Introduction
Non-alcoholic fatty liver disease (NAFLD) is a
condition where the accumulation of lipids exceeds 5% of
hepatocytes and is not generated by alcohol, drug consumption or
does not damage hepatocytes (1).
The global prevalence of NAFLD is increasing, with ~20–30% of
patients presenting with early-stage disease (2,3).
This disease is currently of great concern as it may increase the
risk of developing other subsequent anomalies, as for example type
2 diabetes mellitus (T2DM) and cardiovascular disease (CVD)
(4).
It has been revealed that de novo lipogenesis
(DNL) may be crucial for the development of NAFLD (5). It occurs primarily in hepatocytes
and is triggered mainly by a high intake of glucose or fructose.
DNL turns excessive glucose or fructose into fatty acid and
triglycerides (6). DNL is a
normal process for the maintenance of homeostasis in the body, and
its increased activation may potentially cause hepatic steatosis
(7). Therefore, the inhibition of
DNL is highly pursued as a therapeutic target for lipid
metabolism-related disease.
The sterol regulatory element-binding protein 1c
(SREBP1c) and carbohydrate response element-binding protein
(ChREBP) are key transcription factors that play a crucial role in
DNL (8). Several studies have
revealed that SREBP1c and ChREBP increase the expression of
lipogenic enzymes related to DNL (9–11).
The simultaneous activity of SREBP1c and ChREBP is a normal process
for the maintenance of cell homeostasis; however, at excessive
levels, the cell has a specific mechanism to terminate the
signalling activation. Several proteins are responsible for
reducing DNL, including AMP-activated protein kinase (AMPK)
(12).
AMPK regulates DNL through several mechanisms,
phosphorylating and inactivating acetyl-CoA carboxylase (ACC), thus
inhibiting fatty acid biosynthesis (13). Furthermore, AMPK also inhibits
transcriptional regulators, including SREBP1c and ChREBP. The
activation of AMPK has been reported to be blocked the nuclear
translocation of SREBP1c and attenuates aberrant lipogenesis in
diabetic mice (14). In another
study on 3T3-L1 cells, AMPK was revealed to phosphorylate the
precursor of SREBP1c and prevented the conversion of SREBP1c into
its mature form (15). It also
regulates the activity of ChREBP, as demonstrated in an
ethanol-induced fatty liver experiment, where AMPK was inhibited by
ethanol, while ChREBP activity increased significantly (16). Therefore, AMPK is considered one
of the proteins that can maintain cell balance, specifically
concerning lipid metabolism.
Sirtuin 1 (SIRT1) is also well-known as a regulatory
protein (17). Several studies
have reported the activation of SIRT1 in lowering the expression of
DNL enzymes (18,19). Furthermore, the increased activity
of SIRT1 decreases the expression of SREBP1c, while the knockout
SIRT1 has been reported to elevated the expression of ChREBP in
HepG2 cells (19). This
demonstrates the importance of SIRT1 in the regulation of lipid
metabolism, specifically in DNL.
The effects of AMPK and SIRT1 activation on lipid
metabolism are well known; however, there are still concerns as to
whether the combination of their activators is beneficial for
pathological lipid metabolism-related diseases, including fatty
liver disease. Therefore, the present review article aimed to
summarize the role of AMPK and SIRT1 in NAFLD, based on evidence
obtained from randomized control studies.
