Introduction
Acute mountain sickness (AMS) is a potentially
lethal condition caused by acute hypoxia after ascending to
altitudes higher than 2,500 m in a short time. It is common in
high-altitude travelers, and may lead to life-threatening
high-altitude conditions such as high-altitude cerebral edema
(HACE) or high-altitude pulmonary edema (HAPE) (1,2).
Previous studies suggested that hypoxia stress can
induce an oxidative stress reaction, which leads to increases in
endogenous reactive oxygen species (ROS), and oxidative stress
damage, resulting in body damage (3–5). ROS
are very active and unstable, and indirectly reflect the level of
oxidative stress, as well as the degree of injury through the
enzyme activity of superoxide dismutase (SOD), glutathione
peroxidase (GSH-Px) and the expression of malondialdehyde (MDA)
(6,7).
Previous studies demonstrated that oxidative stress
can induce the production of inflammatory factors (8). The presence of inflammatory cells,
immunoglobulins and complement components in the broncho-alveolar
lavage fluid of patients with HAPE suggests that inflammatory
immune responses can lead to increased permeability of the
pulmonary blood barrier and are involved in the occurrence of HAPE
(9). In addition, key factors such
as nuclear factor-κB (NF-κB), interleukin (IL)-1β, IL-6, and tumor
necrosis factor-α (TNF-α) serve an important role in the
development of exacerbated immune responses, among which IL-1β is
an important pro-inflammatory factor (10–13).
Activation of the above factors can stimulate the proliferation of
vascular smooth muscle cells, which can lead to pulmonary
hypertension (14–16). During oxidative stress, vascular
endothelial cells can release adhesion factors and monocytes,
causing lymphocytes to aggregate into endothelial cells through
these two inflammatory factors (17). TNF-α can activate neutrophils,
release a large number of oxygen-free radicals and proteases,
resulting in tissue damage (18).
NF-κB is a transcription regulator. Activation of NF-κB promotes
the transcription of cytokines and the activation of inflammatory
transmitter gene transcription (19,20).
IL-17C is a member of the IL-17 family. Previous studies
demonstrated that activated T cells do not synthesize IL-17C
(21), and that epithelial cells
from various tissues and organs are the main source of IL-17C
(22). IL-17C can promote the
secretion of IL-1β, IL-6, and TNF-α, activate the NF-κB pathway
(23), and induce inflammation and
immune regulation (24–26). As research has progressed,
microorganisms have been shown to promote the expression of IL-17C
in epithelial cells in inflammatory environments (27), yet the role of IL-17C and AMS has
not been established.
In the present study, it was hypothesized that
oxidative stress and inflammatory reactions induced by hypoxia
could serve an important role in the onset and development of AMS.
To verify this hypothesis and further explore the underlying
regulatory mechanism, an AMS rat model was used to study the role
and the mechanism of oxidative stress response and inflammatory
factors, such as IL-17C, in the occurrence and development of
AMS.
Materials and methods
Establishment of the animal model of
AMS
A total of 48 male Sprague-Dawley rats weighing
180–220 g and aged 12 weeks were purchased from the Laboratory
Animal Center of The Medical Department of Xi'an Jiaotong
University [SCXK (Shaanxi) 2018-001], and kept in a specific
pathogen-free at 20°C environment. Rats were provided with water
and food ad libitum under a 12-h light/dark cycle. Rats were
randomly divided into a control group (980 hPa) and simulated acute
hypoxic stress groups (475 hPa) for 24, 48 and 72 h (n=12 in each
group). The animals in the simulated hypoxia group were placed in a
hypobaric oxygen chamber at a simulated altitude of 6,000 m for the
aforementioned periods of time. The altitude in the chamber
increased at a uniform speed of 10 m/sec. The rats were given free
access to standard rodent food and water. Then, 30 mg/kg
pentobarbital sodium was injected intraperitoneally to anesthetize
the rats. When the breathing of the rat became slow and smooth and
muscles were loose, the tissue of both lungs was removed if no
traction reflex was observed, and the rat was then decapitated
(28). The morphological changes in
the right upper lobe lung tissue were detected using hematoxylin
and eosin (H&E) staining under light microscopy and
transmission electron microscopy. The remaining lung tissues were
immediately stored in three cryopreservation tubes and then
subjected to transcriptome sequencing and oxidative stress response
evaluation (measurement of SOD levels). The levels of MDA, GSH-Px
and inflammatory factors (IL-1β, IL-6, IL-17C, NF-κB, and TNF-α)
were also assessed. The measures for handling animals involved in
the sampling process were implemented in accordance with the
National Regulations on the Administration of Laboratory Animals
(GB14923-2010; http://www.cmu.edu.cn/sydwb/info/1835/1388.htm). The
animal experiment scheme was assessed and approved by the Ethics
Committee of School of Medicine of Qinghai University.
Histomorphological examination of lung
tissue
Tissue was harvested from the right upper lobe of
the lung, then fixed in 4% paraformaldehyde for 4 weeks at 4°C,
embedded in paraffin, and sectioned to 5-µm thickness. Lung
sections were concurrently stained with H&E (~95 min) for
histopathological examination. Images were captured using a light
microscope (magnification, ×400).
