In 2017, stroke was recorded as the second leading
cause of death in people >60 years old worldwide and the leading
cause of permanent disability (1,2).
The condition has become a huge global health problem (3). Ischemic stroke is the most common
type of stroke and the third leading cause of disability worldwide
(4,5). Ischemic stroke is a pathological
state of insufficient blood supply in specific parts of the brain,
particularly in the middle cerebral artery, due to sudden rupture
of cerebral vessels or local ischemia caused by cerebral artery
thrombosis or embolism, resulting in an insufficient supply of
nutrients, oxygen and glucose, energy imbalance, and finally,
neuronal cell death (6–9). The pathogenesis of ischemic stroke
is complex, involving numerous mechanisms, including oxidative
stress, neuroinflammation, excitatory neurotoxicity, ion imbalance,
energy metabolism and apoptosis (10–12). At present, recombinant tissue
plasminogen activator is the only drug approved by the Food and
Drug Administration for the treatment of acute ischemic stroke
(13). Therefore, the search for
alternative treatment strategies for ischemic stroke has attracted
increasing attention. Endogenous protection is an important
mechanism of protection and recovery after cerebral ischemia. The
hypoxia-inducible factor-1α (HIF-1α) signaling pathway serves an
important role in endogenous protection. HIF-1α regulates
angiogenesis, neuroprotection, neurogenesis, migration of neuronal
stem cells to the ischemic area and proliferation to functional
neurons by regulating the transcription of downstream target genes
(14). Strengthening the
endogenous compensatory response may become an interesting
potential treatment strategy in stroke.
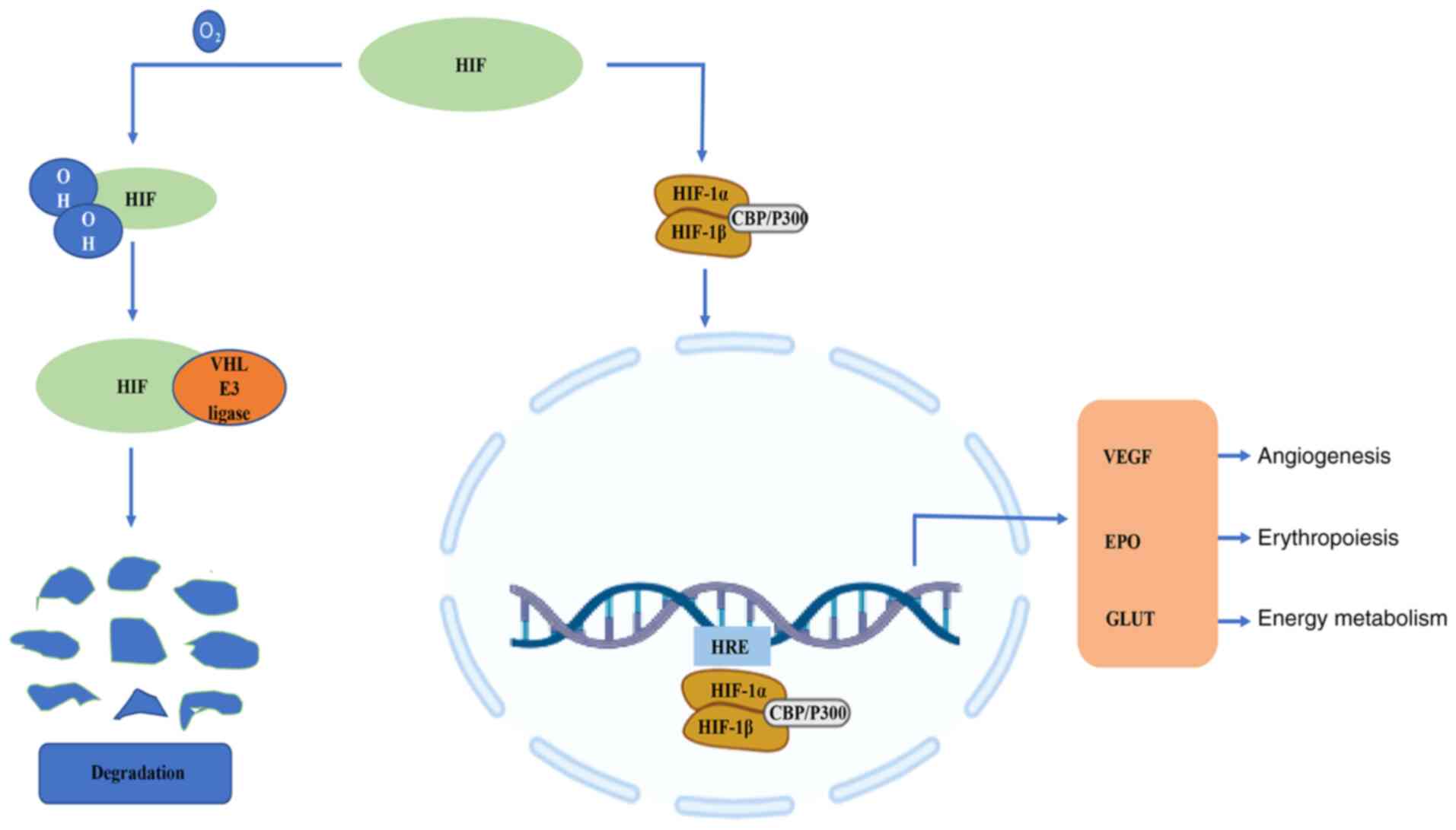
It is worth noting that the HIFs are a family of
transcription factors involved in the hypoxia response and one of
the key regulatory mechanisms of hypoxic stress at the cellular
level (15,16). HIF-1 consists of an
oxygen-regulated α subunit, HIF-1α, and a constitutively expressed
β subunit, HIF-1β (17). Although
mammals have a number of hypoxia adaptation mechanisms, including
those that have a faster response time than the HIF-1α system, the
unique degree of influence of the HIF system makes it a more
important hypoxia response regulation mechanism (18). Under normoxic conditions, the
proline and lysine residues on the oxygen-dependent degradation
domain of HIF-1α are hydroxylated, and the modified HIF-1α
interacts with the Von Hippel-Lindau E3 ubiquitin ligase complex
via ubiquitin-proteasome pathway degradation (19). However, HIF-1α is stable under
hypoxic conditions (20). With
the assistance of co-activators, such as cyclic adenosine
monophosphate response element binding protein and
acetyltransferase, HIF-1α forms a heterodimer with HIF-1β (21), and then HIF-1α is transferred to
the nucleus and combines with the target gene hypoxia response
element (HRE) to induce the expression of downstream genes
(Fig. 1). HIF-1α regulates the
transcription of >100 genes (22); its target genes encode molecules
involved in vasomotor control, angiogenesis, erythropoiesis, cell
proliferation and energy metabolism, and complex physiological and
pathological processes, such as cell death and inflammation
(23–26). During cerebral ischemia, HIF-1α is
expressed in the chronic hypoxic area around the infarct area
(27). Therefore, HIF-1α may
become a novel and valuable therapeutic target.
The literature was searched using PubMed and
ScienceDirect databases. With use of ‘pathogenesis of cerebral
ischemia’, ‘HIF-1α’ and ‘cerebral ischemia and HIF-1α’ as search
terms, 2,199, 1,550 and 42 relevant articles were retrieved,
respectively. The resulting articles were then screened according
to the clarity and specificity of the research objectives, and the
date of publication (from December 2000 to present). A total of 132
articles were selected for assessment in the current study.
The role of HIF-1α in cerebral ischemia is related
to hypoxia. During cerebral ischemia, the oxygen supply is
insufficient and partial oxygen pressure in the tissue decreases,
leading to the activation of HIF-1α (28). HIF-1α is mainly induced in the
penumbra of the cerebral ischemic region and serves an important
role in angiogenesis, glucose metabolism and cell survival
following an ischemic stroke (29–31).
