Introduction
Osteoporosis (OP) is a systemic skeletal disease
characterized by decreased bone mass and microarchitectural
deterioration in the bone tissue, which predisposes affected
individuals to increased bone fragility and fracture risk (1). The diagnosis of OP is based on bone
mineral density (BMD) measurement using dual-energy X-ray
absorptiometry (DXA). According to the World Health Organization, a
BMD T-score of <-2.5, as measured by DXA, is defined as OP
(2). Approximately 50% of women
experience at least one bone fracture following menopause worldwide
in 2020; OP has become a significant public health problem and
received considerable attention worldwide (3).
Type-2 diabetes mellitus (T2DM) has been
hypothesized to cause decreased BMD (4). A large prospective study of
post-menopausal women recruited into the Study of Osteoporotic
Fractures confirmed that female patients with T2DM experienced
higher fracture rates compared with that in female patients without
T2DM (5). In addition, BMD
decreased as diabetes progressed, suggesting that hyperglycemia is
an important risk factor for OP (6).
Advanced glycation end products (AGEs) are a group
of modified proteins and/or lipids with damaging potential, that
are formed under hyperglycemic conditions (7). AGEs have been largely reported to be
involved in the pathogenesis of T2DM and diabetic complications
(7). In addition, increasing
evidence indicates that AGEs play a role in aggravating bone
fragility, as observed during the progression of classic diabetic
complications (8,9). AGEs have been found to significantly
inhibit osteoblast proliferation, differentiation and
mineralization and induce osteoblast apoptosis (10,11). AGEs can affect osteoblast
proliferation and function by modulating autophagy via the receptor
of AGEs (RAGE) pathway, indicating that autophagy is involved in
these processes (12). These
observations suggest that the AGE/RAGE axis may play an important
role in bone formation.
A recent study have revealed that another form of
regulated cell death, ferroptosis, may be involved in the
pathogenesis of OP (13).
Ferroptosis (iron-dependent cell death) differs from apoptosis,
autophagy, necrosis, necroptosis and pyroptosis, and is
characterized by the accumulation of intracellular iron and lipid
reactive oxygen species (14).
The increase in osteoblast ferroptosis plays an important role in
hormone-induced OP (15), and
iron overload plays an important role in general OP and osteoblast
function (16). In a rat model of
T2DM-related OP (OP-T2DM), ferroptosis was observed in the bone
tissue (17). However, none of
the aforementioned studies directly studied the level and changes
of ferroptosis in osteoblasts.
Therefore, the aim of the present study was to
investigate the role of ferroptosis in AGE-regulated dysfunction of
human osteoblasts. Ex vivo experiments using clinical
samples from patients with OP and healthy individuals were
performed. In addition, in vitro experiments using the
hFOB1.19 osteoblast cell line, which was cultured in media
containing AGEs and serum from healthy individuals or patients with
OP and a normal fasting blood glucose (FBG) level and patients with
OP and T2DM to observe cell differentiation, mineralization,
apoptosis and ferroptosis, were also performed.
Materials and methods
Sample collection
The present study was approved by the Ethics
Committee of the Nanjing Pukou District Central Hospital (Nanjing,
China). Written informed consent was obtained from all
participants. A total of 10 healthy individuals and 10 patients
with OP (among which 4 were diagnosed with T2DM) were enrolled into
the present study. Participants with a femoral neck and/or lumbar
spine T score ≤-2.5 were considered to have OP, while a T score of
>-1 was considered normal. Patients with malignancy, coronary
heart disease or other severe diseases in the previous 5 years were
excluded. Images of lumbar vertebra and femur, along with the BMD
from healthy individuals and patients with OP, were obtained using
DXA absorptiometry (Hologic, Inc.). Blood samples from the enrolled
participants were centrifuged at 1,000 × g for 10 min at 4°C, and
the serum was stored at −80°C.
Measurement of AGEs and FBG
All the participants fasted from 10:00 pm the
previous night, and 10 ml venous blood was collected at ~9:00 am
the next morning. The blood samples were centrifuged at 1,000 × g
for 10 min at 4°C, and the serum was used to assess AGEs and FBG or
stored at −80°C for subsequent experiments. The concentration of
AGEs and FBG in the serum samples was measured using an AGE assay
kit (cat no ab238539; Abcam) and a blood glucose kit (Glucose
Oxidase Peroxidase Assay; cat no WD-0108 Shanghai Rongbai
Biotechnology Co., Ltd.), following the manufacturer's
instructions.
Preparation of AGEs
AGEs were prepared as previously described (18). Briefly, human serum albumin (15
mg/ml; Sigma-Aldrich; Merck KGaA) was incubated with 1 mM
3-deoxygluconaldeone in distilled water at 37°C for 14 days.
Cell culture and grouping
The human hFOB1.19 osteoblast cell line (American
Type Culture Collection; ATCC) was cultured in medium containing
1:1 mixture of Ham's F12 medium and DMEM (both from Thermo Fisher
Scientific, Inc.) with 2.5 mM L-glutamine (ATCC), supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin, 100 U/ml streptomycin (Merck KGaA) and 0.3 mg/ml G418
(Merck KGaA). The cells were cultured at 37°C in a humidified
incubator with 5% CO2.
The hFOB1.19 cells were divided into different
groups: Blank (cultured in normal medium), AGEs (cultured in medium
containing 50 pg/ml AGEs), control (cultured in medium containing
10% serum from healthy individuals), OP-normal (cultured in medium
containing 10% serum from patients with OP and a normal FBG level),
OP-T2DM (cultured in medium containing 10% serum from patients with
OP and T2DM), AGEs + deferoxamine (DFO; cultured in medium
containing 50 pg/ml AGE and 80 µM DFO), OP-T2DM + DFO (cultured in
medium containing 10% serum from patients with OP and T2DM, and 80
µM DFO).
Cell counting kit (CCK)-8 assay
The hFOB1.19 cell line, in the logarithmic growth
phase, were seeded into 96-well plates, at a density of
3×103 cells/well, with five replicate wells in each
group. Following culture for 24 h in normal medium, the medium was
replaced with new medium according to the aforementioned groups.
Following culture for 0, 24, 48 and 72 h, 10 µl CCK-8 solution
(Beyotime Institute of Biotechnology) was added to each well and
the samples were incubated at 37°C in the dark for 2 h. The
absorbance value of each well was measured at 450 nm using a
microplate reader (Bio-Rad Laboratories, Inc.).
