Cathepsins are proteases involved in multiple
physiological roles ranging from ion channel activity, apoptosis,
autophagy, immune regulation and complement system activation. They
are divided into several subtypes-serine cathepsins, cysteine
cathepsins and aspartic cathepsins (1). Various pathologies that can be
attributed to dysregulation of cathepsins include pancreatitis,
acute and chronic kidney disease, arthritis, auto-inflammatory
diseases, stroke and IR injury.
Lysosomes are responsible for catabolism and
recycling of multiple macromolecules and are key degradative
compartments in the cell. There are two major proteins that mediate
lysosomal activity; lysosomal membrane proteins and lysosomal
hydrolases including the cathepsins (2). Their activity is facilitated by the
acidic lysosomal environment. In addition to intracellular
functions, cathepsins exert extracellular roles in proteolysis,
antigen activation, extracellular matrix remodeling and tumor
invasion (3,4).
In mammals, there are 3 classes of cathepsins based
on the amino acid each specific cathepsin breaks down including
serine cathepsins, cysteine cathepsins and aspartic cathepsins
(1). Of these subtypes, serine
and cysteine cathepsins are most abundant (7).
There are at least 11 cysteine cathepsin subtypes in
mouse and human cells (B, C, F, H, K, L, O, S, V, W and X)-all of
which have specific roles, but some share similar functions
creating a system with certain redundancy (3).
Cathepsin B is one of the most abundant cysteine
cathepsins within the lysosome. It also is the most stable protease
at physiological pH which translates to its excellent ability to
function outside the lysosome in the cytoplasm and in the
extracellular space (8).
Cathepsin B also plays a major role in apoptosis (8). Cellular injury leads to lysosomal
permeabilization or rupture leading to Cathepsin B release which in
turn causes degradation of Bid (BH3-interacting domain death
agonist, pro-apoptotic member of Bcl-2 protein family) resulting in
its activation and translocation into the mitochondria with
subsequent mitochondrial cytochrome c release and caspases 3 and 9
activation (9). Cathepsin B also
regulates inflammation via NLRP3 (NOD-leucine rich repeat and pyrin
containing protein 3) inflammasome activation (10). NLRP3 inflammasome is a critical
component of the innate immune system, has multiple activators,
such as, SiO2, MonoSodium Urate (MSU), calcium
pyrophosphate dehydrate crystals, nigerecin and ATP, upon
stimulation of these activators, NLRP3 is able to form a transient
complex with Cathepsin B which causes the inflammasome to recruit
and activate pro-inflammatory caspase 1 and cytokines IL-1β and
IL-18 (11). Moreover, the NLRP3
inflammasome complex modulates kidney inflammation due to renal IR
injury (12).
Cathepsin C cleaves protease zymogens to their
active form. Among the proteases cleaved by Cathepsin C are
elastase, proteinase 3, neutrophil serine protease 4, chymase,
tryptase, granzymes A and B (13). These proteases have been linked to
causing early primary lung graft dysfunction in transplant patients
(14).
Cathepsin L like Cathepsin B is abundant in the
lysosomes and can cleave Bid directly promoting cytochrome c
release to initiate apoptosis. It also shares similar molecular
structure to Cathepsin B, explaining redundancy in various
physiological functions. One of the more prominent roles for
Cathepsin L is its role in autophagy; specifically, degradation of
the autophagosome as Cathepsin L knockout mice have pathological
accumulation of autophagosomes (15).
Cathepsin W is mainly expressed in cytotoxic T
lymphocytes and Nature Killer (NK) cells and is upregulated via
IL-2 (22). There is some debate
as to whether Cathepsin W has an important functional role in
cytotoxicity, given mixed reports on this subject; some stating
that Cathepsin W is upregulated in Cytotoxic NK cells (23); whilst others report inhibition of
Cathepsin W showed no changes in cytotoxicity (24).