Data collection methods
The present review summarizes the result of
randomized control studies related to the effect of AMPK and SIRT1
activators on NAFLD. Articles were obtained from the PubMed
database identified using the key words ‘SIRT1 activator AND
NAFLD’, ‘Resveratrol AND fatty liver’, ‘AMPK activator AND NAFLD’,
as well as ‘Metformin AND fatty liver’. Only clinical or randomized
control trial articles published over the last 10 years were
included. By contrast, articles that did not include SIRT1 and AMPK
activators in patients with NAFLD were excluded. The method used
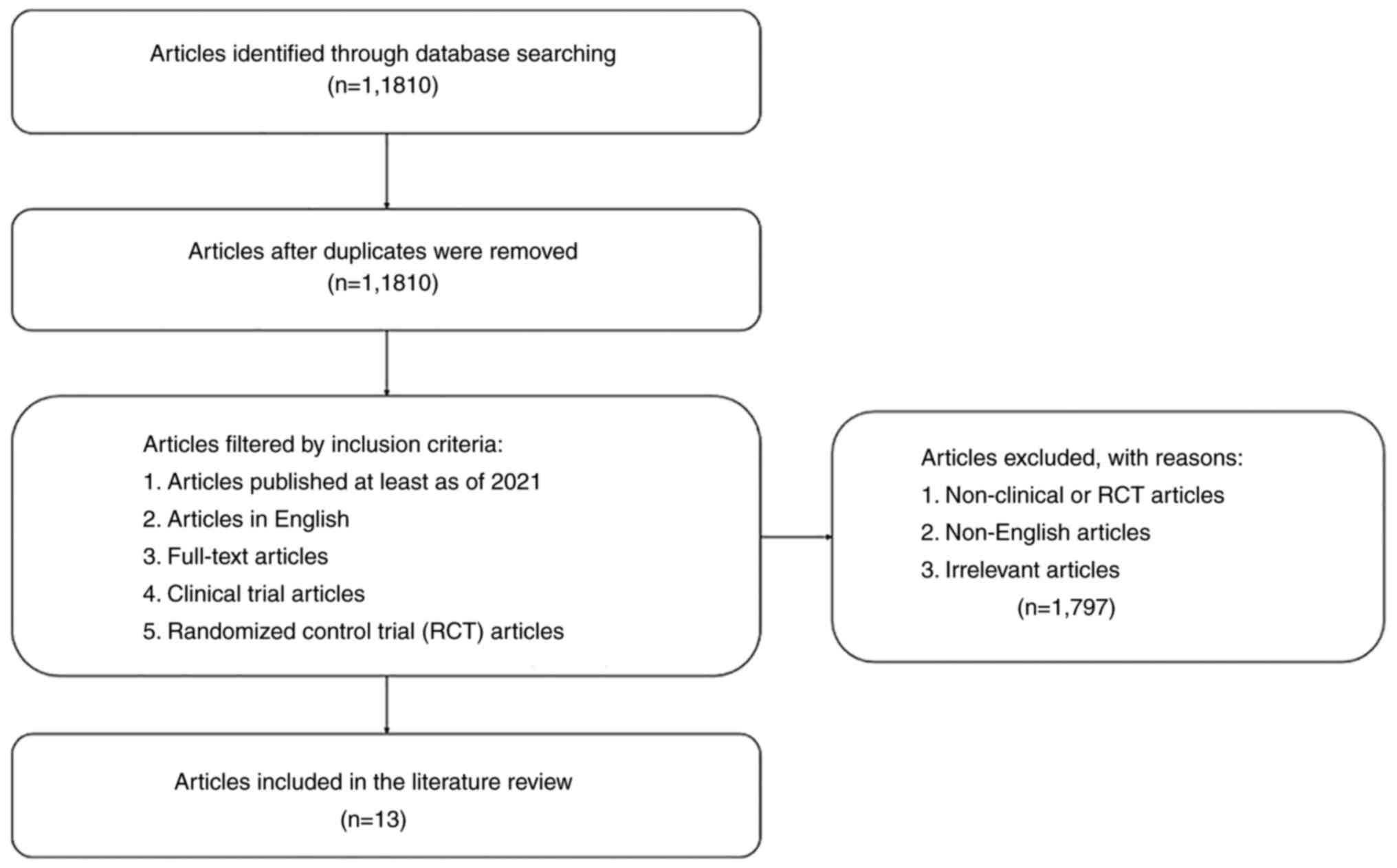
for data collection is summarized in Fig. 1. In total, 13 articles were
collected, and the data are presented in Table I, arranged by the protein, its
activator name, subject, treatment, duration, type of study,
outcome and references, and the results of these studies were then
discussed.
 | Table I.Clinical trials of AMPK and SIRT1
activators on patients with NAFLD. |
Table I.