Alkaline phosphatase (ALP)
The ALP level in the hFOB1.19 cell lysate from each
group was measured using a commercial ALP Assay kit (cat no
ab83369; Abcam), following the manufacturer's instructions.
Briefly, following culture in different media for 48 h, the cell
supernatant was removed and the production of p-nitrophenyl
phosphate was determined by measuring the absorbance at 405 nm
using a microplate reader (Bio-Rad Laboratories, Inc.).
Alizarin red staining
The bone mineralization of the hFOB1.19 cells in
each group was detected using Alizarin red staining (Merck KGaA),
following the manufacturer's instructions. Briefly, the cells were
stained with 2% Alizarin red (pH 4.2) for 10 min at room
temperature, then washed with distilled water. The mineralized
nodules were examined using phase-contrast microscopy after
culturing for 21 days.
TUNEL staining
Cell apoptosis was evaluated using a TUNEL assay
(TUNEL Assay kit-FITC; cat. no. ab66108; Abcam), according to the
manufacturer's instructions. Briefly, following culture in
different media for 48 h, the cells were trypsinized, washed with
PBS and fixed in 1% paraformaldehyde at 4°C for 2 h. The cells were
then incubated with DNA labeling solution for 60 min at 37°C,
followed by the addition of PI/RNase A solution for 30 min. The
images of the TUNEL-labeled cells from 3 random fields were
captured using a fluorescent microscope (magnification, ×200; Evos
FL Cell Imaging System; Thermo Fisher Scientific, Inc.).
Determination of iron level
The ferrous iron level in the cells was determined
using a commercially available indicator (cat. no. P14312; Thermo
Fisher Scientific, Inc.), as previously described (19). Briefly, following treatment, the
cells were resuspended in normal medium, then Phen Green™ SK
fluorescent iron chelator was added for 15 min. The fluorescence
represented the release of free iron and was observed at the
excitation/emission values of 490/520 nm using a fluorescence
microscope (magnification ×200; Evos FL Cell Imaging System; Thermo
Fisher Scientific, Inc.).
Malondialdehyde (MDA) and oxidized low
density lipoprotein (oxLDL) assessment
The relative MDA and oxLDL levels in the hFOB1.19
cells were detected using a Lipid Peroxidation (cat. no. ab118970)
and oxLDL (cat. no. ab242302) (both from Abcam) assay kits,
respectively, according to the manufacturer's instructions.
Glutathione (GSH)/oxidized GSH (GSSG)
assay
The relative GSH concentration in the cells was
assessed using a GSH/GSSG Ratio Detection Assay kit
(Fluorometric-Green; cat. no. ab138881; Abcam), according to the
manufacturer's instructions. Briefly, the whole cell lysates were
diluted at 1:80 for GSH analysis, and a series dilution of GSH and
GSSG stock standards were prepared as standards. A one-step
fluorimetric reaction was performed, with the assay buffer and
probes, for 30 min at room temperature in the dark. Fluorescence
intensity was monitored at the excitation/emission values of
490/520 nm. GSH was calculated from the standard curve. GSSG was
determined using the following equation: GSSG=(total
GSH-GSH)/2.
Western blot analysis
Following treatment, the cells were lysed in cold
RIPA buffer (Beyotime Institute of Biotechnology). The protein
lysates were then centrifuged at 12,000 × g for 10 min at 4°C and
the supernatant was collected. Protein concentration was determined
using a BCA assay kit (Beyotime Institute of Biotechnology). A
total of 10 µg total protein was separated using 10–15% SDS-PAGE,
then transferred onto PVDF membranes. The membranes were then
blocked for 1 h at room temperature with 5% skimmed milk and
subsequently incubated with the primary antibodies overnight at
4°C. Finally, the membranes were incubated with a HRP-conjugated
secondary antibody (goat anti-rabbit IgG; cat no. ab6721; Abcam;
1:10,000) for 2 h at room temperature. The bands were detected
using the ECL Advance reagent (Cytiva) and quantified using ImageJ
software (version 1.8.0; National Institutes of Health). The
following primary antibodies were used: GSH peroxidase (GPX4;
ab125066; Abcam; 1:5,000), solute carrier family 7 member 11
(SLC7A11; ab175186; Abcam; 1:5,000), nuclear receptor coactivator 4
(NCOA4; ab86707; Abcam; 1:3,000), acyl-CoA synthetase long-chain
family member 4 (ACSL4; ab15282; Abcam; 1:10,000), transferrin
receptor (TFR1), divalent metal transporter 1 (DMT1; ab214039;
Abcam; 1:1,000) and GAPDH (ab181602; Abcam; 1:1,000).
Statistical analysis
All the data are from 3 experiments and are
presented as the mean ± SD. Comparisons between two groups were
analyzed using an unpaired Student's t-test. Comparisons among
multiple groups were evaluated using one-way ANOVA followed by
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference. All statistical analysis was
performed using SPSS software (version 11.5; SPSS, Inc.).
