Introduction
Chronic thromboembolic pulmonary hypertension
(CTEPH) is one of the leading causes of pulmonary hypertension
(PH), which presents in 2–4% of patients who have acute pulmonary
embolism (1,2). A previous study revealed that vascular
remodeling in the small pulmonary arteries is associated with the
progression of CTEPH (3).
Additionally, pulmonary artery endothelial cells (PAECs) were
revealed to exhibit hyperproliferative potential while inhibiting
apoptosis, implying that dysfunctional PAECs are involved in CTEPH
(4). To date, CTEPH remains
underdiagnosed and the exact prevalence and incidence of CTEPH in
patients with PH remain unclear (1). Although vascular disobliteration via
pulmonary endarterectomy is thought to be a potential strategy for
CTEPH treatment, it is not suitable for all patients. Therefore, it
is urgent to investigate the mechanism underlying the pathology of
CTEPH and develop novel strategies for diagnosing and treating
CTEPH.
Autophagy is an evolutionarily conserved but dynamic
process associated with the turnover of aggregated or dysfunctional
cytoplasmic proteins, intracellular pathogens and aged organelles
through lysosome-dependent degradation pathways (5). The primary function of autophagy is to
protect the living cell against various pathologies, including
aging, heart disease, cancer and virus infections (6). On the basis of PH-associated diseases,
Lee et al (7) reported that
autophagic protein microtubule-associated protein-1 light chain 3B
(LC3B) functions as a protective factor in hypoxic-induced PH. In
addition, Lahm et al (8)
demonstrated that enhanced autophagy is associated with the
17β-estradiol-mediated protective role in hypoxia-induced PH.
Furthermore, Long et al (9)
reported that the suppression of autophagy with Chloroquine serves
mitigative roles in a rat pulmonary arterial hypertension (PAH)
model. Taken together, these results indicated that autophagy may
be an essential regulator in the development and progression of
PH-induced pathological processes.
Spermidine (SP) is an achiral organic polycation
that is found in all eukaryotic cells in low concentrations
(millimolar) (10). Additionally,
SP exerts multifunctional roles in a series of cellular activities,
including anti-inflammatory, anti-oxidant, mitochondrial function
and proteostasis (11). In recent
years, it has been demonstrated that the physiological effect of SP
is tightly associated with its inductive effect on cytoprotective
autophagy (10,12). Exogenous SP promotes autophagy in
yeast, flies, worms and human immune cells, which in turn increases
the lifespan of these species (13). In humans and mice, vascular
endothelial cell aging is associated with aberrant expression of
autophagy marker proteins, including LC3-II and Beclin1, arterial
endothelium-associated dilatation and enhanced oxidative stress,
which is reversed by SP (14).
However, the mechanism underlying such positive effects of SP
requires further investigation.
Long non-coding RNAs (lncRNAs) are a class of
transcribed RNAs >200 nt in length (15). The lncRNAs modulate gene expression
through various mechanisms and serve essential roles in various
biological and pathological processes (16). As a multifunctional lncRNA, growth
arrest-specific transcript 5 (GAS5) participates in several
pathological processes, including cancer cell apoptosis and
proliferation (17,18), cardiac fibroblast fibrosis (19) and epithelial-mesenchymal transitions
in osteosarcoma (20). Furthermore,
GAS5 is involved in the regulation of autophagy in breast cancer
and non-small-cell lung cancer (NSCLC) (21,22),
suggesting a regulatory role of GAS5 in the autophagic process.
Therefore, based on previous findings, the present study aimed to
investigate the effects of SP on PAECs of patients with CTEPH and
elucidate the role of GAS5 in this process and associated signaling
pathways.
Materials and methods
Ethics statement
All patients provided written informed consent prior
to inclusion in the present study. All experimental protocols were
performed in accordance with the Declaration of Helsinki. The
experimental protocols were approved by the Ethics Committee of
Cangzhou Central Hospital (Cangzhou, China). All experimental
procedures involving animals were approved by the Institutional
Animal Care and Use Committee of Cangzhou Central Hospital. All
animal experiments were performed according to the guidelines of
the local regulatory agencies and conformed to the Regulations for
the Management of Laboratory Animals published by the Ministry of
Science and Technology of the People's Republic of China.
Patients and samples
Proximal pulmonary vascular tissues were obtained
from six patients with CTEPH who had undergone lung
transplantations or surgical pulmonary endarterectomy (PEA) at
Cangzhou Central Hospital (Cangzhou, China), between June 2017 and
June 2018. The clinical features of the patients are summarized in
Table I. Patients enrolled in this
study were previously diagnosed with CTEPH by ventilation-perfusion
lung scans, right heart cardiac catheterization, or computed
tomography pulmonary angiography. Patients with the following
conditions were excluded from this study, including psychosis,
severe obstructive pulmonary disease, chronic liver disease,
chronic kidney disease, amyloidosis, portal hypertension, drug
addiction history, intellectual disability, etc. In addition,
patients who had taken prostacyclin, L-arginine, sildenafil and
endothelin receptor antagonist were also excluded. The tissues used
to isolate cells were free of thrombotic material and contained
neointima and media. The tissues were transported from the
operating room to the laboratory for subsequent processes
immediately after.
 | Table I.Clinical features of the
patients. |
Table I.
Clinical features of the
patients.
| Patient ID | Age, year | Sex | PaO2, mm
Hg | PaCO2,
mm Hg | Ppa, mm Hg | CI,
L/min/m2 | PVR, dyne × s/cm
5 |
|---|
| 001 | 43 | F | 72 | 38.9 | 40 | 3.65 | 590 |
| 002 | 65 | M | 55 | 39.5 | 43 | 3.11 | 510 |
| 003 | 61 | F | 58 | 34.6 | 57 | 2.89 | 879 |
| 004 | 54 | M | 63 | 37.6 | 42 | 3.78 | 654 |
| 005 | 37 | F | 78 | 39.8 | 51 | 4.03 | 397 |
| 006 | 46 | F | 64 | 33.6 | 63 | 3.87 | 898 |
Cell culture
The PAECs were isolated by collagenase digestion
followed by immunomagnetic separation using anti-CD31 monoclonal
antibody-labeled beads (Abcam). The PAECs were cultured in M199
medium (Life Technologies; Thermo Fisher Scientific, Inc.),
supplemented with 20% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), streptomycin (100 µg/ml), fungizone (1.25
µg/ml), penicillin (100 U/ml), heparin (10 U/ml) and α-FGF (5
ng/ml; Abcam) at 37°C in a humidified atmosphere with 5%
CO2. The PAECs (at least 6 passages) were used for
subsequent experiments.
Immunofluorescence assay
The PAECs were fixed with 4% paraformaldehyde for 15
min at room temperature and washed with 0.1 M PBS three times.
Next, the PAECs were permeabilized with 0.1% Triton X-100
(Sigma-Aldrich; Merck KGaA) and washed with 0.1 M PBS three times.
Cells were blocked with 10% normal donkey serum (Sigma-Aldrich;
Merck KGaA) for 20 min at room temperature. The PAECs were
incubated with the primary antibody against LC3B (dilution, 1:100;
cat. no. sc-376404; Santa Cruz Biotechnology, Inc.) overnight at
4°C and then incubated with the fluorescein-conjugated secondary
antibody (dilution, 1:200; cat. no. sc-516102; Santa Cruz
Biotechnology, Inc.) for 1 h at 37°C. Following washing three times
with 0.1 M PBS, cells were administered with
4′,6-diamidino-2-phenylindole (Santa Cruz Biotechnology, Inc.) for
10 min and then washed three times with 0.1 M PBS. Cells were
imaged by confocal fluorescence microscopy (×20 magnification)
(Olympus Corporation).
Western blotting
Total protein was extracted from PAECs using cell
lysis buffer (Thermo Fisher Scientific, Inc.). Western blotting was
performed as previously described (23). The primary antibodies against LC3B
(dilution, 1:1,000; cat. no. sc-271625), Beclin-1 (dilution, 1:500;
cat. no. sc-48341), ATG7 (dilution, 1:1,000; cat. no. sc-376212)
and ACTB (dilution, 1:5,000; cat. no. sc-8432) were purchased from
Santa Cruz Biotechnology, Inc. and NAT8L (dilution, 1:1,000; cat.
no. ab76842) was purchased from Abcam. The primary antibodies were
applied overnight at 4°C. Mouse IgG secondary antibody (dilution,
1:10,000; cat. no. sc-525409) (Santa Cruz Biotechnology, Inc.) and
goat anti-rabbit IgG H&L (HRP) (dilution, 1:10,000; cat. no.
ab205718; Abcam) were applied as the secondary antibody for 1 h at
37°C. The optical density of protein bands was quantified using the
ImageJ software version 1.48 (National Institutes of Health)
(24).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNAs were isolated from PAECs using the TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). First-strand
cDNAs were synthesized using the M-MLV Reverse Transcriptase (RNase
H) kit (GeneCopoeia, Inc.), according to the manufacturer's
protocols. Real-time PCR reactions were conducted on an ABI StepOne
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientiifc,
Inc.) using the recommended reaction conditions in the
manufacturer's protocols. The primers were synthesized by Shanghai
GenePharma Co., Ltd. and summarized in Table II. The abundance of miRNA-31-5p was
quantified by Hairpin-it microRNA and RNU6 snRNA Normalization
RT-PCR Quantitation kit (Shanghai GenePharma Co., Ltd.).
Additionally, GAPDH was used as the internal control for mRNA and
lncRNA, and U6 was used as the internal control for miRNA. Data
were analyzed using the 2−ΔΔCt method (25).
| Table II.Reverse transcription-quantitative
polymerase chain reaction primer sequences. |
Table II.
Reverse transcription-quantitative
polymerase chain reaction primer sequences.
| Gene name | Primer
sequence |
|---|
| LncRNA GAS5
forward |
5′-GTGTGGCTCTGGATAGCAC-3′ |
| LncRNA GAS5
reverse |
5′-ACCCAAGCAAGTCATCCATG-3′ |
| miRNA-31-5p |
5′-AGGCAAGATGCTGGCATAG-3′ |
| RNU6 forward |
5′-CGCTTCGGCAGCACATATACTAAAATTGGAAC-3′ |
| RNU6 reverse |
5′-GCTTCACGAATTTGCGTGTCATCCTTGC-3′ |
| GAPDH forward |
5′-AGAAGGCTGGGGCTCATTTG-3′ |
| GAPDH reverse |
5′-AGGGGCCATCCACAGTCTTC-3′ |
Nuclear/cytoplasmic RNA separation
analysis
Nuclear and cytoplasmic RNA isolation in PAECs were
performed as previously described (26). The abundance of GAS5 in each
cellular fraction was assessed by RT-qPCR.
