Introduction
Globally, the estimated statistics for the year 2018
relating to 36 different types of cancer in 185 countries revealed
that lung cancer remains the leading cause of cancer incidence and
mortality, with reported percentages of 11.6% of the total cases
and 18.4% of the total cancer deaths, respectively (1). Histological classification
distinguishes two main subtypes of lung cancer: Small cell lung
cancer (SCLC) and non-SCLC (NSCLC). The latter has an incidence
rate exceeding 85% (2).
Metastases are a frequent complication of NSCLC, which most
commonly spreads to the bones, brain, liver and adrenal glands
(2). As the majority of patients
with metastatic NSCLC are inoperable at the time of diagnosis, the
prognosis is generally poor, with a 5-year survival rate of <20%
(3).
Epithelial-mesenchymal transition (EMT) is an
evolutionarily conserved developmental program that confers
metastatic properties to malignancies by enhancing the adhesion,
migration and invasion of cancer cells (4,5).
The EMT encompasses dynamic changes in cellular organization,
leading to the loss of epithelial markers and the gain of
mesenchymal phenotypes. E-cadherin is among the most important
molecules that regulate cell-cell adhesion in epithelial tissues.
By contrast, N-cadherin and vimentin are mesenchymal hallmarks that
represent the acquisition of an aggressive tumor phenotype
(6). Sirtuin-1 (SIRT1) is also an
evolutionarily conserved, NAD+-dependent class III
histone deacetylase that plays a key role in epigenetic regulation
by deacetylating both histone and non-histone targets (7). SIRT1 has gained significant interest
due to its ability to promote genomic stability and metabolic
regulation; however, mounting evidence has suggested an oncogenic
role for SIRT1 (8). Particularly,
SIRT1 is associated with tumor metastasis via the positive
regulation of EMT, promoting the migratory capability of cancer
cells in vitro and tumor metastasis in vivo (9).
B7-H3 (CD276), a member of the B7 superfamily, is
upregulated on different types of malignant cells, including NSCLC
cells (10–13). The membrane-bound B7-H3 not only
inhibits tumor-specific T cell activation (14,15), but it also transduces
intracellular signals to promote tumor cell migration and invasion,
angiogenesis, drug resistance and the Warburg effect (16–21). Therefore, B7-H3 may act as a novel
target for cancer immunotherapy (17–22). B7-H3 has been shown to promote the
EMT process of NSCLC cells (16);
however, the underlying signaling mechanisms have yet to be fully
elucidated. Therefore, the aim of the present study was to analyze
the potential role exerted by SIRT1 in B7-H3-mediated EMT, and the
possible mechanisms through which B7-H3-induced signaling may
regulate SIRT1 expression in NSCLC A549 cells.
Materials and methods
Cell lines and cell culture
The A549 NSCLC cell line (cat. no. FH0045) was
obtained from the Cell Culture Center of FuHeng Biology, and the
A549 cells were maintained in Invitrogen® RPMI-1640
medium (Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Lonsera; Shanghai Shuangru Biotechnology Co., Ltd.) and penicillin
(100 IU/ml)/streptomycin (100 µg/ml) (MedChemExpress). The A549
cell line was authenticated by using short tandem repeat (STR)
analysis in combination with sex-typing gene amelogenin detection,
and the A549 cell line was compared with the DSMZ STR cell line
profile before use. The cells were incubated in a humidified
atmosphere containing 5% CO2 at 37°C.
Genome editing of B7-H3 and gene
silencing of SIRT1
Knockout (KO) of the B7-H3 gene in the
A549 cell line was performed using CRISPR/Cas9 guide constructs, on
the basis of a previously published protocol (23). The sequence of single guide RNA
(sgRNA) was TTGATGTGCACAGCGTCCTGCGG, which was designed by using
the online tool available from ZHANG LAB (https://zlab.bio/guide-design-resources). The target
sequences for sgRNA were the N-terminal 250 bp of exon 4 (315 bp),
which encode the transmembrane domain of B7-H3. The
sgRNA-expressing plasmid was constructed by annealing the custom
sgRNA (Sangon Biotech Co., Ltd.) to a lentiviral vector
U6-MCS-sgRNA-SV40-EGFP (cat. no. GV504) developed by Genechem, Inc.
The lentivirus expressing only Cas9 was used to generate negative
control (mock) KO cells. SIRT1 knockdown was performed by
transfection of specific small interfering RNAs (siRNAs) using
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The sequences of the
on-target siRNAs were as follows: siSIRT1 #1,
5′-GCGGGAAUCCAAAGGAUAATT-3′ and siSIRT1 #2,
5′-UUAUCCUUUGGAUUCCCGCTT-3′. Those of the negative control siRNAs
oligos were: siControl #1, 5′-UUCUCCGAACGUGUCACGUTT-3′ and
siControl #2, 5′-ACGUGACACGUUCGGAGAATT-3′. All the oligos were
purchased from Shanghai GenePharma Co., Ltd. The final
concentration of Lipofectamine 3000 and siRNAs were 2 µl/ml and 50
nM, respectively. The siRNA-transfected A549 cells were incubated
at 37°C in 5% CO2, and SIRT1 knockdown was verified at
48 h post-transfection via western blotting, as described
below.
Cell proliferation assay
B7-H3 KO and mock A549 cells were cultured
for 0, 24, 48 and 72 h, and the fold-change of cell proliferation
was assayed by quantitation of the uptake and digestion of
WST-8/Cell Counting Kit-8 (CCK-8) according to the manufacturer's
instructions (Dojindo Laboratories, Inc.). Cell proliferation at 0
h was normalized to 1.0. All experiments were performed in
triplicate.
Western blot analysis
B7-H3 KO and mock-edited, siSIRT1 and
siControl A549 cells were left untreated, whereas the wild-type
A549 cells were either treated with the PI3K-specific inhibitor,
LY294002 (50 µΜ; MedChemExpress), or DMSO control at 37°C for 24 h.
Cells were lysed with a radio-immunoprecipitation assay (RIPA)
buffer containing 1 mM protease inhibitor phenylmethylsulfonyl
fluoride (PMSF) (cat. no. R0020; Beijing Solarbio Science &
Technology Co., Ltd.), and 1% (v/v) protein phosphatase inhibitor
(All-in-one) mixture (cat. no. P1260; Beijing Solarbio Science
& Technology Co., Ltd.). The total cell lysate was measured by
bicinchoninic acid (BCA) assay and 30 µg protein was separated via
SDS-PAGE (12% running, 5% stacking). The separated proteins were
then transferred to a polyvinylidene difluoride (PVDF) membrane,
blocked with 5% (w/v) skimmed milk in Tris-HCL buffer solution
(TBS) for 2 h at room temperature, and incubated at 4°C overnight
with primary antibodies against human B7-H3/CD276 (cat. no.
ab227670, 1:100), E-cadherin (cat. no. ab40772, 1:10,000),
N-cadherin (cat. no. ab76011, 1:5,000), vimentin (cat. no. ab92547,
1:1,000), total (t)-ERK1/2 (cat. no. ab54230, 1:1,000),
phosphorylated (p)-ERK1/2 (T202+T204; cat. no. ab214362, 1:1,000)
and t-STAT-3 (cat. no. ab109085, 1:5,000) (all these antibodies
were purchased from Abcam); or with anti-p-STAT-3 (Tyr705; cat. no.