Results
Level of serum AGEs and FBG is
increased in patients with OP
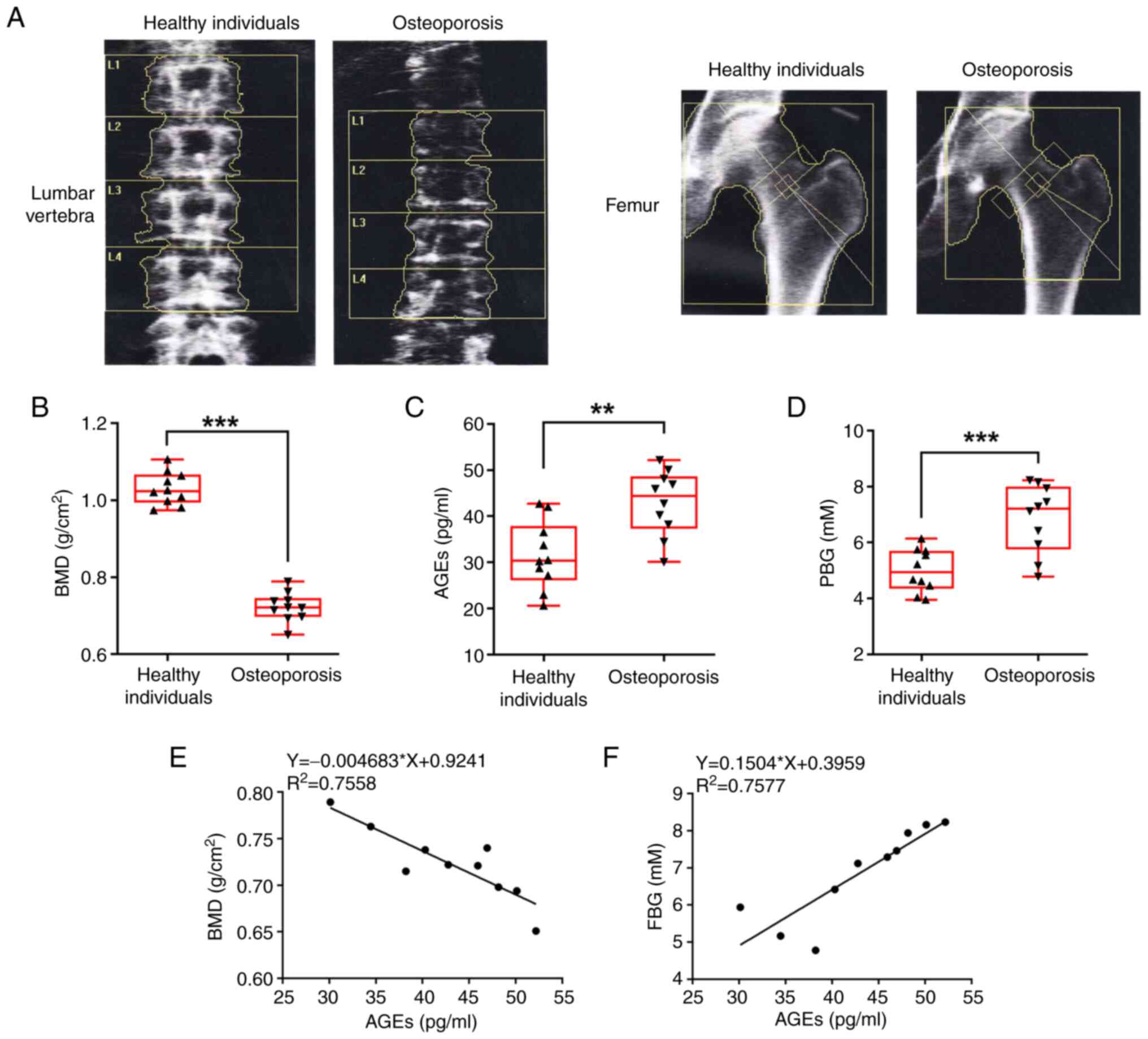
Fig. 1A shows the
images of lumbar vertebra and femur in healthy individuals and
patients with OP, which were collected during DXA absorptiometry.
Compared with that in the healthy individuals, patients with OP
exhibited a decreased BMD in both the lumbar vertebra and femur.
Fig. 1B reveals the significantly
lower BMD in patients with OP compared with that in the healthy
individuals. Next, the level of AGE and FBG in the serum of the
participants were measured. Patients with OP exhibited a
significantly higher level of AGEs (Fig. 1C) and FBG (Fig. 1D). The correlation between BMD and
AGEs, and between FBG and AGEs in patients with OP was analyzed and
the results showed that in patients with OP, the higher the AGE
level, the lower the BMD level, but the higher the FBG level
(Fig. 1E and F, respectively).
These results suggested that an increase in the concentration of
AGEs and FBG may be associated with the decrease in BMD, and even
contribute to OP.
AGEs and serum from patients with OP
inhibit the proliferation, differentiation and mineralization of
hFOB1.19 osteoblasts
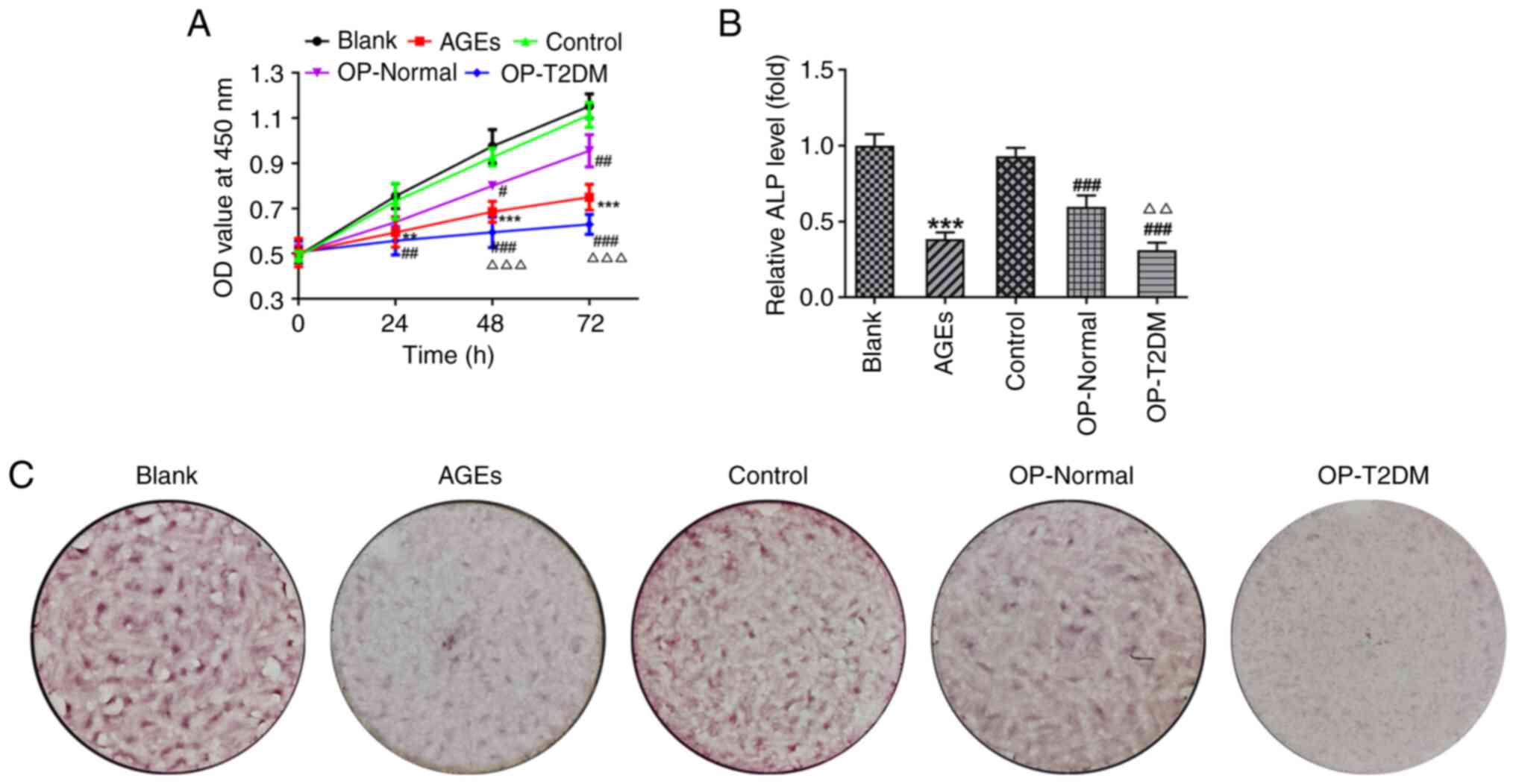
To validate the aforementioned hypothesis in
vitro, the hFOB1.19 osteoblast cell line was cultured in normal
medium (blank), medium containing AGEs (AGEs), or media containing
serum from healthy individuals (control), OP patients with a normal
FBG level (OP-normal) and OP patients with T2DM (OP-T2DM). Cell
proliferation, differentiation and mineralization were then
assessed. The cells that were treated with AGE, and serum from
patients with OP and T2DM exhibited significantly reduced cell
proliferation ability 24 h post-treatment, compared with that in
cells in the blank and control groups, respectively (Fig. 2A). Compared with that in serum
from the healthy individuals, serum from patients with OP and a
normal FBG level had significantly lower cell proliferation 48 h
post-treatment (P<0.05). In addition, the serum from patients
with OP and T2DM resulted in a relatively higher decrease in cell
proliferation compared with that in serum from patients with OP and
a normal FBG level (Fig. 2A). A