5′ and 3′ rapid amplification of cDNA
ends (RACE)
To determine the transcriptional initiation and
termination sites of GAS5, 5′-RACE and 3′-RACE assays were
performed using the 5′/3′ RACE kit (2nd Generation; cat. no.
3353621001; Sigma-Aldrich; Merck KGaA), according to the
manufacturer's protocols.
Cell transfection
The plasmid pcDNA3.1-GAS5 (2 µg) containing the full
length of GAS5 (2,651 bp) were obtained from Shanghai GenePharma
Co., Ltd. The pcDNA3.1-empty vector (2 µg) was used as the negative
control (pcDNA3.1). Small interfering RNAs (siRNAs) of GAS5
(5′-UCUUCAAUCAUGAAUUCUGAG-3′; 20 pmol) and NAT8L
(5′-CUUUAAUUCUUGGGACAAA-3′; 20 pmol) were also obtained from
Shanghai GenePharma Co., Ltd. Scramble siRNA was used as the
negative control (5′-ACGUGACACGUUCGGAGAATT-3′; 20 pmol;
siRNA-control). miRNA-31-5p mimics (100 nM) and inhibitor (100 nM)
were synthesized by Shanghai GenePharma Co., Ltd. and the sequences
were as following: Mimics, forward 5′-AGGCAAGAUGCUGGCAUAGCU-3′ and
reverse 5′-CUAUGCCAGCAUCUUGCCUUU-3′; inhibitor,
5′-AGCUAUGCCAGCAUCUUGCCU-3′; mimic control, forward
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse
5′-ACGUGACACGUUCGGAGAATT-3′; inhibitor control,
5′-CAGUACUUUUGUGUAGUACAA-3′. Cell transfection was conducted using
the Lipofectamine™ 3000 Transfection Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocols. After 24 h of transfection at 37°C, the
PAECs were immediately used for subsequent experiments.
Bioinformatics and luciferase reporter
assay
The GAS5 sequence was analyzed by an online
database, lncipedia (27). The
subcellular location of GAS5 was predicted by an online tool,
lncLocator (28). The putative
binding sites of miRNA-31-5p were predicted by online tools,
StarBase 2.0 (29), TargetScan
(30) and Miranda (31). Based on the predictions from online
databases and preliminary experiments, targeting genes were
selected for subsequent experiments. The luciferase vectors,
including wild-type or mutant 3′-UTR of GAS5 and NAT8L containing
the miRNA-31-5p binding site were synthesized by Shanghai
GenePharma Co., Ltd. Plasmid DNA and miRNA-31-5p mimic were
co-transfected in PAECs using the Lipofectamine™ 3000
Transfection Reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocols. After 48 h of
transfection at 37°C, luciferase activity was quantified using the
Dual-Light Chemiluminescent Reporter Gene assay system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and normalized to the
Renilla luciferase activity.
RNA-binding protein
immunoprecipitation (RIP) assay
The RIP assay was performed using a RNA
immunoprecipitation kit (cat. no. ab206996; Abcam), according to
the manufacturer's protocols. The PAECs were lysed and incubated
with magnetic beads llama monoclonal to GFP VHH single domain (cat.
no. ab193983; Abcam) conjugated with human anti-AGO2 antibody or
anti-mouse IgG (cat. nos. ab186733 and ab150113; Abcam). The
abundance of immunoprecipitated RNA was quantified by qPCR assay as
aforementioned.
CTEPH rat model establishment
Twelve healthy male Sprague Dawley rats (6 weeks
old; body weight, 220±15 g) were obtained from Beijing Vital River
Laboratory Animals Co., Ltd. The animals were housed in an isolated
room at 20–24°C and 65–70% humidity under a 12-h light/dark cycle.
All rats had free access to food and water. The rats' activity was
restricted one week before the surgical procedure to mimic blood
stasis in the clinical setting. Rats were randomly divided into
three groups (n=4): sham, SP and SP+siRNA-GAS5. Autologous blood
clots were prepared one day ahead of the injection day followed by
the previously reported procedure (32). During surgery, rats were fixed on an
operating table and were anesthetized by intraperitoneally
injecting 10% chloral hydrate (0.3 g/kg). No signs of peritonitis
or pain were observed in rats intraperitoneally injected with
chloral hydrate. Autologous blood clots were aspirated into a
syringe with 2 ml saline containing tranexamic acid (200 mg/kg/rat)
and injected into the left external jugular vein of experimental
rats three times on day 1, 3 and 7, respectively. Saline without
tranexamic acid was injected into rats in the sham group. During
the experiment, endogenous fibrinolysis was suppressed by
intraperitoneal injection of tranexamic acid (200 mg/kg) once per
day. Following the operation, penicillin (10,000 U/kg/day) was
administrated to rats for 3 days to prevent infection. Pulmonary
arterial pressure of rats was measured to validate the CTEPH rat
model. Three days after the last injection, rats in the SP group
were intraperitoneally injected with SP (5 mg/kg) dissolved in PBS
daily. Rats in the SP+siRNA-GAS5 group received an injection of the
adenovirus siRNA-GAS5 following a slightly adjusted procedure as
previously reported (33). After 5
days of adenovirus delivery, rats were euthanized by exsanguination
under deep chloral hydrate anesthesia (0.3 g/kg), and the proximal
pulmonary vascular tissues were collected for subsequent
experiments.
Histopathology
Proximal pulmonary vascular tissues collected from
CTEPH rats were fixed with 10% formaldehyde for 24 h at room
temperature. Next, the tissues were embedded in paraffin and
stained with hematoxylin and eosin (H&E) for 3 min at room
temperature. The tissue sections were imaged using an optical
microscope (×40 magnification; DMI3000M; Leica Microsystems,
Inc.).
GFP-LC3 adenovirus assay
The PAECs collected from CTEPH rats were transfected
with GFP-LC3 adenovirus (Hanbio Biotechnology Co., Ltd.) for 24 h,
according to the manufacturer's protocols. The GFP-labeled
autophagic vesicles in PAECs were evaluated using the ImageJ
software (24).
GFP and RFP tandemly tagged LC3
(tfLC3) assay
The PAECs collected from CTEPH rats were transfected
with mRFP-GFP-LC3 plasmids for 24 h using the Premo™
Autophagy Tandem Sensor RFP-GFP-LC3B kit (Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocols. Next, the PAECs
were imaged by fluorescence microscopy (×20 magnification; Hitachi
High-Technologies Corporation) and quantified using ImageJ software
(24).
Transmission electron microscopy
(TEM)
PAECs were collected by trypsinization and fixed in
0.1 M cacodylate buffer containing 4% paraformaldehyde and 2.5
gluteraldehyde for 24 h at 4°C, and then further fixed in 1% osmium
tetroxide buffer for 1 h at 4°C. Next, PAECs were dehydrated by a
graded series of acetone and embedded in spur resin. The slices
were sectioned (500 nm) using a HM 355S Automatic Microtome (Thermo
Fisher Scientific, Inc.) and then stained with saturated solution
of uranyl acetate and lead citrate in the dark for 1 h at room
temperature. Images of autophagic vacuoles were captured using a
JEM-2100Plus Transmission Electron Microscope (×10,000
magnification; JEOL, Ltd.).
Statistical analysis
In the present study, each treatment group consisted
of 3–5 replicates. Statistical analysis was performed using SPSS
v.19.0 software (IBM Corp.). Data are presented as the mean ±
standard error of mean. Differences between two groups were
analyzed by Student's t-test, and multiple comparisons analysis was
performed by one-way analysis of variance and Tukey's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
GAS5 increases in SP-induced autophagy
in PAECs derived from patients with CTEPH
Spermidine is an autophagy inducer in numerous types
of cells (10,12). Using immunofluorescence assays,
Western blotting and TEM to test the effect of SP on PAECs derived
from patients with CTEPH, the results revealed that concentrations
of SP (10 and 100 µM) could significantly increase the level of
autophagy in PAECs derived from CTEPH in a dose-dependent manner
(Fig. 1A and B). Additionally,
protein expression of autophagy markers, beclin-1 and ATG7, was
increased by SP treatment (Fig. 1C)
(34–36). For the subsequent experiments, the
10 µM dose of SP was used. Since GAS5 acts as an anti-cancer
regulator through modulating autophagy (21,22),
and PAECs and tumor cells exhibit hyper-proliferation and
hypo-apoptosis (37,38), GAS5 may be involved in SP-induced
autophagy in PAECs. To verify our hypothesis, the expression of
GAS5 was measured in SP-treated PAECs. The SP treatment increased
the expression of GAS5 in a dose-dependent pattern in PAECs
(Fig. 1D). The sequence of GAS5 was
analyzed using RACE assays which demonstrated that GAS5 was located
in the reverse strand of chromosome 1 (hg38), was 723 bp in full
length and consisted of 13 exons (Fig.
1E). The protein coding potential of GAS5 was predicted to be
non-coding in four of the five predictive metrics (Fig. 1F). On a lncLocator platform, GAS5
was identified to be primarily distributed in the cytoplasm
(Fig. 1G), which was verified using
the nuclear/cytoplasmic RNA separation analysis. The expression of
cytoplasmic GAS5 was significantly higher than that in the nucleus
(Fig. 1H). The SP increased the
expression of GAS5 in the cytoplasm, but not in the nucleus
(Fig. 1I). Taken together, these
results suggested that GAS5 may be involved in the SP-induced
autophagy in PAECs originated from CTEPH.