AF3293, 1:1,000), anti-GAPDH (cat. no. AF7021, 1:3,000) (both from
Affinity Biosciences, Ltd.), anti-SIRT1 (cat. no. 2493, 1:1,000),
anti-t-AKT (cat. no. 4691, 1:1,000) and anti-p-AKT (Ser473; cat.
no. 4060, 1:2,000) (the latter three antibodies were purchased from
Cell Signaling Technology, Inc.). Subsequently, HRP-conjugated goat
anti-rabbit IgG (H+L) (cat. no. S0001, 1:5,000; Affinity
Biosciences, Ltd.) was used as the secondary antibody and incubated
for 1 h at room temperature in the dark. The membranes were washed
four times with TBS with 0.1% (v/v) Tween-20 and then incubated
with ECL substrate (Thermo Fisher Scientific, Inc.), and visualized
using the Tanon Fine-Do X6 Chemi-Image System (Tanon Science and
Technology Co., Ltd.). The expression levels of the signaling
proteins were assessed using ImageJ software (v1.8.0; National
Institutes of Health), and were normalized against GAPDH. All
experiments were performed in triplicate.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from B7-H3 KO and
mock, and SIRT1 silencing and siControl A549 cells with
RNAiso Plus (Takara Bio, Inc.) according to the manufacturer's
instructions. Next, cDNA was synthesized using PrimeScript RT
Master Mix reverse transcription kit (Takara Bio, Inc.), according
to the manufacturer's instructions, and qPCR was performed with
QuantStudio 5 (Thermo Fisher Scientific, Inc.) by using TaqMan gene
expression assay kit/probe sets (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocols. The primers used in this
study were as follows: B7-H3 forward (F),
5′-AGGGCAGCCTATGACATTCCC-3′ and reverse (R),
5′-AGCTCCTGCATTCTCCTCCTCA-3′; SIRT1 F, 5′-ACCTTCTGTTCGGTGATG-3′ and
R, 5′-TATGGACCTATCCGTGGC-3′; and β-actin F,
5′-GGAAGGTGAAGGTCGGAGTC-3′ and R, 5′-CGTTCTCAGCCTTGACGGT-3′. The
thermocycling conditions included an initial denaturation at 94°C
for 10 min; followed by 40 cycles of denaturation at 95°C for 15
sec, annealing at 55°C for 15 sec and extension at 72°C for 1 min.
The relative expression level of B7-H3 and SIRT1 mRNA was
normalized against β-actin expression and calculated using the
2−ΔΔCq method (24).
Cell migration and invasion
assays
For the cell migration assay, 5×104 A549
cells from different groups were resuspended in 200 µl serum-free
medium and placed in the upper Transwell chamber (Corning, Inc.),
and 600 µl medium containing 30% FBS was added to the lower
chamber. Cells were incubated at 37°C in an atmosphere of 5%
CO2, and allowed to migrate for 24 h. Subsequently, the
migrated cells on the lower surface of the membrane were fixed with
4% (w/v) paraformaldehyde in PBS buffer for 40 min and stained with
0.1% crystal violet for 15 min (both at room temperature). The
migrated cells were then counted, and images were captured at a
magnification of ×100 using a light microscope (Olympus
Corporation). In total, five random fields of the membrane in each
well were counted, and the average number/field from triplicate
wells was plotted. For the cell invasion assay, a Transwell chamber
(Corning, Inc.) was pre-coated with 50 µl Matrigel matrix (BD
Biosciences) in serum-free medium for 3 h at 37°C and 5%
CO2, and 1×105 A549 cells from the different
treatment groups were placed in the upper chamber. After 48 h, the
lower surface of the membrane was fixed with 4% (w/v)
paraformaldehyde, stained with 0.1% crystal violet and counted as
described above for the cell migration assay. These experiments
were performed three times.
Statistical analysis
A549 cells in the different experimental treatment
groups (B7-H3 KO, siSIRT1 or LY294002-treated) were
compared with the corresponding control groups, where indicated.
The results are presented as the mean ± SEM of three representative
experiments, where the significance was calculated using an
unpaired two-tailed Student's t-test. Statistical analyses were
performed using SPSS software version 20.0 (IBM Corp.). P<0.05
was considered to indicate a statistically significant
difference.
Results
B7-H3 ablation reduces the
proliferation, migration and invasion of A549 cells
To determine the potential mechanisms via which
B7-H3 could promote metastasis in NSCLC, A549 cells were
genome-edited to generate a B7-H3 KO using
CRISPR/Cas9 technology. Subsequently, a stable B7-H3
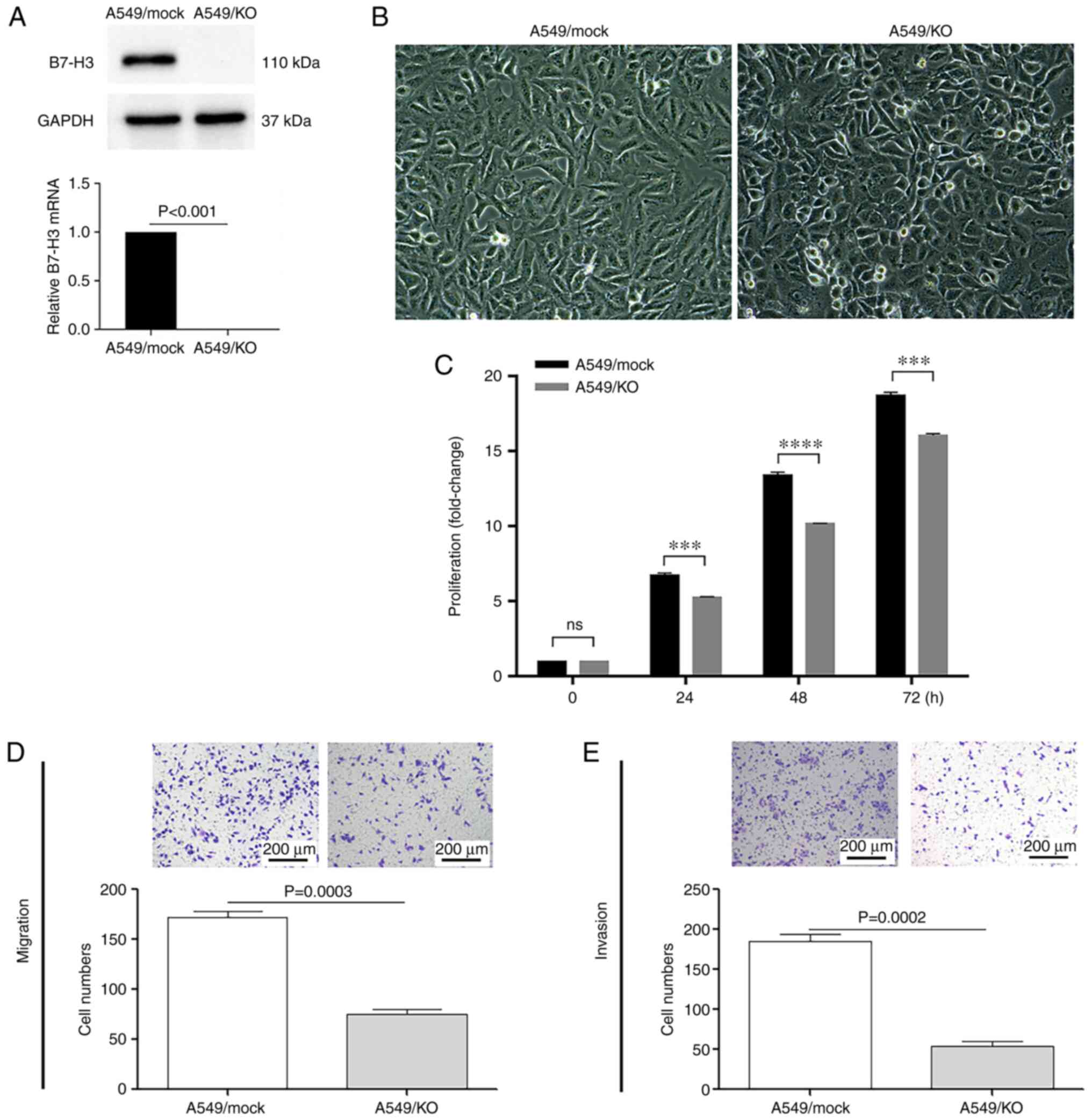
KO A549 strain was established, and confirmed via western blotting
and RT-qPCR (P<0.001; Fig.