Cathepsin E is mainly found in dendritic cells,
microglia and macrophages and has a role in regulating
endosomal/lysosomal microenvironment and protein sorting in these
compartments (25). Multiple
studies also show that it can play a role in MHC-II mediated
antigen processing, which enhances the inflammatory response in the
reperfusion phase of IR by increased tumor necrosis factor α
(26,27). Intracellular functions of some
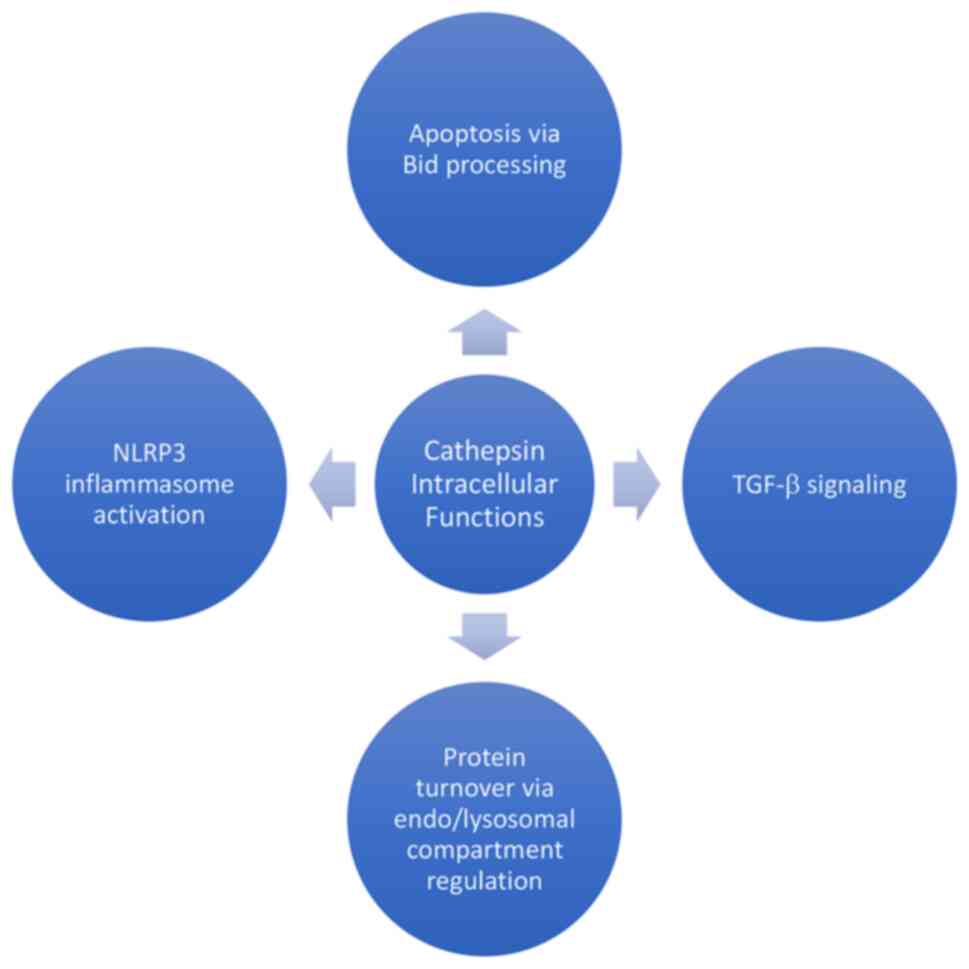
cathepsins are summarized in Fig.
1.
Cathepsin D plays a role in MHC-II (Major
histocompatibility complex class II) mediated antigen presentation
to CD4+ cells responsible for initiating
antigen-specific immune response, similar to Cathepsin E and B
(28). In addition, Cathepsin D
shares a redundant function alongside Cathepsin L in autophagy for
its role for initiation, autophagosome formation and fusion with
lysosomes this is evidenced due to Cathepsin D upregulation in
Cathepsin L knock out mice (10).
Similar to Cathepsin B; Cathepsin D has been implicated in
degradation of Bid, thus initiating apoptosis (29). Furthermore, Cathepsin D has been
implicated in initiating apoptosis via caspase-8 activation
(30); specifically in T cells it
activates Bax, releasing cytochrome c and apoptosis-inducing
factor; this process initiates apoptosis independent of Bid
(31).
Cathepsin A (also known as Lysosomal Protective
Protein) regulates blood pressure via regulation endothelin-1
(ET-1) a potent vasoconstrictor and forms part of the
elastin-binding complex which is responsible of the biogenesis of
elastin fibers (32).
Furthermore, upregulation of Cathepsin A can degrade peptide
hormones such as bradykinin and angiotensin II, both which promote
release of norepinephrine and enhance arrhythmias in myocardial IR
(33). Cathepsin A in cardiac
tissue shows a potential role in IR injury associated
pathologies.
The ischemic phase of a stroke not only can cause
irreversible damage, but after treatment the reperfusion phase is
capable of further enhancing neuronal cell death, brain edema even
hemorrhage (40). Multiple
studies have shown the role of cathepsins in brain IR, specifically
their function in inducing neuronal cell death. Cathepsins B, L and
D have been the major proteases studied after seeing major
increases in their levels in the hippocampus after induction of
experimental cerebral IR (41).