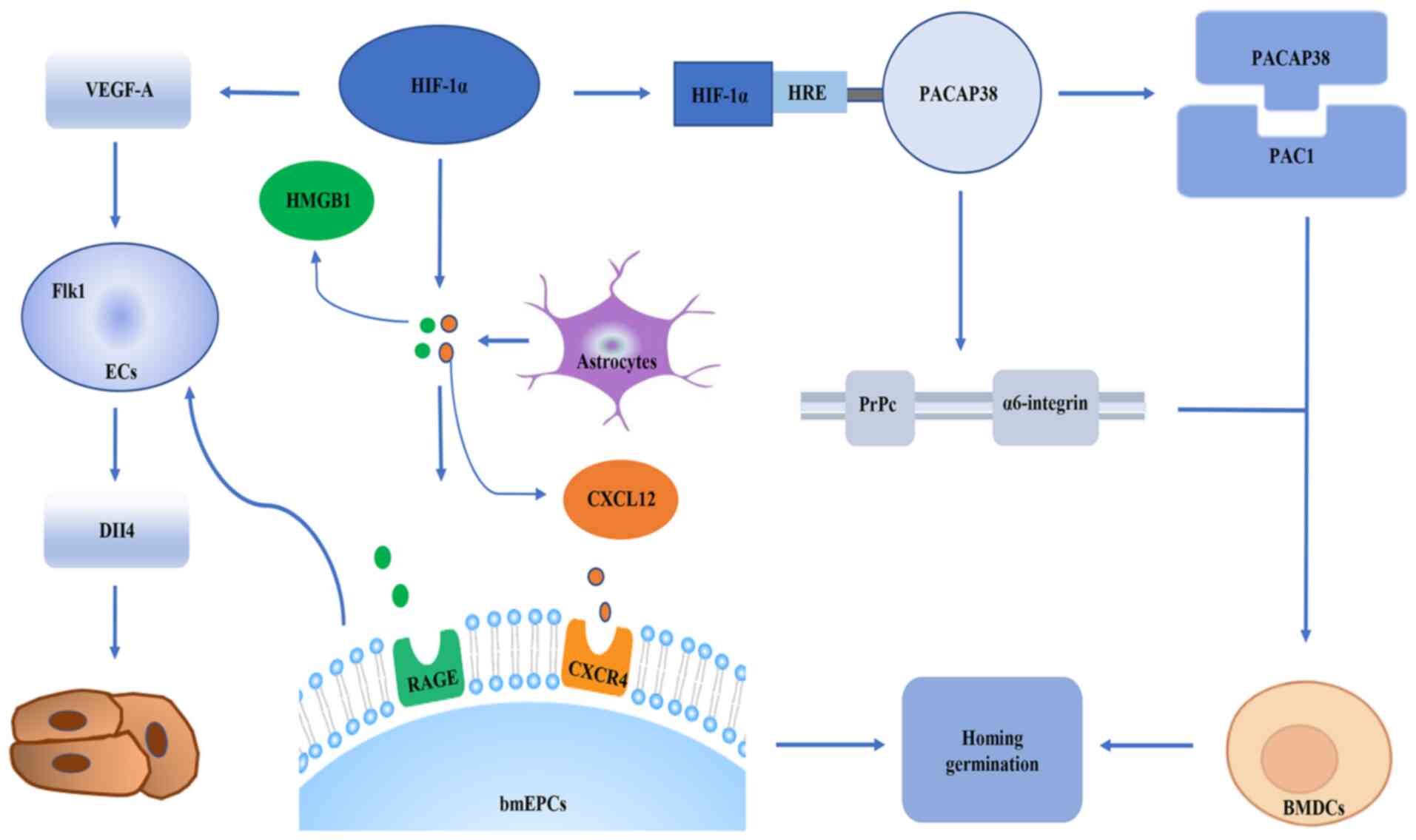
The formation of neovascularization promotes the
nerve recovery of ischemic injury after cerebral ischemia (32). Angiogenesis is one of the most
important modes of neovascularization, which depends on endothelial
progenitor cells (EPCs) (32).
HIF-1α is a transcription factor that regulates angiogenesis. It
has been widely accepted that HIF-1α serves a role in regulating
angiogenesis by regulating endothelial cells (ECs). For example, in
the acute phase of ischemia, HIF-1α serves an important role in
homing and germination of bone marrow-derived EPCs (bmEPC) in
Sprague Dawley rat brain tissues. This effect is related to
maintaining proper astrocyte responses in the ischemic brain. From
a molecular perspective, the signals of the chemokine (C-X-C motif)
ligand 12 (CXCL12)/chemokine C-X-C-motif receptor 4 (CXCR4) axis,
high mobility group protein B1 (HMGB1) and vascular endothelial
growth factor A (VEGF-A)/vascular endothelial growth factor
receptor 2 (Flk1)-neuropilin-1 (Nrp1)/delta-like ligand-4 (Dll4)
axis between astrocytes and bmEPCs are the basis for HIF-1α to
regulate the homing and sprouting of bmEPCs (33). After a stroke, bmEPCs are
mobilized from the bone marrow to the peripheral blood and
recruited to the ischemic brain (34), and CXCL12 and HMGB1 expression is
upregulated. CXCL12/CXCR4 and HMGB1 are important for the homing of
EPCs (35,36). EPCs then attach to damaged blood
vessels and repair them or migrate outward to start angiogenesis
(37). Bud angiogenesis is
another important mode of angiogenesis guided by apical cells
(38,39). Apical cells are usually mobile and
located at the top of vascular buds (38,39). Morphologically, apical cells are
rich in filamentous pseudopodia, which contributes to the formation
of vascular tubes (40). ECs
serve as a bank for apical cells. The differentiation of ECs into
apical cells is controlled by the VEGF-A/Flk1/Dll4 signaling
pathway (38,39). Specifically, when Flk1-positive
ECs are stimulated by VEGF-A, the levels of Dll4 are increased in
these cells, and thus, these cells begin to differentiate into tip
cells (38,39). In addition, Nrp1 is a co-receptor
of Flk1 and is necessary for apical cell formation (41,42). The role of astrocytes in
angiogenesis is that activated astrocytes recruit bmEPCs by
secreting HMGB1 and CXCL12 (35).
These two substances are ligands of advanced glycosylation end
product-specific receptor and recombinant CXCR4, which are
expressed on the cell membrane of EPCs (35,43). It has been revealed that
inhibition of HIF-1α can reduce the expression levels of CXCR4 in
bmEPCs, as well as reducing the expression levels of VEGF-A,
VEGF-A/Flk-1, NRP1 and Dll4 in bmEPCs. These findings suggest that
HIF-α may participate in the homing of bmEPCs via CXCL12/CXCR4 and
HMGB1, and promotes the germination of bmEPCs via
VEGF-A/Flk1-Nrp1/Dll4 (33).
Additionally, knockdown of HIF-1α in vivo reduces the number
of reactive astrocytes in the ischemic brain (33,44). Furthermore, a previous study
(31) found that the number of
reactive astrocytes increased in the brain of ischemic mice with
insufficient prolyl hydroxylases (PHDs), and insufficient PHDs led
to the stabilization of HIF-1α. This indicated that astrocytes
serve a key role in the homing and germination of bmEPCs, and
HIF-1α serves a direct role in the homing and germination of bmEPCs
via regulation of the pathway between astrocytes and bmEPCs.
Conversely, affecting the number of astrocytes has an indirect
effect on the homing and germination of bmEPCs. In addition, HIF-1α
induces bone marrow dendritic cell (BMDC) homing to ECs and
regulates angiogenesis (45). In
the molecular mechanism of BMDC transport, pituitary adenylate
cyclase-activating peptide 38 (PACAP38) increases the expression
levels of adhesion/migration-related proteins cellular prion
protein (PrPc), α6-integrin, B1 integrin, focal adhesion kinase and
CXCR4 (46–52), enhances the activities of MMP9 and
MMP2 in BMDCs, and promotes the homing and migration of BMDCs. The
PACAP38-pituitary adenylate cyclase-activating polypeptide type I
receptor isoform 1 (PAC1) signal is an important part of the homing
mechanism. During ischemia and hypoxia, HIF-1α upregulates PACAP38
by binding to the HRE on the PACAP38 promoter. The PACAP38 receptor
PAC1 is widely expressed on BMDCs. PACAP38 binds to the receptor
and promotes the homing of BMDCs to the ischemic area. In addition,
PACAP38 upregulates the expression levels of α6-integrin and PrPc
on the surface of BMDCs. This may stimulate the bone marrow
mesenchymal cells to move to the blood vessels and increase their
binding to laminin. Laminin is concentrated on the surface of the
blood vessel (53,54). The interaction can enable BMDCs to
integrate into the vascularized parenchymal area of the ischemic
brain to promote tissue repair (Fig.
2) (53,54).