After dissection, the remaining fresh lung tissues
were immersion-fixed in 2.5% glutaraldehyde for 24 h at 4°C. The
samples were post-fixed in 1% osmium tetroxide for 1.5 h at 4°C,
dehydrated through a series of graded ethanol solutions and 1:1
EPON-812 epoxy resin, and then embedded in EPON-812 epoxy resin. 5
nm semi-thin sections were stained with toluidine blue for 10 min,
blocks trimmed, and ultrathin sections stained with lead citrate
and uranyl acetate at room temperature. Specimens were examined
using a transmission electron microscope (magnification, ×10,000 or
20,000).
RNA sequencing and differential gene
screening
Briefly, the total RNA of lung tissue was prepared
with an RNA TRIzol® reagent (Thermo Fisher Scientific,
Inc.) in accordance with the manufacturer's instructions and
agarose gel electrophoresis of extracted RNA performed to ensure
sample RNA integrity and inexistence of DNA contamination. Finally,
the differentially expressed genes were analyzed by functional
annotation and enrichment analysis using clusterProfiler software
(http://www.bioconductor.org/packages/release/bioc/html/clusterProfiler.html)
for Gene Ontology functional enrichment analysis of the
differential gene sets and Kyoto Encyclopedia of Genes and Genomes
(KEGG) (29) pathway enrichment
analysis and then the differentially related genes associated with
AMS in the IL-17 signaling pathway screened.
Reverse transcription-quantitative PCR
(RT-qPCR)
A total of 0.1 g lung tissue was ground in liquid
nitrogen and then 1 ml TRIzol® (Thermo Fisher
Scientific, Inc.) added to extract the total RNA. The cDNA was
obtained using a PrimeScript RT reagent kit with gDNA Eraser
(Takara Bio, Inc.). RT-qPCR was performed using a TB Green Premix
Ex Taq II (Takara Bio, Inc.) with a PIKORed 96 RT-PCR detection
system (Thermo Fisher Scientific, Inc.). All operations were
performed in accordance with the manufacturer's protocols. Relative
gene expression levels of IL-1, IL-6, IL-17C, NF-κB, and TNF-α were
determined using the comparative 2−ΔΔCq method using
β-actin as endogenous control (30). The thermocycling conditions were:
denaturation temperature 94°C for 30 sec, annealing temperature
56°C for 35 sec and extension temperature 72°C for 30 sec (36
cycles). The primer sequences in this study are shown in Table I. Each experiment was repeated more
than three times per tissue to ensure consistency of the
experimental results.
 | Table I.Primer sequences for IL-17C, NF-κB,
IL-1β, IL-6 and TNF-α. |
Table I.
Primer sequences for IL-17C, NF-κB,
IL-1β, IL-6 and TNF-α.
| Analyte | Forward primer
sequence, 5′-3′ | Reverse primer
sequence, 5′-3′ |
|---|
| IL-1β |
AAACAGATGAAGTGCTCCTTCCAGG |
TGGAGAACACCACTTGTTGCTCCA |
| IL-17C |
GCCTATTTGCCCACCTACAA |
AAATTCAGACGGCAAACGAC |
| IL-6 |
TGTGTGAAAGCAGCAAAG |
AGTCTCCTCATTGAATCCA |
| TNF-α |
CGATGAACCACGCCAGTCGCC |
GGATGAACACGCCAGTCGCC |
| NF-κB |
CGACGTATTGCTGTGCCTTC |
TTGAGATCTGCCCAGGTGGTA |
| β-actin |
GAGACCTTCAACACCCAGCC |
GCGGGGCATCGGAACCGCTCA |
Western blot analysis
The protein expressions of IL-1, IL-6, IL-17C,
NF-κB, and TNF-α in lung tissues were detected by WB. The lung
tissue was homogenized in RIPA containing protease and phosphatase
inhibitors (Beyotime Institute of Biotechnology) and lysed on ice
for 10 min. The lysate was centrifuged at 12,000 × g at 4°C for 10
min to collect the supernatant. The total protein concentration was
detected using the BCA protein quantification kit (Beyotime
Institute of Biotechnology). Subsequently, protein sample was
separated on 7% SDS-PAGE gel with 40 µg protein loaded per lane and
transferred onto PVDF membranes (Sigma-Aldrich; Merck KGaA). PVDF
membrane was placed in 5% skimmed milk diluted with TBST (0.1%
Tween) Buffer, incubation box and agitated for 2 h. The PVDF
membrane was washed three times with TBST for 5 min each. The above
operations were all performed at room temperature. The primary
antibodies anti-β-actin (1:100,000; cat. no. AC026; Abclonal),
anti-IL-1 (1:1,000; cat. no. ab234437; Abcam), anti-IL-6 (1:1,000;
cat. no. ab9324; Abcam), anti-IL-17C (1:1,000; cat. no. bs26112;
BIOSS), anti-NF-κB, (1:1,000; cat. no. ab32536; Abcam), and
anti-TNF-α (1:1,000; cat. no. ab55275; Abcam) were used to detect
the relative protein expression in samples and the secondary
antibody was Goat Anti-Rabbit IgG H&L (1:5,000; cat. no.
ab6721; Abcam). Images were acquired using Tanon GIS chassis
control software v2.0 (Shanghai, China).
