantly expressed lncRNA and the associated
TF-mRNA network in hepatocellular carcinoma. J Cell Biochem.
121:1491–1503. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Su R, Cao S, Ma J, Liu Y, Liu X, Zheng J,
Chen J, Liu L, Cai H, Li Z, et al: Knockdown of SOX2OT inhibits the
malignant biological behaviors of glioblastoma stem cells via
up-regulating the expression of miR-194-5p and miR-122. Mol Cancer.
16:1712017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang K, Jin W, Song Y and Fei X: LncRNA
RP11-436H11.5, functioning as a competitive endogenous RNA,
upregulates BCL-W expression by sponging miR-335-5p and promotes
proliferation and invasion in renal cell carcinoma. Mol Cancer.
16:1662017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lu S, Zhou J, Sun Y, Li N, Miao M, Jiao B
and Chen H: The noncoding RNA HOXD-AS1 is a critical regulator of
the metastasis and apoptosis phenotype in human hepatocellular
carcinoma. Mol Cancer. 16:1252017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dong H, Jian P, Yu M and Wang L: Silencing
of long noncoding RNA LEF1-AS1 prevents the progression of
hepatocellular carcinoma via the crosstalk with
microRNA-136-5p/WNK1. J Cell Physiol. 235:6548–6562. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang W, Shi J, Cheng C and Wang H:
CircTIMELESS regulates the proliferation and invasion of lung
squamous cell carcinoma cells via the miR-136-5p/ROCK1 axis. J Cell
Physiol. 235:5962–5971. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu L, Liu Y, Chen Q, Yu G, Chen J, Chen
K, Yang N, Zeng T, Yan S, Huang A and Tang H: Long-noncoding RNA
colorectal neoplasia differentially expressed gene as a potential
target to upregulate the expression of IRX5 by miR-136-5P to
promote oncogenic properties in hepatocellular carcinoma. Cell
Physiol Biochem. 50:2229–2248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jeong HY, Park SY, Kim HJ, Moon S, Lee S,
Lee SH and Kim SH: B3GNT5 is a novel marker correlated with
stem-like phenotype and poor clinical outcome in human gliomas. CNS
Neurosci Ther. 26:1147–1154. 2020. View Article : Google Scholar : PubMed/NCBI
|