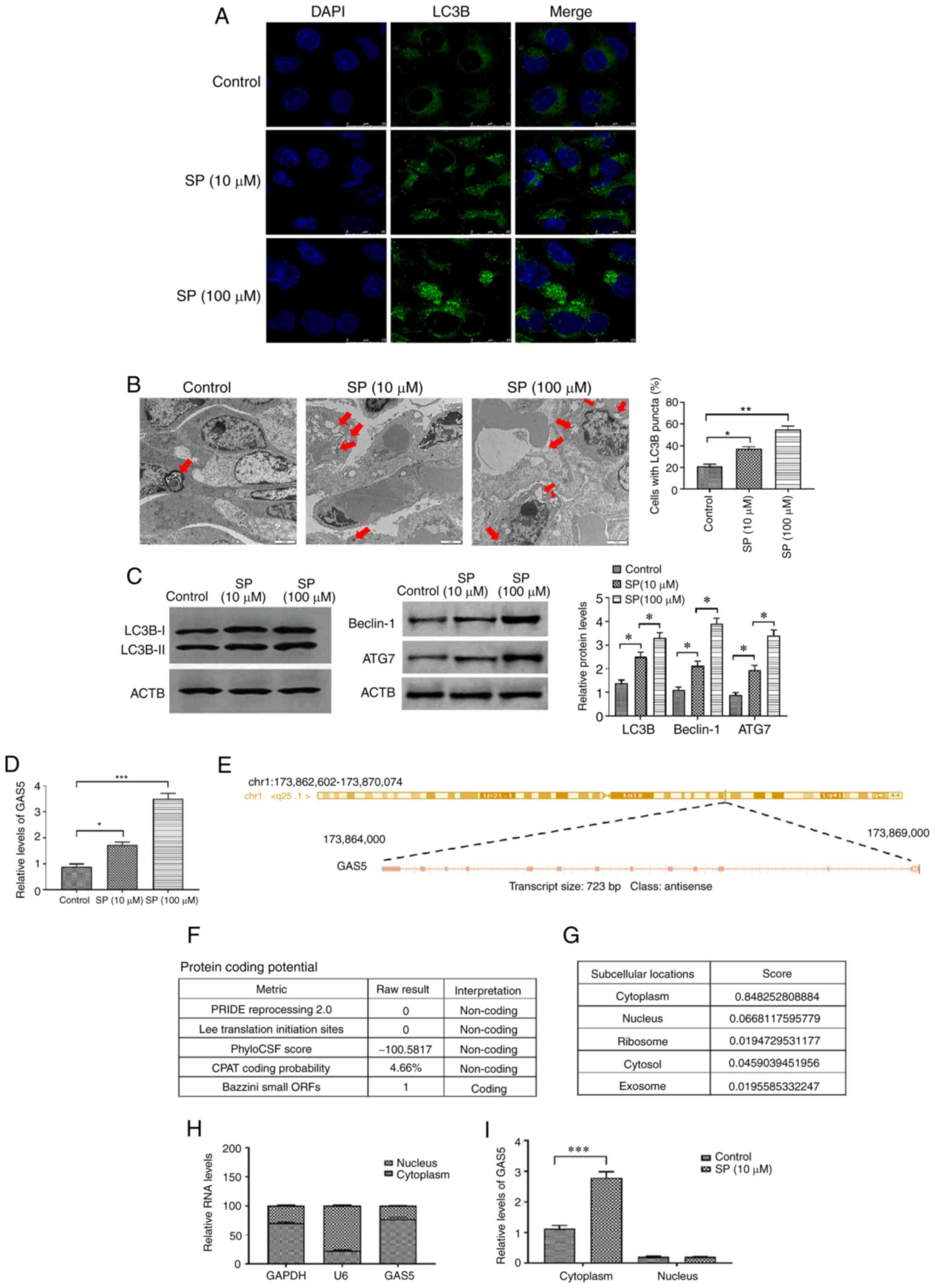 | Figure 1.SP promotes autophagy and increases
GAS5 in PAECs. (A) Abundance of LC3B in SP (10 and 100 µM)-treated
PAECs, as assessed by immunofluorescence assays (scale bar, 25 µm;
magnification, ×20). (B) Autophagic vacuoles in the cellular
cytoplasm of SP (10 and 100 µM)-treated PAECs, as evaluated by
transmission electron microscopy (scale bar, 2 µm; magnification,
×10,000). Red arrows indicate the autophagic vacuoles. (C) Protein
expression of LC3B, Beclin-1 and ATG7 in SP (10 and 100 µM)-treated
PAECs were assessed by Western blotting. (D) Expression of GAS5 in
PAECs treated with SP (10 and 100 µM). (E) Sequence information of
GAS5. (F) Prediction of protein-coding potential of GAS5. (G)
Prediction of subcellular location of GAS5. (H) The subcellular
location of GAS5 was assessed by nuclear/cytoplasmic RNA separation
assays. (I) Effect of SP (10 µM) on the subcellular location of
GAS5. *P<0.05; **P<0.01; ***P<0.001. There were at least
three replicates in each group available for analysis. SP,
spermidine; GAS5, growth arrest-specific transcript 5; PAECs,
pulmonary artery endothelial cells. |
GAS5 promotes SP-induced autophagy in
PAECs
To investigate the effect of GAS5 on SP-induced
autophagy in PAECs, pcDNA3.1-GAS5 and siRNA-GAS5 were transfected
into PAECs to overexpress and knockdown the abundance of GAS5,
respectively. The transfection efficiency was evaluated by RT-qPCR
(Fig. 2A and B), and the most
effective pcDNA3.1 plasmid and siRNA were used for subsequent
experiments. Following overexpressing GAS5 in SP-treated PAECs, the
stimulatory effect of SP on autophagy in PAECs was enhanced by
increased expression of GAS5 (Fig.
2C-E). By contrast, the overexpression of GAS5 was reversed by
the downregulation of GAS5 through transfecting with the siRNA
(Fig. 2E). Similarly, the effect of
GAS5 on autophagy was also observed when PAECs were not given the
SP treatment (Fig. 2F). In
conclusion, GAS5 may function as an autophagy enhancer in
PAECs.
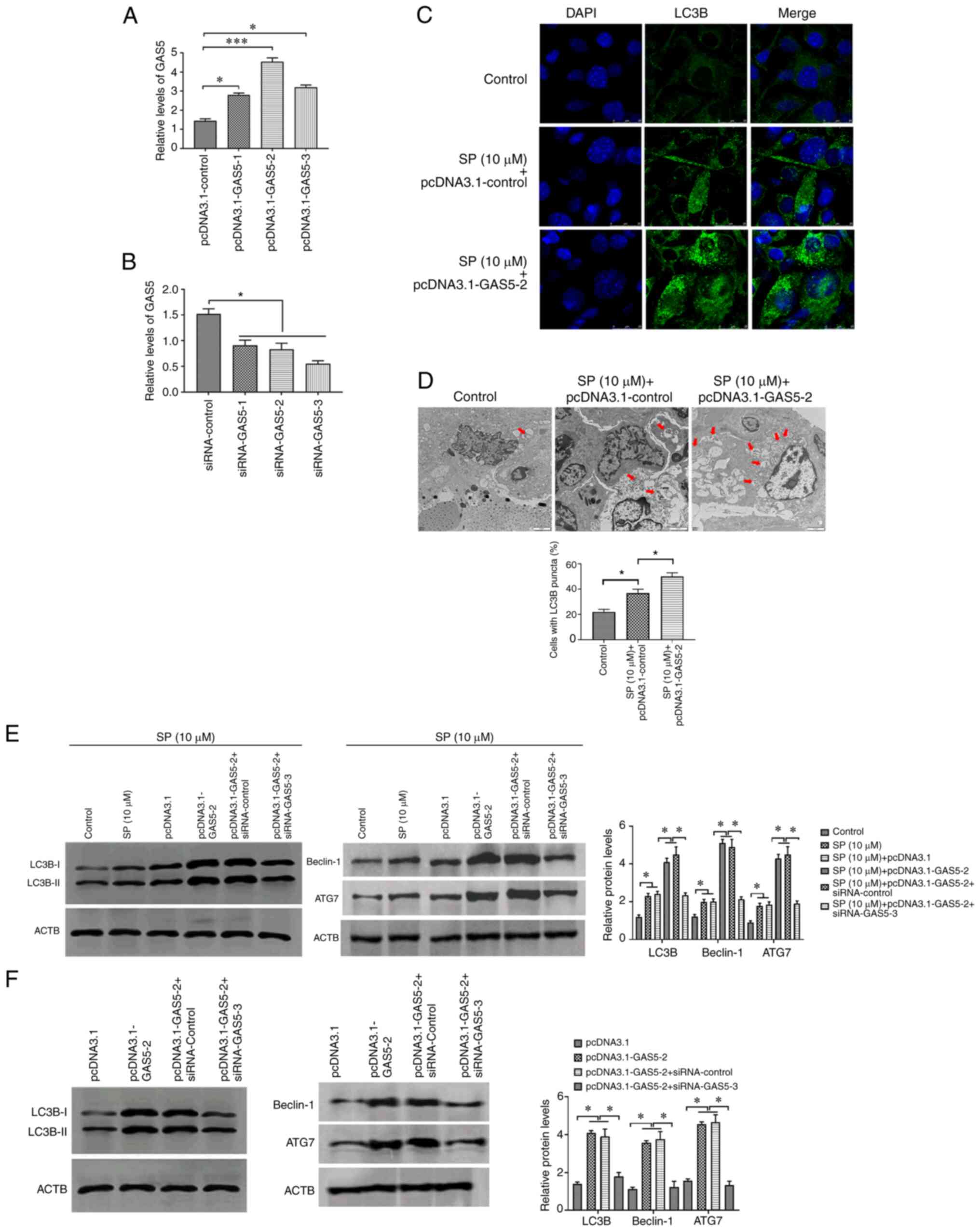 | Figure 2.GAS5 promotes SP-induced autophagy in
PAECs. (A) Transfection efficiency of pcDNA3.1-GAS5 in PAECs. (B)
Transfection efficiency of siRNA-GAS5 in PAECs. (C) The abundance
of LC3B in PAECs treated with the combination of SP (10 µM) and
pcDNA3.1-GAS5, as assessed by immunofluorescence assays (scale bar,
25 µm; magnification, ×20). (D) Autophagic vacuoles in the cellular
cytoplasm of PAECs treated with the combination of SP (10 µM) and
pcDNA3.1-GAS5, as evaluated by transmission electron microscopy
(scale bar, 2 µm; magnification, ×10,000). Red arrows indicate the
autophagic vacuoles. (E) Protein expressions of LC3B, Beclin-1 and
ATG7 in SP (10 µM)-treated PAECs transfected with pcDNA3.1-GAS5 and
the combination of pcDNA3.1-GAS5 and siRNA-GAS5, as assessed by
Western blotting. (F) Abundance of LC3B in PAECs transfected with
pcDNA3.1-GAS5 and the combination of pcDNA3.1-GAS5 and siRNA-GAS5,
as assessed by Western blotting. *P<0.05. There were at least
three replicates in each group available for analysis. GAS5, growth