Detection of relevant indicators of
oxidative stress
The levels of GSH-Px, SOD and MDA in lung tissue
were determined using ELISA kits purchased from Shanghai Enzyme
Union Biotechnology Co. Ltd. (cat. nos. ml059387-C, ml097316-C and
ml077384-C). Standard curve pore and detection pore were
established. Optical density was measured at 450 nm wavelength
using a microplate reader. GSH-Px, SOD and MDA levels were
calculated using a standard curve.
Statistical analysis
All data were analyzed using SPSS 22.0 (IBM Corp.)
statistical analysis software, and shown as mean ± standard
deviation (SD). One-way analysis of variance (ANOVA) was used with
the Tukey HSD post hoc test. Two-tailed Student's t-test were
applied to analyze the significant differences between the groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Morphological changes in rat lung
tissue at different time points following hypoxic stress
Lung tissue of the 24 h group (Fig. 1B) showed thickening of the alveolar
septum and inflammatory cell infiltration not visible in the
control group (Fig. 1A). The
alveolar structure of the 48-h group was severely damaged (Fig. 1C), with a large number of
inflammatory cells and red blood cells diffused in the field of
vision. Lung tissue injury was still apparent in the 72-h group
(Fig. 1D). However, alveolar
structure showed none of the damage visible in the 48-h group,
suggesting recovery from hypoxic stress at the 72-h time point.
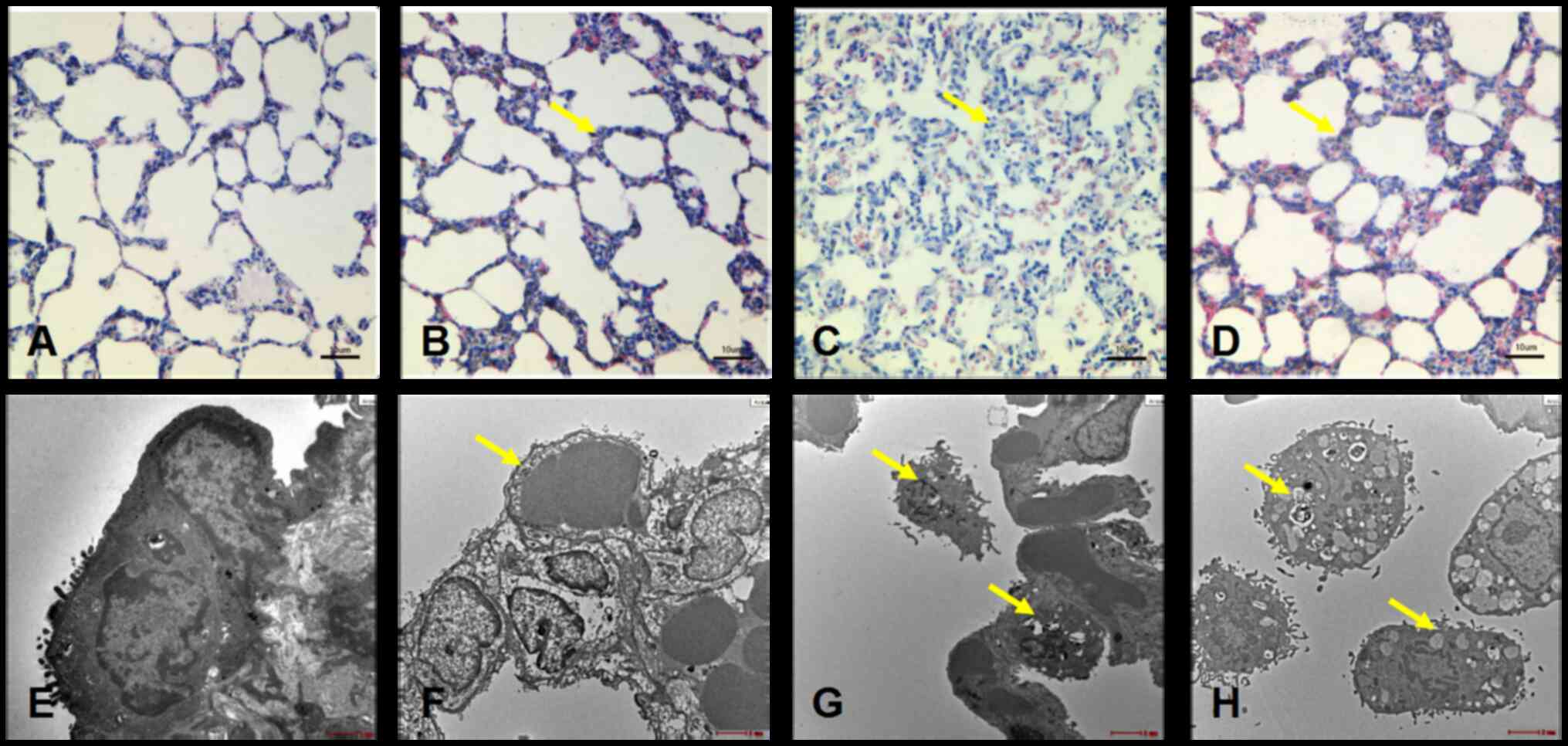 | Figure 1.Histomorphological changes in the
lungs of Sprague-Dawley rats of different groups. Light microscopy
images for the (A) 0 h group, (B) 24 h group, (C) 48 h group and
(D) 72 h group. Scale bar, 10 µm. Electron microscope for the (E) 0
h group, scale bar, 1 µm, (F) 24 h group, scale bar, 500 nm, (G) 48
h group, scale bar, 1 µm) and (H) 72 h group, scale bar, 500 nm.