similar result was found for ALP levels in the hFOB1.19 cells
(Fig. 2B), in which ALP level was
decreased by AGEs and serum from patients with OP (OP-Normal and
OP-T2DM). Alizarin red staining was performed to determine the
mineralization of osteoblasts. Compared with that in cells in the
blank and control groups, cells in the AGE, OP-Normal and OP-T2DM
groups exhibited a decrease in the number of mineralized nodules
(Fig. 2C). These results
suggested that AGEs and serum from patients with OP could inhibit
the proliferation, differentiation and mineralization of hFOB1.19
osteoblasts. In addition, the effect of serum from patients with OP
and T2DM was stronger compared with that of serum from patients
with OP and a normal FBG level.
AGEs and serum from patients with OP
induce ferroptosis in hFOB1.19 osteoblasts
Subsequently, changes in cell ferroptosis were
investigated. TUNEL staining was used to observe cell apoptosis. As
shown in Fig. 3A, the AGEs group
had a higher percentage of apoptotic cells compared with that in
the blank group. The OP-Normal and OP-T2DM groups also exhibited
increased apoptosis compared with the control group. Next, cellular
free iron levels were analyzed (Fig.
3B) and the results showed that AGEs and serum from patients
with OP significantly increased the cellular free iron level of
hFOB1.19 osteoblasts. The concentrations of MDA and oxLDL was also
significantly increased by AGEs and serum from patients with OP
(OP-Normal and OP-T2DM), compared with those in the blank and
control groups, respectively (Fig. 3C
and D). However, the opposite effect was caused by AGEs and
serum from OP patients (OP-Normal and OP-T2DM) on GSH/GSSG level
(Fig. 3E). Following the
detection of the expression level of ferroptosis-associated
proteins, it was found that AGEs and serum from patients with OP
resulted in a decrease in the protein expression level of GPX4 and
SLC7A11, but an increase in the protein expression level of NCOA4,
ACSL4, TFR1 and DMT1 (Fig. 3F and
G). The results revealed that AGEs and serum from patients with
OP significantly increased ferroptosis in hFOB1.19 osteoblasts, and
that the effect of the serum from patients with OP and T2DM was
stronger compared with that from patients with OP and a normal FBG