arrest-specific transcript 5; SP, spermidine; PAECs, pulmonary
artery endothelial cells; siRNA, small interfering RNA. |
GAS5 promotes SP-induced autophagy in
PAECs via targeting miRNA-31-5p
To further determine the signaling pathway
underlying the effect of GAS5 in PAECs, bioinformatic analysis was
used to predict the potential miRNAs that interact with GAS5. The
results demonstrated that the 3′-UTR region of GAS5 contained a
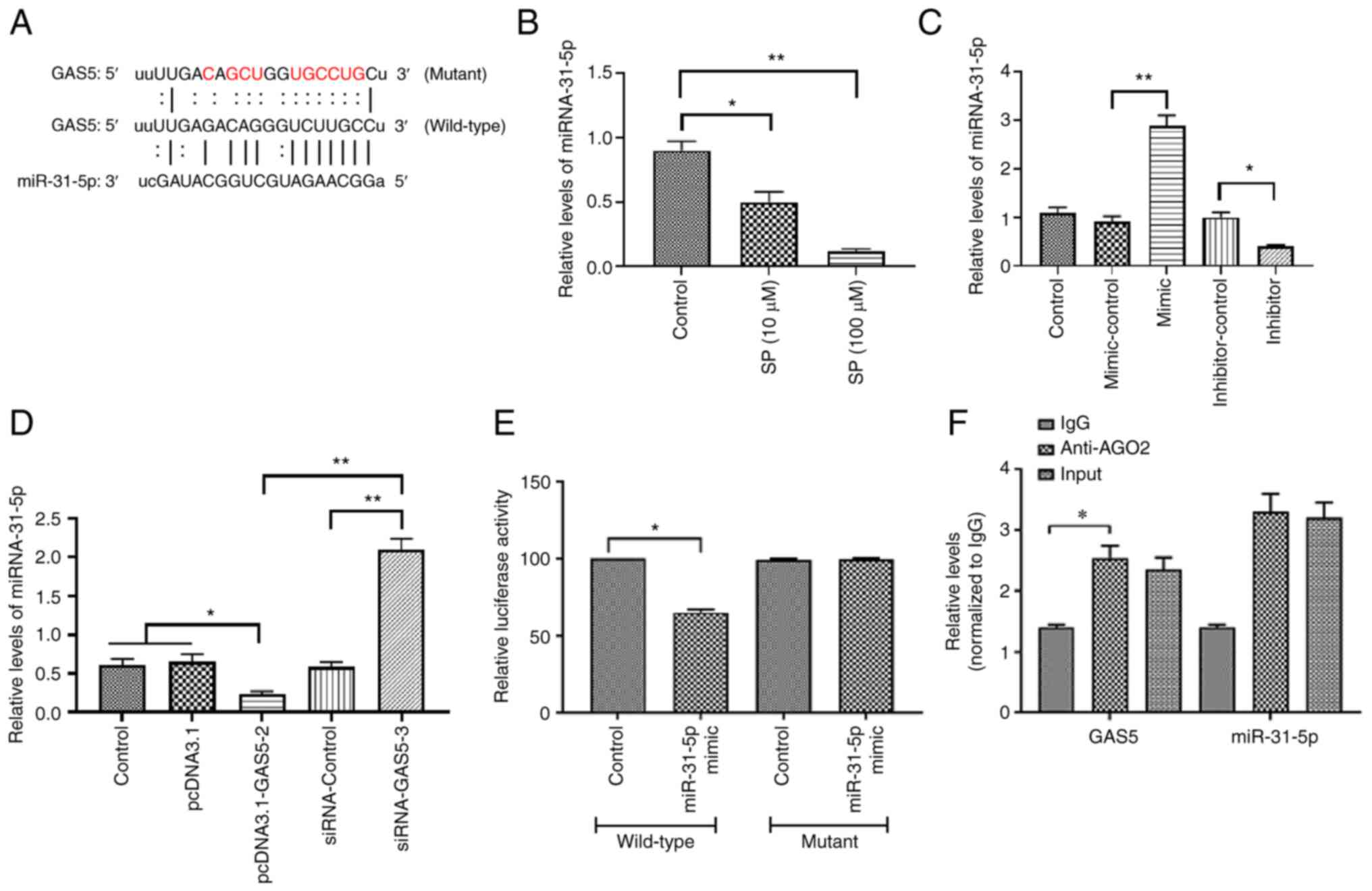
putative binding site of miRNA-31-5p (Fig. 3A). The expression of miRNA-31-5p in
SP-treated PAECs was measured and the results revealed that the
level of miRNA-31-5p was decreased by the SP treatment (Fig. 3B), suggesting that miRNA-31-5p may
be associated with the effect of SP on PAECs. To determine the
function of miRNA-31-5p in PAECs, miRNA-31-5p mimic was used to
increase the expression of miRNA-31-5p while the inhibitor was
applied to decrease the expression of miRNA-31-5p. The transfection
efficiency was evaluated by RT-qPCR (Fig. 3C). The overexpression of GAS5
decreased the miRNA-31-5p expression and the knockdown of GAS5
increased the miRNA-31-5p expression (Fig. 3D), suggesting a possible negative
association between GAS5 and miRNA-31-5p. The luciferase reporter
assays demonstrated that the luciferase activity of PAECs that were
co-transfected with the wild-type GAS5 plasmid and miRNA-31-5p
mimic decreased, while the co-transfection of the mutant GAS5
plasmid was not affected (Fig. 3E),
suggesting a physical interaction between GAS5 and miRNA-31-5p.
This interactive association was also verified by RIP assays, in
which GAS5 and miRNA-31-5p were enriched in AGO2 immunoprecipitates
(Fig. 3F). Taken together, these
results suggested that miRNA-31-5p may participate in the
regulation of SP-induced autophagy in PAECs.
MiRNA-31-5p suppresses SP-induced
autophagy in PAECs
To investigate the function of miRNA-31-5p in
autophagy in PAECs, miRNA-31-5p mimic and inhibitor were applied to
SP-treated PAECs, respectively. The results demonstrated that the
miRNA-31-5p mimic may reverse the effect of SP on autophagy in
PAECs (Fig. 4A-C). By contrast,
miRNA-31-5p inhibitor was found to enhance the autophagic level of
SP-treated PAECs (Fig. 4D-F).
Therefore, the miRNA-31-5p may act as a target gene of GAS5 to
regulate SP-induced autophagy in PAECs.
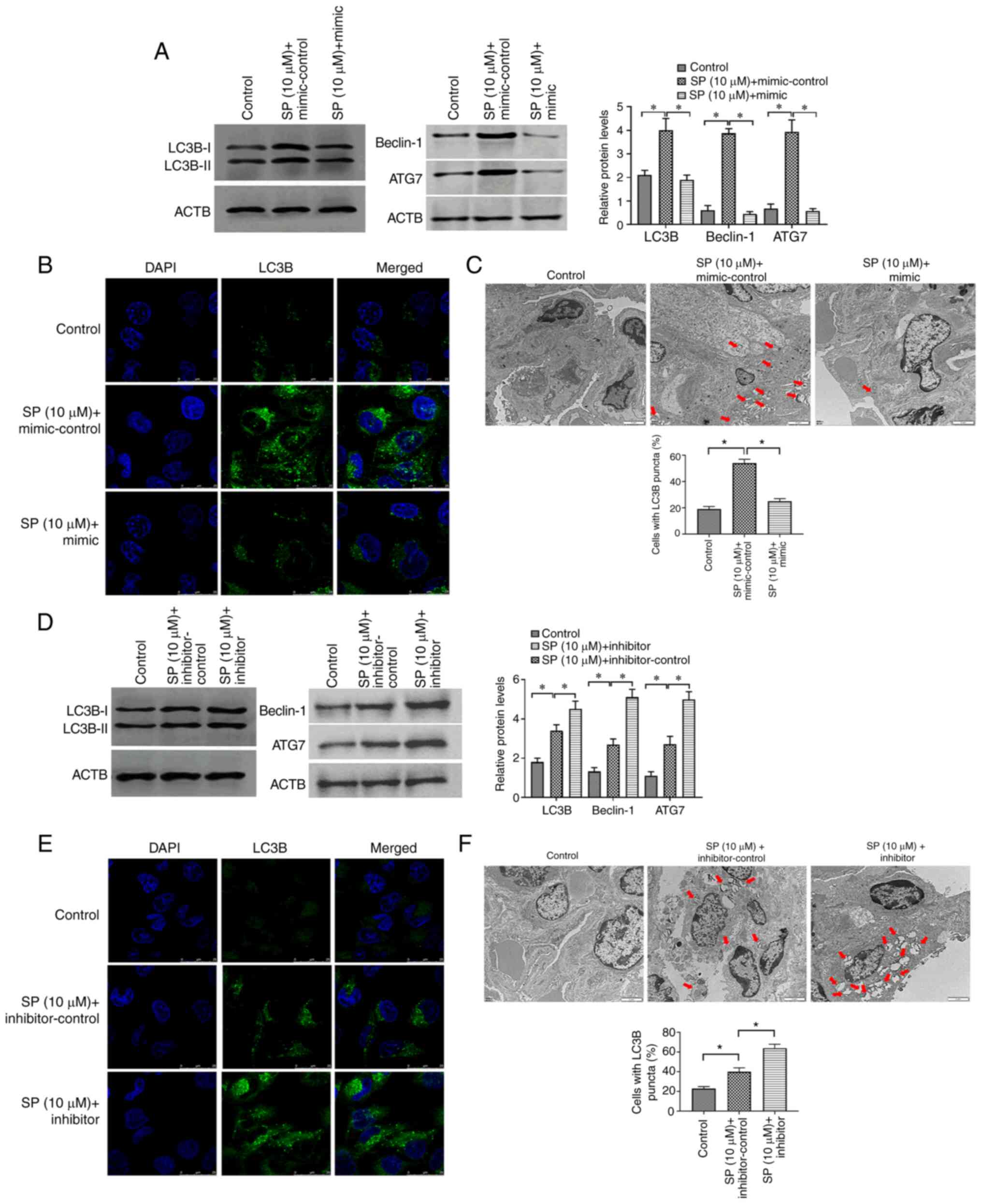 | Figure 4.MiRNA-31-5p inhibits SP-induced
autophagy in PAECs. (A) Abundance of LC3B in PAECs treated the
combination of SP (10 µM) and miRNA-31-5p mimic, as assessed by
Western blotting. (B) Protein expression of LC3B, Beclin-1 and ATG7
in PAECs treated with the combination of SP (10 µM) and miRNA-31-5p
mimic, as assessed by immunofluorescence assays (scale bar, 25 µm;
magnification, ×20). (C) Autophagic vacuoles in the cellular
cytoplasm of PAECs treated the combination of SP (10 µM) and
miRNA-31-5p mimic, as evaluated by transmission electron microscopy
(scale bar, 2 µm; magnification, ×10,000). Red arrows indicate the
autophagic vacuoles. (D) Protein expression of LC3B, Beclin-1 and
ATG7 in PAECs treated with the combination of SP (10 µM) and
