Introduction
The liver plays a central role in metabolic
homeostasis (1). Although hepatic
metabolism is decreased by acute liver injuries, such as hepatitis
and transplantation, this function is restored by liver
regeneration based on the rapid proliferation of hepatocytes
(2). The 70% partial hepatectomy
(PHx) model has been widely used to examine the mechanisms of liver
regeneration, repair after tissue injury, and hepatocytes cell
cycle dynamics in vivo (3). Particularly, nonparenchymal liver
cells are known to promote liver regeneration by the production of
inflammatory cytokines, such as interleukin-6 (IL-6) and tumor
necrosis factor-alpha (TNF-α), via the TLR/MyD88/NF-kB axis
(4,5). In addition, the TLR/MyD88/NF-kB
pathway is activated by a gut-derived lipopolysaccharide (LPS) and
damage-associated molecular pattern in a partial hepatectomy (PHx)
(6). Furthermore, heat-shock
proteins, which are endogenous TLR ligands, are involved in liver
regeneration after PHx (7).
Liver regeneration is promoted not only by
inflammatory cytokines but also by other factors, such as
tryptophan (Trp) metabolites. The expression of indoleamine
2,3-dioxygenase (Ido) 1, which catalyzes the oxidation of Trp to
kynurenine (Kyn) in the first step of the Kyn pathway (8,9),
is induced in various immune cells and tissues under inflammatory
conditions. Ido1-derived Kyn regulates various immune responses
through its interaction with the aryl hydrocarbon receptor (Ahr),
which inhibits NF-κB activation (10,11). We recently reported that
inhibition of Ido1 accelerates early liver regeneration after PHx
by increasing the expression of cell cycle and pro-inflammatory
cytokine genes (12).
Furthermore, Kyn treatment suppressed liver regeneration in WT
mice. In contrast, Ido2, an isozyme of Ido1, is constitutively
expressed in the liver, placenta, central nervous system, and
macrophages, and its expression is regulated by various
inflammatory mediators, including interferon-γ, IL-10, and
prostaglandin E2 (13).
Constitutive expression of Ido2 is crucial for the prevention of
inflammatory diseases, such as psoriasis and endotoxin shock
(14,15). We previously reported that
Ido2-derived Kyn in hepatocytes prevents severe hepatocellular
damage and liver fibrosis induced by CCl4 through activation of Ahr
signaling-dependent inflammatory responses (16). However, the biological and
physiological roles of Ido2 under liver regeneration conditions
remain unclear. Thus, we investigated the role of Ido2 expression
in intrahepatic MNCs after PHx.
Materials and methods
Ethical approval
The ethics governing the use and conduct of
experiments on animals were strictly observed, and the experimental
protocol was approved by the Animal Care of Fujita Health
University. The protocol for all animal experiments was approved by
the Animal Experimentation Committee of Fujita Health University
(approval no. AP20031-R21). Procedures involving mice and their
care conformed to international guidelines, as described in
Principles of Laboratory Animal Care (National Institutes of Health
publication 85–23, revised 1985).
Mice
Ido2-KO mice on a C57BL/6 N background were obtained
from the Knockout Mouse Project (KOMP, CA, USA). Homozygous Ido2-KO
mice generated by intercrossing heterozygous mice were used for the
following experiments. These mice were housed in a specific
pathogen-free environment in our facility, maintained at 25°C room
temperature, 40–60% humidity, on a 12 h light/dark cycle (lights on
at 08:00), and had free access to food and water.
Animal experiment
Mice underwent 70% PHx, as previously described
(17). Briefly, mice were incised
in the upper abdominal wall, the liver was exposed, and three
anterior lobes (right medial, left medial, and left lateral) were
rejected by proximal ligation under inhalational anesthetic with
isoflurane (anesthetic induction on 4% isoflurane, followed by
anesthetic maintenance on 2% isoflurane). Mice were kept warm at
37°C until postoperative awakening. The control group undergone
midline laparotomy incision without liver lobe resection. Mice were
randomly selected for the partial hepatectomy treatment group at
each time point. Mice were sacrificed by cervical dislocation at
each time point and analyzed (17,18). The experiments complied with the
ARRIVE guidelines (19).
Immunohistochemistry
The livers were fixed in 10% buffered formalin and
embedded in paraffin. Immunohistochemical staining for PCNA were
used to evaluate hepatocyte proliferation in the liver as
previously described (18).
Briefly, 4-µm thick sections of the livers were deparaffinized and
treated with 3% hydrogen peroxide to inactivate endogenous
peroxidases. The sections were heated in 0.1 M citrate buffer (pH
6.0) using the autoclave. The sections were treated with 10% goat
serum (Invitrogen, Carlsbad, CA, USA) for 60 min to prevent
non-specific antibody binding and then incubated with anti-PCNA
(1:2,000, catalog no. LS-B14132, LSBio, Seattle, WA, USA) overnight
at 4°C. After washing, primary antibody-stained sections were
incubated with horseradish peroxidase (HRP)-conjugated rabbit
immunoglobulin antibody solution (1:1, Histofine Simple Stain
MAX-PO(R), Nichirei, Tokyo, Japan), followed by
3,3′-diaminobenzidine staining (catalog no. K3468, DAKO, Tokyo,
Japan). Finally, the sections were counterstained using Mayer's
hematoxylin.