1A). Morphologically, the B7-H3-deleted cells
were characterized by a bright outline, with a round blunt shape
(Fig. 1B). Furthermore, the KO of
B7-H3 expression led to significantly decreased proliferation of
A549 cells at 24, 48 and 72 h after plating (P<0.001; Fig. 1C).
Next, a Boyden chamber assay was then used to
evaluate the in vitro migratory and invasive capabilities of
the B7-H3 KO A549 cells. As shown in Fig. 1D and E, B7-H3
ablation reduced the percentages of migrating and invasive cells by
>50% (P=0.0003) and 70% (P=0.0002), respectively, compared with
the mock control. These results suggested that B7-H3 is associated
with the fast-growing and aggressive status of NSCLC by promoting
the proliferation and metastasis of cancer cells.
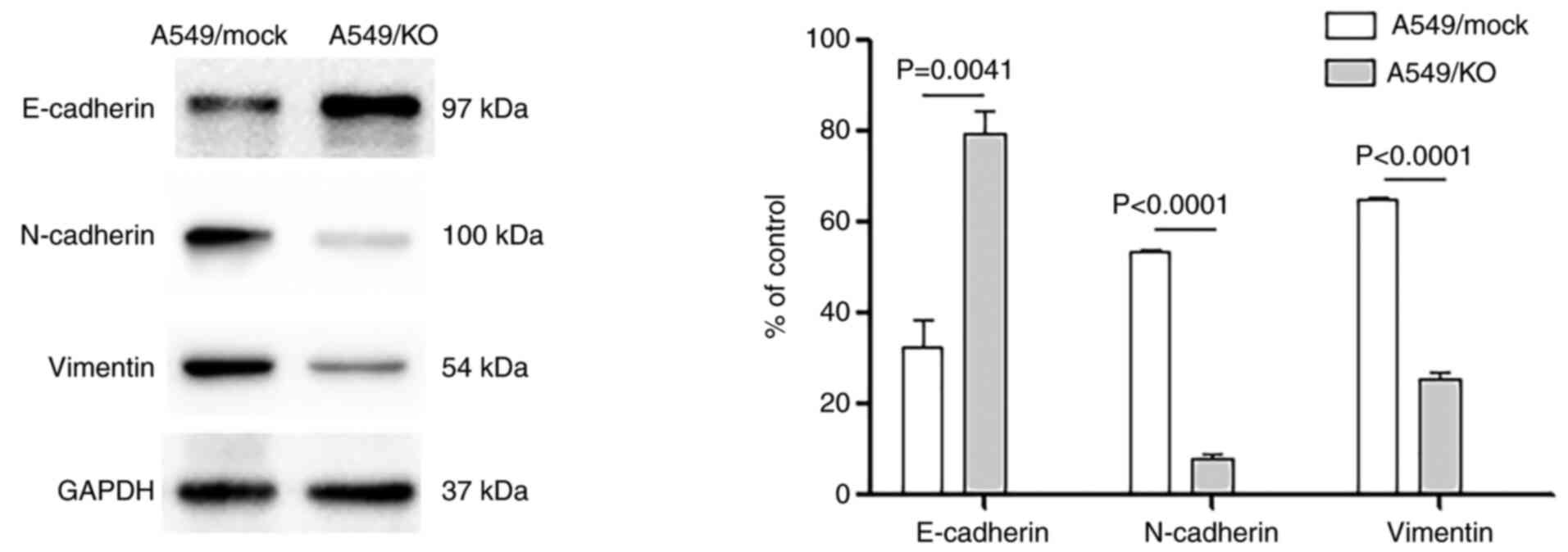
B7-H3 deletion regulates the
expression of EMT-associated factors
EMT is highly dysregulated in malignancy, leading to
functional changes in tumor cell migration and invasion.
Consequently, the effects of B7-H3 KO on aspects
typically associated with EMT were studied, including the
epithelial marker E-cadherin and the mesenchymal phenotypic
markers, vimentin and N-cadherin. B7-H3 deletion was
found to elicit a significant suppression of N-cadherin and
vimentin expression (both P<0.0001), whereas the expression of
E-cadherin was significantly increased compared with the mock
control A549 cells (P=0.0041) (Fig.
2). These results further demonstrated that B7-H3
silencing decreases the EMT process, hindering the loss of adhesive
junctions and the gain of mesenchymal activities.