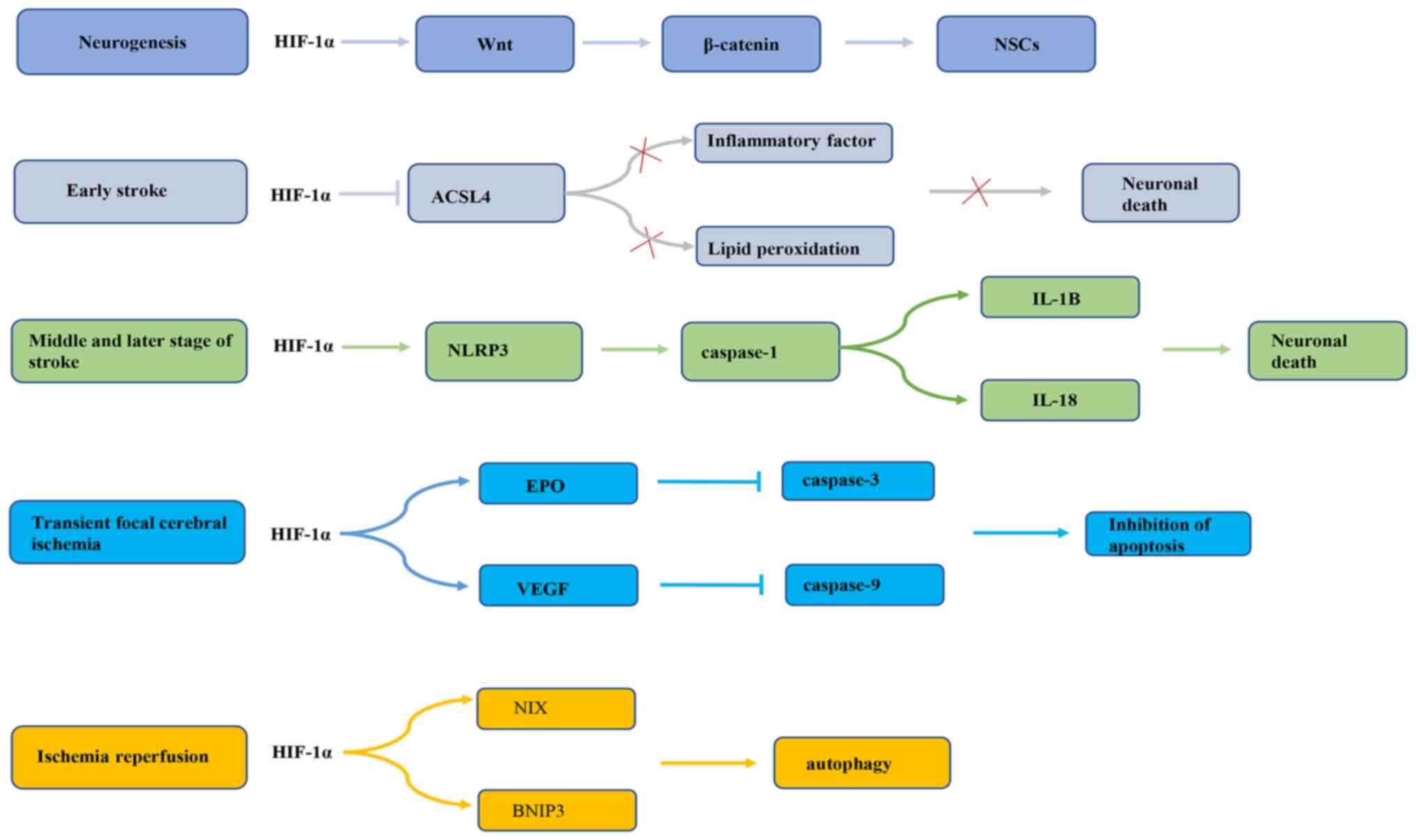
Previous research has revealed that HIF-1α serves a
dual role in promoting survival or death of nerve cells during
cerebral ischemia. First, endogenous neurogenesis is enhanced
during cerebral ischemia and hypoxia (55), which may be related to the
activation of endogenous neural stem cells (NSCs) during cerebral
ischemia (56,57). Previous studies have demonstrated
that both global and focal cerebral ischemia can increase the
proliferation and neural differentiation of NSCs located in the
subgranular area of the dentate gyrus, the anterior subventricular
area and the posterior peripheral area of the ventricle adjacent to
the hippocampus (58,59). HIF-1α, by increasing activation of
the Wnt/β-catenin signaling pathway, stimulates NSC proliferation
(60). The Wnt signaling pathway
regulates the embryonic NSC pattern, cell fate determination and
cell proliferation (61). Wnt
signaling may regulate hippocampal neurogenesis in adult rats
(62). In fact, Wnt3α mutant mice
exhibit hippocampal hypoplasia due to a lack of proliferation
(62). Wnt family members are
expressed in hippocampal astrocytes, while hippocampal
stem/progenitor cells express Wnt protein receptors and signal
components (63). It has been
reported that HIF-1α signaling is inhibited under oxygen-deprived
conditions, which may reduce β-catenin nuclear translocation and
cyclin D1 expression, delaying NSC proliferation (60). In addition, HIF-1α has different
effects on the occurrence of apoptosis in different periods of
cerebral ischemia (64,65). A previous study has demonstrated
that HIF-1α may have a neuroprotective effect in the early stage of
an ischemic stroke (66). HIF-1α
is highly expressed in rat brain tissue in the early stage of
ischemic stroke and may markedly reduce infarct cell apoptosis.
This effect may be related to the inhibition of acyl CoA synthase
long chain family member 4 (ACSL4) by HIF-1α (67). ACSL4 is an important metabolic
isoenzyme of polyunsaturated fatty acids. ACSL4 promotes neuronal
death by enhancing lipid peroxidation (a marker of iron drop
disease). In addition, ACSL4 may promote the microglia mediated
inflammatory response. HIF-1α inhibits ACSL4 expression, thereby
reducing lipid peroxidation and inflammation, and exhibiting a
neuroprotective effect on cerebral ischemia (67). On the contrary, a previous study
conducted by Panchision (68)
demonstrated that HIF-1α expression could promote neuronal
apoptosis after long-term severe ischemia and hypoxia. HIF-1α may
regulate the inflammatory response through the NLR family
containing pyrin domain protein 3 (NLRP3) inflammasome complex,
thereby promoting apoptosis and pyrophagocytic cell death after
stroke (69). NLRP3 inflammatory
bodies are the main mediators of the inflammatory response during
ischemic stroke (70,71). Upregulation of NLRP3 inflammatory
bodies activates pre-caspase-1 by cleavage (72), thus promoting the maturation of
IL-1B and IL-18. HIF-1α regulates the NLRP3 inflammatory focal
pathway, resulting in brain cell death (69). In addition, in the transient focal
cerebral ischemia model, on the one hand, HIF-1α upregulates
erythropoietin (EPO) expression, thereby inhibiting the expression
of activated caspase-3 in neurons and inhibiting neuronal apoptosis
to improve the recovery of nerve function (73). On the other hand, HIF-1α exerts a
neuroprotective effect on transient focal cerebral ischemia by
upregulating VEGF and downregulating caspase-9 (74). A previous study also found that
the anti-apoptotic effect of HIF-1α gene therapy effectively
reduced the neurological deficit score and brain edema at 24 and 72
h after reperfusion, and inhibited the pathological damage and
apoptosis of nerve cells in a rat middle cerebral artery occlusion
model (75). Additionally, a
previous study reported that HIF-1α may improve brain damage after
ischemia/reperfusion (I/R) via BCL2/adenovirus E1B interacting
protein 3 (BNIP3) and Bcl-2 family proteins containing BH3
domain-dependent enhancement of autophagy cell survival (76). Taken together, these observations
suggest that HIF-1α may induce cell death in severe and long-term
ischemia, but that activation in mild ischemic stress could promote
cell survival (Fig. 3). This may
be related to different mechanisms being involved in the regulation
of the response to ischemic stroke by HIF-1α. The role of HIF-1α in
neuroprotection requires further study.
During cerebral ischemia, endogenous factors, such
as neurotransmitters, amino acids and inorganic salts, serve a
protective role against cerebral ischemia by regulating HIF-1α to
influence the angiogenesis and neuroprotection of ischemic brain
tissue (77–80). For example, choline may increase
the levels of α7 nicotinic acetylcholine receptor, induce the
expression of HIF-1α and VEGF, and promote the formation of
cerebral arteries and cerebral cortex capillaries, thereby
effectively reducing cerebral ischemic damage in permanent middle
cerebral artery occlusion (MCAO) rats (77). Peroxynitrite promotes neurogenesis
by activating HIF-1α and enhancing the Wnt/β-catenin signaling
pathway (78). Arginine reduces
the inflammatory response mediated by HIF-1α and protects against
the death of ischemic neurons after I/R injury in rats (79). Glycine inhibits HIF-1α by
inhibiting the upregulation of NF-κB/p65 after I/R injury, thereby
inhibiting pro-inflammatory activity (80).
MicroRNAs (miRNAs/miRs) are a type of small
endogenous non-coding single-stranded RNA that regulate protein
expression by inducing mRNA degradation or interfering with
translation. miRNAs have been found to be involved in the
pathogenesis of stroke. To date, numerous miRNAs have been
determined to be involved in the molecular process of the ischemic
cascade (81,82). Several miRNAs, including
miR-376b-5p, miR-433, miR-335, miRNA-210 and miR-155-5p, have also
been demonstrated to regulate HIF-1α during cerebral ischemia.
Notably, among these miRNAs, miRNA-210 is positively associated
with HIF-1n expression. When miRNA-210 expression is upregulated,
the gene and protein expression levels of HIF-1α and VEGF are
increased, and their expression trends are consistent (83). Recently, it has been revealed that
elevated levels of miRNA-210 increase neuronal cell apoptosis by
activating the HIF-1α-VEGF signaling pathway. By contrast,
downregulation of miRNA-210 expression markedly inhibits the gene
and protein expression of HIF-1α and VEGF (83). The expression levels of
miR-376b-5p, miR-433, miR-335 and miR-155-5p are negatively
associated with the expression levels of HIF-1α (84–87). miR-376b-5p inhibits angiogenesis
after cerebral ischemia via the HIF-1α-mediated VEGFA-Notch1
signaling pathway (84).
Overexpression or downregulation of miR-433 alters the mRNA and
protein levels of HIF-1α and its downstream genes, VEGF, glucose
transporter 1 (GLUT1) and angiopoietin 2 (Angpt2) (85). In addition, miR-335, as a direct
regulator of HIF-1α, serves different roles in different periods of
cerebral ischemia, which may be related to the different effects of
HIF-1α in different periods of cerebral ischemia. In the early
stage of cerebral ischemia, miR-335 mimic may reduce the area of
cerebral infarction, while the levels of HIF-1α protein are lower.