RNA extraction and real-time qPCR
analysis
Total RNA was extracted from cell lines with
ISOGENII (Nippon gene, Tokyo, Japan) and reverse transcription-PCR
was performed using Revatra Ace Kits (Toyobo, Tokyo, Japan). PCR
amplification was performed using Sso advanced SYBR Green Supermix
(Bio-Rad, Hercules, CA, USA). The thermocycling conditions were as
follows: denaturation at 95°C for 2 min, annealing at 60°C for 30
sec and elongation at 72°C for 20 sec. The qPCR primers were as
follows: IL-6 sense 5′-GATACCACTCCCAACAGA-3′ and antisense
5′-GCCATTGCACAACTCTTT-3′; TNF-α sense 5′-TCATGCACCACCATCAAG-3′ and
antisense 5′-CAGAACTCAGGAATGGACAT-3′; Ido1 sense
5′-AGTTGGGCCTGCCTCCTATTC-3′ and antisense
5′-GAAGAAGCCCTTGTCGCAGTC-3′; Ido2 sense 5′-CATACCAGGCAATTGCTCCAC-3′
and antisense 5′-GCCTGGGCTAAAGAGCTCAATAC-3′; 18s sense,
5′-GGATTGACAGATTGATAGC-3′ and antisense 5′-TATCGGAATTAACCAGACAA-3′.
CyclinD1 sense, 5′-AGTGCGTGCAGAAGGAGATT-3′ and antisense
5′-CACAACTTCTCGGCAGTCAA-3′; Birc5 sense
5′-TACCTCAAGAACTACCGCATCG-3′ and antisense
5′-AAGGCTCAGCATTAGGCAGC-3′; Foxm1 sense
5′-GGAGGAAATGCCACACTTAGCG-3′ and antisense
5′-TAGGACTTCTTGGGTCTTGGGGTG-3′. The mRNA expression levels of IL-6,
TNF-α, Ido1, Ido2, and 18s rRNA were analyzed on a 7500 Fast
Real-Time PCR Systems in conjunction with Applied Biosystems™
software (Applied Biosystems).
Measurements of kynurenine pathway
metabolites
Trp and Kyn levels were measured as previously
reported (16). Briefly, MNCs and
liver were homogenized (1:4, w/v) in 10% perchloric acid. Next, 50
µl of the supernatants were subjected to high-performance liquid
chromatography (HPLC; SHIMADU, Kyoto, Japan).
Measurement of serum cytokines
The concentration of IL-6 and TNF-α in the sera was
measured by Mouse ELISA Kit (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer's instructions.
MNCs isolation
MNCs were isolated as previously described (17,18). The livers of mice treated with or
without PHx were perfused slowly via the inferior vena cava with 10
ml of cold PBS. Briefly, the excised liver was cut into small
pieces with scissors in digestion medium (RPMI-1640 containing 10%
fetal bovine serum and 20 U/ml collagenase Type I) and then
incubated at 37°C for 45 min with shaking at 180 rpm. After
incubation, collagenase-digested hepatic cells were pressed through
70 µm cell strainer and were centrifuged at 50 × g for 3 min at
4°C. Then, the supernatant was collected and centrifuged at 600 × g
for 5 min at 4°C. Pellet was applied to a discontinuous 70 and 40%
Percoll density gradient (Sigma-Aldrich, St Louis, MO, USA), and
the cells at the interface were used as mononuclear cells.
Cell culture and immunocytochemical
analysis
Murine resident peritoneal macrophages were
stimulated with LPS (1 ng/ml, Escherichia coli O55:B5;
Sigma-Aldrich) for indicated time periods. After cultivation, the
cells were fixed with 4% paraformaldehyde/PBS for 10 min and
permeabilized with 0.2% TritonX-100/PBS for 10 min. After being
blocked with 5% bovine serum albumin (BSA)/PBS for 60 min, the
cells were incubated with rat monoclonal anti-F4/80 antibody (1:50,
catalog no. ab16911, Abcam Cambridge, U.K.) or rabbit monoclonal
anti-NF-κB (1:400, catalog no. #8242, Cell Signaling Technology,
Danvers, MA) at 4°C, overnight. After washing, the cells were
incubated with goat anti-rat IgG (H+L) antibody (1:200, catalog no.
ab150157, Abcam) or rabbit anti-rat IgG (H+L) antibody (1:500,
catalog no. 611-142-122, Rockland Immunochemicals, Inc. Limerick,
PA) for 1 h, respectively. Nuclei were stained with
4,6-diamidino-2-phenylindole (Dojindo, Tokyo, Japan).
Western blotting and nuclear
extraction
The livers were washed with cold PBS.
Nuclear/cytosolic fractions were isolated using Lysopure™ Nuclear
and Cytoplasmic Extractor Kit (Wako Chemical), according to the
manufacturer's protocol. Ten micrograms of protein were loaded on
10% Mini-PROTEAN TGX gels (Bio-Rad) and transferred to PVDF
membranes. The membranes were incubated with anti-IκB (1:1,000,
catalog no. #4812, Cell Signaling Technology), anti-NF-κB p65
(1:1,000, catalog no. #8242, Cell Signaling Technology), anti-Lamin
B1 (1:2,000, catalog No. 66095-1-lg, proteintech), β-actin (catalog
no. A5441, Sigma-Aldrich), anti-Ido2 (1:1,000, catalog no.
ab214214, abcam), anti-Ido1 (1:1,000, catalog no. MABF850, Merck
Millipore, Darmstadt, Germany), or anti-Tdo2 (1:1,000, catalog no.