Cathepsin B has been heavily implicated in a potential role to
induce apoptosis via TNF-α and cytochrome c release (42,43). Several cathepsin inhibitors
attenuate cerebral IR injury including E64d (aloxistatin, cathepsin
B/L inhibitor), CA-074me (membrane permeable cathepsin B
inhibitor), Ginkgolide-B (Ginkgo biloba derivative, showed reduced
Cathepsin B/L activity), Cysteine Protease Inhibitor 1 (CP-1,
cathepsin B and L inhibitor), Probenecid (gout medication, showed
reduced cathepsin B activity), Sevoflurane (volatile anesthetic,
showed reduced cathepsin B activity), Propofol (intravenous
anesthetic, showed reduced cathepsin B/D activity) and Tomatidine
(green tomato extract, enhanced cathepsin B/D activity) (5,41,44–49). Furthermore; the specific mechanism
responsible for the release of cathepsins in the brain come from
lysosomal membrane permeabilization or lysosomal membrane rupture
(50). In terms of mechanisms of
cathepsins in cerebral IR injury, there is evidence that Heat shock
protein 70 (Hsp70) and Lysosome Associated Membrane Protein 1 and 2
(LAMP-1/2) are major players in maintaining lysosomal membrane
stability; therefore, preventing major cathepsin leakage which can
have the potential to attenuate cerebral IR injury in addition to
other organ systems IR injuries (51–53).
Myocardial ischemia is one of the leading causes of
death in the world, the earlier the ischemic period is treated
results in better prognosis; yet the reperfusion phase is
responsible for another sequela of stressors such as myocardial
stunning, inflammation, apoptosis, necrosis and microvascular
obstruction (54,55). The inflammatory damage that occurs
in the early reperfusion period is mediated by early activation of
neutrophils and mast cells, which their products serve as major
chemoattractant for other leukocytes (56,57). Cathepsin G has been shown to be a
modulator for neutrophil chemoattractant along with the ability to
cause morphological changes that disrupt focal adhesion and
intracellular contacts in cardiomyocytes (58–60). Inhibition of Cathepsin G with DCCI
(dual cathepsin G and chymase inhibitor) attenuated collagen
deposition within the myocardium, marked improvement in left
ventricle (LV) function, implicating that these could decrease
further progression of heart failure (61). Cathepsin S has also been linked
with LV remodeling post ischemia specifically as a potent protease
in degrading collagen and fibronectin (62,63). Furthermore, Cathepsin S inhibition
preserves LV function via TGF-β1 signalling, myofibroblast
trans-differentiation and extracellular matrix (ECM) synthesis
(63) and possible role in
apoptosis and inflammation via regulation of caspase 8, caspase-3
and TNF-α (64). Similarly,
Cathepsin A has shown a role in atrial fibrosis via potential in
ECM remodeling (65). In
addition, Cathepsin A inhibition in cardiac IR preserved greater LV
viable myocardium, along with reduced ECM remodeling which in turn
decreased the risk of developing arrhythmias, specifically atrial
arrhythmias (66). Moreover, both
Cathepsins D and L have been studied in conjunction for their
potential role in degradation of myofibrillar proteins,
specifically in IR injury via coronary artery bypass graft (CABG);
which demonstrated increased levels of both D and L occur in the
intralysosomal compartment during the ischemic period vs. increased
levels in the extralysosomal compartment during reperfusion phase;
indicating their potential for being chemoattractant in the
reperfusion period (67,68). Taken together, cathepsins seem to
share a system of roles in ECM remodeling specifically for cardiac
IR injury, furthermore linking all the cathepsins in a potential
redundant role.
Hepatic IR is the major cause for primary
non-function of liver grafts after liver transplantation,
hemorrhagic shock and liver resections (69). Moreover, Cathepsin B and its link
to the lysosomal pathway of apoptosis via TNF-α upregulation has
been found to play a large role in hepatic IR, where studies of
both pharmacological and gene knockout have shown to attenuate
apoptosis (70,71). Similarly, Cathepsin E knockout
mice in combination with TGR5 (G-protein coupled bile acid
receptor) deficiency have attenuated hepatic IR injury by
restraining macrophage migration and facilitating macrophage M2
polarization (72).
Taken together, cathepsins are proteases with
various roles ranging from apoptosis, immunity and metastasis.