This results in the decreased expression of downstream target genes
of HIF-1α, including Angpt2, BNIP3, MMP9, plasminogen activator
inhibitor-1 and VEGF-A. By contrast, in the middle and late stages
of cerebral ischemia, the use of anti-miR-335 is beneficial. HIF-1α
protein is upregulated and then its downstream gene expression
levels increase (86). miR-335
regulates HIF-1α expression and also affects neurovascular
permeability, cell death and the blood-brain barrier, resulting in
a reduction in infarct volume (86). Furthermore, a recent study has
demonstrated that miR-155-5p targets HIF-1α in NSCs (87). Inhibition of miR-155-5p may
promote the viability of NSCs and inhibit cell apoptosis induced by
oesophago-gastro-duodenoscopy (OGD). Additionally, after
transplantation, NSCs inhibit miR-155-5p, and this also enhances
the inhibition of inflammation and oxidative stress, which enhances
the protection against cerebral infarction. Long non-coding RNAs
(lncRNAs) are a relatively newly discovered class of non-coding
RNA, ranging in length from ~200 nucleotides to several kilobases
(88). lncRNAs are dynamically
expressed in tissues, based on differentiation stages and cell
type-specific patterns, and participate in numerous normal cellular
processes. lncRNAs can compete with specific mRNAs for the same
miRNA pool. The result is that the binding of miRNAs to target
mRNAs is inhibited or reduced, and the function of miRNA
post-transcriptional silencing is impaired (89,90). HIF-1α-AS2 is an antisense lncRNA
derived from the natural antisense transcript of HIF-1α (91). A previous study has revealed that
the expression levels of HIF-1α-AS2 are upregulated in hypoxic
human umbilical vein ECs (HUVECs) (92). Upregulation of HIF-1α-AS2 leads to
downregulation of miR-153-3p, which can reduce the
post-transcriptional silencing of HIF-1α. MCAO reduces the level of
miR-153-3p RNA in the infarct area and increases the protein levels
of HIF-1α, VEGF-A and Notch1. This function of HIF-1α-AS2 promotes
the activation of the HIF-1α/VEGFA/Notch1 cascade, thereby
promoting the vitality, migration and tube formation of HUVECs
(92).
Preconditioning has a certain protective effect on
cerebral ischemia. To date, several preconditioning methods have
been used to induce ischemic tolerance (IT). Studies have revealed
that during pretreatment, HIF-1α is activated to serve a
neuroprotective role (93,94).
Hypoxic preconditioning is a phenomenon in which mild hypoxia may
induce a strong state of IT to resist the subsequent damage caused
by severe hypoxia, which exists in a number of organs, especially
the brain (95–98). Hypoxic preconditioning improves
the survival rate of rats with cerebral ischemia, reduces
neurological deficits, increases the object recognition and social
recognition memory of the rat, and inhibits the inflammatory
response caused by cerebral ischemia. These effects are regulated
by HIF-1α (99). During hypoxia
preconditioning, IT levels increase, which is related to the
activation of HIF-1α and the expression of its target genes GLUT1,
EPO, VEGFα, Bcl-2 and inducible nitric oxide synthase. Also during
hypoxic preconditioning, olfactory mucosal mesenchymal stem cells
activate HIF-1α in vitro to inhibit the pyrolysis and
apoptosis of microglia after cerebral I/R injury (100). In addition to hypoxic
preconditioning, IPC is also used as a means of cerebral ischemia
protection. IPC is defined as a transient sublethal ischemic
injury, which may mobilize protective mechanisms to improve
neuronal damage following fatal ischemia (101). A previous study has revealed
that IPC protects CA1 pyramidal neurons from non-IPC lethal
ischemia by increasing HIF-1α expression in CA1 pyramidal neurons,
thereby enhancing the expression of VEGF and the activation of
NF-κB (102). The increase of
HIF-1α may be related to the continuous increase of P2X7 receptors
in astrocytes. Activation of P2X7 receptors leads to the increase
of HIF-1α. This hypoxia-independent, but P2X7 receptor-dependent
mechanism could induce persistent expression of HIF-1α in
astrocytes, thereby effectively inducing IT and neuroprotection
against ischemia (103). Studies
have also demonstrated that remote IPC (RIPC) could improve the
response of peripheral immune cells by regulating the upregulation
of HIF-1α (104,105). This effect may be the protective
effect of cerebral ischemia mediated by RIPC activation of the
HIF-1α/AMP-activated protein kinase (AMPK)/70-kilodalton heat shock
protein (HSP70) signaling pathway. Highly conserved HSPs, as
molecular chaperones of abnormally folded proteins in cellular
stress, are induced through the HIF-1α pathway during hypoxia, and
the existing evidence also demonstrates that, in newborns, HSPs
transform into the mature conformation, and HSP70 and HSP90 serve
an important role in the post-translational process (106). The AMPK-histone deacetylase 5
signaling pathway promotes HIF-1α through the deacetylation of
HSP70 in the cytoplasm (107,108). The nuclear accumulation of
HIF-1α and the activation of HIF-1α function indicate that RIPC
mediates ischemic protection through the interaction between AMPK,
HIF-1α and HSP70 (109). In
addition, exercise preconditioning is a special neuronal IPC, which
may induce brain tolerance to ischemia, enhance neuroprotection and
resist a series of brain damages caused by ischemia (110,111). Previous studies have
demonstrated that exercise pretreatment 3 weeks before stroke
improves the structural integrity of the brain microvascular
structure in rats (112–114). This effect may be related to
HIF-1α. The pre-IPC exercise induces cerebral IT mediated by
neurons and astrocytes by increasing the expression levels of
HIF-1α (115,116). In addition, HIF-1α triggers the
expression of endothelin 1, increases the expression of B-type
natriuretic peptide and has a neuroprotective effect (117).
Platelet drugs and thrombolytic drugs are the only
ischemic stroke drugs supported by strong clinical evidence
(118). However, the application
of these two treatment methods is often limited by the potential
risk of cerebral hemorrhage and a narrow treatment time window. In
general, after decades of practice and investigation, the number of
effective interventions for ischemic stroke is limited. Therefore,
it is necessary to study further feasible and effective treatment
options. A number of studies (119–122) have demonstrated that certain
natural compounds, chemical drugs and traditional Chinese medicine
compounds may alleviate cerebral ischemic injury through HIF-1α. A
previous study found that certain compounds and traditional Chinese
medicines serve a role in angiogenesis and vascular protection
through the HIF-1α/VEGF signaling pathway, thereby protecting
against cerebral ischemic injury (123). In vivo and in
vitro studies have demonstrated that catalpol directly
activates the HIF-1α/VEGF signaling pathway in the brain and
primary cerebral microvascular ECs of rats with cerebral ischemia,
protects the vascular structure and promotes blood vessel
generation (120). Astragaloside
IV activates the HIF-1α/VEGF/Notch signaling pathway through
miRNA-210 to promote angiogenesis (121). Fluoxetine induces a cascade of
events leading to the upregulation of the expression of
HIF-1α-Netrin/VEGF protein, promotes angiogenesis after ischemic
stroke and improves long-term functional recovery after ischemic
stroke (122,123). Racemic dl-3-n-butylphthalide
treatment could also promote functional recovery after focal
transient cerebral ischemia, and this recovery and dl-NBP may
upregulate the expression of HIF-1α-VEGF and Notch-Dll4, and then
affect the integrity of white matter, the number of capillaries and
the expression of tight junction protein occludin (124). In addition, certain compounds
serve a neuroprotective role by regulating HIF-1α. Berberine
pretreatment enhances the accumulation of HIF-1α by activating
PI3K/Akt and induces the production of sphingosine-1-phosphate
(S1P) by promoting HIF-1α-mediated sphingosine kinase 2 (Sphk2)
transcription and activation of Sphk2. S1P protects neuronal cells
against hypoxia and ischemia by activating high-affinity G
protein-coupled receptors, and serves an important protective role
in tissue I/R injury (125).