level.
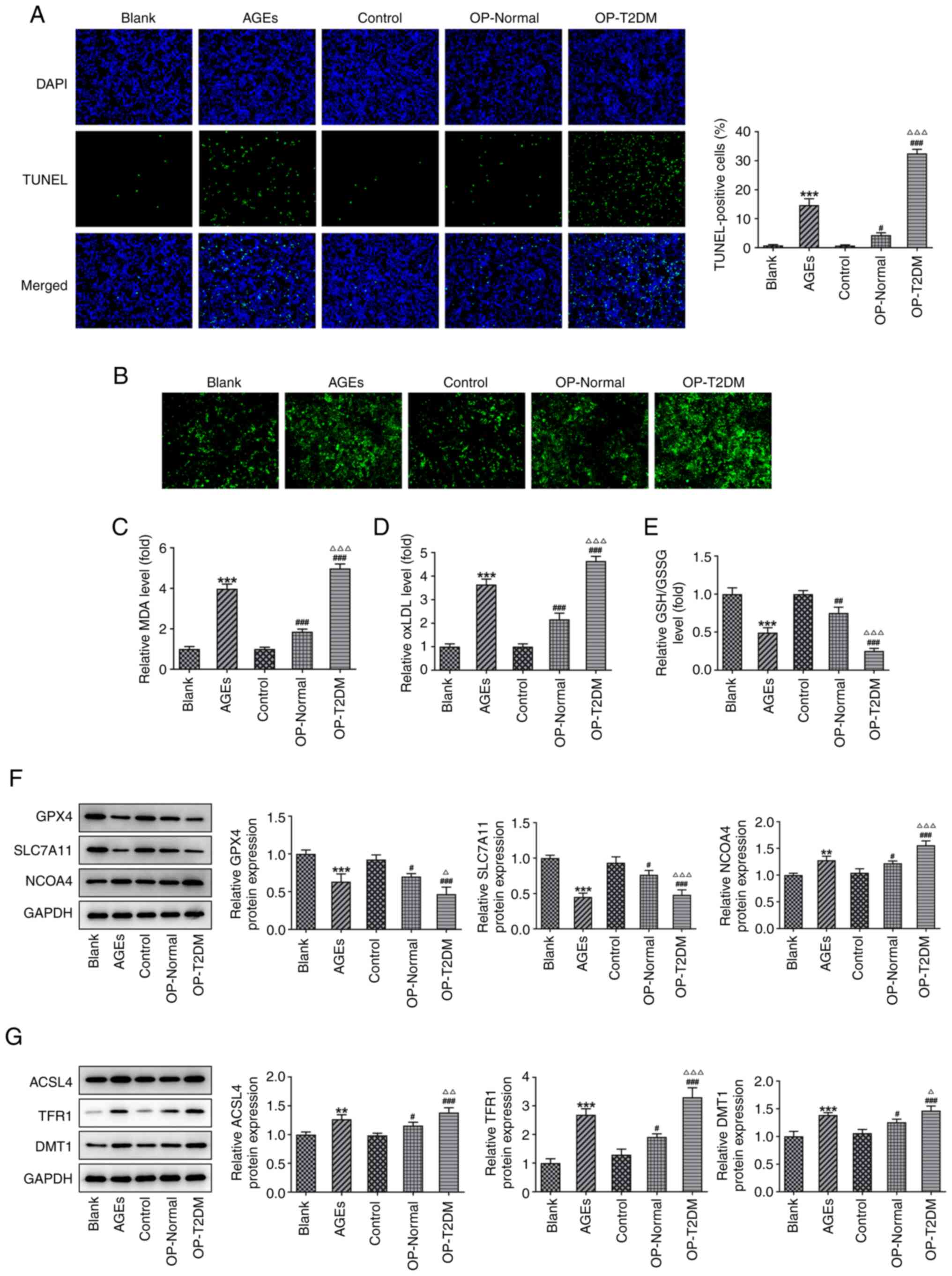 | Figure 3.AGEs and serum from patients with OP
induces apoptosis and ferroptosis in hFOB1.19 osteoblasts. (A)
Representative images and quantitative analysis of TUNEL staining
in hFOB1.19 cells from different treatment groups. (B) Cellular
free iron levels in hFOB1.19 osteoblasts in different treatment
groups. The relative levels of (C) cellular MDA, (D) ox-LDL and (E)
GSH/GSSG in hFOB1.19 osteoblasts in different treatment groups. (F
and G) The expression levels of proteins associated with
ferroptosis was measured using western blot analysis. **P<0.01
and ***P<0.001 vs. Blank; #P<0.05,
##P<0.01 and ###P<0.001 vs. Control;
∆P<0.05, ∆∆P<0.01 and
∆∆∆P<0.001 vs. OP-Normal. AGEs, advanced glycation
end products; OP, osteoporosis; MDA, malondialdehyde; ox-LDL,
oxidized low density lipoprotein; GSH/GSSG, glutathione/oxidized
GSH; T2DM, type-2 diabetes mellitus; GPX4, GSH peroxidase; SLC7A11,
solute carrier family 7 member 11; NCOA4, nuclear receptor
coactivator 4; ACSL4, acyl-CoA synthetase long-chain family member
4; TFR1, transferrin receptor; DMT1, divalent metal transporter
1. |
Ferroptosis inhibition reverses the
effect of AGEs and serum from patients with OP and T2DM on hFOB1.19
osteoblasts
To determine the role of ferroptosis in the
impairment of hFOB1.19osteoblast biological activities induced by
AGEs and serum from patients with OP and T2DM, the ferroptosis
inhibitor, DFO was added to the culture medium in the hFOB1.19
cells treated with AGEs and serum from patients with OP and T2DM
(20). The CCK-8 and ALP assays,
and Alizarin red staining results showed that the presence of DFO
promoted cell proliferation, ALP production and mineralized nodule
formation in the hFOB1.19 osteoblasts from the AGE and OP-T2DM
groups (Fig. 4A-C). Cell
apoptosis was measured using TUNEL staining, and the results showed
that DFO decreased the number of apoptotic cells in the AGEs and
OP-T2DM groups; however, the result was not significant
(P>0.05), (Fig. 4D). By
contrast, DFO significantly reduced cellular iron, MDA and oxLDL
levels, and increased GSH/GSSG levels in the hFOB1.19 osteoblasts
from the AGE and OP-T2DM groups (Fig.
4E-H), indicating a DFO-induced inhibition of ferroptosis.