MABN1537, Merck Millipore). The membranes were then incubated with
HRP-conjugated anti-mouse IgG (1:10,000, catalog no. NA931A,
Cytiva., Tokyo, Japan) or HRP-conjugated anti-rabbit IgG (1:5,000,
catalog no. NA934V, Cytiva., Tokyo, Japan), respectively. The
detection of target proteins was performed with Amersham ECL Prime
Western Blotting Detection Reagent (Cytiva). Protein levels were
quantified by ImageJ software (National Institutes of Health,
Bethesda, MD). To re-probe the PVDF membranes, the antibodies bound
to the membranes were removed by a commercial stripping
solution.
Statistical analysis
Statistics were analyzed with GraphPad Prism 6
Software (GraphPad Software Inc., San Diego, USA). All data are
expressed as means ± SEM. Differences between WT and knockout mouse
groups were analyzed by the Kruskal-Wallis test followed by
Scheffe's test. A value of P<0.05 was considered statistically
significant (Fig. 1A). For the
values obtained in time course experiments, statistical analysis
was performed using two-way ANOVA followed by Tukey's
multiple-comparison test. A P-value of <0.05 was considered
statistically significant.
Results
Ido2 deficiency promotes liver
regeneration after PHx
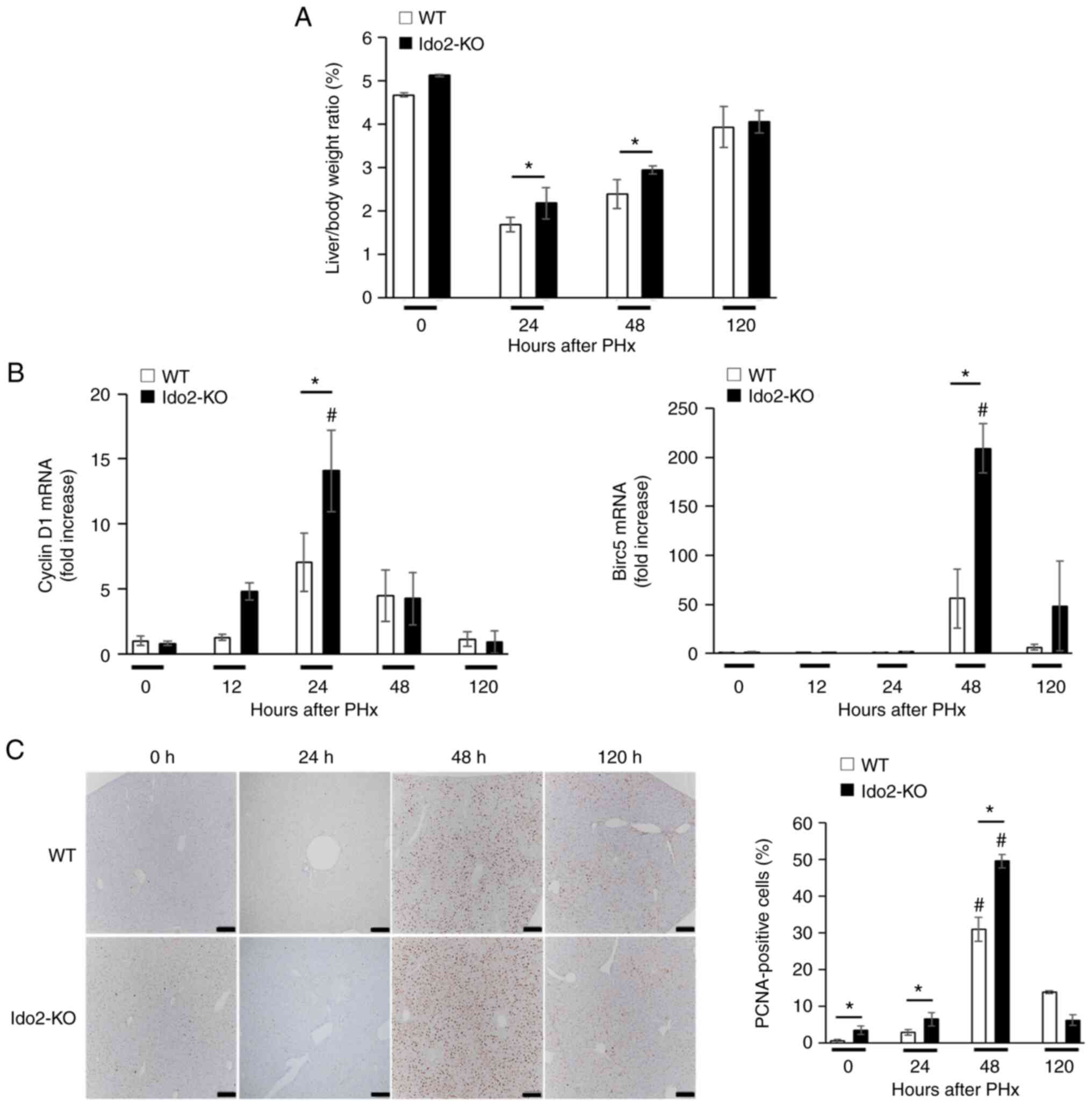
To investigate the roles of Ido2 in liver
regeneration, we compared liver weight and hepatocyte growth
between WT and Ido2-KO mice that were subjected to PHx. Although
the ratio of liver to body weight after PHx was significantly
higher in Ido2-KO mice than in WT mice at 24 and 48 h after PHx,
the ratio was comparable between these mice at 120 h (Fig. 1A). The expression of
pro-proliferative genes (Cyclin D1 and Birc5) was significantly
higher in Ido2-KO mice than that in WT mice. (Fig. 1B). Consistent with these results,
the frequency of PCNA+ cells were significantly
increased in Ido2-KO mice compared to WT mice 48 h after PHx
(Fig. 1C). These results suggest
that Ido2 deficiency accelerates liver regeneration after PHx.