Methylene blue protects hippocampal-derived neuronal cells from
OGD-reoxygenation damage by increasing the content of HIF-1α
protein and activating the EPO signaling pathway (126). Salidroside induces the
production of HIF-1α subunit and EPO via the PI3K/Akt signaling
pathway, and exerts anti-inflammatory effects on cerebral ischemia
and reperfusion (127). Certain
drugs inhibit HIF-1α to exert neuroprotective effects, while others
upregulate HIF-1α to produce the same effect. Curcumin exerts a
neuroprotective effect by inhibiting the interaction between HIF-1α
and autophagy in cerebral I/R injury (128). A recent study has reported that
nateglinide stabilizes HIF-1 cerebral I/R injury by inhibiting
STAT-3 phosphorylation and stops the expression of HIF-1α-dependent
inflammation and mediators of apoptosis, namely
phorbol-12-myristate-13-acetate-induced protein 1 and NF-κB
(129). Different drugs have a
neuroprotective effect through the opposite regulation of HIF-1α,
which may be due to the different mechanisms of HIF-1α in the
neuroprotective effect of cerebral ischemia. Different drugs affect
different pathways by activating or inhibiting HIF-1α, and serve a
role in cerebral ischemia. Angelica sinensis has a
neuroprotective effect on astrocyte-mediated infarct expansion
through HIF-1α-mediated angiogenesis, as well as HIF-1α-mediated
anti-apoptotic effects. Angelica sinensis activates
p38/MAPK/HIF-1α/VEGF-A/cAMP-response element binding protein/von
Willebrand factor signaling to mediate angiogenesis (130). In addition, the anti-apoptotic
effect of Angelica sinensis has been attributed to the
activation of the HIF-1α/VEGF-A/p-Bad signaling pathway mediated by
p38/MAPK (130). This activation
could lead to Bad inactivation, maintain the integrity of the outer
mitochondrial membrane and prevent cytochrome caspase-3-mediated
apoptosis in the cortical ischemic penumbra, thereby exerting an
anti-apoptotic effect (130). In
addition, a previous study demonstrated that ligustilide can
inhibit the upregulation of HIF-1α, VEGF and aquaporin 4 in an
OGD-induced blood-brain barrier model and reduce the permeability
of the OGD-induced blood-brain barrier in an in vitro model
(131). Bu Yang Huan Wu
decoction has a protective effect on the brain I/R injury in MCAO
rats by inhibiting the activation of the HIF-1α/VEGF signaling
pathway in the brain and stabilizing the β epithelial
Na+ channel ion channel, suggesting that Bu Yang Huan Wu
decoction may be used to treat acute brain injury during stroke
(Table I) (132).
An increasing number of studies have demonstrated
that HIF-1α serves a key role in cerebral ischemia. The present
review describes the means by which HIF-1α is activated during
cerebral ischemia and how it serves a protective role in cerebral
ischemic tissues in terms of angiogenesis and neuroprotection. In
terms of neuroprotection, since HIF-1α expression in ischemic
stroke may be controlled by different mechanisms, HIF-1α has a dual
effect. In addition, when cerebral ischemia occurs, endogenous
regulatory factors directly or indirectly regulate HIF-1α, which
may be the key mechanism of endogenous protection during cerebral
ischemia. Recent research has also revealed that preconditioning
has a positive therapeutic effect on cerebral ischemia and may
become a novel clinical treatment for cerebral ischemia. Natural
medicines and traditional Chinese medicines could be used to treat
cerebral ischemia by regulating HIF-1α. HIF-1α is expected to
become a novel target for the treatment of cerebral ischemic
diseases, and identifying the effect of natural products on HIF-1α
is also a future research direction.
Whether the signals and pathways initiated by HIF-1α
in hypoxia (or hypoxic diseases) serve the same role in cerebral
ischemia needs to be further confirmed. As HIF-1α regulates
multiple downstream target genes, and the related pathways and
mechanisms are complex, the current literature on the mechanism of
HIF-1α in cerebral ischemia is not comprehensive and in-depth.
Therefore, it is necessary to further study the role and mechanism
of HIF-1α in the pathophysiology of cerebral ischemia.
Not applicable.
The present study was supported by the National Natural Science
Foundation of China (grant no. 81973588) and the Joint Guidance
Project of Provincial Natural Science Foundation (grant no.
LH2020H094).
Not applicable.
HH conceived and designed the review. QL retrieved
the relevant literature and wrote the manuscript. PD reviewed and
edited the manuscript. Data authentication is not applicable. All
authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
WHO publishes definitive atlas on global
heart disease and stroke epidemic. Indian J Med Sci. 58:405–406.
2004.PubMed/NCBI
|
|
2
|
Cramer SC, Wolf SL, Adams HP Jr, Chen D,
Dromerick AW, Dunning K, Ellerbe C, Grande A, Janis S, Lansberg MG,
et al: Stroke recovery and rehabilitation research: Issues,
opportunities, and the national institutes of health strokeNet.
Stroke. 48:813–819. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
GBD 2016 Causes of Death Collaborators, .
Global, regional, and national age-sex specific mortality for 264
causes of death, 1980–2016: A systematic analysis for the global
burden of disease study 2016. Lancet. 390:1151–1210. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Benjamin EJ, Blaha MJ, Chiuve SE, Cushman
M, Das SR, Deo R, de Ferranti SD, Floyd J, Fornage M, Gillespie C,
et al: Heart disease and stroke statistics-2017 update: A report
from the American Heart Association. Circulation. 135:e146–e603.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jianrong S, Yanjun Z, Chen Y and Jianwen
X: DUSP14 rescues cerebral ischemia/reperfusion (IR) injury by
reducing inflammation and apoptosis via the activation of Nrf-2.
Biochem Biophys Res Commun. 509:713–721. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim JY, Kawabori M and Yenari MA: Innate
inflammatory responses in stroke: Mechanisms and potential
therapeutic targets. Curr Med Chem. 21:2076–2097. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tobin MK, Bonds JA, Minshall RD,
Pelligrino DA, Testai FD and Lazarov O: Neurogenesis and
inflammation after ischemic stroke: What is known and where we go
from here. J Cereb Blood Flow Metab. 34:1573–1584. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang H, Sun X, Xie Y, Zan J and Tan W:
Isosteviol sodium protects against permanent cerebral ischemia
injury in mice via inhibition of NF-κB-mediated inflammatory and
apoptotic responses. J Stroke Cerebrovasc Dis. 26:2603–2614. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ishrat T, Sayeed I, Atif F and Stein DG:
Effects of progesterone administration on infarct volume and
functional deficits following permanent focal cerebral ischemia in
rats. Brain Res. 1257:94–101. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hazell AS: Excitotoxic mechanisms in
stroke: An update of concepts and treatment strategies. Neurochem
Int. 50:941–953. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rothman SM and Olney JW: Glutamate and the
pathophysiology of hypoxic-ischemic brain damage. Ann Neurol.
19:105–111. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lo EH, Dalkara T and Moskowitz MA:
Mechanisms, challenges and opportunities in stroke. Nat Rev
Neurosci. 4:399–415. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wardlaw JM, Murray V, Berge E, del Zoppo
G, Sandercock P, Lindley RL and Cohen G: Recombinant tissue
plasminogen activator for acute ischaemic stroke: An updated
systematic review and meta-analysis. Lancet. 379:2364–2372. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Prabhakar NR and Semenza GL: Adaptive and
maladaptive cardiorespiratory responses to continuous and
intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2.
Physiol Rev. 92:967–1003. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kruschewski M, Foitzik T, Perez-Cantó A,
Hübotter A and Buhr HJ: Changes of colonic mucosal microcirculation
and histology in two colitis models: An experimental study using
intravital microscopy and a new histological scoring system. Dig
Dis Sci. 46:2336–2343. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Semenza GL and Wang GL: A nuclear factor
induced by hypoxia via de novo protein synthesis binds to the human
erythropoietin gene enhancer at a site required for transcriptional
activation. Mol Cell Biol. 12:5447–5454. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harris AL: Hypoxia-a key regulatory factor
in tumour growth. Nat Rev Cancer. 2:38–47. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wilkins SE, Abboud MI, Hancock RL and
Schofield CJ: Targeting protein-protein interactions in the HIF
system. ChemMedChem. 11:773–786. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pereira T, Zheng X and Poellinger L:
Degradation of the hypoxia-inducible factor 1alpha: Where does it
happen? Cell Cycle. 5:2720–2722. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee JW, Bae SH, Jeong JW, Kim SH and Kim
KW: Hypoxia-inducible factor (HIF-1)alpha: Its protein stability
and biological functions. Exp Mol Med. 36:1–12. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rabie T and Marti HH: Brain protection by
erythropoietin: A manifold task. Physiology (Bethesda). 23:263–274.
2008.PubMed/NCBI
|
|
22
|
Schito L and Semenza GL: Hypoxia-inducible
factors: Master regulators of cancer progression. Trends Cancer.
2:758–770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Semenza GL: Targeting HIF-1 for cancer
therapy. Nat Rev Cancer. 3:721–732. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sharp FR and Bernaudin M: HIF1 and oxygen
sensing in the brain. Nat Rev Neurosci. 5:437–448. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ke XJ and Zhang JJ: Changes in HIF-1α,
VEGF, NGF and BDNF levels in cerebrospinal fluid and their
relationship with cognitive impairment in patients with cerebral
infarction. J Huazhong Univ Sci Technolog Med Sci. 33:433–437.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kuang S, Zheng J, Yang H, Li S, Duan S,
Shen Y, Ji C, Gan J, Xu XW and Li J: Structure insight of GSDMD
reveals the basis of GSDMD autoinhibition in cell pyroptosis. Proc
Natl Acad Sci USA. 114:10642–10647. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chavez JC and LaManna JC: Activation of
hypoxia-inducible factor-1 in the rat cerebral cortex after
transient global ischemia: Potential role of insulin-like growth
factor-1. J Neurosci. 22:8922–8931. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Masoud GN and Li W: HIF-1α pathway: Role,
regulation and intervention for cancer therapy. Acta Pharm Sin B.