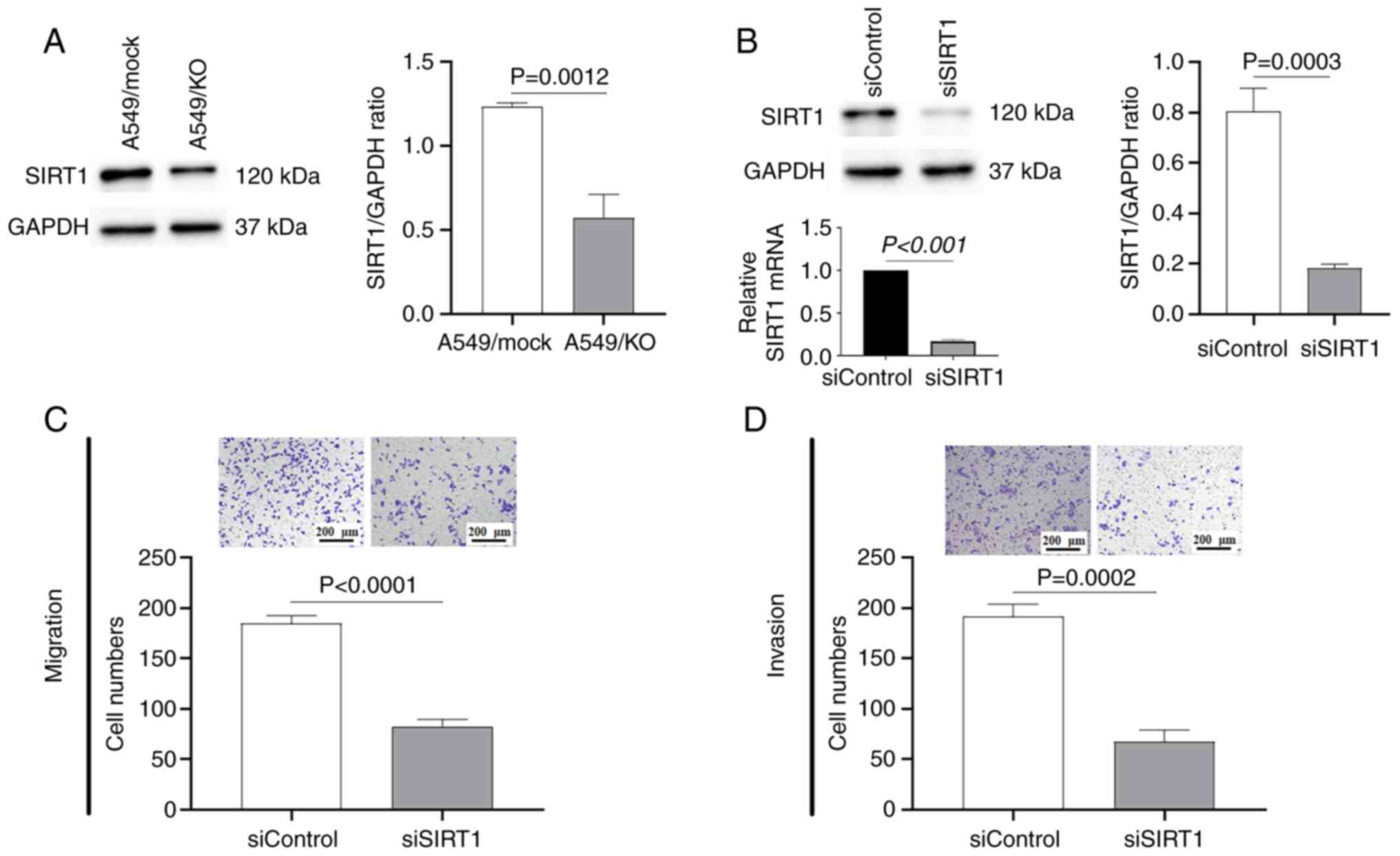
SIRT1 is involved in the B7-H3-induced
EMT process
SIRT1 is a NAD+-dependent, class III
histone deacetylase involved in the epigenetic regulation of tissue
homeostasis and numerous diseases, including tumorigenesis
(7). SIRT1 dysregulation has
already been demonstrated in various cancer cells (8,9);
however, its precise role in cancer development has yet to be fully
elucidated. In the present study, it was first demonstrated that
B7-H3 ablation led to a significant downregulation of
SIRT1 expression compared with the mock control (P=0.0012; Fig. 3A). Subsequently, the effects of
SIRT1 on biological processes relevant to metastatic activity,
migration and invasion were studied in A549 cells transfected with
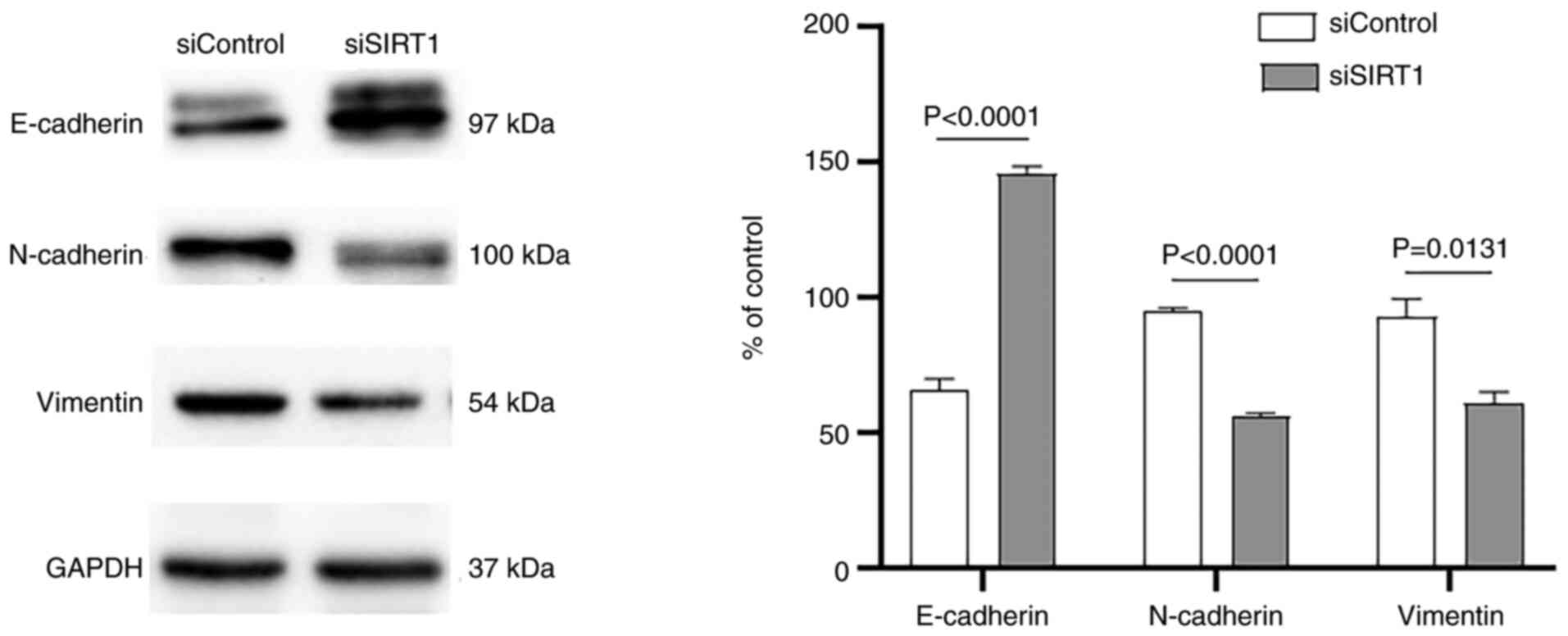
either siSIRT1 or the scrambled siControl (Fig. 3B). As shown in Fig. 3C and D, SIRT1 silencing
produced effects that were similar to those of B7-H3
deletion in A549 cells, reducing the numbers of migrating and
invasive cells by >50% (P<0.0001) and 60% (P=0.0002),
respectively, compared with the mock siControl. Consistent with
these findings, SIRT1 silencing also led to a significant decrease
in the expression levels of vimentin (P=0.0131) and N-cadherin
(P<0.0001), whereas E-cadherin was observed to be upregulated
(P<0.0001) in comparison with the scrambled control (Fig. 4). Therefore, these results
suggested that SIRT1 was involved in the B7-H3-induced EMT process
in A549 cells.