Elevated Ido2 expression in
intrahepatic mononuclear cells after PHx
Next, we examined Ido2 expression during liver
regeneration after PHx. Ido2 expression was reduced in the total
liver 24 h after PHx but was restored within 48 h (Fig. 2A). In contrast, Ido2 expression in
intrahepatic MNCs was increased 24 h after PHx and was decreased to
baseline levels within 48 h (Fig.
2B). Intriguingly, the levels of Ido1 mRNA expression and Kyn
in total liver or intrahepatic MNCs after PHx were comparable
between Ido2-KO mice and WT mice (Figs. 2C, D, and Fig. S1). Since Ido2 expression was
mainly induced in intrahepatic MNCs after PHx (Fig. 2A and B), we focused on
intrahepatic MNCs in the following experiments.
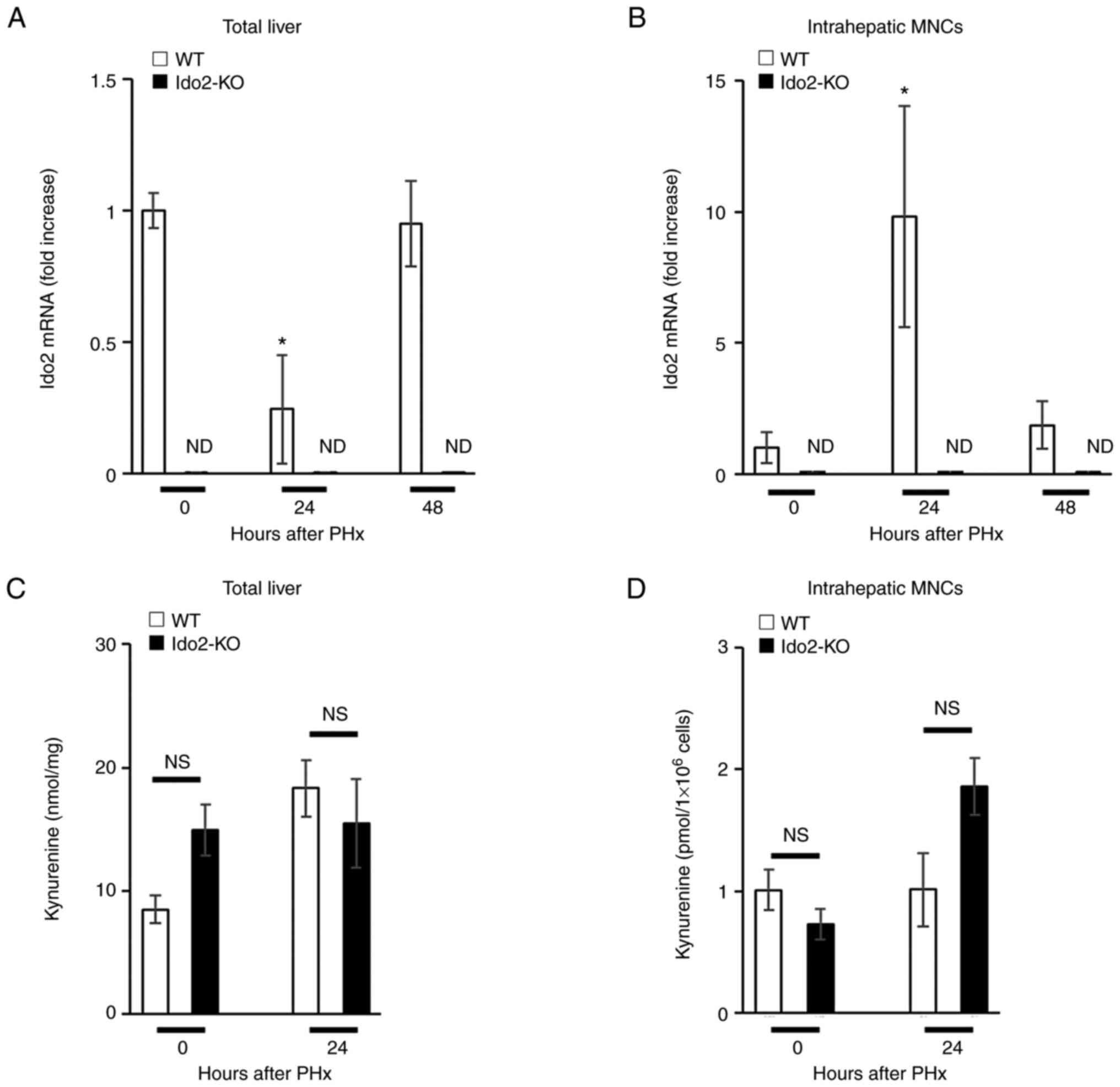 | Figure 2.Expression of the Ido2 gene is
increased in intrahepatic MNCs following PHx. WT (n=3) and Ido2-KO
(n=3) mice were subjected to 70% PHx. After each time point, the
mice were sacrificed and total liver and intrahepatic MNCs were
isolated. (A) Total liver and (B) intrahepatic MNCs were analyzed
to assess Ido2 mRNA expression using reverse