Serine, cysteine and aspartic cathepsins share among them some
similar structural and physiological roles, yet also have distinct
differences. The role of cathepsins in IR injury has gained
attention biomedical research, establishing them as potential
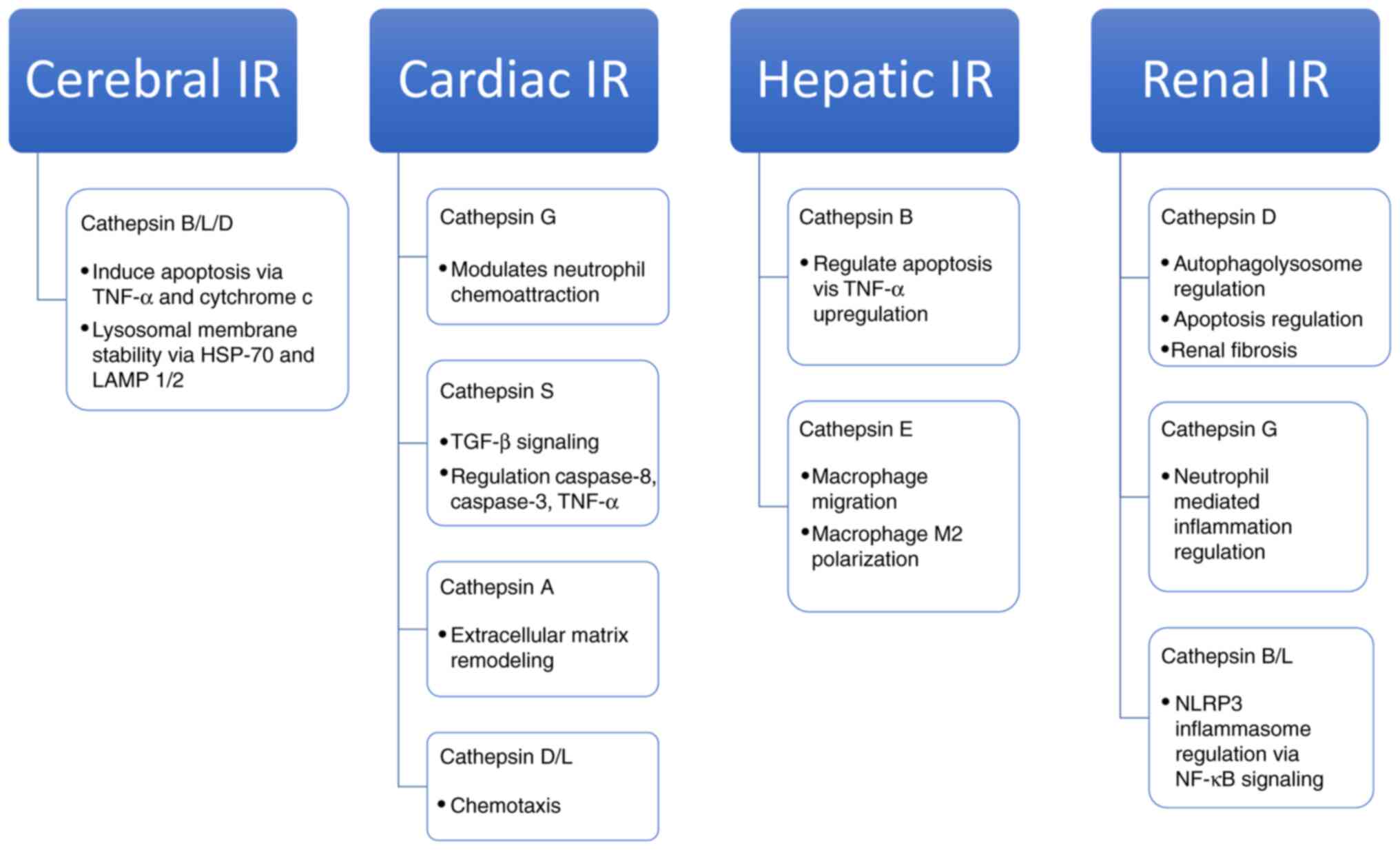
pharmacological targets. In summary, cerebral IR injury the
cathepsins most studied were B, L and D; for cardiac IR were
cathepsins G, S, A, D and L; hepatic IR cathepsins B and E; and
renal IR were cathepsins D, G, B and L. Summarized roles of
cathepsins in different organ systems are mentioned in Fig. 2. However, multiple cathepsins are
yet to be studied under different IR models, future research should
focus to test other cathepsin targets and understanding the
specific pathway involvement of cathepsins in IR injury across
multiple organ systems. Specifically, test cathepsins C, X and W
which have been associated with different mechanisms within the
immune system and inflammation, both components of IR injury.
Furthermore, there is a need to investigate cathepsins F, H, K, L,
O and V in the setting of inflammation or other components of the
immune system to see if there is potential to them being also
researched in the setting of IR injury.
Not applicable.
This work was supported by National Institute of Health Grant
(grant nos. DK115694, DK109544, DK129252 and T32GM8464-29).
Not applicable.
JH acquired, analyzed and compiled all cathepsin
articles pertaining to ischemia reperfusion injury and manuscript
drafting. HTL contributed to manuscript drafting, critical
revisions of intellectual content and approved final manuscript
version to be published. Both authors have read and approved the
final manuscript. Data sharing is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Patel S, Homaei A, El-Seedi HR and Akhtar
N: Cathepsins: Proteases that are vital for survival but can also
be fatal. Biomed Pharmacother. 105:526–532. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cocchiaro P, De Pasquale V, Della Morte R,
Tafuri S, Avallone L, Pizard A, Moles A and Pavone LM: The
multifaceted role of the lysosomal protease cathepsins in kidney
disease. Front Cell Dev Biol. 5:1142017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brix K, Dunkhorst A, Mayer K and Jordans
S: Cysteine cathepsins: Cellular roadmap to different functions.