Pathological changes are indicated by arrows. |
Electron microscopy demonstrated that the thickness
of the basement membrane of the capillary increased, compared with
the control group (Fig. 1E) to
different extents. The control group was also used as a marker to
measure the pathological damage of each group of rats. In the lung
tissue of the 24 h group, the thickness of the capillary basement
membrane increased to varying degrees. Alveolar septum capillary
thickness also increased, and microvilli shedding was observed
(Fig. 1F). In the 48 h group,
alveolar wall capillary endothelial cell edema became more apparent
(Fig. 1G). In the 72 h group, basal
membrane partial shedding and blood-air barrier obviously thinning
are still evident (Fig. 1H).
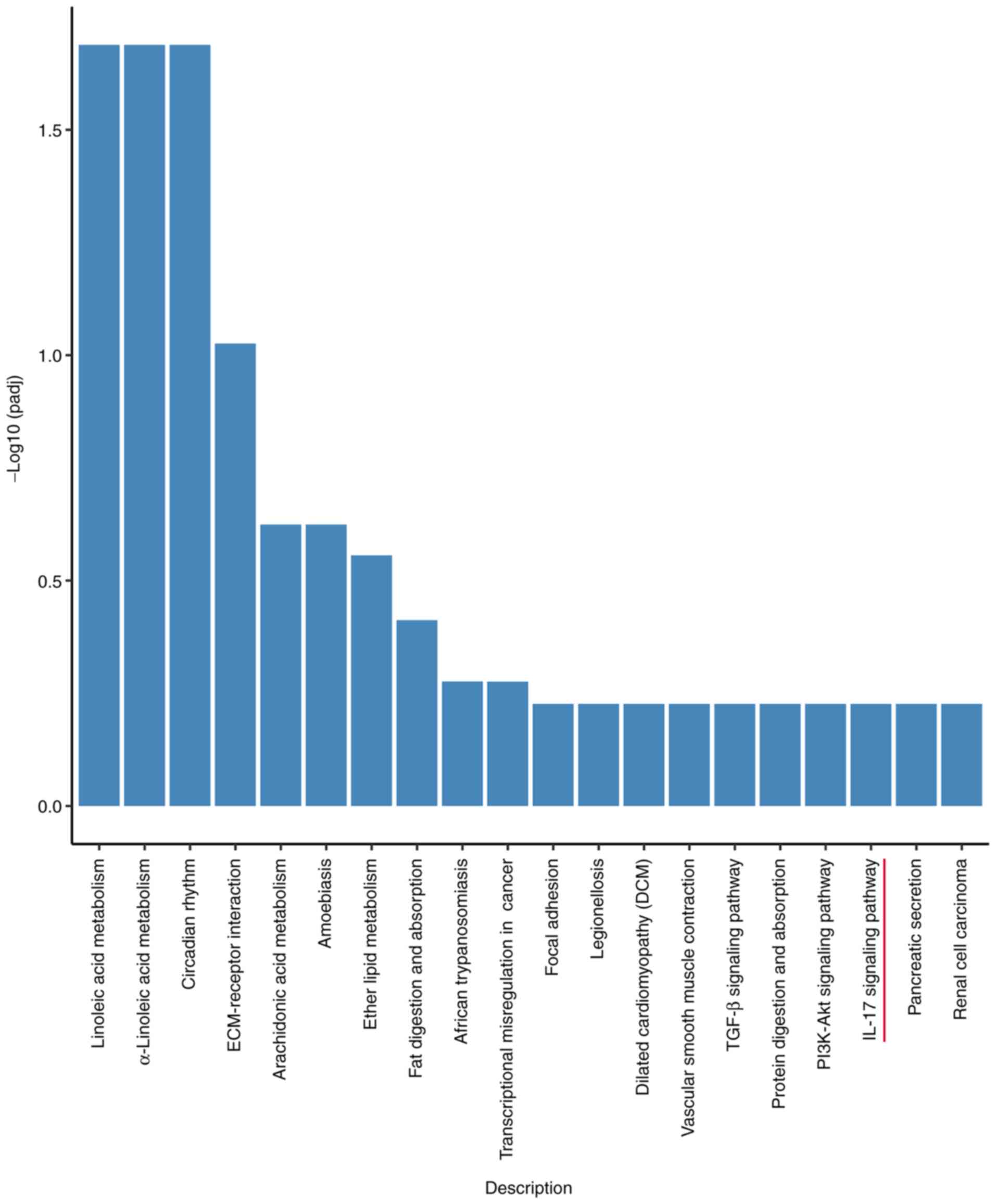
Transcriptome sequencing results
RNA sequencing analysis of lung tissues from rats in
each group identified 1,728 differentially expressed genes in the
24 h, and 2,148 differentially expressed genes were found in the 48
h group, and there were 8,201 genes of 72 h group differentially
expressed relative to the control group. The differentially
expressed genes were obtained by KEGG (Fig. 2). The present study focused on the
IL-17 signaling pathway. The main differentially expressed genes in
this pathway were IL-17C, NF-κB, IL-1β, IL-6, and TNF-α.