transcription-quantitative PCR. (ND, not detected). The results
were normalized to the expression of 18S rRNA. (C) Kyn
concentration in total liver from WT and Ido2-KO mice was
determined using HPLC (0 h group, n=3; 24 h group, n=8). (D) Kyn
concentration in intrahepatic MNCs from WT (n=3) and Ido2-KO (n=3)
mice was determined using HPLC. *P<0.05 compared with 0 h in
each mouse. NS, not significant; HPLC, high-performance liquid
chromatography; WT, wild-type; Ido2, indoleamine 2,3-dioxygenase 2;
KO, knockout; PHx, partial hepatectomy; MNC, mononuclear cells;
rRNA, ribosomal RNA; Kyn, kynurenine. |
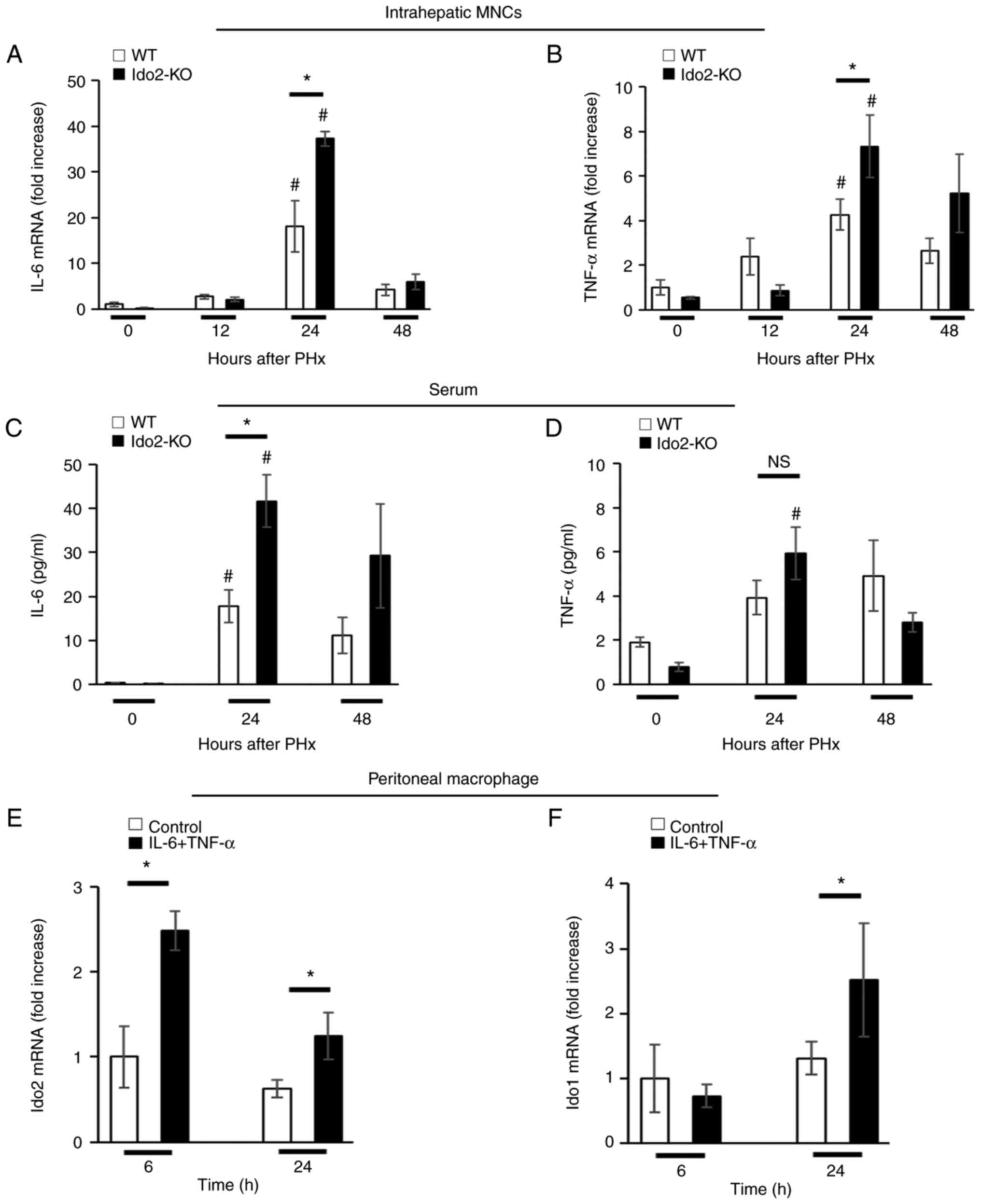
Ido2 deficiency enhances inflammatory
cytokine production after PHx
Hepatic expression of TNF-α and IL-6 is essential
for hepatocyte proliferation (1,2).
We measured the expression of these inflammatory cytokines in
intrahepatic MNCs after PHx. The expression of IL-6 and TNF-α mRNA
in intrahepatic MNCs was significantly higher in Ido2-KO mice than
in WT mice 24 h after PHx (Fig. 3A
and B). Similar results were obtained in the sera after PHx.