Biochimie. 90:194–207. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vidak E, Javoršek U, Vizovišek M and Turk
B: Cysteine cathepsins and their extracellular roles: Shaping the
microenvironment. Cells. 8:2642019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xu Y, Wang J, Song X, Wei R, He F, Peng G
and Luo B: Protective mechanisms of CA074-me (other than
cathepsin-B inhibition) against programmed necrosis induced by
global cerebral ischemia/reperfusion injury in rats. Brain Res
Bull. 120:97–105. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stoka V, Turk V and Turk B: Lysosomal
cysteine cathepsins: Signaling pathways in apoptosis. Biol Chem.
388:555–560. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan GJ, Peng ZK, Lu JP and Tang FQ:
Cathepsins mediate tumor metastasis. World J Biol Chem. 4:91–101.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hook G, Jacobsen JS, Grabstein K, Kindy M
and Hook V: Cathepsin B is a new drug target for traumatic brain
injury therapeutics: Evidence for E64d as a promising lead drug
candidate. Front Neurol. 6:1782015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ben-Ari Z, Mor E, Azarov D, Sulkes J, Tor
R, Cheporko Y, Hochhauser E and Pappo O: Cathepsin B inactivation
attenuates the apoptotic injury induced by ischemia/reperfusion of
mouse liver. Apoptosis. 10:1261–1269. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yadati T, Houben T, Bitorina A and
Shiri-Sverdlov R: The Ins and outs of cathepsins: Physiological
function and role in disease management. Cells. 9:16792020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chevriaux A, Pilot T, Derangère V, Simonin
H, Martine P, Chalmin F, Ghiringhelli F and Rébé C: Cathepsin B is
required for NLRP3 inflammasome activation in macrophages, through
NLRP3 interaction. Front Cell Dev Biol. 8:1672020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang TT, Lv LL, Pan MM, Wen Y, Wang B, Li
ZL, Wu M, Wang FM, Crowley SD and Liu BC: Hydroxychloroquine
attenuates renal ischemia/reperfusion injury by inhibiting
cathepsin mediated NLRP3 inflammasome activation. Cell Death Dis.
9:3512018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Korkmaz B, Caughey GH, Chapple I, Gauthier
F, Hirschfeld J, Jenne DE, Kettritz R, Lalmanach G, Lamort AS,
Lauritzen C, et al: Therapeutic targeting of cathepsin C: From
pathophysiology to treatment. Pharmacol Ther. 190:202–236. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rehm SRT, Smirnova NF, Morrone C,
Götzfried J, Feuchtinger A, Pedersen J, Korkmaz B, Yildirim AÖ and
Jenne DE: Premedication with a cathepsin C inhibitor alleviates
early primary graft dysfunction in mouse recipients after lung
transplantation. Sci Rep. 9:99252019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dennemärker J, Lohmüller T, Müller S,
Aguilar SV, Tobin DJ, Peters C and Reinheckel T: Impaired turnover
of autophagolysosomes in cathepsin L deficiency. Biol Chem.
391:913–922. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McComb S, Shutinoski B, Thurston S,
Cessford E, Kumar K and Sad S: Cathepsins limit macrophage
necroptosis through cleavage of Rip1 kinase. J Immunol.
192:5671–5678. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Figueiredo JL, Aikawa M, Zheng C, Aaron J,
Lax L, Libby P, de Lima Filho JL, Gruener S, Fingerle J, Haap W, et
al: Selective cathepsin S inhibition attenuates atherosclerosis in
apolipoprotein E-deficient mice with chronic renal disease. Am J
Pathol. 185:1156–1166. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakanishi H: Neuronal and microglial
cathepsins in aging and age-related diseases. Ageing Res Rev.
2:367–381. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kos J, Sekirnik A, Premzl A, Zavasnik
Bergant V, Langerholc T, Turk B, Werle B, Golouh R, Repnik U, Jeras
M and Turk V: Carboxypeptidases cathepsins X and B display distinct
protein profile in human cells and tissues. Exp Cell Res.
306:103–113. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Polcyn R, Capone M, Hossain A, Matzelle D,
Banik NL and Haque A: Neuron specific enolase is a potential target
for regulating neuronal cell survival and death: Implications in
neurodegeneration and regeneration. Neuroimmunol Neuroinflamm.
4:254–257. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Obermajer N, Premzl A, Zavasnik Bergant T,
Turk B and Kos J: Carboxypeptidase cathepsin X mediates
beta2-integrin-dependent adhesion of differentiated U-937 cells.
Exp Cell Res. 312:2515–2527. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ondr JK and Pham CT: Characterization of
murine cathepsin W and its role in cell-mediated cytotoxicity. J
Biol Chem. 279:27525–27533. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wex T, Wex H, Hartig R, Wilhelmsen S and
Malfertheiner P: Functional involvement of cathepsin W in the
cytotoxic activity of NK-92 cells. FEBS Lett. 552:115–119. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stoeckle C, Gouttefangeas C, Hammer M,
Weber E, Melms A and Tolosa E: Cathepsin W expressed exclusively in
CD8+ T cells and NK cells, is secreted during target cell killing
but is not essential for cytotoxicity in human CTLs. Exp Hematol.