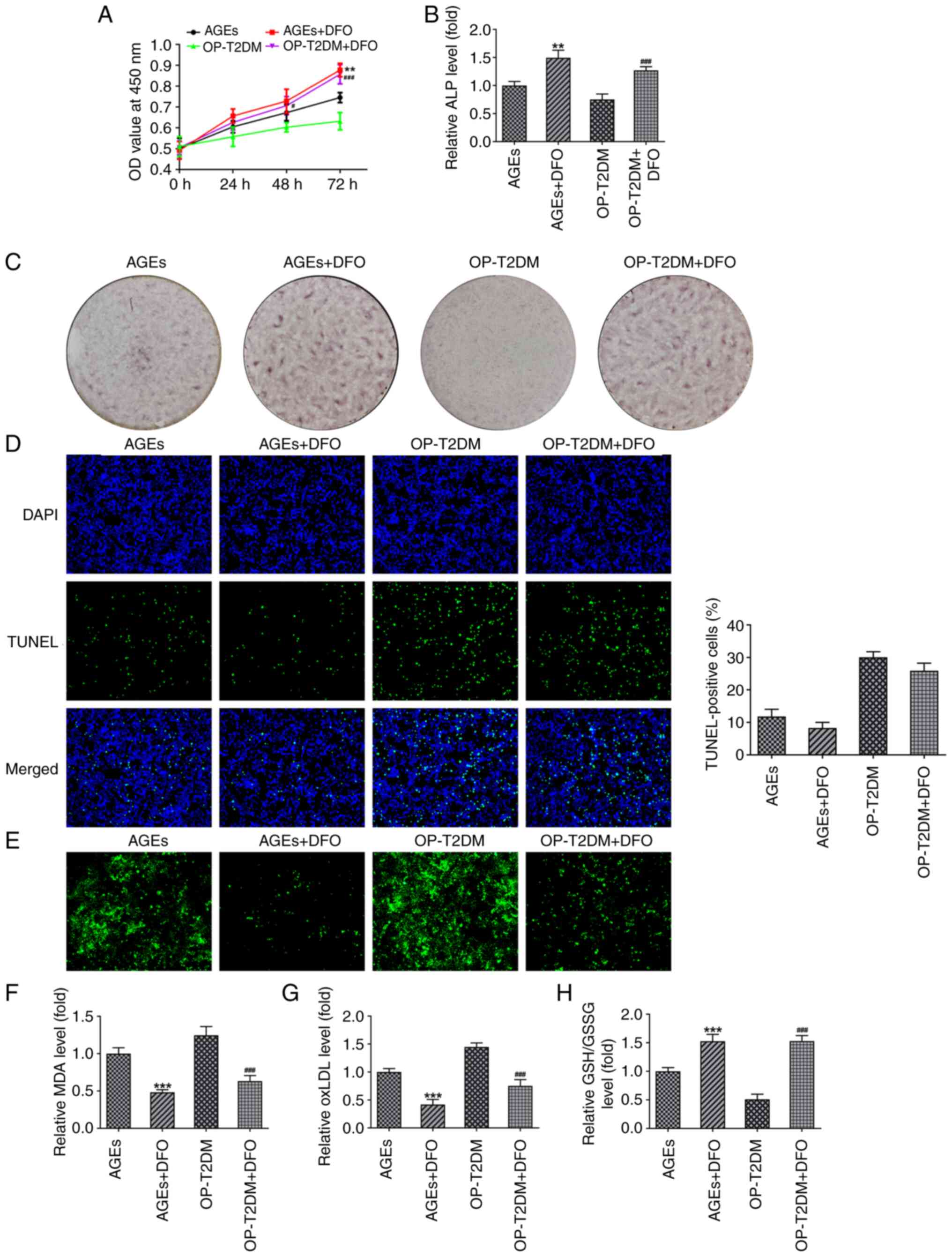 | Figure 4.Ferroptosis inhibitor, DFO, reverses
the effect of AGEs and serum from patients with OP and T2DM on
hFOB1.19 osteoblasts. (A) A Cell Counting Kit-8 assay was used to
detect proliferation in hFOB1.19 cells in different treatment
groups. (B) Cellular ALP level of hFOB1.19 osteoblasts was measured
using a ALP kit in different treatment groups. (C) Alizarin red
staining was used to observe the formation of mineralized nodules
in different treatment groups. (D) Representative TUNEL staining
and quantitative analysis for hFOB1.19 osteoblasts in different
treatment groups. (E) The cellular free iron level inhFOB1.19
osteoblasts in different treatment groups. The relative level of
cellular (F) MDA, (G) ox-LDL and (H) GSH/GSSG in hFOB1.19
osteoblasts in different treatment groups. **P<0.01 and
***P<0.001 vs. AGEs; #P<0.05 and
###P<0.001 vs. OP-T2DM. DFO, deferoxamine; AGEs,
advanced glycation end products; OP, osteoporosis; T2DM, type-2
diabetes mellitus; ALP, alkaline phosphatase; MDA, malondialdehyde;
ox-LDL, oxidized low density lipoprotein; GSH/GSSG,
glutathione/oxidized GSH; OD, optical density. |
Discussion
OP is a common debilitating bone disease
characterized by loss of bone mass and degradation of bone
architecture. The involvement of AGEs in OP, particularly
diabetes-related OP has been previously reported (21–23). In the present study, the
inhibitory effect of AGEs on osteoblast proliferation,
differentiation and mineralization was demonstrated, and the role
of ferroptosis in these effects was identified. The results
suggested that therapeutically targeting AGEs and AGE-induced
ferroptosis in osteoblasts may promote bone formation and even
relieve OP.
Patients with T2DM are more susceptible to OP, and
the formation and accumulation of AGEs have been reported to cause
diabetes-related OP (24).
Increased AGE production is observed in diabetic rats, and AGE
levels are negatively correlated with BMD (25). Higher levels of serum AGEs are
found in patients with OP and osteopenia compared with those in
healthy women (26).
Consistently, the results in the present study showed increased
serum AGE levels in patients with OP and a negative correlation
with BMD. Furthermore, a higher concentration of FBG was observed
in patients with OP compared with that in healthy individuals, and
a positive correlation between AGEs and FBG was observed in
patients with OP. The results further confirmed the crucial role of
AGEs and hyperglycemia in OP.
OP is also a disease caused by an imbalance between
osteoclasts and osteoblasts. Osteoblasts can contribute to bone
formation by secreting various proteins that guide the deposition
of bone extracellular matrix, such as ALP, osteocalcin and
osteopontin (27). The results
from the present study showed that AGEs and serum from patients
with OP, which showed a higher concentration of serum AGEs compared
with that in healthy individuals, significantly inhibited hFOB1.19
osteoblast proliferation, differentiation and mineralization.
Furthermore, the effect of serum from patients with OP and T2DM was
markedly stronger compared with serum from patients with OP and a
normal FBG level, demonstrating the detrimental effect of
hyperglycemia on human bone formation. These results were in
accordance with those of previous studies, which showed that
hyperglycemia and AGEs suppressed osteoblast differentiation and
mineralization, accompanied by an enhanced RAGE expression
(28–30). It has been reported that AGEs are
diverse compounds that are generated via a non-enzymatic reaction
between reducing sugars and the amine residues on proteins, lipids
and nucleic acids (9). Intra- and
extracellular AGEs accumulate in an age-dependent manner in human
tissue including cartilage, thus contributing to bone fragility in
patients with diabetes (31).
Therefore, the disruption of osteoblast function caused by serum
from patients with OP and T2DM may also be attributed to the high
production of AGEs.
To elucidate the mechanisms underlying the
destructive effect of AGEs on bone function, ferroptosis was
investigated, as it is a process that has been found to be involved
in OP pathogenesis in recent years (13,15,17,32). The results from the present study
showed that AGEs and serum from patients with OP triggered
apoptosis, and increased the cellular level of free iron, MDA and
oxLDL levels, and reduced GSH/GSSG level in hFOB1.19 osteoblasts.