(Fig. 3C and D).
Next, we assessed whether Ido2 expression was
induced by these inflammatory cytokines. Therefore, we used
peritoneal resident macrophages instead of intrahepatic MNCs
because the frequency of MNCs in the liver is low to isolate as
single cells. Ido2 and Ido1 mRNA expression was induced in
peritoneal macrophages by a mixture of IL-6 and TNF-α (Fig. 3E and F). These results suggest
that Ido2 is rapidly induced by inflammatory cytokines.
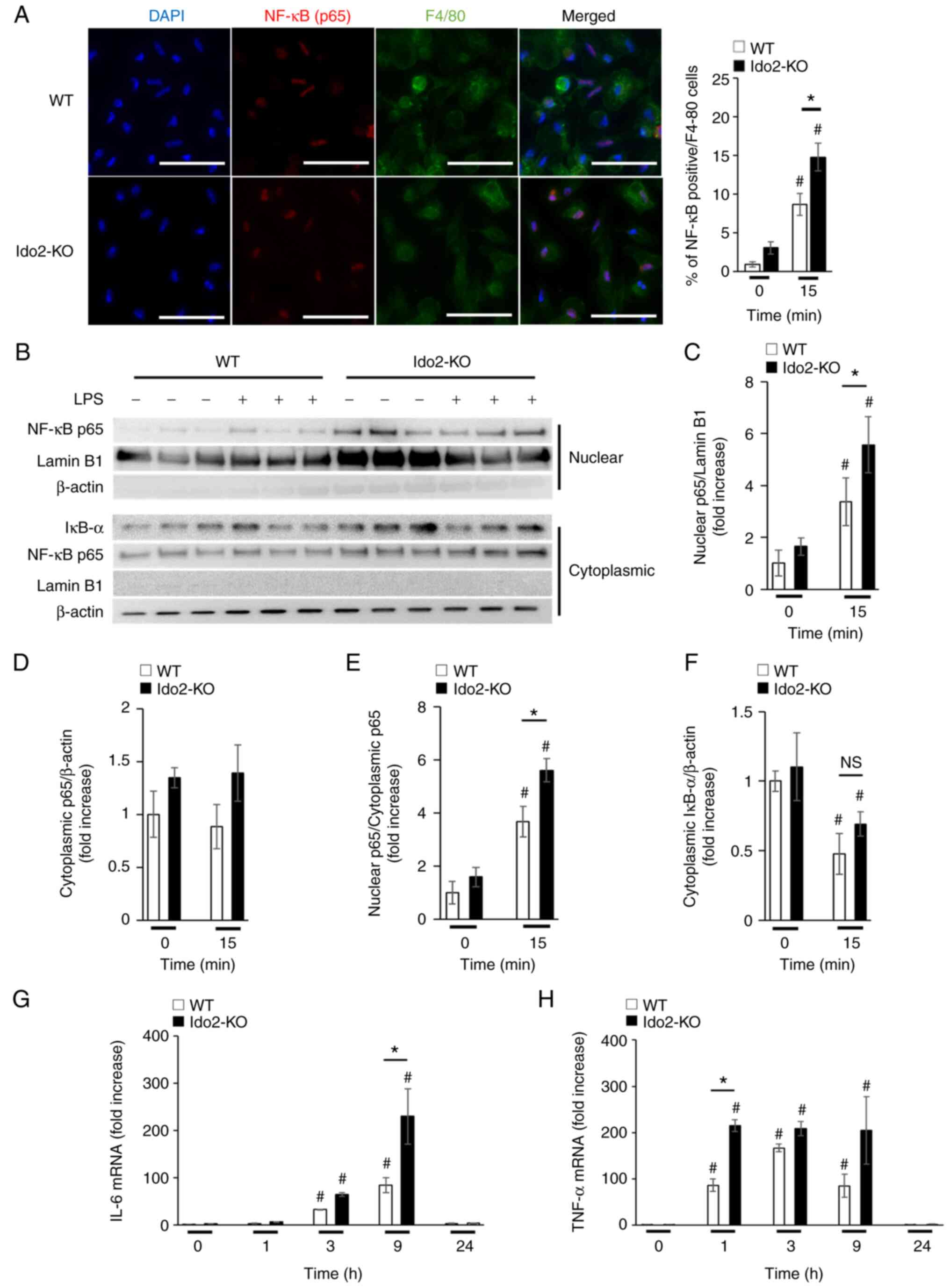
Ido2 deficiency enhances NF-κB
intranuclear transport in intrahepatic MNCs
The production of TNF-α and IL-6 after PHx is
dependent on NF-κB signaling (4,5).
To determine whether NF-κB-mediated production of these cytokines
is enhanced in Ido2-KO mice, peritoneal macrophages were stimulated
with LPS, which mimics stimulation with the endogenous TLR ligands
that are produced after PHx. Upon stimulation with LPS, the nuclear
transport of NF-κB (p65) in macrophages was increased in Ido2-KO
mice compared to WT mice, as measured by immunocytochemical
(Fig. 4A) and western blotting
analysis (Fig. 4B-F). Moreover,
the mRNA expression levels of IL-6 (Fig. 4G) and TNF-α (Fig. 4H) in peritoneal macrophages from
Ido2-KO mice after LPS stimulation were significantly enhanced
compared with those in WT mice. Importantly, the levels of Ido1 and
Tdo2 protein in peritoneal macrophages were comparable between WT
and Ido2-KO mice at this time point (Fig. S2). These results suggest that
increased Ido2 expression in peritoneal macrophages may regulate
inflammatory cytokine production by suppressing NF-κB
signaling.