37:266–275. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kakehashi H, Nishioku T, Tsukuba T,
Kadowaki T, Nakamura S and Yamamoto K: Differential regulation of
the nature and functions of dendritic cells and macrophages by
cathepsin E. J Immunol. 179:5728–5737. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chain BM, Free P, Medd P, Swetman C, Tabor
AB and Terrazzini N: The expression and function of cathepsin E in
dendritic cells. J Immunol. 174:1791–1800. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamamoto K, Kawakubo T, Yasukochi A and
Tsukuba T: Emerging roles of cathepsin E in host defense
mechanisms. Biochim Biophys Acta. 1824:105–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deussing J, Roth W, Saftig P, Peters C,
Ploegh HL and Villadangos JA: Cathepsins B and D are dispensable
for major histocompatibility complex class II-mediated antigen
presentation. Proc Natl Acad Sci USA. 95:4516–4521. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Droga-Mazovec G, Bojic L, Petelin A,
Ivanova S, Romih R, Repnik U, Salvesen GS, Stoka V, Turk V and Turk
B: Cysteine cathepsins trigger caspase-dependent cell death through
cleavage of bid and antiapoptotic Bcl-2 homologues. J Biol Chem.
283:19140–19150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Conus S, Perozzo R, Reinheckel T, Peters
C, Scapozza L, Yousefi S and Simon HU: Caspase-8 is activated by
cathepsin D initiating neutrophil apoptosis during the resolution
of inflammation. J Exp Med. 205:685–698. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bidère N, Lorenzo HK, Carmona S, Laforge
M, Harper F, Dumont C and Senik A: Cathepsin D triggers Bax
activation, resulting in selective apoptosis-inducing factor (AIF)
relocation in T lymphocytes entering the early commitment phase to
apoptosis. J Biol Chem. 278:31401–31411. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Seyrantepe V, Hinek A, Peng J, Fedjaev M,
Ernest S, Kadota Y, Canuel M, Itoh K, Morales CR, Lavoie J, et al:
Enzymatic activity of lysosomal carboxypeptidase (cathepsin) A is
required for proper elastic fiber formation and inactivation of
endothelin-1. Circulation. 117:1973–1981. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jackman HL, Massad MG, Sekosan M, Tan F,
Brovkovych V, Marcic BM and Erdös EG: Angiotensin 1–9 and 1–7
release in human heart: Role of cathepsin A. Hypertension.
39:976–981. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Burster T, Macmillan H, Hou T, Boehm BO
and Mellins ED: Cathepsin G: Roles in antigen presentation and
beyond. Mol Immunol. 47:658–665. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Meyer-Hoffert U: Neutrophil-derived serine
proteases modulate innate immune responses. Front Biosci (Landmark
Ed). 14:3409–3418. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pintucci G, Iacoviello L, Castelli MP,
Amore C, Evangelista V, Cerletti C and Donati MB: Cathepsin
G-induced release of PAI-1 in the culture medium of endothelial
cells: A new thrombogenic role for polymorphonuclear leukocytes? J
Lab Clin Med. 122:69–79. 1993.PubMed/NCBI
|
|
37
|
Richter R, Bistrian R, Escher S, Forssmann
WG, Vakili J, Henschler R, Spodsberg N, Frimpong-Boateng A and
Forssmann U: Quantum proteolytic activation of chemokine CCL15 by
neutrophil granulocytes modulates mononuclear cell adhesiveness. J
Immunol. 175:1599–1608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Miao Z, Premack BA, Wei Z, Wang Y, Gerard
C, Showell H, Howard M, Schall TJ and Berahovich R: Proinflammatory
proteases liberate a discrete high-affinity functional FPRL1
(CCR12) ligand from CCL23. J Immunol. 178:7395–7404. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Brignone C, Munoz O, Batoz M,
Rouquette-Jazdanian A and Cousin JL: Proteases produced by
activated neutrophils are able to release soluble CD23 fragments
endowed with proinflammatory effects. FASEB J. 15:2027–2029. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang Z, Weian C, Susu H and Hanmin W:
Protective effects of mangiferin on cerebral ischemia-reperfusion
injury and its mechanisms. Eur J Pharmacol. 771:145–151. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Seyfried DM, Veyna R, Han Y, Li K, Tang N,
Betts RL, Weinsheimer S, Chopp M and Anagli J: A selective cysteine
protease inhibitor is non-toxic and cerebroprotective in rats
undergoing transient middle cerebral artery ischemia. Brain Res.
901:94–101. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Benchoua A, Braudeau J, Reis A, Couriaud C
and Onténiente B: Activation of proinflammatory caspases by
cathepsin B in focal cerebral ischemia. J Cereb Blood Flow Metab.