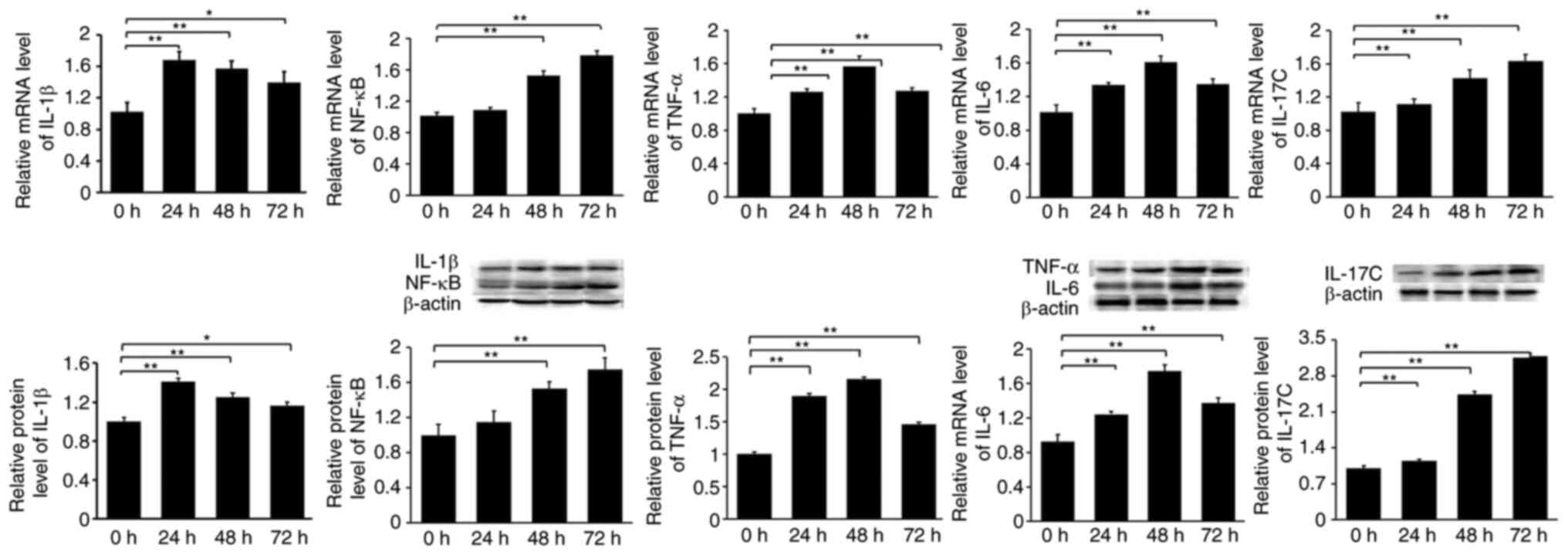
Expression of IL-1β, IL-6, IL-17C,
NF-κB, and TNF-α in rats subjected to 24, 48 or 72 h of hypoxic
stress
The expression of IL-17C was significantly
upregulated in the 24, 48 and 72 h groups, compared with the
control group. NF-κB expression was a significantly upregulated
after 48 and 72 h of hypoxia. The expression of IL-1β, TNF-α and
IL-6 was also significantly upregulated at all time points
(Fig. 3).
Levels of GSH-Px, SOD and MDA in rats
under hypoxic stress in different groups
The activity of SOD and GSH-Px decreased
significantly after 24, 48 and 72 h of hypoxia (P<0.01), while
the expression of MDA increased significantly after 48 and 72 h of
hypoxic stress (P<0.01; Table
II).
| Table II.Experimental results of oxidative
stress in rats of each group. |
Table II.