Discussion
In the present study, we demonstrated that Ido2
deficiency elevated the production of inflammatory cytokines and
promoted liver regeneration after PHx. Elevated expression of
inflammatory cytokines is suppressed by the intrinsic Ido2
expression in immune cells (such as intrahepatic MNCs and possibly
macrophages). Further, Ido2 expression in intrahepatic MNCs may
regulate NF-κB signaling and control inflammatory cytokine
production after PHx.
Absence of Ido2 leads to the development of
inflammatory diseases such as endotoxin shock and psoriasis via the
aberrant production of inflammatory cytokines (14,15). Similarly, although aberrant
production of inflammatory cytokines is induced in intrahepatic
MNCs from PHx-treated Ido2-KO mice, these cytokines are essential
for hepatocyte growth, which leads to liver regeneration.
Therefore, Ido2 deficiency may have beneficial effects on liver
regeneration.
Kyn is known to inhibit NF-κB signaling-mediated
production of inflammatory cytokines through its interaction with
Ahr (20,21). We recently reported that absence
of Ido2 suppresses Ahr signaling in hepatocytes (16). In this context, we showed that
Ido1 mRNA expression and production of Kyn did not differ between
WT and Ido2-KO mice after PHx. Therefore, regulation of NF-κB by
Kyn cannot be ruled out, but is considered to be small. Based on
our findings, the role of Ido2 may differ between intrahepatic MNCs
and hepatocytes in terms of Ahr/NF-κB signaling.
We found that intrahepatic MNC-intrinsic Ido2
regulates hepatocyte proliferation after PHx, which raises some
questions. First, does hepatocyte-intrinsic Ido2 regulate
hepatocyte proliferation? Under steady-state conditions, the
frequencies of PCNA+ hepatocytes were found to be
significantly higher in Ido2-KO mice than in WT mice regardless of
inflammatory cytokine production, suggesting that
hepatocyte-intrinsic Ido2 is dispensable for regeneration after
PHx. Second, how does Ido2 affect NF-κB activation in a
Kyn-independent manner? Ido1 and possibly Ido2 are reported to play
non-enzymatic signaling role (13,22,23). In a transforming growth factor β
(TGF-β)-dominated microenvironment, Ido1, which is phosphorylated
by Fyn kinase, interacts with SH2 domain-containing phosphatases
and phosphatidylinositol-3 kinase (p85), leading to the activation
of non-canonical NF-κB (22).
Recently, the non-enzymatic signaling capacity of Ido2 has been
reported to be required for autoimmune arthritis development
(23). According to this report,
Ido2 interacts with Runx1, GAPDH, RANbp10, and Mgea5, all of which
may be involved in immune signaling. Interestingly, Runx1 interacts
with NF-κB (p50) to produce inflammatory cytokines in
LPS-stimulated macrophages (24),
implying that Ido2 might suppress NF-κB activation via regulation
of Runx1. Although we did not identify Ido2-interacting proteins in
a PHx model, our findings provide a promising potential for Ido2 as
a non-enzymatic signaling protein that regulates the NF-κB pathway
through a negative feedback loop. Further investigation is needed
to identify the mechanism by which Ido2 and its product Kyn
regulate liver regeneration through inflammatory cytokine
production.
In conclusion, this study suggests that Ido2
deficiency in intrahepatic MNCs augments inflammatory cytokine
production and hepatocyte proliferation following PHx. Ido2 may
inhibit inflammatory cytokine production by regulating NF-κB in
intrahepatic MNCs. Therefore, Ido2 may be a novel target for liver
regeneration in various hepatic injuries.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by Grants-in-Aids for Research Activity
Start-up (grant no. 19K23873) from the Japan Society for the
Promotion of Science (JSPS) and the Private University Research
Branding Project from the Ministry of Education, Culture, Sports,
Science and Technology of Japan (MEXT).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TA and MH designed the study. TA, MH and HT
performed the experiments. TA, MH, and KN were responsible for data
integrity and analyses. TA, MH, HT, HI, KN, YY and KS discussed and
interpreted the data. TA, MH, HT and HI drafted the manuscript. TA,
MH, HT, YY and KS conducted the study. TA and MH confirm the
authenticity of all the raw data. KS had the primary responsibility
for the final content. All authors read and approved the
manuscript.
Ethics approval and consent to
participate
The experimental protocol was approved by the Animal
Care of Fujita Health University. The protocol for all animal
experiments was approved by the Animal Experimentation Committee of
Fujita Health University (approval no. AP20031-R21).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Taub R: Liver regeneration: From myth to
mechanism. Nat Rev Mol Cell Biol. 5:836–847. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fausto N: Liver regeneration. J Hepatol.
32 (1 Suppl):S19–S31. 2000. View Article : Google Scholar
|
|
3
|
Mitchell C and Willenbring H: A
reproducible and well-tolerated method for 2/3 partial hepatectomy
in mice. Nat Protoc. 3:1167–1170. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Seki E, Tsutsui H, Iimuro Y, Naka T, Son
G, Akira S, Kishimoto T, Nakanishi K and Fujimoto J: Contribution
of Toll-like receptor/myeloid differentiation factor 88 signaling
to murine liver regeneration. Hepatology. 41:443–450. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Y and Sun R: Toll-like receptors in
acute liver injury and regeneration. Int Immunopharmacol.