Clinical trials of AMPK and SIRT1
activators on patients with NAFLD.
| Protein | Activator | Subjects | Treatment and
duration of study | Type of study | Outcome | (Refs.) |
|---|
| AMPK | Metformin | 173 children with
NAFLD | 500 mg twice a day
for 96 weeks | Randomized,
placebo-controlled, double blind |
| (57) |
|
| Metformin combined
with N-acetylcysteine | 53 patients with
NAFLD | 850-1,500 mg/day
for 48 weeks | Open-label
multicenter randomized trial |
| (58) |
|
| PXL770 | 12 patients with
NAFLD |
| Randomized,
double-blind, placebo |
| (65) |
|
| Metformin | 63 patients with
NAFLD | 500 mg metformin
once a day for 4–12 month | Randomized,
placebo-controlled |
| (68) |
|
| Metformin | 10 patients at a
risk of developing NAFLD | 500 mg once a day
for 12 weeks | Single center, open
label trial |
| (69) |
|
| Metformin | 35 patients with
NAFLD | 850 mg daily for 24
weeks | Prospective
controlled trial |
| (70) |
|
| Metformin | 29 patients with
type 2 diabetes and NAFLD |
| Single center,
open-label, prospective, randomized trial |
| (71) |
| SIRT1 | Resveratrol | 50 patients with
NAFLD | 500 mg once a day
for 12 weeks | Randomized,
placebo-controlled, double blind |
-
– Reduced steatosis
grade
-
– Reduce level of ALT
and AST
-
– Reduced inflammatory
markers
| (75) |
|
| Resveratrol | 60 patients with
NAFLD | 150 mg Twice a day
for 3 months | Randomized,
placebo-controlled, double blind |
-
– Decreased ALT, AST,
LDL-c, glucose level
-
– Reduction of
adiponectin level and TNF-α
| (76) |
|
| Resveratrol | 25 patients with
NAFLD | 500 mg once a day
for 12 weeks | Randomized,
placebo-controlled, double blind |
| (48) |
|
| Resveratrol | 28 patients with
NAFLD | 1,5 g daily for 6
months | Randomized,
placebo-controlled, double blind |
| (49) |
|
| Resveratrol | 44 patients with
NAFLD | 50 mg and 200 mg
once a day for 6 months | Randomized |
| (86) |
| AMPK and SIRT1 | Metformin + leucine
+ sildenafil (NS-0200) | 91 patients with
NAFLD |
| Randomized,
placebo-controlled, double blind |
| (103) |
De novo lipogenesis
DNL is considered the primary factor in the
development of fatty liver disease (7). In a pathological condition, such as
NAFLD, DNL activation increases, generating excessive fat and
culminating in intrahepatic lipid accumulation (5,20).
Furthermore, DNL is a biosynthetic pathway for the productions of
fatty acids and triglycerides from a non-lipid source, triggered by
a high presence of carbohydrates or by insulin receptor-mediated
signalling. The pathway is highly regulated by two significant
factors, namely transcriptional regulation of DNL enzyme and
allosteric regulation of ACC (21).
The transcriptional regulation of the DNL enzyme
includes two transcription factor proteins, namely SREBP1c and
ChREBP (Fig. 2). The influx of
glucose and the signalling from insulin induce the activation of
ChREBP and SREBP1c, respectively. Under basal conditions, the
binding of SREBP1c to SREBP cleavage-activating protein (SCAP) and
insulin-induced gene 1 (INSIG1) protein on the endoplasmic
reticulum, prevents its translocation to the nucleus (8). Subsequently, INSIG1 is dissociated
via the phosphorylation of SREBP1c and SCAP is cleaved by S1 and S2
proteases in the Golgi apparatus, and eventually, SREBP1c
expresssion is released (8).
Additionally, ChREBP is anchored by 14-3-3 protein and the
phosphorylation of this complex permits the free ChREBP entry the
nucleus (22). Furthermore,
SREBP1c and ChREBP bind to the promoter gene target in the nucleus
and start the transcription of lipogenic genes, including fatty
acid synthase (FAS), stearoyl-CoA desaturase 1 (SCD1) and ACC
(23).