Experimental results of oxidative
stress in rats of each group.
| Group | n | GSH-Px (mg/l) | SOD (Um/l) | MDA (nmol/ml) |
|---|
| 0
h | 6 | 4.18±0.16 | 2.62±0.11 | 0.91±0.04 |
| 24 h | 6 |
3.63±0.12a |
2.31±0.15a | 0.91±0.02 |
| 48 h | 6 |
3.47±0.25a |
2.25±0.18a |
0.95±0.04a |
| 72 h | 6 |
3.93±0.36a |
2.36±0.16a |
0.95±0.04a |
Discussion
After rapid exposure to a hypoxic environment, a
series of reactions as oxidative stress response occur to adapt to
the hypoxic environment and even damage the lung tissue. The main
manifestations include diffuse swelling and hyperemia of both
lungs, considerable leakage of protein-rich liquid into the
alveolar cavity, thickening of the alveolar cavity (31), increasing the activity of
macrophages and infiltration of inflammatory cells (32). Luo et al (33) and Guo et al (34) demonstrated that lung tissue injury
was most serious after exposure to a simulated altitude of 5,000 m
and 48 h of hypoxia. Li et al (35) also suggested that lung tissue injury
was the most serious after exposure to a simulated altitude of
6,000 m and hypoxic stress lasting 72 h. In the present study,
Sprague-Dawley rats were exposed to a simulated altitude of 6,000 m
and hypoxia stress lasting 24, 48 or 72 h. Morphological
examination of the lung tissues in each group suggested that the
alveolar structure of rats in hypoxic stress groups was damaged to
varying degrees. Lung tissue was damaged most severely after
hypoxic stress lasting 48 h.
The production of ROS and the elimination of
antioxidants create a stable dynamic balance. When the organism is
subjected to hypoxic stress, oxygen-free radicals accumulate in
cells and damage lipids, proteins and DNA, thus causing lung tissue
damage (36–38). The present study suggested that the
activities of SOD and GSH-Px decreased and the expression of MDA
increased as hypoxic stress was prolonged; that is, oxidative
damage occurred in lung tissue under hypoxic stress.
Oxidative damage can directly stimulate macrophages
to release IL-1β. As a pro-inflammatory factor, IL-1β can initiate
immune response and recruit centralized granulocytes, macrophages
and other inflammatory cells, which in turn can to infiltrate
tissues and release IL-6 and TNF-α (39). The results of the present study
demonstrated that IL-1β levels were highest after 24 h of hypoxic
stress, while IL-6 and TNF-α expression was highest after 48 h of
hypoxic stress. GSH-Px and SOD activity were the lowest after 48 h
of hypoxia stress, and MDA was highly expressed, suggesting that
inflammation and the exudation of tissue fluid in lung tissue were
further aggravated. The injury to alveolar epithelial cells and
pulmonary interstitial edema was most pronounced at 48 h. Previous
studies demonstrated that IL-17C was not produced by activated T
cells (22); rather, it is secreted
by the epithelial cells of various tissues (40). In the present study, it was
hypothesized that, given the aforementioned three factors,
pulmonary epithelial cells could also participate in the immune
response. The presence of large quantities of IL-17C activates the
NF-κB pathway and participates in the onset and development of AMS.
According to the experimental results of the present study, IL-17C
stimulation could increase the expression of NF-κB in the AMS rat
model. The combined stimulation of IL-17C and NF-κB also increased
the expression of IL-17C, NF-κB, TNF-α, IL-1β, and IL-6 in alveolar
epithelial cells, which also reflects the positive feedback
regulation of IL-17C on the acute inflammatory response of
pulmonary epithelial cells.
The innovation of the current study was the
combination of hypoxia-induced oxidative stress and excessive
inflammation to study the damage to the body. Under hypoxic stress,
rats quickly initiate the oxidative stress response and immune
response, but it is worth noting that, with the prolongation of
hypoxic stress time, excessive oxidative stress can further
stimulate the immune system, and release a large number of
inflammatory factors, which accumulate in the body, and even lead
to the occurrence of inflammatory storms.
According to the literature, IL-17C can also
regulate the expression of IL-1β, TNF-α, and IL-6. However, the
expression of these three factors was downregulated in the 72-h
group. The specific molecular mechanism needs further study, and it
may be related to the immune regulatory function of
1,25-dihydroxyvitamin D3 (41),
studied earlier by our group.
Acknowledgements
Not applicable.
Funding
The present study was supported by The Second Tibetan Plateau
Scientific Expedition and Research Program (grant no.
2019QZKK0606), The Chinese National Natural Science Foundation
(grant no. 81460051), The Key Laboratory of Medicinal Animal and
Plant resources of The Qinghai-Tibetan Plateau (grant no.