24:1272–1279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Guicciardi ME, Deussing J, Miyoshi H,
Bronk SF, Svingen PA, Peters C, Kaufmann SH and Gores GJ: Cathepsin
B contributes to TNF-alpha-mediated hepatocyte apoptosis by
promoting mitochondrial release of cytochrome c. J Clin Invest.
106:1127–1137. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tsubokawa T, Solaroglu I, Yatsushige H,
Cahill J, Yata K and Zhang JH: Cathepsin and calpain inhibitor E64d
attenuates matrix metalloproteinase-9 activity after focal cerebral
ischemia in rats. Stroke. 37:1888–1894. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Qin XF, Lu XJ, Ge JB, Xu HZ, Qin HD and Xu
F: Ginkgolide B prevents cathepsin-mediated cell death following
cerebral ischemia/reperfusion injury. Neuroreport. 25:267–273.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wei R, Wang J, Xu Y, Yin B, He F, Du Y,
Peng G and Luo B: Probenecid protects against cerebral
ischemia/reperfusion injury by inhibiting lysosomal and
inflammatory damage in rats. Neuroscience. 301:168–177. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhu YM, Gao X, Ni Y, Li W, Kent TA, Qiao
SG, Wang C, Xu XX and Zhang HL: Sevoflurane postconditioning
attenuates reactive astrogliosis and glial scar formation after
ischemia-reperfusion brain injury. Neuroscience. 356:125–141. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cui DR, Wang L, Jiang W, Qi AH, Zhou QH
and Zhang XL: Propofol prevents cerebral ischemia-triggered
autophagy activation and cell death in the rat hippocampus through
the NF-κB/p53 signaling pathway. Neuroscience. 246:117–132. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ahsan A, Zheng Y, Ma S, Liu M, Cao M, Li
Y, Zheng W, Zhou X, Xin M, Hu WW, et al: Tomatidine protects
against ischemic neuronal injury by improving lysosomal function.
Eur J Pharmacol. 882:1732802020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kilinc M, Gürsoy-Ozdemir Y, Gürer G,
Erdener SE, Erdemli E, Can A and Dalkara T: Lysosomal rupture,
necroapoptotic interactions and potential crosstalk between
cysteine proteases in neurons shortly after focal ischemia.
Neurobiol Dis. 40:293–302. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang F, Gómez-Sintes R and Boya P:
Lysosomal membrane permeabilization and cell death. Traffic.
19:918–931. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kirkegaard T, Roth AG, Petersen NH,
Mahalka AK, Olsen OD, Moilanen I, Zylicz A, Knudsen J, Sandhoff K,
Arenz C, et al: Hsp70 stabilizes lysosomes and reverts Niemann-Pick
disease-associated lysosomal pathology. Nature. 463:549–553. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yamashima T: Hsp70.1 and related lysosomal
factors for necrotic neuronal death. J Neurochem. 120:477–494.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Heusch G and Gersh BJ: The pathophysiology
of acute myocardial infarction and strategies of protection beyond
reperfusion: A continual challenge. Eur Heart J. 38:774–784.
2017.PubMed/NCBI
|
|
55
|
Vander Heide RS and Steenbergen C:
Cardioprotection and myocardial reperfusion: Pitfalls to clinical
application. Circ Res. 113:464–477. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Frangogiannis NG: Regulation of the
inflammatory response in cardiac repair. Circ Res. 110:159–173.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kain V, Prabhu SD and Halade GV:
Inflammation revisited: Inflammation versus resolution of
inflammation following myocardial infarction. Basic Res Cardiol.
109:4442014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Meyer-Hoffert U and Wiedow O: Neutrophil
serine proteases: Mediators of innate immune responses. Curr Opin
Hematol. 18:19–24. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sabri A, Alcott SG, Elouardighi H, Pak E,
Derian C, Andrade-Gordon P, Kinnally K and Steinberg SF: Neutrophil
cathepsin G promotes detachment-induced cardiomyocyte apoptosis via
a protease-activated receptor-independent mechanism. J Biol Chem.
278:23944–23954. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Iacoviello L, Kolpakov V, Salvatore L,
Amore C, Pintucci G, de Gaetano G and Donati MB: Human endothelial
cell damage by neutrophil-derived cathepsin G. Role of cytoskeleton
rearrangement and matrix-bound plasminogen activator inhibitor-1.
Arterioscler Thromb Vasc Biol. 15:2037–2046. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hooshdaran B, Kolpakov MA, Guo X, Miller
SA, Wang T, Tilley DG, Rafiq K and Sabri A: Dual inhibition of
cathepsin G and chymase reduces myocyte death and improves cardiac
remodeling after myocardial ischemia reperfusion injury. Basic Res
Cardiol. 112:622017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Taleb S, Cancello R, Clément K and Lacasa
D: Cathepsin S promotes human preadipocyte differentiation:
Possible involvement of fibronectin degradation. Endocrinology.