B7-H3 regulates SIRT1 via the PI3K/AKT
pathway
The PI3K/AKT, JAK2/STAT3 and Raf/MEK/ERK1/2
signaling cascades have been reported to be involved in
B7-H3-induced signaling in different types of cancer cells
(17–21). In view of this, the underlying
mechanisms governing how B7-H3-induced signaling may regulate SIRT1
expression was investigated in the present study. First, the
putative functional signaling pathway in A549 cells was assessed.
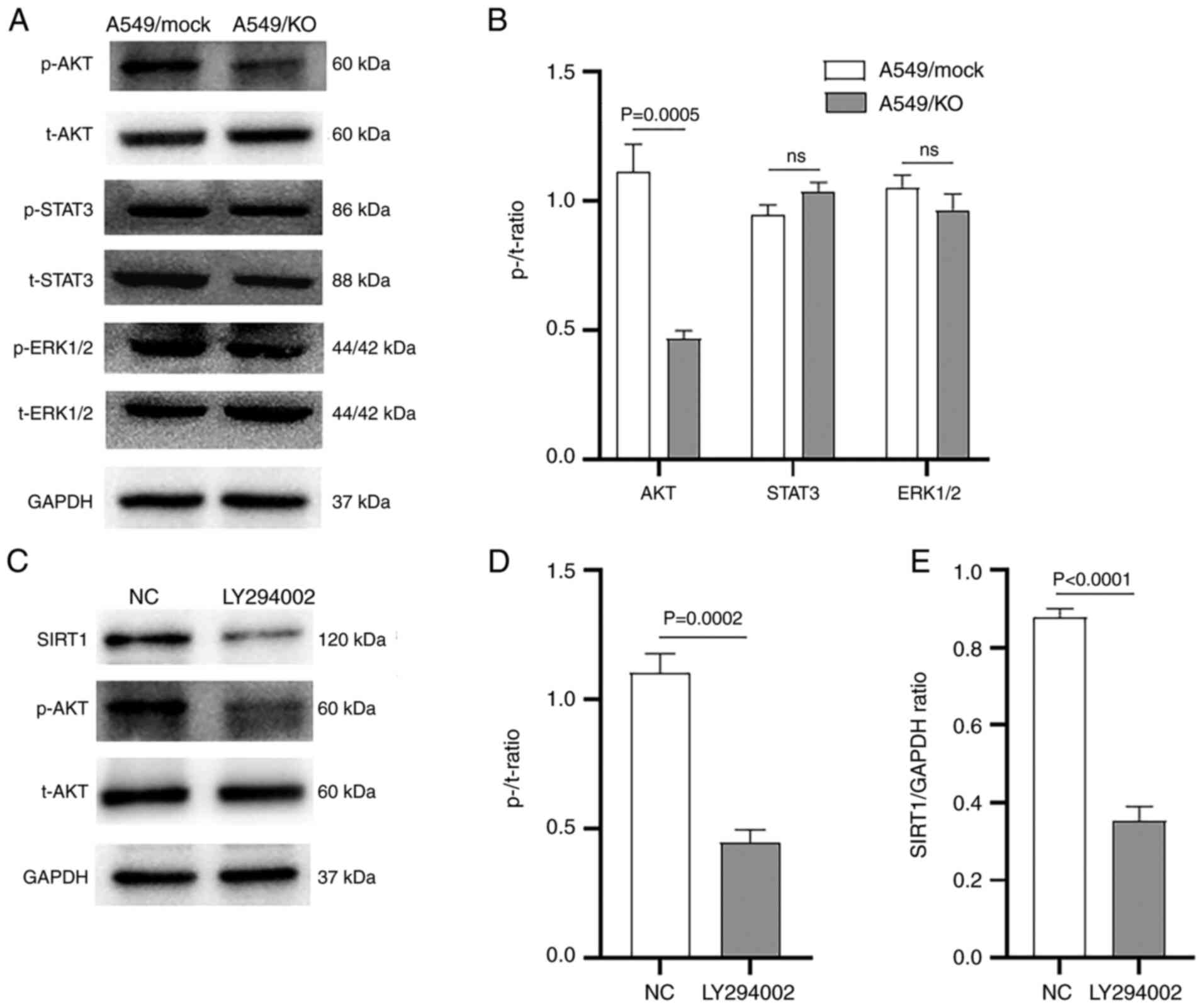
Western blot analysis revealed that the expression level of p-AKT,
but not of p-STAT3 or p-ERK1/2, was significantly downregulated in
B7-H3 KO A549 cells compared with the mock control
(P=0.0005) (Fig. 5A and B).
Secondly, to explore whether the PI3K/AKT pathway was involved in
B7-H3 mediated SIRT1 regulation, the specific PI3K inhibitor
LY294002 was used to observe its effects on SIRT1. Western blot
analysis showed that LY294002 resulted in significantly decreased
expression levels of SIRT1 (P<0.0001) and p-AKT (P=0.0002) in
A549 cells compared with those measured upon treatment with DMSO
(Fig. 5C-E). Taken together,
these results suggested that B7-H3 regulates SIRT1 via the PI3K/AKT
pathway in A549 cells.
Discussion
Data from the SEER Cancer Statistics Review
(25) showed that only ~16% of
all NSCLC cases diagnosed between the years 2009 and 2015 were of
localized disease, whereas 22% were diagnosed with regional
involvement and 57% of the cases exhibited dissemination to distant
metastasis. By contrast, the 5-year relative survival rates of
localized, regional and distant NSCLC were 57.4, 30.8 and 5.2%,
respectively (25). Therefore,
developing an understanding of metastases is critical, since
unmanageable migration and invasiveness are responsible for ~90% of
cases of death attributable to NSCLC. In the present study, in
vitro evidence has been provided to support that B7-H3 may be
closely associated with the aggressive status of NSCLC, since
B7-H3 ablation reduced the numbers of migrating and
invading cells by >50 and 70%, respectively. This result was
consistent with the findings of Yu et al (16), who had previously shown in in
vitro experiments the pro-metastasis function of B7-H3 in A549
cells through the siRNA silencing method. The significant effects
of B7-H3 on metastases have likewise been demonstrated in cervical
cancer (26), bladder cancer
(27) and gastric adenocarcinoma
cells (28). In vitro
B7-H3 knockdown via RNA interference in gastric adenocarcinoma
cells led to decreased rates of cell migration and Transwell
invasion by ≤50% (28).
Collectively, these results highlighted an important role for B7-H3
in promoting cancer cell metastasis, with life-threatening
consequences.