2017-ZJ-Y13), The Department of Science and Technology of Qinghai
Province (grant no. 2019-ZJ-7042), The Scientific Research for
Middle-aged and Young people of the Medical College of Qinghai
University (grant no. 2018-kyy-1), and The National Students'
Platform for Innovation and Entrepreneurship Training Program
(grant no. 201910743003).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
XP contributed to the idea of the study and served
an important role in interpreting the results. FL contributed
significantly to analysis and manuscript preparation. XL performed
data analysis and wrote the manuscript. ZC and RW helped perform
the analysis with constructive discussion. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The animals involved in the present study were
handled in accordance with the National Regulations on the
Administration of Laboratory Animals (GB14923-2010). The animal
experiment scheme was approved and examined by the Ethics Committee
of School of Medicine of Qinghai University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tang E, Chen Y and Luo Y: Dexamethasone
for the prevention of acute mountain sickness: Systematic review
and meta-analysis. Int J Cardiol. 173:133–138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guo P, Luo H, Fan Y, Luo Y and Zhou Q:
Establishment and evaluation of an experimental animal model of
high altitude cerebral edema. Neurosci Lett. 547:82–86. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Swenson ER, Maggiorini M, Mongovin S,
Gibbs JS, Greve I, Mairbäurl H and Bärtsch P: Pathogenesis of
high-altitude pulmonary edema: Inflammation is not an etiologic
factor. JAMA. 287:2228–2235. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao S, Zhang L, Lian G, Wang X, Zhang H,
Yao X, Yang J and Wu C: Sildenafil attenuates LPS-induced
pro-inflammatory responses through down-regulation of intracellular
ROS-related MAPK/NF-κB signaling pathways in N9 microglia. Int
Immunopharmacol. 11:468–474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wyse C, Cathcart A, Sutherland R, Ward S,
McMillan L, Gibson G, Padgett M and Skeldon K: Effect of maximal
dynamic exercise on exhaled ethane and carbon monoxide levels in
human, equine, and canine athletes. Comp Biochem Physiol A Mol
Integr Physiol. 141:239–246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Balan DJ, Rajavel T, Das M, Sathya S,
Jeyakumar M and Devi KP: Thymol induces mitochondrial
pathway-mediated apoptosis via ROS generation, macromolecular
damage and SOD diminution in A549 cells. Pharmacol Rep. 73:240–254.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma Y, Wang D, Xu X, Yang X, Wang X, Zhu Z,
Zhao Y, Chen M, Xu F, Fu L, et al: Dynamic changes of ROS, MDA and
SOD during arsenic-induced neoplastic transformation in human
keratinocytes. Wei Sheng Yan Jiu. 44:456–461. 2015.(In Chinese).
PubMed/NCBI
|
|
8
|
Zi Y, Jiang B, He C and Liu L: Lentinan
inhibits oxidative stress and inflammatory cytokine production
induced by benzo(a)pyrene in human keratinocytes. J Cosmet
Dermatol. 19:502–507. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Altamura S, Bärtsch P, Dehnert C,
Maggiorini M, Weiss G, Theurl I, Muckenthaler MU and Mairbäurl H:
Increased hepcidin levels in high-altitude pulmonary edema. J Appl
Physiol (1985). 118:292–298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gao H, Liu L, Zhao Y, Hara H, Chen P, Xu
J, Tang J, Wei L, Li Z, Cooper DK, et al: Human IL-6, IL-17, IL-1β,
and TNF-α differently regulate the expression of pro-inflammatory
related genes, tissue factor, and swine leukocyte antigen class I
in porcine aortic endothelial cells. Xenotransplantation. 24:2017.
View Article : Google Scholar
|
|
11
|
Sun Y, Hu G, Zhang X and Minshall RD:
Phosphorylation of caveolin-1 regulates oxidant-induced pulmonary
vascular permeability via paracellularand transcellular pathways.
Circ Res. 105:676–685. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shukla D, Saxena S, Jayamurthy P, Sairam
M, Singh M, Jain SK, Bansal A and Ilavazaghan G: Hypoxic
preconditioning with cobalt attenuates hypobaric hypoxia-induced
oxidative damage inratlungs. High Alt Med Biol. 10:57–69. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gonzalez NC and Wood JG: Leukocyte
endothelial interactions in environmental hypoxia. Adv Exp Med
Biol. 502:39–60. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao-Lin MA, Jin RB, Zhang XH, Cui GY, Ren
XJ, Shi H and Sun YQ: Study on management of 726 victims in
military hospitals following Yushu earthquake in Qinghai province.
J Traumat Surg. 4:339–343. 2010.(In Chinese).
|
|
15
|
Tang C, Luo Y, Li S, Huang B, Xu S and Li
L: Characteristics of inflammation process in monocrotaline-induced
pulmonary arterial hypertension in rats. Biomed Pharmacother.
133:1110812021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han Z, Li X, Cui X, Yuan H and Wang H: The
roles of immune system and autoimmunity in pulmonary arterial
hypertension: A review. Pulm Pharmacol Ther 102094. Nov
2–2021.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu DL, Yu YX, Liang R, Zhou SY, Duan SL,
Jiang ZY, Meng CY, Jiang W, Wang H, Sun YX and Fang LS: Regulation
of hypoxia inducible factor-1α on permeability of vascular
endothelial cells and the mechanism. Zhonghua Shao Shang Za Zhi.
35:209–217. 2019.(In Chinese). PubMed/NCBI
|
|
18
|
Zhou Q, Luo Y, Li H, Li S, Zhang X, Gao W,
Zheng B, Yang D, Liu F and Yuqi G: Epidemiological study of
mountain sickness complicated with multiple organ dysfunction
syndrome on the Qinghai-Tibetan Plateau: report of 103 cases. Sci
Res Essays. 5:2010.
|
|
19
|
Althubiti M, Rada M, Samuel J, Escorsa JM,
Najeeb H, Lee KG, Lam KP, Jones GD, Barlev NA and Macip S: BTK
modulates p53 activity to enhance apoptotic and senescent
responses. Cancer Res. 76:5405–5414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Barbarulo A, Grazioli P, Campese AF,
Bellavia D, Di Mario G, Pelullo M, Ciuffetta A, Colantoni S, Vacca
A, Frati L, et al: Notch3 and canonical NF-kappaB signaling
pathways cooperatively regulate Foxp3 transcription. J Immunol.