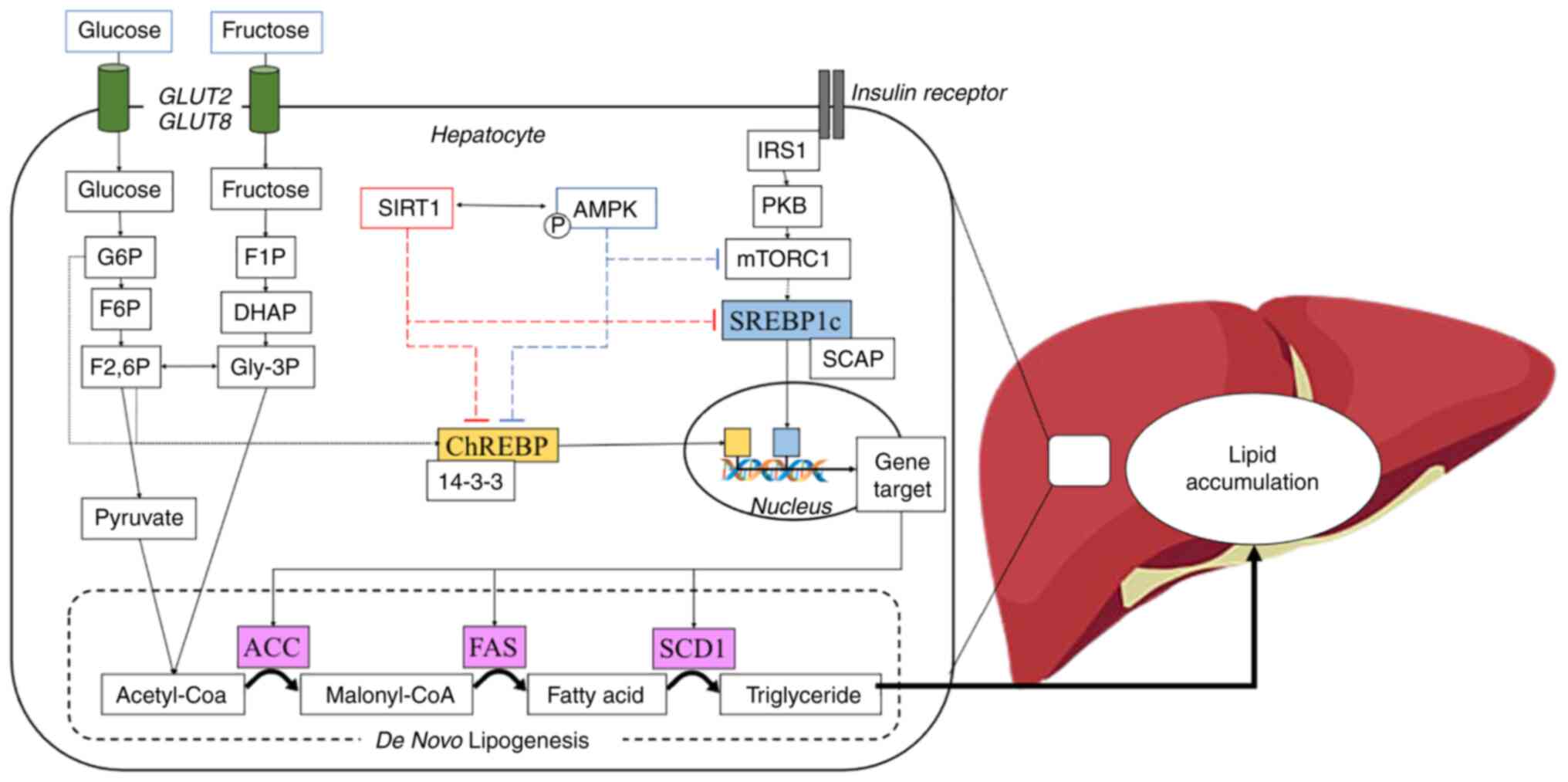 | Figure 2.Schematic overview of DNL and its
transcriptional regulation. Carbohydrates, including glucose or
fructose enter hepatocyte cells and become a sensor for DNL
activation. Glucose is converted to G6P followed by isomerization
to F6P and F2,6P through the glycolysis process. By contrast,
fructose also converts to Gly-3P through fructolysis and further
converts to F2,6P. G6P and F2,6P induce dephosphorylation of
ChREBP, and it detaches from 14-3-3 protein into an active form.