The invasiveness and metastatic activity acquired
during tumor progression have been shown to be due to EMT. After
activation of EMT, tumor cells either lose, or have a reduced
expression level of, epithelial-specific E-cadherin, whereas the
expression levels of N-cadherin and vimentin are increased as the
cells gain mesenchymal activities (6). The results from the present study
showed that B7-H3 deletion led to a substantial
suppression of vimentin and N-cadherin expression. By contrast,
B7-H3 KO significantly increased the expression of
E-cadherin compared with the mock control A549 cells. This result
was consistent with observations from a previous study (16), further demonstrating that
B7-H3 deletion decreases the EMT process by hindering
the loss of adhesive junctions and the gain of mesenchymal
activities. The promotion of tumor aggressiveness and invasion
resulting from B7-H3 targeting the EMT has also been reported in
glioma (18), hepatocellular
carcinoma (19), cervical cancer
(29) and salivary gland adenoid
cystic carcinoma (30). As a
result of the cells acquiring infiltrating and metastasizing
properties, high expression levels of B7-H3 have been shown to be
closely associated with poor prognosis of NSCLC (13). Therefore, as with programmed
death-ligand 1 (31), B7-H3 is an
attractive target for cancer immunotherapy of NSCLC. However, the
current strategies available are limited due to the utilization of
B7-H3 as an antibody-based tumor antigen (32). Alternative strategies based on
non-immunological pro-tumorigenic functions of B7-H3, such as
migration and invasion, are in need of further development, since
B7-H3-induced signaling in cancer cells remains poorly understood.
Thus, the in vivo experimentation to evaluate migration and
invasion in animal models, e.g., through magnetic resonance imaging
tracking is the focus of future research (33).
In the present study, it was shown that
B7-H3 ablation downregulated SIRT1 expression in A549
cells. Furthermore, SIRT1 silencing produced similar effects to
those of B7-H3 deletion, reducing the numbers of
migrating and invasive A549 cells by >50 and 60%, respectively.
SIRT1 silencing also caused a substantial decrease in the
expression levels of vimentin and N-cadherin, whereas E-cadherin
was observed to be upregulated in comparison with the scrambled
siControl. Therefore, these results suggested that SIRT1 may be
involved in the B7-H3-induced EMT process. A recent study has
highlighted that SIRT1 is a key regulator of cancer metastasis by
promoting EMT, since the dysregulation of SIRT1 substantially
altered the in vitro migration ability of cancer cells, and
in vivo tumor metastasis (9). In NSCLC, experiments using both
SIRT1-targeted miR-138 and its ‘sponge’, the circular RNA
hsa_circ_0006571, mutually confirmed the potential role of SIRT1 in
promoting EMT (34,35). However, a study by Li et al
(36) demonstrated that SIRT1
could serve a role as a tumor suppressor in NSCLC through
attenuating osteopontin-induced NF-κB p65 acetylation and the EMT.
Given that osteopontin is an extracellular matrix protein secreted
by both tumor cells and non-tumor stromal cells, it is likely that
the observable effects of SIRT1 on EMT are associated with the
tumor microenvironment. More recently, in studying colorectal
carcinoma, Meng et al (37) presented results that demonstrated
alterations in the SIRT1/NF-κB/B7-H3/TNF-α signaling axis,
suggesting that the protein level of B7-H3 is downregulated via
SIRT1-mediated NF-κB deactivation. However, whether a negative
feedback cycle exists between B7-H3 and SIRT1 requires further
investigation. More recently, Yu et al (38) demonstrated that there was a
fibroblast growth factor 21-induced SIRT1/PI3K/AKT signaling
pathway in A549 cells that could promote cell growth and migration.
Thus, positive feedback may also occur to increase the output by
mutual promotion between PI3K/AKT and SIRT1.
The results of the present study demonstrated that
the PI3K/AKT signaling pathway, but not the JAK2/STAT3 and
Raf/MEK/ERK1/2 pathways, was functionally affected by B7-H3 in A549
cells. Furthermore, treatment with the specific PI3K inhibitor
LY294002 resulted in significantly decreased SIRT1 and p-AKT
expression levels, which suggested that B7-H3 regulates SIRT1 via
the PI3K/AKT pathway. The finding that B7-H3 promotes cell
migration and invasion (39),
drug resistance (20,40) and aerobic glycolysis (21) via the PI3K/AKT pathway has also
been reported in ovarian cancer (20), oral squamous carcinoma (21), bladder cancer (39) and colorectal cancer cells
(40). On the other hand, B7-H3
has been shown to promote growth and invasion of multiple myeloma
(17), glioma (18) and hepatocellular carcinoma
(19) by activating the
JAK2/STAT3 signaling pathway, and to promote the angiogenesis of
colorectal cancer through activating the NF-κB pathway (41). Therefore, it is likely that
B7-H3-induced signaling is transduced through diverse signaling
cascades for the purpose of fulfilling several different functions.
Tumoral B7-H3 has multiple effects in tumors, by promoting cancer
invasion, migration, angiogenesis, drug sensitivity and the Warburg
effect. The other possibility is that the signaling cascades
downstream of B7-H3 operate differently according to the different
cancer types and subtypes. It should be noted that our previous
study demonstrated that B7-H3-induced signaling is dissimilar,
comparing among lung adenocarcinoma cell lines with divergent
epidermal growth factor receptor mutation patterns (23).