186:6199–6206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li H, Chen J, Huang A, Stinson J, Heldens
S, Foster J, Dowd P, Gurney AL and Wood WI: Cloning and
characterization of IL-17B and IL-17C, two new members of the IL-17
cytokine family. Proc Natl Acad Sci USA. 97:773–778. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Song X, He X, Li X and Qian Y: The roles
and functional mechanisms ofinterleukin-17 family cytokines in
mucosal immunity. Cell Mol Immunol. 13:418–431. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Song X, Zhu S, Shi P, Liu Y, Shi Y, Levin
SD and Qian Y: IL-17RE is the functional receptor for IL-17C and
mediates mucosal immunity to infection with intestinal pathogens.
NatImmunol. 12:1151–1158. 2011.
|
|
24
|
Akitsu A and Iwakura Y:
Interleukin-17-producing γδ T (γδ17) cells in inflammatory
diseases. Immunology. 155:418–426. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamaguchi Y, Fujio K, Shoda H, Okamoto A,
Tsuno NH, Takahashi K and Yamamoto K: IL-17B and IL-17C are
associated with TNF-alpha production and contribute to the
exacerbation of inflammatory arthritis. J Immunol. 179:7128–7136.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ruiz de Morales JMG, Puig L, Daudén E,
Cañete JD, Pablos JL, Martín AO, Juanatey CG, Adán A, Montalbán X,
Borruel N, et al: Critical role of interleukin (IL)-17 in
inflammatory and immune disorders: An updated review of the
evidence focusing in controversies. Autoimmun Rev. 19:1024292020.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu S, Luo X, Yang B, Xiao L, Wu X, Li H
and Wu C: Poly-functional T helper cells in human tonsillar
mononuclear cells. Eur Cytokine Netw. 30:114–122. 2019.PubMed/NCBI
|
|
28
|
De-Doncker L, Picquet F, Petit J and
Falempin M: Characterization of spindle afferents in rat soleus
muscle using ramp-and-hold and sinusoidal stretches. J
Neurophysiol. 89:442–449. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ryan CF, Lowe AA and Fleetham JA: Nasal
continuous positive airway pressure (CPAP) therapy for obstructive
sleep apnea in Hallermann-Streiff syndrome. Clin Pediatr (Phila).
29:122–124. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Maggiorini M, Mélot C, Pierre S, Pfeiffer
F, Greve I, Sartori C, Lepori M, Hauser M, Scherrer U and Naeije R:
High-altitude pulmonary edema is initially caused by an increase in
capillary pressure. Circulation. 103:2078–2083. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Luo X, Guo W, Xu R, et al: Effect of high
altitude and hypoxia in simulated environment on HPT axis and
expression of VEGF and HIF-1 in lung tissues of rats. Acad J Chin
PLA Med Sch. 37:864–868. 2016.(In Chinese).
|
|
34
|
Guo W: The dynamic observation of
hypothalamic-pituitary-adrenala axis/hypothalamic-pituitary-thyroid
axis stress and the effect of lung and brain tissue under simulated
high altitude hypoxia. Gan Su University of Chinese Medicine; 2016,
(In Chinese).
|
|
35
|
Li Y: The research of hypoxia related
genes in high altitude pulmonary edema. Qing Hai University; 2017,
(In Chinese).
|
|
36
|
Brandon M, Baldi P and Wallace DC:
Mitochondrial mutations in cancer. Oncogene. 25:4647–4662. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Morrow JD: Quantification of isoprostanes
as indices of oxidant stress and the risk of atherosclerosis in
humans. Arterioscler Thromb Vasc Biol. 25:279–286. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou L, Aon MA, Almas T, Cortassa S,
Winslow RL and O'Rourke B: A reaction-diffusion model of
ROS-induced ROS release in a mitochondrial network. PLoS Comput
Biol. 6:e10006572010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xiong R, Jiang W, Li N, Liu B, He R, Wang
B and Geng Q: PM2.5-induced lung injury is attenuated in
macrophage-specific NLRP3 deficient mice. Ecotoxicol Environ Saf.
221:1124332021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ramirez-Carrozzi V, Sambandam A, Luis E,
Lin Z, Jeet S, Lesch J, Hackney J, Kim J, Zhou M, Lai J, et al:
IL-17C regulates the innate immune function of epithelial cells in
an autocrine manner. Nat Immunol. 12:1159–1166. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Uberti F, Lattuada D, Morsanuto V, Nava U,
Bolis G, Vacca G, Squarzanti DF, Cisari C and Molinari C: Vitamin D
protects human endothelial cells from oxidative stress through the
autophagic and survival pathways. J Clin Endocrinol Metab.
99:1367–1374. 2014. View Article : Google Scholar : PubMed/NCBI
|