5:378–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Singh N, Sharma G and Mishra V: Hypoxia
inducible factor-1: Its potential role in cerebral ischemia. Cell
Mol Neurobiol. 32:491–507. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu J, Jiang L, Zhu H, Zhang L and Wang T:
Hypoxia-inducible factor-1α and erythropoietin expression in the
hippocampus of neonatal rats following hypoxia-ischemia. J Nanosci
Nanotechnol. 14:5614–5619. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li L, Saliba P, Reischl S, Marti HH and
Kunze R: Neuronal deficiency of HIF prolyl 4-hydroxylase 2 in mice
improves ischemic stroke recovery in an HIF dependent manner.
Neurobiol Dis. 91:221–235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ishikawa H, Tajiri N, Shinozuka K,
Vasconcellos J, Kaneko Y, Lee HJ, Mimura O, Dezawa M, Kim SU and
Borlongan CV: Vasculogenesis in experimental stroke after human
cerebral endothelial cell transplantation. Stroke. 44:3473–3481.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Y, Ran H, Xiao Y, Wang H, Chen Y, Chen
W and Xu X: Knockdown of HIF-1α impairs post-ischemic vascular
reconstruction in the brain via deficient homing and sprouting
bmEPCs. Brain Pathol. 28:860–874. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Borlongan CV, Glover LE, Tajiri N, Kaneko
Y and Freeman TB: The great migration of bone marrow-derived stem
cells toward the ischemic brain: Therapeutic implications for
stroke and other neurological disorders. Prog Neurobiol.
95:213–228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hayakawa K, Pham LD, Katusic ZS, Arai K
and Lo EH: Astrocytic high-mobility group box 1 promotes
endothelial progenitor cell-mediated neurovascular remodeling
during stroke recovery. Proc Natl Acad Sci USA. 109:7505–7510.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Miller JT, Bartley JH, Wimborne HJ, Walker
AL, Hess DC, Hill WD and Carroll JE: The neuroblast and angioblast
chemotaxic factor SDF-1 (CXCL12) expression is briefly up regulated
by reactive astrocytes in brain following neonatal hypoxic-ischemic
injury. BMC Neurosci. 6:632005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang ZG, Zhang L, Jiang Q and Chopp M:
Bone marrow-derived endothelial progenitor cells participate in
cerebral neovascularization after focal cerebral ischemia in the
adult mouse. Circ Res. 90:284–288. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Eilken HM and Adams RH: Dynamics of
endothelial cell behavior in sprouting angiogenesis. Curr Opin Cell
Biol. 22:617–625. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jakobsson L, Franco CA, Bentley K, Collins
RT, Ponsioen B, Aspalter IM, Rosewell I, Busse M, Thurston G,
Medvinsky A, et al: Endothelial cells dynamically compete for the
tip cell position during angiogenic sprouting. Nat Cell Biol.
12:943–953. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Phng LK, Stanchi F and Gerhardt H:
Filopodia are dispensable for endothelial tip cell guidance.
Development. 140:4031–4040. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Aspalter IM, Gordon E, Dubrac A, Ragab A,
Narloch J, Vizán P, Geudens I, Collins RT, Franco CA, Abrahams CL,
et al: Alk1 and Alk5 inhibition by Nrp1 controls vascular sprouting
downstream of Notch. Nat Commun. 6:72642015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fantin A, Vieira JM, Plein A, Denti L,
Fruttiger M, Pollard JW and Ruhrberg C: NRP1 acts cell autonomously
in endothelium to promote tip cell function during sprouting
angiogenesis. Blood. 121:2352–2362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Domanska UM, Kruizinga RC, Nagengast WB,
Timmer-Bosscha H, Huls G, de Vries EG and Walenkamp AM: A review on
CXCR4/CXCL12 axis in oncology: No place to hide. Eur J Cancer.
49:219–230. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang C, Lin G, Luan Y, Ding J, Li PC, Zhao
Z, Qian C, Liu G, Ju S and Teng GJ: HIF-prolyl hydroxylase 2
silencing using siRNA delivered by MRI-visible nanoparticles
improves therapy efficacy of transplanted EPCs for ischemic stroke.
Biomaterials. 197:229–243. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lin CH, Chiu L, Lee HT, Chiang CW, Liu SP,
Hsu YH, Lin SZ, Hsu CY, Hsieh CH and Shyu WC: PACAP38/PAC1
signaling induces bone marrow-derived cells homing to ischemic
brain. Stem Cells. 33:1153–1172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Muller WA: Mechanisms of transendothelial
migration of leukocytes. Circ Res. 105:223–230. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ferrero E, Belloni D, Contini P, Foglieni
C, Ferrero ME, Fabbri M, Poggi A and Zocchi MR: Transendothelial
migration leads to protection from starvation-induced apoptosis in
CD34+CD14+ circulating precursors: Evidence
for PECAM-1 involvement through Akt/PKB activation. Blood.
101:186–193. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Honczarenko M, Le Y, Swierkowski M, Ghiran
I, Glodek AM and Silberstein LE: Human bone marrow stromal cells
express a distinct set of biologically functional chemokine
receptors. Stem Cells. 24:1030–1041. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Torzicky M, Viznerova P, Richter S, Strobl
H, Scheinecker C, Foedinger D and Riedl E: Platelet endothelial
cell adhesion molecule-1 (PECAM-1/CD31) and CD99 are critical in
lymphatic transmigration of human dendritic cells. J Invest
Dermatol. 132:1149–1157. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
de la Rosa G, Longo N, Rodríguez-Fernández
JL, Puig-Kroger A, Pineda A, Corbí AL and Sánchez-Mateos P:
Migration of human blood dendritic cells across endothelial cell
monolayers: Adhesion molecules and chemokines involved in
subset-specific transmigration. J Leukoc Biol. 73:639–649. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kaneider NC, Kaser A, Dunzendorfer S, Tilg
H and Wiedermann CJ: Sphingosine kinase-dependent migration of
immature dendritic cells in response to neurotoxic prion protein
fragment. J Virol. 77:5535–5539. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Muller WA: Leukocyte-endothelial-cell
interactions in leukocyte transmigration and the inflammatory
response. Trends Immunol. 24:327–334. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang CC, Steele AD, Lindquist S and
Lodish HF: Prion protein is expressed on long-term repopulating
hematopoietic stem cells and is important for their self-renewal.
Proc Natl Acad Sci USA. 103:2184–2189. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gnecchi M and Melo LG: Bone marrow-derived
mesenchymal stem cells: Isolation, expansion, characterization,
viral transduction, and production of conditioned medium. Methods
Mol Biol. 482:281–294. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Miles DK and Kernie SG: Hypoxic-ischemic
brain injury activates early hippocampal stem/progenitor cells to
replace vulnerable neuroblasts. Hippocampus. 18:793–806. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Santilli G, Lamorte G, Carlessi L, Ferrari
D, Rota Nodari L, Binda E, Delia D, Vescovi AL and De Filippis L:
Mild hypoxia enhances proliferation and multipotency of human
neural stem cells. PLoS One. 5:e85752010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang P, Liu Y, Li J, Kang Q, Tian Y, Chen
X, Shi Q and Song T: Cell proliferation in ependymal/subventricular
zone and nNOS expression following focal cerebral ischemia in adult
rats. Neurol Res. 28:91–96. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Bürgers HF, Schelshorn DW, Wagner W,
Kuschinsky W and Maurer MH: Acute anoxia stimulates proliferation
in adult neural stem cells from the rat brain. Exp Brain Res.
188:33–43. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Park KI, Hack MA, Ourednik J, Yandava B,
Flax JD, Stieg PE, Gullans S, Jensen FE, Sidman RL, Ourednik V and
Snyder EY: Acute injury directs the migration, proliferation, and
differentiation of solid organ stem cells: Evidence from the effect
of hypoxia-ischemia in the CNS on clonal ‘reporter’ neural stem
cells. Exp Neurol. 199:156–178. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Qi C, Zhang J, Chen X, Wan J, Wang J,
Zhang P and Liu Y: Hypoxia stimulates neural stem cell
proliferation by increasing HIF-1α expression and activating
Wnt/β-catenin signaling. Cell Mol Biol (Noisy-le-grand). 63:12–19.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ciani L and Salinas PC: WNTs in the
vertebrate nervous system: From patterning to neuronal
connectivity. Nat Rev Neurosci. 6:351–362. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lee SM, Tole S, Grove E and McMahon AP: A
local Wnt-3a signal is required for development of the mammalian
hippocampus. Development. 127:457–467. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lie DC, Colamarino SA, Song HJ, Désiré L,
Mira H, Consiglio A, Lein ES, Jessberger S, Lansford H, Dearie AR
and Gage FH: Wnt signalling regulates adult hippocampal
neurogenesis. Nature. 437:1370–1375. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cheng YL, Park JS, Manzanero S, Choi Y,
Baik SH, Okun E, Gelderblom M, Fann DY, Magnus T, Launikonis BS, et
al: Evidence that collaboration between HIF-1α and Notch-1 promotes
neuronal cell death in ischemic stroke. Neurobiol Dis. 62:286–295.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yang Z, Zhao TZ, Zou YJ, Zhang JH and Feng
H: Hypoxia induces autophagic cell death through hypoxia-inducible
factor 1α in microglia. PLoS One. 9:e965092014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Sun Y, He W and Geng L: Neuroprotective
mechanism of HIF-1α overexpression in the early stage of acute
cerebral infarction in rats. Exp Ther Med. 12:391–395. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Cui Y, Zhang Y, Zhao X, Shao L, Liu G, Sun
C, Xu R and Zhang Z: ACSL4 exacerbates ischemic stroke by promoting
ferroptosis-induced brain injury and neuroinflammation. Brain Behav
Immun. 93:312–321. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Panchision DM: The role of oxygen in
regulating neural stem cells in development and disease. J Cell
Physiol. 220:562–568. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Jiang Q, Geng X, Warren J, Eugene Paul
Cosky E, Kaura S, Stone C, Li F and Ding Y: Hypoxia inducible
factor-1α (HIF-1α) mediates NLRP3 inflammasome-dependent-pyroptotic
and apoptotic cell death following schemic stroke. Neuroscience.