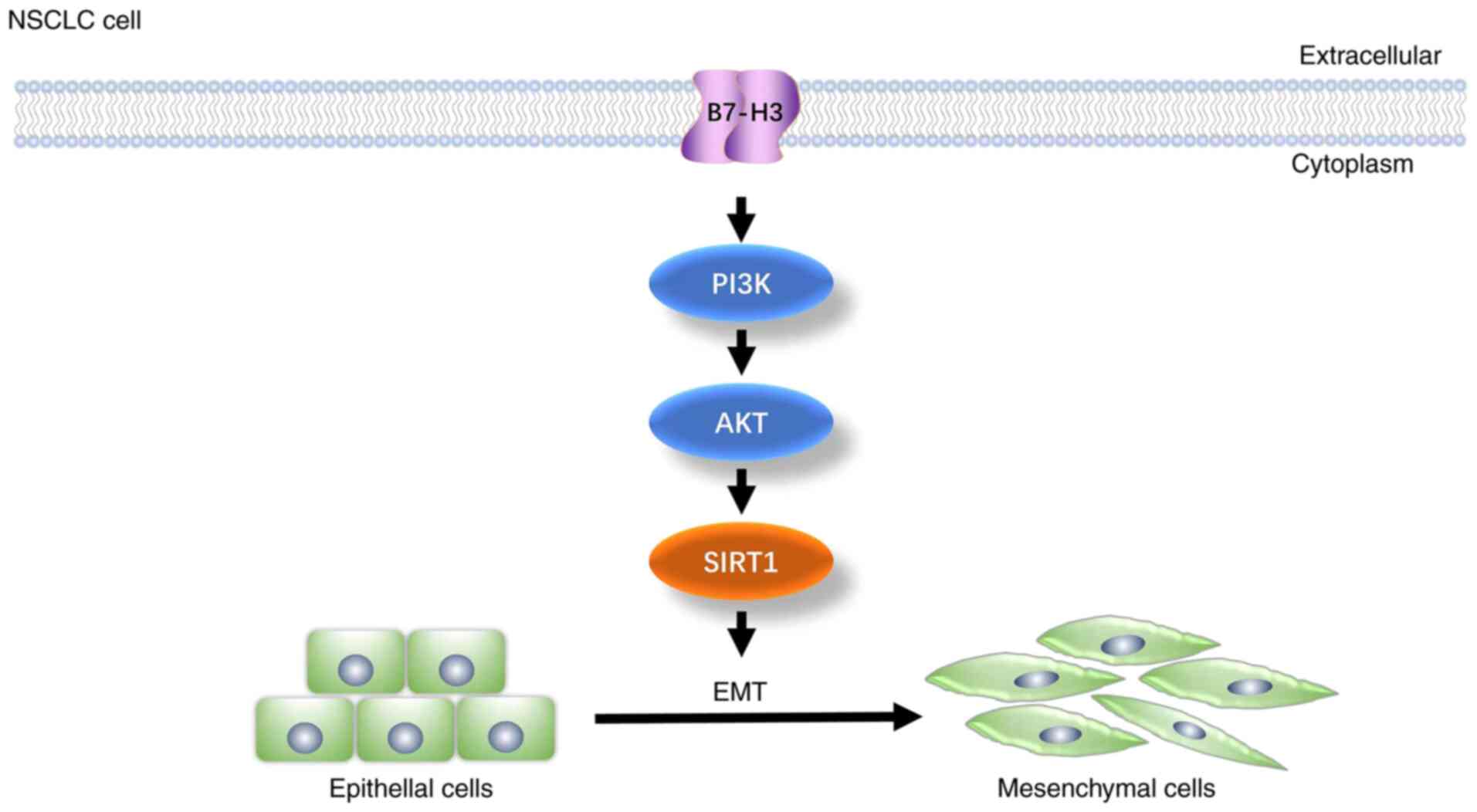
Taken together, the present results have confirmed
that B7-H3 contributes to the migration and invasion of NSCLC cells
by promoting the EMT process. The present study has provided
evidence that the class III histone deacetylase SIRT1 may be
involved in B7-H3-induced EMT, and also that B7-H3 regulates SIRT1
via the PI3K/AKT signaling pathway in NSCLC A549 cells (Fig. 6). These findings may prove to be
useful in terms of identifying novel strategies for therapeutic
intervention in NSCLC metastasis. Further studies are required,
however, to provide in vivo evidence of the effectiveness
and therapeutic feasibility of targeting B7-H3-mediated EMT in
NSCLC.
Acknowledgements
Not applicable.
Funding
This work was funded by the Natural Science Foundation of Anhui
Province (grant no. 1808085MH229) and Key Research and Development
Program of Anhui Province (grant no. 202004j07020027).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LC and HL designed the present study and analyzed
the data. HL and MD performed the experiments and drafted the
article. NZ and YY contributed to the analysis and interpretation
of all data. LC revised the manuscript and performed the manuscript
submission. HL and MD confirm the authenticity of all the raw data.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
EMT
|
epithelial-mesenchymal transition
|
|
NSCLC
|
non-small cell lung cancer
|
|
SCLC
|
small cell lung cancer
|
|
SIRT1
|
Sirtuin 1
|
|
AKT/PKB
|
Protein kinase B
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
KO
|
knockout
|
|
mock
|
negative control
|
|
siControl
|
negative control siRNAs oligos
|
|
DMSO
|
dimethyl sulfoxide
|
|
t-
|
total
|
|
p-
|
phosphorylated
|
|
ECL
|
enhanced chemiluminescent
|
|
HRP
|
horseradish peroxidase
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Travis WD, Brambilla E, Burke AP, Marx A
and Nicholson AG: Introduction to the 2015 World Health
Organization Classification of tumors of the lung, pleura, thymus,
and heart. J Thorac Oncol. 10:1240–1242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang Z, Su W, Lu T, Wang Y, Dong Y, Qin
Y, Liu D, Sun L and Jiao W: First-line immune-checkpoint inhibitors
in non-small cell lung cancer: Current landscape and future
progress. Front Pharmacol. 11:5780912020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mittal V: Epithelial mesenchymal
transition in tumor metastasis. Annu Rev Pathol. 13:395–412. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang W, Yan Y, Liu Y, Lin M, Ma J, Zhang
W, Dai J, Li J, Guo Q, Chen H, et al: Exosomes with low miR-34c-3p
expression promote invasion and migration of non-small cell lung
cancer by upregulating integrin α2β1. Signal Transduct Target Ther.
5:392020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jørgensen CLT, Forsare C, Bendahl PO,
Falck AK, Fernö M, Lövgren K, Aaltonen K and Rydén L: Expression of
epithelial-mesenchymal transition-related markers and phenotypes
during breast cancer progression. Breast Cancer Res Treat.
181:369–381. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sauve AA, Wolberger C, Schramm VL and
Boeke JD: The biochemistry of sirtuins. Annu Rev Biochem.
75:435–465. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao B, Li X, Zhou L, Wang Y and Shang P:
SIRT1: A potential tumour biomarker and therapeutic target. J Drug
Target. 27:1046–1052. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choupani J, Mansoori DS, Bayat S, Alivand
MR and Shekari KM: Narrower insight to SIRT1 role in cancer: A
potential therapeutic target to control epithelial-mesenchymal
transition in cancer cells. J Cell Physol. 233:4443–457. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nehama D, Di Ianni N, Musio S, Du H,
Patane M, Pollo B, Finocchiaro G, Park JJ, Dunn DE, Edwards DS, et
al: B7-H3-redirected chimeric antigen receptor T cells target
glioblastoma and neurospheres. EBioMedicine. 47:33–43. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Z, Jiang C, Liu Z, Yang M, Tang X,
Wang Y, Zheng M, Huang J, Zhong K, Zhao S, et al: B7-H3-targeted
CAR-T cells exhibit potent antitumor effects on hematologic and
solid tumors. Mol Ther Oncolytics. 17:180–189. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Seaman S, Zhu Z, Saha S, Zhang XM, Yang
MY, Hilton MB, Morris K, Szot C, Morris H, Swing DA, et al:
Eradication of tumors through simultaneous ablation of
CD276/B7-H3-positive tumor cells and tumor vasculature. Cancer
Cell. 31:501–515. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Altan M, Pelekanou V, Schalper KA, Toki M,
Gaule P, Syrigos K, Herbst RS and Rimm DL: B7-H3 expression in
NSCLC and its association with B7-H4, PD-L1 and tumor-infiltrating
lymphocytes. Clin Cancer Res. 23:5202–5209. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yonesaka K, Haratani K, Takamura S, Sakai
H, Kato R, Takegawa N, Takahama T, Tanaka K, Hayashi H, Takeda M,
et al: B7-H3 negatively modulates CTL-mediated cancer immunity.