miRNA-31-5p inhibitor, as assessed by Western blotting. (E) The
abundance of LC3B in PAECs treated with the combination of SP (10
µM) and miRNA-31-5p inhibitor, as assessed by immunofluorescence
assays (scale bar, 25 µm; magnification, ×20). (F) Autophagic
vacuoles in the cellular cytoplasm of PAECs treated with the
combination of SP (10 µM) and miRNA-31-5p inhibitor, as evaluated
by transmission electron microscopy (scale bar, 2 µm;
magnification, ×10,000). Red arrows indicate the autophagic
vacuoles. *P<0.05. There were at least three replicates in each
group available for analysis. SP, spermidine; PAECs, pulmonary
artery endothelial cells. |
MiRNA-31-5p inhibits SP-induced
autophagy in PAECs through targeting NAT8L
Based on the sponge role of miRNAs for mRNAs
(39), bioinformatic analysis was
used to further determine the possible target gene of miRNA-31-5p
associated with SP-induced autophagy. The results demonstrated that
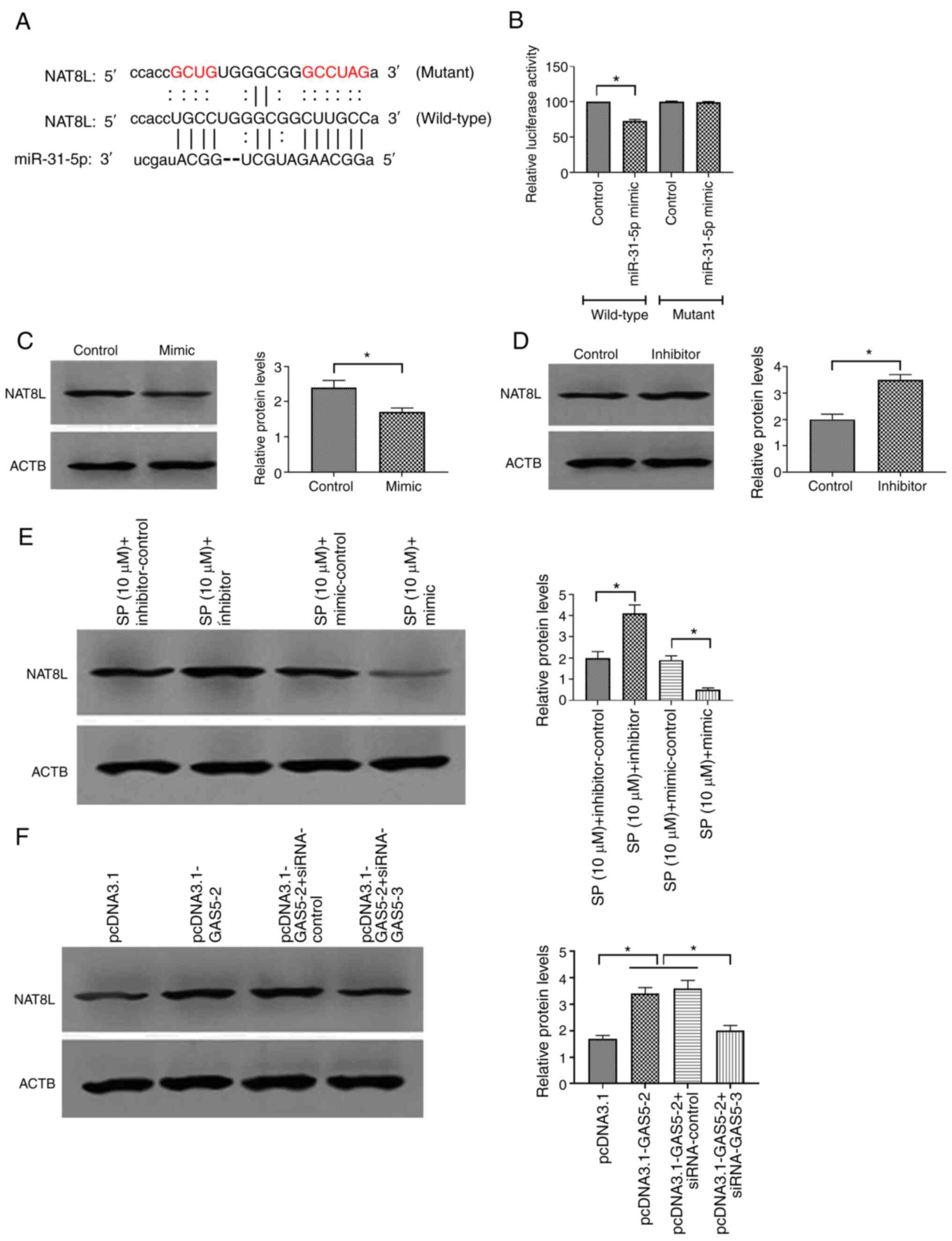
there was a binding site of miRNA-31-5p in the 3′-UTR sequence of
NAT8L (Fig. 5A). Luciferase
reporter assays demonstrated that PAECs co-transfected with the
wild-type NAT8L plasmid and miRNA-31-5p mimic decreased the
luciferase activity compared with those transfected with the mutant
NAT8L plasmid and miRNA-31-5p mimic (Fig. 5B), suggesting a physical interaction
between miRNA-31-5p and NAT8L. Functional studies revealed that
miRNA-31-5p mimic decreased and the inhibitor increased the protein
level of NAT8L (Fig. 5C and D,
respectively). Furthermore, similar effects of miRNA-31-5p mimic
and inhibitor on NAT8L were found in SP-treated PAECs, which
suggested that NAT8L may be involved in the regulation of
SP-induced autophagy in PAECs (Fig.
5E). Furthermore, the overexpression of GAS5 increased and the
knockdown of GAS5 decreased the protein expression of NAT8L in
PAECs (Fig. 5F). Therefore, GAS5
may modulate autophagy of PAECs through the miRNA-31-5p/NAT8L
axis.
GAS5/miRNA-31-5p/NAT8L axis regulates
SP-induced autophagy in PAECs
To verify the regulatory role of the
GAS5/miRNA-31-5p/NAT8L axis in SP-induced autophagy, siRNA-NAT8L
was used to downregulate the expression of NAT8L and the
transfection efficiency was confirmed by Western blotting (Fig. 6A). Using Western blotting,
immunofluorescence assays and TEM, SP-treated PAECs decreased
protein expression of NAT8L, which was associated with inhibited
autophagic levels and was reversed by the overexpression of GAS5
through transfecting with pcDNA3.1-GAS5 (Fig. 6B-D, respectively). Taken together,
the results demonstrated that GAS5 may promote SP-induced autophagy
in PAECs through the miRNA-31-5p/NAT8L signaling axis.
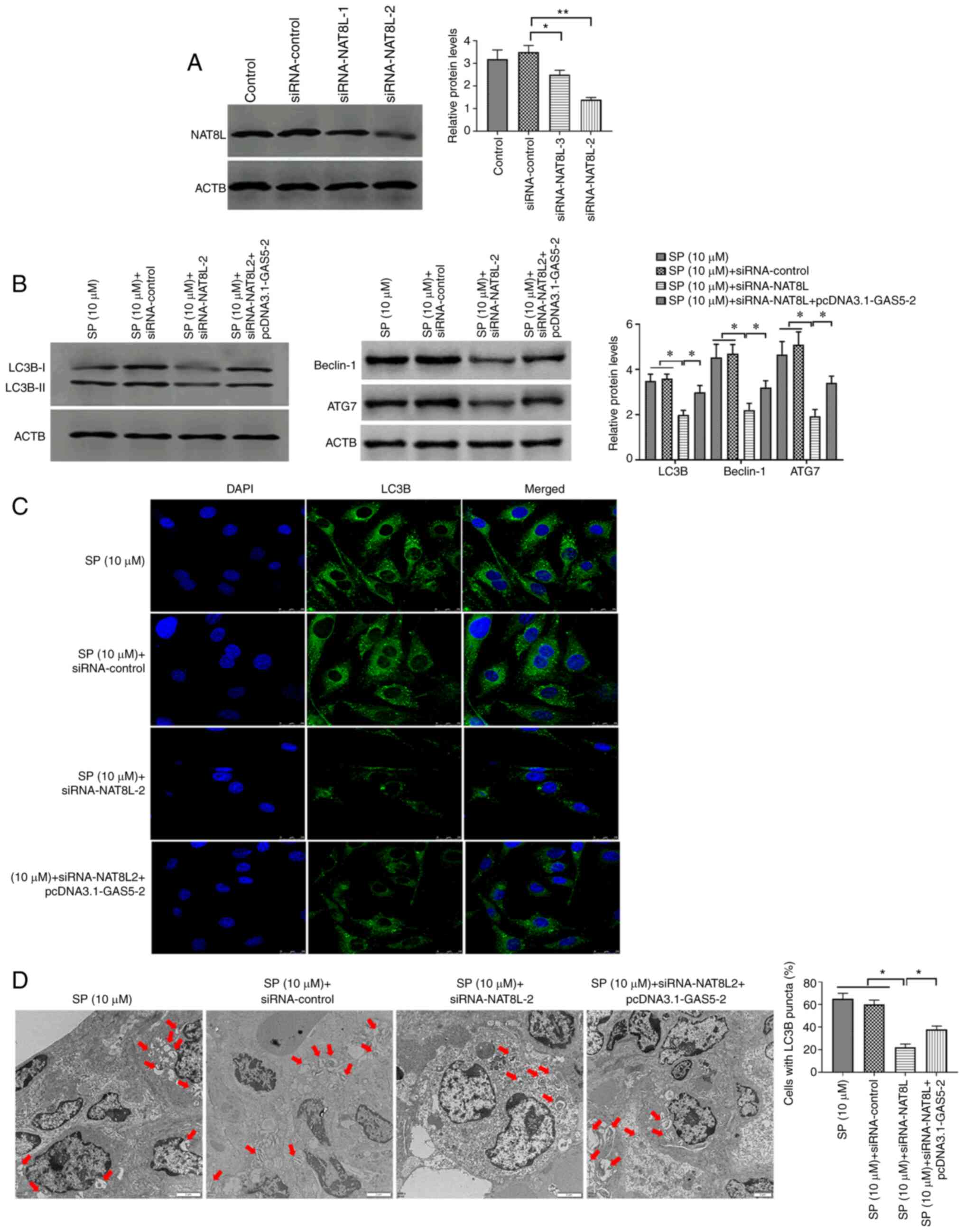 | Figure 6.NAT8L promotes SP-induced autophagy
in PAECs. (A) Transfection efficiency of siRNA-NAT8L in PAECs. (B)
Protein expression of LC3B, Beclin-1 and ATG7 in PAECs transfected
with siRNA-NAT8L and the combination of siRNA-NAT8L and
pcDNA3.1-GAS5, as assessed by Western blotting. (C) The abundance
of LC3B in PAECs, as assessed by immunofluorescence assays (scale
bar, 25 µm; magnification, ×20). (D) Autophagic vacuoles in the