448:126–139. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
An P, Xie J, Qiu S, Liu Y, Wang J, Xiu X,
Li L and Tang M: Hispidulin exhibits neuroprotective activities
against cerebral ischemia reperfusion injury through suppressing
NLRP3-mediated pyroptosis. Life Sci. 232:1165992019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Tang B, Tang WJ, Tang YH and Deng CQ:
Astragaloside IV attenuates cerebral ischemia and reperfusion
injury and reduces activation of NLRP3 inflammasome and NF-κB
phosphorylation in rats following a transient middle cerebral
artery occlusion. Sheng Li Xue Bao. 71:424–430. 2019.(In Chinese).
PubMed/NCBI
|
|
72
|
Davis BK, Wen H and Ting JP: The
inflammasome NLRs in immunity, inflammation, and associated
diseases. Annu Rev Immunol. 29:707–735. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Li J, Tao T, Xu J, Liu Z, Zou Z and Jin M:
HIF-1α attenuates neuronal apoptosis by upregulating EPO expression
following cerebral ischemia-reperfusion injury in a rat MCAO model.
Int J Mol Med. 45:1027–1036. 2020.PubMed/NCBI
|
|
74
|
Zhu T, Zhan L, Liang D, Hu J, Lu Z, Zhu X,
Sun W, Liu L and Xu E: Hypoxia-inducible factor 1α mediates
neuroprotection of hypoxic postconditioning against global cerebral
ischemia. J Neuropathol Exp Neurol. 73:975–986. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yang ML, Tao T, Xu J, Liu Z and Xu D:
Antiapoptotic effect of gene therapy with recombinant adenovirus
vector containing hypoxia-inducible factor-1α after cerebral
ischemia and reperfusion in rats. Chin Med J (Engl). 130:1700–1706.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Guo Y: Role of HIF-1a in regulating
autophagic cell survival during cerebral ischemia reperfusion in
rats. Oncotarget. 8:98482–98494. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Jin X, Wang RH and Wang H, Long CL and
Wang H: Brain protection against ischemic stroke using choline as a
new molecular bypass treatment. Acta Pharmacol Sin. 36:1416–1425.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chen X, Zhou B, Yan T, Wu H, Feng J, Chen
H, Gao C, Peng T, Yang D and Shen J: Peroxynitrite enhances
self-renewal, proliferation and neuronal differentiation of neural
stem/progenitor cells through activating HIF-1α and Wnt/β-catenin
signaling pathway. Free Radic Biol Med. 117:158–167. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chen SF, Pan MX, Tang JC, Cheng J, Zhao D,
Zhang Y, Liao HB, Liu R, Zhuang Y, Zhang ZF, et al: Arginine is
neuroprotective through suppressing HIF-1α/LDHA-mediated
inflammatory response after cerebral ischemia/reperfusion injury.
Mol Brain. 13:632020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Liu R, Liao XY, Pan MX, Tang JC, Chen SF,
Zhang Y, Lu PX, Lu LJ, Zou YY, Qin XP, et al: Glycine exhibits
neuroprotective effects in ischemic stroke in rats through the
inhibition of M1 microglial polarization via the NF-κB p65/Hif-1α
signaling pathway. J Immunol. 202:1704–1714. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Jeyaseelan K, Lim KY and Armugam A:
MicroRNA expression in the blood and brain of rats subjected to
transient focal ischemia by middle cerebral artery occlusion.
Stroke. 39:959–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wang Y, Wang Y and Yang GY: MicroRNAs in
cerebral ischemia. Stroke Res Treat. 2013:2765402013.PubMed/NCBI
|
|
83
|
Sun JJ, Zhang XY, Qin XD, Zhang J, Wang MX
and Yang JB: miRNA-210 induces the apoptosis of neuronal cells of
rats with cerebral ischemia through activating HIF-1α-VEGF pathway.
Eur Rev Med Pharmacol Sci. 23:2548–2554. 2019.PubMed/NCBI
|
|
84
|
Li LJ, Huang Q, Zhang N, Wang GB and Liu
YH: miR-376b-5p regulates angiogenesis in cerebral ischemia. Mol
Med Rep. 10:527–535. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zhang L, Zhang Y, Zhang X, Zhang Y, Jiang
Y, Xiao X, Tan J, Yuan W and Liu Y: MicroRNA-433 inhibits the
proliferation and migration of HUVECs and neurons by targeting
hypoxia-inducible factor 1 alpha. J Mol Neurosci. 61:135–143. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Liu FJ, Kaur P, Karolina DS, Sepramaniam
S, Armugam A, Wong PT and Jeyaseelan K: miR-335 regulates Hif-1α to
reduce cell death in both mouse cell line and rat ischemic models.
PLoS One. 10:e01284322015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang D, Wang L, Bai L, Du Y, Liu L and
Chen X: Effects of inhibition of miR-155-5p in neural stem cell
subarachnoid transplant on rats with cerebral infarction. Hum Gene
Ther Methods. 30:184–193. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhang X, Li H, Burnett JC and Rossi JJ:
The role of antisense long noncoding RNA in small RNA-triggered
gene activation. RNA. 20:1916–1928. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
PLOS Genetics Staff, . Correction:
fMiRNA-192 and miRNA-204 directly suppress lncRNA HOTTIP and
interrupt GLS1-mediated glutaminolysis in hepatocellular carcinoma.
PLoS Genet. 12:e10058252016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Chen ZH, Wang WT, Huang W, Fang K, Sun YM,
Liu SR, Luo XQ and Chen YQ: The lncRNA HOTAIRM1 regulates the
degradation of PML-RARA oncoprotein and myeloid cell
differentiation by enhancing the autophagy pathway. Cell Death
Differ. 24:212–224. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Mineo M, Ricklefs F, Rooj AK, Lyons SM,
Ivanov P, Ansari KI, Nakano I, Chiocca EA, Godlewski J and Bronisz
A: The Long Non-coding RNA HIF1A-AS2 facilitates the maintenance of
mesenchymal glioblastoma stem-like cells in hypoxic niches. Cell
Rep. 15:2500–2509. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Li L, Wang M, Mei Z, Cao W, Yang Y, Wang Y
and Wen A: lncRNAs HIF1A-AS2 facilitates the up-regulation of
HIF-1α by sponging to miR-153-3p, whereby promoting angiogenesis in
HUVECs in hypoxia. Biomed Pharmacother. 96:165–172. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Fani L, Bos D, Mutlu U, Portegies MLP,
Zonneveld HI, Koudstaal PJ, Vernooij MW, Ikram MA and Ikram MK:
Global brain perfusion and the risk of transient ischemic attack
and ischemic stroke: The rotterdam study. J Am Heart Assoc.
8:e0115652019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Tsai MJ, Kuo YM and Tsai YH: Transient
ischemic attack induced by melted solid lipid microparticles
protects rat brains from permanent focal ischemia. Neuroscience.
275:136–145. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Sprick JD, Mallet RT, Przyklenk K and
Rickards CA: Ischaemic and hypoxic conditioning: Potential for
protection of vital organs. Exp Physiol. 104:278–294. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Rojas DR, Tegeder I, Kuner R and Agarwal
N: Hypoxia-inducible factor 1α protects peripheral sensory neurons
from diabetic peripheral neuropathy by suppressing accumulation of
reactive oxygen species. J Mol Med. 96:1395–1405. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Rodríguez-Reynoso S, Leal-Cortés C,
Portilla-de Buen E and López-De la Torre SP: Ischemic
preconditioning preserves liver energy charge and function on
hepatic ischemia/reperfusion injury in rats. Arch Med Res.
49:373–380. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Meng SS, Xu XP, Chang W, Lu ZH, Huang LL,
Xu JY, Liu L, Qiu HB, Yang Y and Guo FM: LincRNA-p21 promotes
mesenchymal stem cell migration capacity and survival through
hypoxic preconditioning. Stem Cell Res Ther. 9:2802018. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Yang Y, Lu F, Zhuang L, Yang S, Kong Y,
Tan W, Gong Z and Zhan S: Combined preconditioning with hypoxia and
GYKI-52466 protects rats from cerebral ischemic injury by
HIF-1α/eNOS pathway. Am J Transl Res. 9:5308–5319. 2017.PubMed/NCBI
|
|
100
|
Huang Y, Tan F, Zhuo Y, Liu J, He J, Duan
D, Lu M and Hu Z: Hypoxia-preconditioned olfactory mucosa
mesenchymal stem cells abolish cerebral
ischemia/reperfusion-induced pyroptosis and apoptotic death of
microglial cells by activating HIF-1α. Aging (Albany NY).