Furthermore, AGEs and serum from patients with OP resulted in a
decrease in the protein expression levels of GPX4 and SLC7A11, but
an increase in the protein expression levels of NCOA4, ACSL4, TFR1
and DMT1. The accumulation of intracellular iron and lipid
peroxidates, such as MDA, oxLDL and GSH, which are normally
synthesized by ACSL4 and dissipated by the antioxidant enzyme GPX4,
is a key feature of ferroptosis (33). In addition, the downregulation of
SLC7A11 ultimately leads to a suppression of GPX4 activity and
ferroptosis activation. NCOA4 is an autophagy cargo receptor, TFR1
is a transferrin receptor and DMT1 transports metal ions across
membranes in mammals; all are necessary for iron release into the
cytoplasm (34). The potential
association between AGEs level and iron overload has been suggested
in previous studies (35,36). AGEs were also reported to enhance
heme oxygenase-1 (HO-1) mRNA expression in endothelial cells
(37) and inhibited Nrf2
activation in H9C2 cells (38).
Nrf2 is a transcription factor that regulates iron metabolism genes
in response to oxidative stress, and increased HO-1 activity has
been shown to increase cellular iron levels (39). The Nrf2/HO-1 signaling pathway has
been reported to increase the cellular iron levels and promote
ferroptosis (13,40). The aforementioned studies have
found an association between AGEs and ferroptosis; however, to the
best of our knowledge, the present study was the first to
demonstrate the effect of AGEs on ferroptosis. Subsequently, the
ferroptosis inhibitor DFO was found to not only suppress
ferroptosis, but also induce a recovery of hFOB1.19 osteoblast
proliferation, differentiation and mineralization in the AGE, and
OP-T2DM groups. However, DFO had no significant effect on apoptosis
in the hFOB1.19 cells treated with AGE and serum from patients with
OP and T2DM. This may be due to the specific inhibition of DFO on
ferroptosis without affecting apoptosis. This was in line with our
hypothesis, suggesting that AGEs may disrupt osteoblast function by
inducing ferroptosis. Nevertheless, the present results remain to
be validated using in vivo animal models, which will be
performed in future research.
In conclusion, the results from the present study
demonstrated the inhibitory effect of AGEs on bone formation in OP.
AGEs significantly suppressed proliferation, differentiation and
mineralization in hFOB1.19 osteoblasts. In addition, ferroptosis
influenced the effects of AGEs. To the best of our knowledge, the
present study was the first to uncover the mechanisms mediating the
effect of AGEs on osteoblast function and demonstrated the role of
osteoblast ferroptosis in bone formation. Therapies targeting AGEs
and ferroptosis may be valuable in relieving OP.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
WLu, WG and JJ contributed to the conception or
design of the study and performed the experiments. WW, JJ, JY, WLi
and YC contributed to data analysis and interpretation. WLu, WG and
JJ drafted the manuscript and revised it critically for important
intellectual content. All authors read and approved the final
manuscript. WLu, WG and JJ confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
All the experiments were approved by the Ethics
Committee of the Nanjing Pukou District Central Hospital (Jiangsu,
China). Written informed consent was obtained from all
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Baccaro LF, Conde DM, Costa-Paiva L and
Pinto-Neto AM: The epidemiology and management of postmenopausal
osteoporosis: A viewpoint from Brazil. Clin Interv Aging.
10:583–591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Armas LA and Recker RR: Pathophysiology of
osteoporosis: New mechanistic insights. Endocrinol Metab Clin North
Am. 41:475–486. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Reid IR: A broader strategy for
osteoporosis interventions. Nat Rev Endocrinol. 16:333–339. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ensrud KE, Ewing SK, Taylor BC, Fink HA,
Stone KL, Cauley JA, Tracy JK, Hochberg MC, Rodondi N and Cawthon
PM; Study of Osteoporotic Fractures Research Group, : Frailty and
risk of falls, fracture, and mortality in older women: The study of
osteoporotic fractures. J Gerontol A Biol Sci Med Sci. 62:744–751.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Johnston CB and Dagar M: Osteoporosis in
older adults. Med Clin North Am. 104:873–884. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kurra S, Fink DA and Siris ES:
Osteoporosis-associated fracture and diabetes. Endocrinol Metab
Clin North Am. 43:233–243. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nowotny K, Jung T, Höhn A, Weber D and
Grune T: Advanced glycation end products and oxidative stress in
type 2 diabetes mellitus. Biomolecules. 5:194–222. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goh SY and Cooper ME: Clinical review: The
role of advanced glycation end products in progression and
complications of diabetes. J Clin Endocrinol Metab. 93:1143–1152.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamamoto M and Sugimoto T: Advanced
glycation end products, diabetes, and bone strength. Curr
Osteoporos Rep. 14:320–326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Alikhani M, Alikhani Z, Boyd C, MacLellan
CM, Raptis M, Liu R, Pischon N, Trackman PC, Gerstenfeld L and
Graves DT: Advanced glycation end products stimulate osteoblast
apoptosis via the MAP kinase and cytosolic apoptotic pathways.
Bone. 40:345–353. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Notsu M, Yamaguchi T, Okazaki K, Tanaka K,
Ogawa N, Kanazawa I and Sugimoto T: Advanced glycation end product
3 (AGE3) suppresses the mineralization of mouse stromal ST2 cells
and human mesenchymal stem cells by increasing TGF-β expression and
secretion. Endocrinology. 155:2402–2410. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meng HZ, Zhang WL, Liu F and Yang MW:
Advanced glycation end products affect osteoblast proliferation and
function by modulating autophagy via the receptor of advanced
glycation end Products/Raf Protein/Mitogen-activated protein
Kinase/Extracellular Signal-regulated Kinase Kinase/Extracellular
Signal-regulated Kinase (RAGE/Raf/MEK/ERK) pathway. J Biol Chem.
290:28189–28199. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma H, Wang X, Zhang W, Li H, Zhao W, Sun J
and Yang M: Melatonin suppresses ferroptosis induced by high
glucose via activation of the Nrf2/HO-1 signaling pathway in type 2
diabetic osteoporosis. Oxid Med Cell Longev. 2020:90676102020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou B, Liu J, Kang R, Klionsky DJ,
Kroemer G and Tang D: Ferroptosis is a type of autophagy-dependent
cell death. Semin Cancer Biol. 66:89–100. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu J, Yang J, Zheng Y, Chen X and Fang S:
Extracellular vesicles from endothelial progenitor cells prevent
steroid-induced osteoporosis by suppressing the ferroptotic pathway
in mouse osteoblasts based on bioinformatics evidence. Sci Rep.