Clin Cancer Res. 24:2653–2664. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cai D, Li J, Liu D, Hong S, Qiao Q, Sun Q,
Li P, Lyu N, Sun T, Xie S, et al: Tumor-expressed B7-H3 mediates
the inhibition of antitumor T-cell functions in ovarian cancer
insensitive to PD-1 blockade therapy. Cell Mol Immunol. 17:227–236.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu TT, Zhang T, Lu X and Wang RZ: B7-H3
promotes metastasis, proliferation, and epithelial-mesenchymal
transition in lung adenocarcinoma. Onco Targets Ther. 11:4693–4700.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin L, Cao L, Liu Y, Wang K, Zhang X, Qin
X, Zhao D, Hao J, Chang Y, Huang X, et al: B7-H3 promotes multiple
myeloma cell survival and proliferation by ROS-dependent activation
of Src/STAT3 and c-Cbl-mediated degradation of SOCS3. Leukemia.
33:1475–1486. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhong C, Tao B, Chen Y, Guo Z, Yang X,
Peng L, Xia X and Chen L: B7-H3 Regulates Glioma growth and cell
invasion through a JAK2/STAT3/Slug-dependent signaling pathway.
Onco Targets Ther. 13:2215–2224. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kang FB, Wang L, Jia HC, Li D, Li HJ,
Zhang YG and Sun DX: B7-H3 promotes aggression and invasion of
hepatocellular carcinoma by targeting epithelial-to-mesenchymal
transition via JAK2/STAT3/Slug signaling pathway. Cancer Cell Int.
15:452015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou L and Zhao Y: B7-H3 induces ovarian
cancer drugs resistance through an PI3K/AKT/BCL-2 signaling
pathway. Cancer Manag Res. 11:10205–10214. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Z, Liu J, Que L and Tang X: The
immunoregulatory protein B7-H3 promotes aerobic glycolysis in oral
squamous carcinoma via PI3K/Akt/mTOR pathway. J Cancer.
10:5770–5784. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Flem-Karlsen K, Fodstad Ø and Nunes-Xavier
CE: B7-H3 immune checkpoint protein in human cancer. Curr Med Chem.
27:4062–4086. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ding M, Liao H, Zhou N, Yang Y, Guan S and
Chen L: B7-H3-induced signaling in lung adenocarcinoma cell lines
with divergent epidermal growth factor receptor mutation patterns.
Biomed Res Int. 2020:88248052020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Howlader N, Noone AM, Krapcho M, Miller D,
Brest A, Yu M, Ruhl J, Tatalovich Z, Mariotto A, Lewis DR, et al:
SEER Cancer Statistics Review, 1975-2016. National Cancer
Institute; https://seer.cancer.gov/csr/1975_2016/Bethesda, MD:
2019
|
|
26
|
Li Y, Zhang J, Han S, Qian Q, Chen Q, Liu
L and Zhang Y: B7-H3 promotes the proliferation, migration and
invasiveness of cervical cancer cells and is an indicator of poor
prognosis. Oncol Rep. 38:1043–1050. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu ZL, Zhang Y, Wang L, Li F, Man HW, Li
PF and Shan BE: B7-H3 promotes malignant progression of
muscle-invasive bladder cancer. Oncol Rep. 40:2722–2733.
2018.PubMed/NCBI
|
|
28
|
Dai W, Shen G, Qiu J, Zhao X and Gao Q:
Aberrant expression of B7-H3 in gastric adenocarcinoma promotes
cancer cell metastasis. Oncol Rep. 32:2086–2092. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang X, Feng KX, Li H, Wang L and Xia H:
MicroRNA-199a inhibits cell proliferation, migration, and invasion
and activates AKT/mTOR signaling pathway by targeting B7-H3 in
cervical cancer. Technol Cancer Res Treat. Aug 28–2020.(Epub ahead
of print). doi: 10.1177/1533033820942245. View Article : Google Scholar
|
|
30
|
Fan TF, Deng WW, Bu LL, Wu TF, Zhang WF
and Sun ZJ: B7-H3 regulates migration and invasion in salivary
gland adenoid cystic carcinoma via the JAK2/STAT3 signaling
pathway. Am J Transl Res. 9:1369–1380. 2017.PubMed/NCBI
|
|
31
|
Luo M, Wang F, Zhang H, To KKW, Wu S, Chen
Z, Liang S and Fu L: Mitomycin C enhanced the efficacy of PD-L1
blockade in non-small cell lung cancer. Signal Transduct Target
Ther. 5:1412020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kontos F, Michelakos T, Kurokawa T,
Sadagopan A, Schwab JH, Ferrone CR and Ferrone S: B7-H3: An
attractive target for antibody-based immunotherapy. Clin Cancer
Res. 27:1227–1235. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bretschi M, Merz M, Komljenovic D, Berger
MR, Semmler W and Bäuerle T: Cilengitide inhibits metastatic bone
colonization in a nude rat model. Oncol Rep. 26:843–851.
2011.PubMed/NCBI
|
|
34
|
Ye Z, Fang B, Pan J, Zhang N, Huang J, Xie
C, Lou T and Cao Z: miR-138 suppresses the proliferation,
metastasis and autophagy of non-small cell lung cancer by targeting
Sirt1. Oncol Rep. 37:3244–3252. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang HL, Wang HR, Liang Y, Hu AN, Enguita
FJ, Zhou XG and Dong J: Hsa_circ_0006571 promotes spinal metastasis
through sponging microRNA-138 to regulate sirtuin 1 expression in
lung adenocarcinoma. Transl Lung Cancer Res. 9:2411–2427. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li X, Jiang Z, Li X and Zhang X: SIRT1
overexpression protects non-small cell lung cancer cells against
osteopontin-induced epithelial-mesenchymal transition by
suppressing NF-κB signaling. Onco Targets Ther. 11:1157–1171. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Meng F, Yang M, Chen Y, Chen W and Wang W:
miR-34a induces immunosuppression in colorectal carcinoma through
modulating a SIRT1/NF-κB/B7-H3/TNF-α axis. Cancer Immunol
Immunother. 70:2247–2259. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yu X, Li Y, Jiang G, Fang J, You Z, Shao
G, Zhang Z, Jiao A and Peng X: FGF21 promotes non-small cell lung
cancer progression by SIRT1/PI3K/AKT signaling. Life Sci.
269:1188752021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li Y, Guo G, Song J, Cai Z, Yang J, Chen
Z, Wang Y, Huang Y and Gao Q: B7-H3 promotes the migration and
invasion of human bladder cancer cells via the PI3K/Akt/STAT3
signaling pathway. J Cancer. 8:816–824. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang P, Chen Z, Ning K, Jin J and Han X:
Inhibition of B7-H3 reverses oxaliplatin resistance in human
colorectal cancer cells. Biochem Biophys Res Commun. 490:1132–1138.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang R, Ma Y, Zhan S, Zhang G, Cao L,
Zhang X, Shi T and Chen W: B7-H3 promotes colorectal cancer
angiogenesis through activating the NF-κB pathway to induce VEGFA
expression. Cell Death Dis. 11:552020. View Article : Google Scholar : PubMed/NCBI
|