12:10931–10950. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Murry CE, Jennings RB and Reimer KA:
Preconditioning with ischemia: A delay of lethal cell injury in
ischemic myocardium. Circulation. 74:1124–1136. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Lee JC, Tae HJ, Kim IH, Cho JH, Lee TK,
Park JH, Ahn JH, Choi SY, Bai HC, Shin BN, et al: Roles of HIF-1α,
VEGF, and NF-κB in ischemic preconditioning-mediated
neuroprotection of hippocampal CA1 pyramidal neurons against a
subsequent transient cerebral ischemia. Mol Neurobiol.
54:6984–6998. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Hirayama Y and Koizumi S:
Hypoxia-independent mechanisms of HIF-1α expression in astrocytes
after ischemic preconditioning. Glia. 65:523–530. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Yang J, Liu C, Du X, Liu M, Ji X, Du H and
Zhao H: Hypoxia inducible factor 1α plays a key role in remote
ischemia preconditioning against stroke by modulating inflammatory
responses in rats. J Am Heart Assoc. 7:e0075892018. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Liu ZJ, Chen C, Li XR, Ran YY, Xu T, Zhang
Y, Geng XK, Zhang Y, Du HS, Leak RK, et al: Remote ischemic
preconditioning-mediated neuroprotection against stroke is
associated with significant alterations in peripheral immune
responses. CNS Neurosci Ther. 22:43–52. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Xia M, Ding Q, Zhang Z and Feng Q: Remote
limb ischemic preconditioning protects rats against cerebral
ischemia via HIF-1α/AMPK/HSP70 pathway. Cell Mol Neurobiol.
37:1105–1114. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Fath DM, Kong X, Liang D, Lin Z, Chou A,
Jiang Y, Fang J, Caro J and Sang N: Histone deacetylase inhibitors
repress the transactivation potential of hypoxia-inducible factors
independently of direct acetylation of HIF-alpha. J Biol Chem.
281:13612–13619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Liang D, Kong X and Sang N: Effects of
histone deacetylase inhibitors on HIF-1. Cell Cycle. 5:2430–2435.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Chen S, Yin C, Lao T, Liang D, He D, Wang
C and Sang N: AMPK-HDAC5 pathway facilitates nuclear accumulation
of HIF-1α and functional activation of HIF-1 by deacetylating Hsp70
in the cytosol. Cell Cycle. 14:2520–2536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Stetler RA, Leak RK, Gan Y, Li P, Zhang F,
Hu X, Jing Z, Chen J, Zigmond MJ and Gao Y: Preconditioning
provides neuroprotection in models of CNS disease: Paradigms and
clinical significance. Prog Neurobiol. 114:58–83. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zhang F, Wu Y and Jia J: Exercise
preconditioning and brain ischemic tolerance. Neuroscience.
177:170–176. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Ding YH, Ding Y, Li J, Bessert DA and
Rafols JA: Exercise pre-conditioning strengthens brain
microvascular integrity in a rat stroke model. Neurol Res.
28:184–189. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Ding YH, Li J, Yao WX, Rafols JA, Clark JC
and Ding Y: Exercise preconditioning upregulates cerebral integrins
and enhances cerebrovascular integrity in ischemic rats. Acta
Neuropathol. 112:74–84. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Kang KA, Seong H, Jin HB, Park J, Lee J,
Jeon JY and Kim YJ: The effect of treadmill exercise on ischemic
neuronal injury in the stroke animal model: Potentiation of
cerebral vascular integrity. J Korean Acad Nurs. 41:197–203.
2011.(In Korean). View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Otsuka S, Sakakima H, Terashi T, Takada S,
Nakanishi K and Kikuchi K: Preconditioning exercise reduces brain
damage and neuronal apoptosis through enhanced endogenous 14-3-3γ
after focal brain ischemia in rats. Brain Struct Funct.
224:727–738. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Wang L, Deng W, Yuan Q and Yang H:
Exercise preconditioning reduces ischemia reperfusion-induced focal
cerebral infarct volume through up-regulating the expression of
HIF-1α. Pak J Pharm Sci. 28 (Suppl 2):S791–S798. 2015.
|
|
117
|
Wang H, Niu F, Fan W, Shi J, Zhang J and
Li B: Modulating effects of preconditioning exercise in the
expression of ET-1 and BNP via HIF-1α in ischemically injured
brain. Metab Brain Dis. 34:1299–1311. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Hacke W, Kaste M, Bluhmki E, Brozman M,
Dávalos A, Guidetti D, Larrue V, Lees KR, Medeghri Z, Machnig T, et
al: Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic
stroke. N Engl J Med. 359:1317–1329. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Wang H and Xu X, Yin Y, Yu S, Ren H, Xue Q
and Xu X: Catalpol protects vascular structure and promotes
angiogenesis in cerebral ischemic rats by targeting HIF-1α/VEGF.
Phytomedicine. 78:1533002020. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Liang C, Ni GX, Shi XL, Jia L and Wang YL:
Astragaloside IV regulates the HIF/VEGF/Notch signaling pathway
through miRNA-210 to promote angiogenesis after ischemic stroke.
Restor Neurol Neurosci. 38:271–282. 2020.PubMed/NCBI
|
|
121
|
Hu Q, Liu L, Zhou L, Lu H, Wang J, Chen X
and Wang Q: Effect of fluoxetine on HIF-1α-Netrin/VEGF cascade,
angiogenesis and neuroprotection in a rat model of transient middle
cerebral artery occlusion. Exp Neurol. 329:1133122020. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Wang J, Zhou X, Lu H, Song M, Zhao J and
Wang Q: Fluoxetine induces vascular endothelial growth
factor/Netrin over-expression via the mediation of
hypoxia-inducible factor 1-alpha in SH-SY5Y cells. J Neurochem.
136:1186–1195. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Zou J, Fei Q, Xiao H, Wang H, Liu K, Liu
M, Zhang H, Xiao X, Wang K and Wang N: VEGF-A promotes angiogenesis
after acute myocardial infarction through increasing ROS production
and enhancing ER stress-mediated autophagy. J Cell Physiol.
234:17690–17703. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Cheng X, Wang H, Liu C, Zhong S, Niu X,
Zhang X, Qi R, Zhao S, Zhang X, Qu H and Zhao C:
Dl-3-n-butylphthalide promotes remyelination process in cerebral
white matter in rats subjected to ischemic stroke. Brain Res.
1717:167–175. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zhang Q, Bian H, Guo L and Zhu H:
Berberine preconditioning protects neurons against ischemia via
sphingosine-1-phosphate and hypoxia-inducible factor-1[Formula: See
text]. Am J Chin Med. 44:927–941. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Ryou MG, Choudhury GR, Li W, Winters A,
Yuan F, Liu R and Yang SH: Methylene blue-induced neuronal
protective mechanism against hypoxia-reoxygenation stress.
Neuroscience. 301:193–203. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Wei Y, Hong H, Zhang X, Lai W, Wang Y, Chu
K, Brown J, Hong G and Chen L: Salidroside inhibits inflammation
through PI3K/Akt/HIF signaling after focal cerebral ischemia in
rats. Inflammation. 40:1297–1309. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Hou Y, Wang J and Feng J: The
neuroprotective effects of curcumin are associated with the
regulation of the reciprocal function between autophagy and HIF-1α
in cerebral ischemia-reperfusion injury. Drug Des Devel Ther.
13:1135–1144. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Saad MAE, Fahmy MIM, Al-Shorbagy M, Assaf
N, Hegazy AAE and El-Yamany MF: Nateglinide exerts neuroprotective
effects via downregulation of HIF-1α/TIM-3 inflammatory pathway and
promotion of caveolin-1 expression in the rat's hippocampus
subjected to focal cerebral ischemia/reperfusion injury.
Inflammation. 43:401–416. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Cheng CY, Ho TY, Hsiang CY, Tang NY, Hsieh
CL, Kao ST and Lee YC: Angelica sinensis exerts angiogenic
and anti-apoptotic effects against cerebral ischemia-reperfusion
injury by activating p38MAPK/HIF-1[Formula: See text]/VEGF-A
signaling in rats. Am J Chin Med. 45:1683–1708. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Wu S, Wang N, Li J, Wang G, Seto SW, Chang
D and Liang H: Ligustilide ameliorates the permeability of the
blood-brain barrier model in vitro during oxygen-glucose
deprivation injury through HIF/VEGF pathway. J Cardiovasc
Pharmacol. 73:316–325. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Chen ZZ, Gong X, Guo Q, Zhao H and Wang L:
Bu Yang Huan Wu decoction prevents reperfusion injury following
ischemic stroke in rats via inhibition of HIF-1α, VEGF and
promotion β-ENaC expression. J Ethnopharmacol. 228:70–81. 2019.
View Article : Google Scholar : PubMed/NCBI
|