9:161302019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia D, Wu J, Xing M, Wang Y, Zhang H, Xia
Y, Zhou P and Xu S: Iron overload threatens the growth of
osteoblast cells via inhibiting the PI3K/AKT/FOXO3a/DUSP14
signaling pathway. J Cell Physiol. Jan 29–2019.(Epub ahead of
print). doi: 10.1002/jcp.28217. View Article : Google Scholar
|
|
17
|
Wang X, Ma H, Sun J, Zheng T, Zhao P, Li H
and Yang M: Mitochondrial ferritin deficiency promotes osteoblastic
ferroptosis via mitophagy in type 2 diabetic osteoporosis. Biol
Trace Elem Res. 200:298–307. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tsukushi S, Katsuzaki T, Aoyama I,
Takayama F, Miyazaki T, Shimokata K and Niwa T: Increased
erythrocyte 3-DG and AGEs in diabetic hemodialysis patients: Role
of the polyol pathway. Kidney Int. 55:1970–1976. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim HJ and Kim SG: Alterations in cellular
Ca(2+) and free iron pool by sulfur amino acid deprivation: The
role of ferritin light chain down-regulation in prooxidant
production. Biochem Pharmacol. 63:647–657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun Y, Zheng Y, Wang C and Liu Y:
Glutathione depletion induces ferroptosis, autophagy, and premature
cell senescence in retinal pigment epithelial cells. Cell Death
Dis. 9:7532018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng YZ, Yang SL, Wang JY, Ye M, Zhuo XY,
Wang LT, Chen H, Zhang H and Yang L: Irbesartan attenuates advanced
glycation end products-mediated damage in diabetes-associated
osteoporosis through the AGEs/RAGE pathway. Life Sci. 205:184–192.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sanguineti R, Puddu A, Mach F, Montecucco
F and Viviani GL: Advanced glycation end products play adverse
proinflammatory activities in osteoporosis. Mediators Inflamm.
2014:9758722014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Y, Wang L, Zhang M, Huang K, Yao Z, Rao
P, Cai X and Xiao J: Advanced glycation end products inhibit the
osteogenic differentiation potential of adipose-derived stem cells
by modulating Wnt/β-catenin signalling pathway via DNA methylation.
Cell Prolif. 53:e128342020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
de Paula FJ, Horowitz MC and Rosen CJ:
Novel insights into the relationship between diabetes and
osteoporosis. Diabetes Metab Res Rev. 26:622–630. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saito M, Fujii K, Mori Y and Marumo K:
Role of collagen enzymatic and glycation induced cross-links as a
determinant of bone quality in spontaneously diabetic WBN/Kob rats.
Osteoporos Int. 17:1514–1523. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang DH, Chiang TI, Chang IC, Lin FH, Wei
CC and Cheng YW: Increased levels of circulating advanced glycation
end-products in menopausal women with osteoporosis. Int J Med Sci.
11:453–460. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee WC, Guntur AR, Long F and Rosen CJ:
Energy metabolism of the osteoblast: Implications for osteoporosis.
Endocr Rev. 38:255–266. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sanguineti R, Storace D, Monacelli F,
Federici A and Odetti P: Pentosidine effects on human osteoblasts
in vitro. Ann NY Acad Sci. 1126:166–172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Franke S, Rüster C, Pester J, Hofmann G,
Oelzner P and Wolf G: Advanced glycation end products affect growth
and function of osteoblasts. Clin Exp Rheumatol. 29:650–660.
2011.PubMed/NCBI
|
|
30
|
Lee EJ, Kang MK, Kim YH, Kim DY, Oh H, Kim
SI, Oh SY, Na W and Kang YH: Coumarin ameliorates impaired bone
turnover by inhibiting the formation of advanced glycation end
products in diabetic osteoblasts and osteoclasts. Biomolecules.
10:10522020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dariya B and Nagaraju GP: Advanced
glycation end products in diabetes, cancer and phytochemical
therapy. Drug Discov Today. 25:1614–1623. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ni S, Yuan Y, Qian Z, Zhong Z, Lv T, Kuang
Y and Yu B: Hypoxia inhibits RANKL-induced ferritinophagy and
protects osteoclasts from ferroptosis. Free Radic Biol Med.
169:271–282. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Casati L, Pagani F, Fibiani M, Lo Scalzo R
and Sibilia V: Potential of delphinidin-3-rutinoside extracted from
Solanum melongena L. as promoter of osteoblastic MC3T3-E1
function and antagonist of oxidative damage. Eur J Nutr.
58:1019–1032. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hirschhorn T and Stockwell BR: The
development of the concept of ferroptosis. Free Radic Biol Med.
133:130–143. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mirlohi MS, Yaghooti H, Shirali S,
Aminasnafi A and Olapour S: Increased levels of advanced glycation
end products positively correlate with iron overload and oxidative
stress markers in patients with β-thalassemia major. Ann Hematol.
97:679–684. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen SH, Yuan KC, Lee YC, Shih CK, Tseng
SH, Tinkov AA, Skalny AV and Chang JS: Iron and advanced glycation
end products: Emerging role of Iron in androgen deficiency in
obesity. Antioxidants (Basel). 9:2612020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He M, Siow RC, Sugden D, Gao L, Cheng X
and Mann GE: Induction of HO-1 and redox signaling in endothelial
cells by advanced glycation end products: A role for Nrf2 in
vascular protection in diabetes. Nutr Metab Cardiovasc Dis.
21:277–285. 2011.PubMed/NCBI
|
|
38
|
Ko SY, Chang SS, Lin IH and Chen HI:
Suppression of antioxidant Nrf-2 and downstream pathway in H9c2
cells by advanced glycation end products (AGEs) via ERK
phosphorylation. Biochimie. 118:8–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Suttner DM and Dennery PA: Reversal of
HO-1 related cytoprotection with increased expression is due to
reactive iron. FASEB J. 13:1800–1809. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu T, Ding W, Ji X, Ao X, Liu Y, Yu W and
Wang J: Molecular mechanisms of ferroptosis and its role in cancer
therapy. J Cell Mol Med. 23:4900–4912. 2019. View Article : Google Scholar : PubMed/NCBI
|