147:4950–4959. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chen H, Wang J, Xiang MX, Lin Y, He A, Jin
CN, Guan J, Sukhova GK, Libby P, Wang JA and Shi GP: Cathepsin
S-mediated fibroblast trans-differentiation contributes to left
ventricular remodelling after myocardial infarction. Cardiovasc
Res. 100:84–94. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Peng K, Liu H, Yan B, Meng XW, Song SY, Ji
FH and Xia Z: Inhibition of cathepsin S attenuates myocardial
ischemia/reperfusion injury by suppressing inflammation and
apoptosis. J Cell Physiol. 236:1309–1320. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Linz D, Hohl M, Dhein S, Ruf S, Reil JC,
Kabiri M, Wohlfart P, Verheule S, Böhm M, Sadowski T and Schotten
U: Cathepsin A mediates susceptibility to atrial tachyarrhythmia
and impairment of atrial emptying function in Zucker diabetic fatty
rats. Cardiovasc Res. 110:371–380. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hohl M, Erb K, Lang L, Ruf S, Hübschle T,
Dhein S, Linz W, Elliott AD, Sanders P, Zamyatkin O, et al:
Cathepsin A mediates ventricular remote remodeling and atrial
cardiomyopathy in rats with ventricular ischemia/reperfusion. JACC
Basic Transl Sci. 4:332–344. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Turski WA and Zasłonka J: Activity of
cathepsin D and L in the heart muscle of coronary patients during
coronary-aortal bypass graft operation. Med Sci Monit. 6:853–860.
2000.PubMed/NCBI
|
|
68
|
Turski WA and Zasłonka J: Effects of
Bretschneider cardioplegic fluid on the lysosomal cathepsins D and
L of myocardium of coronary patients during coronary-aortal bypass
graft operation. Med Sci Monit. 6:861–866. 2000.PubMed/NCBI
|
|
69
|
Zhai Y, Petrowsky H, Hong JC, Busuttil RW
and Kupiec-Weglinski JW: Ischaemia-reperfusion injury in liver
transplantation-from bench to bedside. Nat Rev Gastroenterol
Hepatol. 10:79–89. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Baskin-Bey ES, Canbay A, Bronk SF,
Werneburg N, Guicciardi ME, Nyberg SL and Gores GJ: Cathepsin B
inactivation attenuates hepatocyte apoptosis and liver damage in
steatotic livers after cold ischemia-warm reperfusion injury. Am J
Physiol Gastrointest Liver Physiol. 288:G396–G402. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Guicciardi ME, Miyoshi H, Bronk SF and
Gores GJ: Cathepsin B knockout mice are resistant to tumor necrosis
factor-alpha-mediated hepatocyte apoptosis and liver injury:
Implications for therapeutic applications. Am J Pathol.
159:2045–2054. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhou H, Zhou S, Shi Y, Wang Q, Wei S, Wang
P, Cheng F, Auwerx J, Schoonjans K and Lu L: TGR5/Cathepsin E
signaling regulates macrophage innate immune activation in liver
ischemia and reperfusion injury. Am J Transplant. 21:1453–1464.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Mehta RL, Kellum JA, Shah SV, Molitoris
BA, Ronco C, Warnock DG and Levin A; Acute Kidney Injury Network, :
Acute Kidney Injury Network: Report of an initiative to improve
outcomes in acute kidney injury. Crit Care. 11:R312007. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Eltzschig HK, Bonney SK and Eckle T:
Attenuating myocardial ischemia by targeting A2B adenosine
receptors. Trends Mol Med. 19:345–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yap SC and Lee HT: Adenosine and
protection from acute kidney injury. Curr Opin Nephrol Hypertens.
21:24–32. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Suzuki C, Tanida I, Ohmuraya M, Oliva
Trejo JA, Kakuta S, Sunabori T and Uchiyama Y: Lack of Cathepsin D
in the renal proximal tubular cells resulted in increased
sensitivity against renal ischemia/reperfusion injury. Int J Mol
Sci. 20:17112019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Cocchiaro P, Fox C, Tregidgo NW, Howarth
R, Wood KM, Situmorang GR, Pavone LM, Sheerin NS and Moles A:
Lysosomal protease cathepsin D; a new driver of apoptosis during
acute kidney injury. Sci Rep. 6:271122016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Fox C, Cocchiaro P, Oakley F, Howarth R,
Callaghan K, Leslie J, Luli S, Wood KM, Genovese F, Sheerin NS and
Moles A: Inhibition of lysosomal protease cathepsin D reduces renal
fibrosis in murine chronic kidney disease. Sci Rep. 6:201012016.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Shimoda N, Fukazawa N, Nonomura K and
Fairchild RL: Cathepsin G is required for sustained inflammation
and tissue injury after reperfusion of ischemic kidneys. Am J
Pathol. 170:930–940. 2007. View Article : Google Scholar : PubMed/NCBI
|