cellular cytoplasm of PAECs, as evaluated by transmission electron
microscopy (scale bar, 2 µm; magnification, ×10,000). Red arrows
indicate the autophagic vacuoles. *P<0.05; **P<0.01. There
were at least three replicates in each group available for
analysis. SP, spermidine; PAECs, pulmonary artery endothelial
cells; siRNA, small interfering RNA. |
GAS5/miRNA-31-5p/NAT8L axis regulates
SP-induced autophagy in PAECs in vivo
To confirm the results from the in vitro
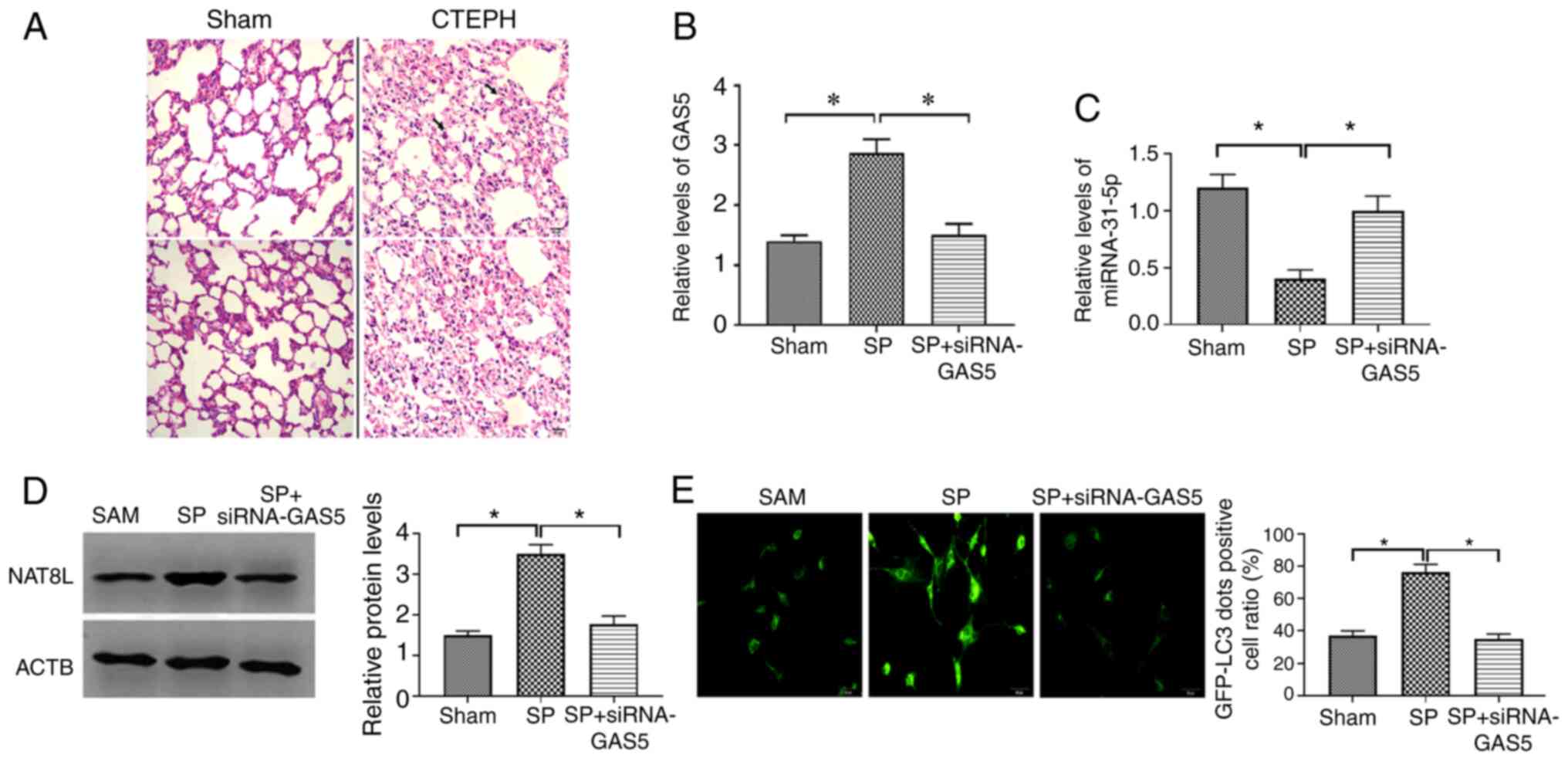
studies, the CTEPH rat model was created through the administration
of autologous blood clots (9) and
confirmed by assessment of pulmonary arterial pressure (Table III). The pathological feature of
the CTEPH rat model was investigated by H&E staining and the
results demonstrated that CTEPH rats had more alveolar exudation
and hemorrhages (Fig. 7A),
suggesting that the CTEPH rat model was successfully established.
The SP and the combination of SP and siRNA-GAS5 was delivered to
CTEPH rats, and the expression of GAS5, miRNA-31-5p and NAT8L was
assessed. The results demonstrated that in PAECs of CTEPH rats, SP
treatment enhanced the expression of GAS5 and NAT8L, as well as
inhibiting the level of miRNA-31-5p, and that this effect of SP
treatment was reversed by the downregulation of GAS5 via siRNA-GAS5
(Fig. 7B-D). To assess the
autophagic levels in SP and SP+siRNA-GAS5-treated rats, western
blotting, immunofluorescence assays and TEM were applied. The
results demonstrated that SP treatment enhanced autophagic levels
in PAECs of rats with CTEPH, while such roles were attenuated by
the downregulation of GAS5 (Fig.
8A-C). Additionally, similar observations were demonstrated by
using two other assays to evaluate the autophagic level, GFP-LC3
adenovirus and tfLC3 (Figs. 7E and
8D). Taken together, the results
demonstrated that GAS5 may regulate SP-induced autophagy in PAECs
through modulating the miRNA-31-5p/NAT8L axis (Fig. 8E).
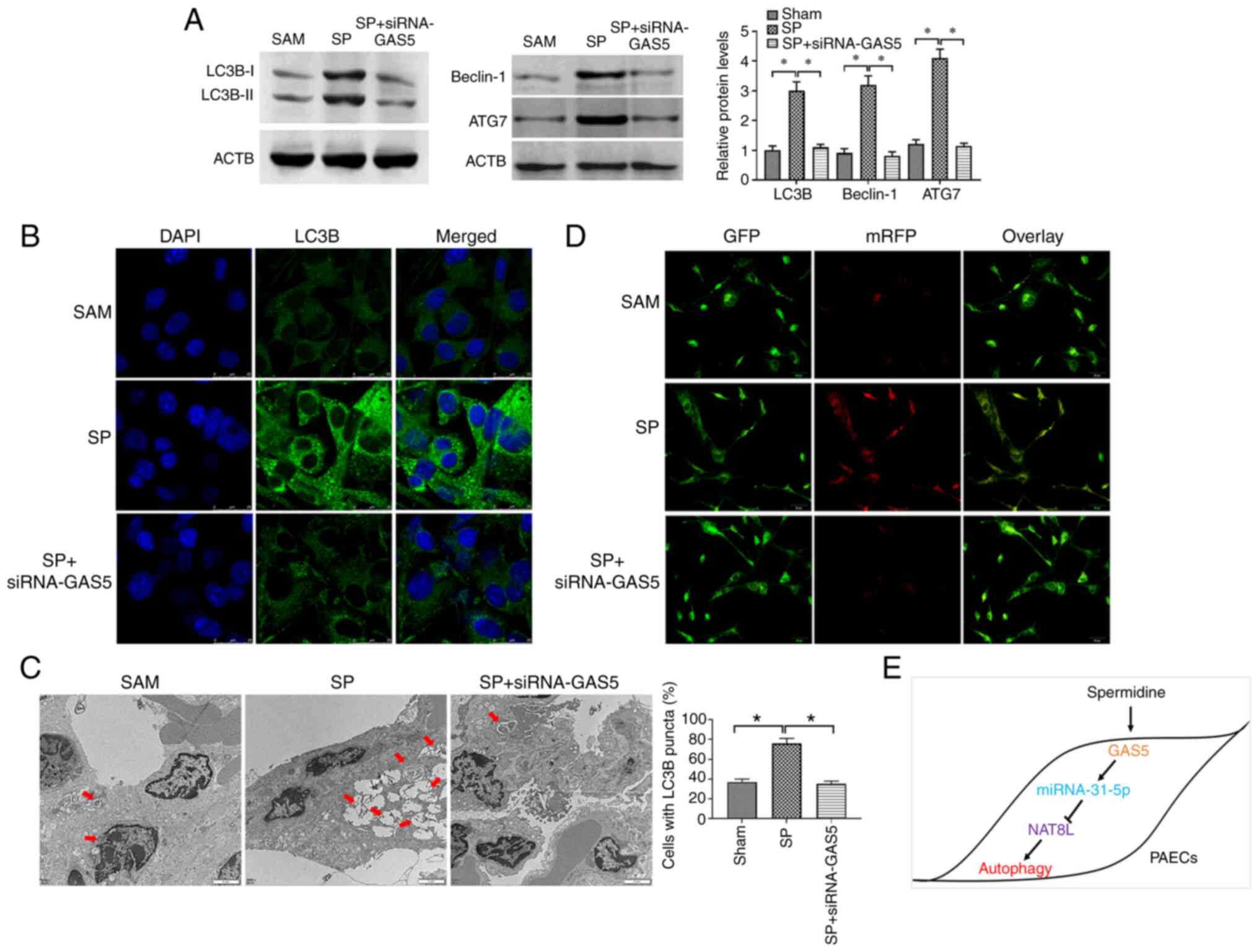 | Figure 8.NAT8L regulates SP-induced autophagy
in PAECs in vivo. (A) Protein expression of LC3B, Beclin-1 and ATG7
in PAECs of CTEPH rats treated with SP and the combination of SP
and siRNA-GAS5. (B) The abundance of LC3B in PAECs of rats with
CTEPH treated with SP and the combination of SP and siRNA-GAS5, as
assessed by immunofluorescence assays (scale bar: 25 µm;
magnification, ×20). (C) Autophagic vacuoles in the cellular
cytoplasm of PAECs of CTEPH rats treated with SP and the
combination of SP and siRNA-GAS5, as evaluated by transmission
electron microscopy (scale bar, 2 µm; magnification, ×10,000). Red
arrows indicate the autophagic vacuoles. (D) GFP signals in PAECs
as assessed by tfLC3 assays. (Scale bar, 50 µm; magnification,
×20). (E) Schematic of GAS5 regulating PAECs autophagy induced by
SP. *P<0.05. There were at least three replicates in each group
available for analysis. SP, spermidine; PAECs, pulmonary artery
endothelial cells; siRNA, small interfering RNA; CTEPH, chronic
thromboembolic pulmonary hypertension. |
| Table III.Pulmonary arterial pressure in the
rat CTEPH model. |
Table III.