11:1433–1441. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cornell RP, Liljequist BL and Bartizal KF:
Depressed liver regeneration after partial hepatectomy of
germ-free, athymic and lipopolysaccharide-resistant mice.
Hepatology. 11:916–922. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wolf JH, Bhatti TR, Fouraschen S,
Chakravorty S, Wang L, Kurian S, Salomon D, Olthoff KM, Hancock WW
and Levine MH: Heat shock protein 70 is required for optimal liver
regeneration after partial hepatectomy in mice. Liver Transpl.
20:376–385. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hoshi M, Matsumoto K, Ito H, Ohtaki H,
Arioka Y, Osawa Y, Yamamoto Y, Matsunami H, Hara A, Seishima M and
Saito K: L-tryptophan-kynurenine pathway metabolites regulate type
I IFNs of acute viral myocarditis in mice. J Immunol.
188:3980–3987. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ito H, Hoshi M, Ohtaki H, Taguchi A, Ando
K, Ishikawa T, Osawa Y, Hara A, Moriwaki H, Saito K and Seishima M:
Ability of IDO to attenuate liver injury in
alpha-galactosylceramide-induced hepatitis model. J Immunol.
185:4554–4560. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bessede A, Gargaro M, Pallotta MT, Matino
D, Servillo G, Brunacci C, Bicciato S, Mazza EM, Macchiarulo A,
Vacca C, et al: Aryl hydrocarbon receptor control of a disease
tolerance defence pathway. Nature. 511:184–190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takenaka MC, Gabriely G, Rothhammer V,
Mascanfroni ID, Wheeler MA, Chao CC, Gutiérrez-Vázquez C, Kenison
J, Tjon EC, Barroso A, et al: Control of tumor-associated
macrophages and T cells in glioblastoma via AHR and CD39. Nat
Neurosci. 22:729–740. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ogiso H, Ito H, Kanbe A, Ando T, Hara A,
Shimizu M, Moriwaki H and Seishima M: The inhibition of indoleamine
2,3-dioxygenase accelerates early liver regeneration in mice after
partial hepatectomy. Dig Dis Sci. 62:2386–2396. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Prendergast GC, Metz R, Muller AJ, Merlo
LMF and Mandik-Nayak L: IDO2 in immunomodulation and autoimmune
disease. Front Immunol. 5:5852014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fujii K, Yamamoto Y, Mizutani Y, Saito K
and Seishima M: Indoleamine 2,3-dioxygenase 2 deficiency
exacerbates imiquimod-induced psoriasis-like skin inflammation. Int
J Mol Sci. 21:55152020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yamamoto Y, Yamasuge W, Imai S, Kunisawa
K, Hoshi M, Fujigaki H, Mouri A, Nabeshima T and Saito K:
Lipopolysaccharide shock reveals the immune function of indoleamine
2,3-dioxygenase 2 through the regulation of IL-6/stat3 signalling.
Sci Rep. 8:159172018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hoshi M, Osawa Y, Nakamoto K, Morita N,
Yamamoto Y, Ando T, Tashita C, Nabeshima T and Saito K: Kynurenine
produced by indoleamine 2,3-dioxygenase 2 exacerbates acute liver
injury by carbon tetrachloride in mice. Toxicology. 438:1524582020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ito H, Ando K, Nakayama T, Taniguchi M,
Ezaki T, Saito K, Takemura M, Sekikawa K, Imawari M, Seishima M and
Moriwaki H: Role of Valpha 14 NKT cells in the development of
impaired liver regeneration In vivo. Hepatology. 38:1116–1124.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ando T, Ito H, Kanbe A, Hara A and
Seishima M: Deficiency of NALP3 signaling impairs liver
regeneration after partial hepatectomy. Inflammation. 40:1717–1725.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Percie du Sert N, Hurst V, Ahluwalia A,
Alam S, Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl
U, et al: The ARRIVE guidelines 2.0: Updated guidelines for
reporting animal research. PLoS Biol. 18:e30004102020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vogel CFA and Matsumura F: A new
cross-talk between the aryl hydrocarbon receptor and RelB, a member
of the NF-kappaB family. Biochem Pharmacol. 77:734–745. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gutiérrez-Vázquez C and Quintana FJ:
Regulation of the immune response by the Aryl hydrocarbon receptor.
Immunity. 48:19–33. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fallarino F, Grohmann U and Puccetti P:
Indoleamine 2,3-dioxygenase: From catalyst to signaling function.
Eur J Immunol. 42:1932–1937. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Merlo LMF, Peng W, DuHadaway JB,
Montgomery JD, Prendergast GC, Muller AJ and Mandik-Nayak L: The
immunomodulatory enzyme IDO2 mediates autoimmune arthritis through
a nonenzymatic mechanism. J Immunol. 208:571–581. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo MC, Zhou SY, Feng DY, Xiao J, Li WY,
Xu CD, Wang HY and Zhou T: Runt-related transcription factor 1
(RUNX1) binds to p50 in macrophages and enhances TLR4-triggered
inflammation and septic shock. J Biol Chem. 291:22011–22020. 2016.
View Article : Google Scholar : PubMed/NCBI
|