Pulmonary arterial pressure in the
rat CTEPH model.
| Parameter | Sham | CTEPH | P-value |
|---|
| mPAP, mmHg | 15.5±2.1 | 33.6±3.1 | 0.021 |
| dPAP, mmHg | 12.6±1.3 | 24.9±3.7 | 0.038 |
| sPAP, mmHg | 22.1±1.2 | 46.3±4.2 | 0.020 |
Discussion
Pulmonary embolism (PE) is characterized by the
obstruction of the pulmonary artery or its branches by embolus, and
the most common PE is pulmonary thromboembolism (PTE) (40). In addition, CTEPH is classified as a
late sequela of PTE (41). At
present, CTEPH remains underdiagnosed and undertreated; therefore,
investigating the pathology of CTEPH has drawn a great amount of
attention in basic and clinical research fields. It has been
demonstrated that there is a tight anatomical correlation between
the thrombus and pulmonary artery endothelium (4), suggesting that there is an essential
involvement of PAECs in CTEPH. Accordingly, PAEC dysfunction,
including hyperproliferation and reduced-apoptosis, is known to
contribute toward the pathogenic vascular remodeling seen in CTEPH
(37). However, very few studies
report the involvement of autophagy, a complicated intracellular
degradation process, in the development and progression of CTEPH.
Therefore, investigating the effect of autophagy on PAECs
originated from CTEPH was a primary objective of the present
study.
In the current study, endogenous SP promoted
autophagy of PAECs of CTEPH in a dose-dependent manner. In the past
decade, studied have revealed that SP, a natural polyamine, is an
autophagy enhancer and promotes immune cell longevity (13), anti-aging (12), cardioprotective effects (42) and stress resistance (43), in which autophagy is required for
such health-facilitating roles. Furthermore, autophagy is tightly
associated with the degradation of impaired organelles,
dysfunctional protein aggregates and the clearance of cytoplasmic
materials that may accumulate during aging, thereby ensuring cell
homeostasis and proteostasis (44).
It has been reported that the mechanism underlying the stimulatory
effect of SP on autophagy is associated with the upregulation of
autophagy-relevant proteins via histone H3 hypoacetylation
(13) and de-acetylation of histone
H3 (45). Currently, the role of SP
on PAECs is unknown. Therefore, the promoting effect of SP on PAECs
reported in the present study extends the understanding of SP
function and provides a potential agent for CTEPH treatment.
To investigate the mechanism of SP action on PAECs,
lncRNAs, a group of non-protein-coding RNAs serving essential roles
in gene expression at transcriptional, post-transcriptional and
epigenetic level (46) were
studied. Based on previous studies, GAS5 functions as an antitumor
factor in multiple cancer types through modulating autophagy. Gu
et al (21) reported that
GAS5 exerts key roles in regulating autophagy in breast cancer via
the miRNA-23a/autophagy related 3 (Atg3) pathway. Additionally,
GAS5 promotes the cisplatin sensitivity of tumor cells via
suppressing autophagy in NSCLC (22). Furthermore, GAS5 is found to blunt
cisplatin resistance through inhibiting autophagy via the mTOR
signaling pathway in glioma cells (47). Since PAECs originating from CTEPH
display certain common characteristics with tumor cells, including
enhanced proliferation and inhibited apoptosis (37,38),
GAS5 may also serve important roles in the regulation of autophagy
in PAECs. Therefore, in the present study, the upregulation of GAS5
was observed in SP-induced autophagy in PAECs of CTEPH.
Additionally, the downregulation of GAS5 through siRNA-GAS5
reversed the effects of SP on autophagy in PAECs. Taken together,
these results indicated that GAS5 may be a key regulator in
SP-induced autophagy in PAECs.
As one of the primary roles, lncRNAs functions as
miRNA sponges to inhibit the regulatory effect of miRNAs on target
genes (48). This lncRNA-miRNA
interaction also serves important roles in various biological and
pathological processes (49).
Therefore, by applying bioinformatics and functional studies, SP
treatment decreased the expression of miRNA-31-5p in PAECs, and
GAS5 modulated the autophagic process in PAECs through sponging
miRNA-31-5p. In addition, miRNA-31-5p has been reported to
participate in a variety of diseases; for example, the upregulation
of miRNA-31-5p functions as a tumor suppressor to inhibit cell
migration, proliferation and invasion through the Sp1 transcription
factor in hepatocellular carcinoma cells (50). Furthermore, miRNA-31-5p caused
endothelial dysfunction, vascular remodeling and hypertension via
inhibiting post-transcription of endothelial nitric-oxide synthase
mRNA (51). To date, the role of
miRNA-31-5p in PAECs has barely been reported. Therefore,
miRNA-31-5p as an essential regulator in autophagy in PAECs may
provide novel insight into the function of miRNA-31-5p.
As a class of short non-coding RNAs, miRNAs are
associated with the regulation of gene expression at the
post-transcriptional level through two main mechanisms: mRNA
degradation and translational repression (39). To further investigate the mechanism
underlying the effect of GAS5 on autophagy in PAECs, bioinformatics
and functional studies were performed to investigate the downstream
target gene of miRNA-31-5p. The results demonstrated that NAT8L was
a direct target gene of miRNA-31-5p and that NAT8L mediated effects
of the GAS5-miRNA-31-5p signaling pathway in PAEC autophagy.
Furthermore, NAT8L is an enzyme associated with synthesizing NAA
that acts as a molecular transporter of acetate in lipid synthesis
in the brain (52). Regarding the
regulation of autophagy, NAT8L is a downstream regulator in lncRNA
TGFB2-OT1-associated autophagy and inflammation of vascular
endothelial cells through modulating mitochondrial function
(53). Additionally, the
overexpression of NAT8L induces autophagy as a compensatory pattern
to modulate energy homeostasis in brown adipose tissue (54). Combined with the results of the
present study, the results of this previous study suggested that
NAT8L-associated regulatory pathway may be a promising target for
modulating autophagy.
In conclusion, these results demonstrated that SP
may promote autophagy in PAECs derived from patients with CTEPH and
a rat model. Additionally, GAS5 may regulate SP-induced autophagy
through the miRNA-31-5p/NAT8L axis. The results of the present
study suggested that GAS5 may be regarded as a potential target for
therapeutic strategies of CTEPH. Furthermore, it is necessary to
highlight that the observations obtained from in vivo
studies should be further verified with a larger sample size due to
the limited sample size in animal experiments.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the first and corresponding authors on
reasonable request.
Authors' contributions
QW designed and performed the majority of
experiments, analyzed and interpreated the data, and was a major
contributor in writing the manuscript. XZ carried out the molecular
experiments, and analyzed and interpreated the data. YW designed
the experiments, and drafted and approved the final version of the
manuscript. YH designed experiments, was a major contributor in
writing the manuscript, and approved the final version of the
manuscript. QW and YH confirm the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All patients provided written informed consent prior
to inclusion in the present study. All experimental protocols were
performed in accordance with the Declaration of Helsinki. The
experimental protocols were approved by the Ethics Committee of
Cangzhou Central Hospital (Cangzhou, China). All experimental
procedures involving animals were approved by the Institutional
Animal Care and Use Committee of Cangzhou Central Hospital. All
animal experiments were performed according to the guidelines of
the local regulatory agencies and conformed to the Regulations for
the Management of Laboratory Animals published by the Ministry of
Science and Technology of the People's Republic of China.
Patient consent to participate
All patients provided written informed consent for
publication.
Competing interests
The authors declare that they have no competining
interests.
References
|
1
|
Pengo V, Lensing AW, Prins MH, Marchiori
A, Davidson BL, Tiozzo F, Albanese P, Biasiolo A, Pegoraro C,
Iliceto S, et al: Incidence of chronic thromboembolic pulmonary
hypertension after pulmonary embolism. N Engl J Med. 350:2257–2264.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Becattini C, Agnelli G, Pesavento R,
Silingardi M, Poggio R, Taliani MR and Ageno W: Incidence of
chronic thromboembolic pulmonary hypertension after a first episode
of pulmonary embolism. Chest. 130:172–175. 2006. View Article : Google Scholar
|
|
3
|
Sakao S and Tatsumi K: Crosstalk between
endothelial cell and thrombus in chronic thromboembolic pulmonary
hypertension: Perspective. Histol Histopathol. 28:185–193.
2013.
|
|
4
|
Lang IM, Pesavento R, Bonderman D and Yuan
JX: Risk factors and basic mechanisms of chronic thromboembolic
pulmonary hypertension: A current understanding. Eur Respir J.
41:462–468. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jin Y and Choi AM: Cross talk between
autophagy and apoptosis in pulmonary hypertension. Pulm Circ.
2:407–414. 2012. View Article : Google Scholar
|
|
6
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar
|
|
7
|
Lee SJ, Smith A, Guo L, Alastalo TP, Li M,
Sawada H, Liu X, Chen ZH, Ifedigbo E, Jin Y, et al: Autophagic
protein LC3B confers resistance against hypoxia-induced pulmonary
hypertension. Am J Respir Crit Care Med. 183:649–658. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lahm T, Albrecht M, Fisher AJ, Selej M,
Patel NG, Brown JA, Justice MJ, Brown MB, Van Demark M, Trulock KM,
et al: 17β-Estradiol attenuates hypoxic pulmonary hypertension via
estrogen receptor-mediated effects. Am J Respir Crit Care Med.
185:965–980. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Long L, Yang X, Southwood M, Lu J,
Marciniak SJ, Dunmore BJ and Morrell NW: Chloroquine prevents
progression of experimental pulmonary hypertension via inhibition
of autophagy and lysosomal bone morphogenetic protein type II
receptor degradation. Circ Res. 112:1159–1170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pegg AE: Mammalian polyamine metabolism
and function. IUBMB Life. 61:880–894. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Madeo F, Eisenberg T, Pietrocola F and
Kroemer G: Spermidine in health and disease. Science.
359:eaan27882018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
LaRocca TJ, Gioscia-Ryan RA, Hearon CM Jr
and Seals DR: The autophagy enhancer spermidine reverses arterial
aging. Mech Ageing Dev. 134:314–320. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Eisenberg T, Knauer H, Schauer A, Büttner
S, Ruckenstuhl C, Carmona-Gutierrez D, Ring J, Schroeder S, Magnes
C, Antonacci L, et al: Induction of autophagy by spermidine
promotes longevity. Nat Cell Biol. 11:1305–1314. 2009. View Article : Google Scholar
|
|
14
|
LaRocca TJ, Henson GD, Thorburn A, Sindler
AL, Pierce GL and Seals DR: Translational evidence that impaired
autophagy contributes to arterial ageing. J Physiol. 590:3305–3316.
2012. View Article : Google Scholar
|
|
15
|
Wilusz JE, Sunwoo H and Spector DL: Long
noncoding RNAs: Functional surprises from the RNA world. Genes Dev.
23:1494–1504. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar
|
|
17
|
Xue D, Zhou C, Lu H, Xu R, Xu X and He X:
LncRNA GAS5 inhibits proliferation and progression of prostate
cancer by targeting miR-103 through AKT/mTOR signaling pathway.
Tumour Biol. 37:16187–16197. 2016. View Article : Google Scholar
|
|
18
|
Pickard M, Mourtada-Maarabouni M and
Williams G: Long non-coding RNA GAS5 regulates apoptosis in
prostate cancer cell lines. Biochim Biophys Acta. 1832:1613–1623.
2013. View Article : Google Scholar
|
|
19
|
Tao H, Zhang JG, Qin RH, Dai C, Shi P,
Yang JJ, Deng ZY and Shi KH: LncRNA GAS5 controls cardiac
fibroblast activation and fibrosis by targeting miR-21 via
PTEN/MMP-2 signaling pathway. Toxicology. 386:11–18. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ye K, Wang S, Zhang H, Han H, Ma B and Nan
W: Long noncoding RNA GAS5 suppresses cell growth and
epithelial-mesenchymal transition in osteosarcoma by regulating the
miR-221/ARHI pathway. J Cell Biochem. 118:4772–4781. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gu J, Wang Y, Wang X, Zhou D, Wang X, Zhou
M and He Z: Effect of the LncRNA GAS5-MiR-23a-ATG3 axis in
regulating autophagy in patients with breast cancer. Cell Physiol
Biochem. 48:194–207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang N, Yang GQ, Shao XM and Wei L: GAS5
modulated autophagy is a mechanism modulating cisplatin sensitivity
in NSCLC cells. Eur Rev Med Pharmacol Sci. 20:2271–2277. 2016.
|
|
23
|
Pulito C, Mori F, Sacconi A, Goeman F,
Ferraiuolo M, Pasanisi P, Campagnoli C, Berrino F, Fanciulli M,
Ford RJ, et al: Metformin-induced ablation of microRNA 21-5p
releases Sestrin-1 and CAB39L antitumoral activities. Cell Discov.
3:170222017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rasband W: ImageJ software. US National
Institutes of Health; Bethesda, Maryland, USA: 2011
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yin QF, Yang L, Zhang Y, Xiang JF, Wu YW,
Carmichael GG and Chen LL: Long noncoding RNAs with snoRNA ends.
Mol Cell. 48:219–230. 2012. View Article : Google Scholar
|
|
27
|
Volders PJ, Anckaert J, Verheggen K,
Nuytens J, Martens L, Mestdagh P and Vandesompele J: LNCipedia 5:
Towards a reference set of human long non-coding RNAs. Nucleic
Acids Res. 47:D135–D139. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao Z, Pan X, Yang Y, Huang Y and Shen HB:
The lncLocator: A subcellular localization predictor for long
non-coding RNAs based on a stacked ensemble classifier.
Bioinformatics. 34:2185–2194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:(Database Issue). D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar
|
|
31
|
Betel D, Koppal A, Agius P, Sander C and
Leslie C: Comprehensive modeling of microRNA targets predicts
functional non-conserved and non-canonical sites. Genome Biol.
11:R902010. View Article : Google Scholar
|
|
32
|
Deng C, Wu D, Yang M, Chen Y, Ding H,
Zhong Z, Lian N, Zhang Q, Wu S and Liu K: The role of tissue factor
and autophagy in pulmonary vascular remodeling in a rat model for
chronic thromboembolic pulmonary hypertension. Respir Res.
17:652016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang K, Liu CY, Zhou LY, Wang JX, Wang M,
Zhao B, Zhao WK, Xu SJ, Fan LH, Zhang XJ, et al: APF lncRNA
regulates autophagy and myocardial infarction by targeting
miR-188-3p. Nat Commun. 6:67792015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao Y and Klionsky DJ: Physiological
functions of Atg6/beclin 1: A unique autophagy-related protein.
Cell Res. 17:839–849. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Komatsu M, Tanida I, Ueno T, Ohsumi M,
Ohsumi Y and Kominami E: The C-terminal region of an Apg7p/Cvt2p is
required for homodimerization and is essential for its E1 activity
and E1-E2 complex formation. J Biol Chem. 276:9846–9854. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ichimura Y, Kirisako T, Takao T, Satomi Y,
Shimonishi Y, Ishihara N, Mizushima N, Tanida I, Kominami E, Ohsumi
M, et al: A ubiquitin-like system mediates protein lipidation.
Nature. 408:488–492. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mercier O, Arthur Ataam J, Langer NB,
Dorfmüller P, Lamrani L, Lecerf F, Decante B, Dartevelle P,
Eddahibi S and Fadel E: Abnormal pulmonary endothelial cells may
underlie the enigmatic pathogenesis of chronic thromboembolic
pulmonary hypertension. J Heart Lung Transplant. 36:305–314. 2017.
View Article : Google Scholar
|
|
38
|
Casero RA Jr and Marton LJ: Targeting
polyamine metabolism and function in cancer and other
hyperproliferative diseases. Nat Rev Drug Discov. 6:373–390. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cai Y, Yu X, Hu S and Yu J: A brief review
on the mechanisms of miRNA regulation. Genomics Proteomics
Bioinformatics. 7:147–154. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kucher N, Rossi E and Derosa M: Massive
pulmonary embolism. J Vasc Surg. 44:684–685. 2006. View Article : Google Scholar
|
|
41
|
Gerges C, Skoro-Sajer N and Lang IM: Right
ventricle in acute and chronic pulmonary embolism (2013 Grover
Conference series). Pulm Circ. 4:378–386. 2014. View Article : Google Scholar
|
|
42
|
Eisenberg T, Abdellatif M, Schroeder S,
Primessnig U, Stekovic S, Pendl T, Harger A, Schipke J, Zimmermann
A, Schmidt A, et al: Cardioprotection and lifespan extension by the
natural polyamine spermidine. Nat Med. 22:1428–1438. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Minois N, Carmona-Gutierrez D, Bauer MA,
Rockenfeller P, Eisenberg T, Brandhorst S, Sigrist SJ, Kroemer G
and Madeo F: Spermidine promotes stress resistance in Drosophila
melanogaster through autophagy-dependent and -independent pathways.
Cell Death Dis. 3:e4012012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mizushima N and Komatsu M: Autophagy:
Renovation of cells and tissues. Cell. 147:728–741. 2011.
View Article : Google Scholar
|
|
45
|
Morselli E, Mariño G, Bennetzen MV,
Eisenberg T, Megalou E, Schroeder S, Cabrera S, Bénit P, Rustin P,
Criollo A, et al: Spermidine and resveratrol induce autophagy by
distinct pathways converging on the acetylproteome. J Cell Biol.
192:615–629. 2011. View Article : Google Scholar
|
|
46
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar
|
|
47
|
Huo JF and Chen XB: Long noncoding RNA
growth arrest-specific 5 facilitates glioma cell sensitivity to
cisplatin by suppressing excessive autophagy in an mTOR-dependent
manner. J Cell Biochem. 120:6127–6136. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Paraskevopoulou MD and Hatzigeorgiou AG:
Analyzing miRNA-lncRNA interactions. Long Non-Coding RNAs. Methods
in Molecular Biology. 1402. Feng Y and Zhang L: Humana Press; New
York, NY: pp. 271–286. 2016, View Article : Google Scholar
|
|
49
|
Yoon J, Abdelmohsen K and Gorospe M:
Functional interactions among microRNAs and long noncoding RNAs.
Seminars in cell and developmental biology. Elsevier; pp. 9–14.
2014, View Article : Google Scholar
|
|
50
|
Hessam S, Sand M, Skrygan M, Gambichler T
and Bechara FG: Expression of miRNA-155, miRNA-223, miRNA-31,
miRNA-21, miRNA-125b, and miRNA-146a in the inflammatory pathway of
hidradenitis suppurativa. Inflammation. 40:464–472. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kim S, Lee KS, Choi S, Kim J, Lee DK, Park
M, Park W, Kim TH, Hwang JY, Won MH, et al: NF-κB-responsive
miRNA-31-5p elicits endothelial dysfunction associated with
preeclampsia via down-regulation of endothelial nitric-oxide
synthase. J Biol Chem. 293:18989–19000. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mehta V and Namboodiri M:
N-acetylaspartate as an acetyl source in the nervous system. Brain
Res Mol Brain Res. 31:151–157. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Huang S, Lu W, Ge D, Meng N, Li Y, Su L,
Zhang S, Zhang Y, Zhao B and Miao J: A new microRNA signal pathway
regulated by long noncoding RNA TGFB2-OT1 in autophagy and
inflammation of vascular endothelial cells. Autophagy.
11:2172–2183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Huber K, Hofer DC, Trefely S, Pelzmann HJ,
Madreiter-Sokolowski C, Duta-Mare M, Schlager S, Trausinger G,
Stryeck S, Graier WF, et al: N-acetylaspartate pathway is nutrient
responsive and coordinates lipid and energy metabolism in brown
adipocytes. Biochim Biophys Acta Mol Cell Res. 1866:337–348. 2019.
View Article : Google Scholar : PubMed/NCBI
|