Moreover, the activation of insulin receptor leads to the
phosphorylation of IRS1, further activating the mTORC pathway and
induces the nuclear translocation of SREBP1c. In the feedback
response, SIRT1 and AMPK prevent the nuclear translocation of
ChREBP and SREBP1c, resulting in the inhibition of DNL
transcriptional regulation. DNL, de novo lipogenesis; G6P,
glucose 6-phosphate; F6P, fructose 6-phosphate; Gly-3P, glycerol
3-phosphate; F2,6P, fructose 2,6-bisphosphate; ChREBP, carbohydrate
response element-binding protein; IRS1, insulin receptor substrate
1; mTORC, mammalian target of rapamycin complex; SREBP1c, sterol
regulatory element-binding protein 1c; SIRT1, Sirtuin 1; AMPK,
AMP-activated protein kinase. |
The inhibition of SREBP1c and ChREBP reduces the
production of lipogenic genes as well as lipogenesis (24,25). Several proteins such as AMPK have
been reported to inhibit the activity of SREBP1c and ChREBP.
Another possible inhibitory mechanism of AMPK is predicted through
SIRT1 which reportedly blocked both SREBP1c and ChREBP (19,23,26).
AMPK
The body has a system to maintain energy balance, in
the form of adenosine triphosphate (ATP). When cellular ATP levels
are reduced, the AMPK pathway is activated, phosphorylating the
growth-regulating enzymes along with proteins, in order to generate
ATP and decrease ATP consumption (27). AMPK is considered the master
regulator of numerous proteins responsible for aging, inflammation,
redox and the metabolism of lipids and glucose (28).
Based on the crystal structure of the protein, AMPK
is a trimeric complex, consisting of a catalytic α subunit and two
regulatory subunits, namely β and γ. The α subunit contains a
kinase domain and an important residue (Thr172), which is
phosphorylated by upstream kinases. The β subunit contains a
binding site for carbohydrates that causes AMPK to associate with
glycogen. Additionally, the γ subunit acts as a sensor for changes
in the AMP/ADP ratio (29). When
AMP increases and ADP decreases, AMP binds to the γ subunit,
activating AMPK through three mechanisms, namely: i) The
phosphorylation of Thr172 by stimulating the upstream proteins or
stabilizing AMPK into a substrate more susceptible to
phosphorylation; ii) AMP prevents the dephosphorylation by the
phosphatase on Thr172; and iii) AMP causes allosteric activation of
Thr172 in the α sub-unit (30,31). The major upstream kinase of AMPK
is liver kinase B1 (LKB1) and Ca2+/calmodulin-dependent
protein (CaMKK) which phosphorylates AMPK in Thr172 (32,33). LKB1 is the main upstream activator
of AMPK. It is activated by the stress signal or by the presence of
activators, including aminoimidazole-4-carboxamide ribonucleoside
and metformin (34). In addition,
CaMKK is highly distributed in neural tissue to respond to neuronal
depolarization (35).
AMPK is known to play an essential role in various
metabolic-related diseases, such as NAFLD. Its activity causes the
inhibition of DNL through the suppression of SREBP1c and ChREBP.
AMPK inhibits the activation of SREBP1c through the phosphorylation
at Ser372 residue and prevents the cleavage process by protease
(14). Furthermore, a recent
study demonstrated that it suppresses SREBP1c expression through
the mTOR and LXRa proteins (36).
ChREBP is also phosphorylated at the Ser568 residue by AMPK,
causing re-binding to 14-3-3 protein and the subsequent conversion
into an inactive form as well as preventing lipid synthesis
(22,36).
SIRT1
SIRT1 is a class III family of histone deacetylases,
and their reactions require nicotinamide adenine (NAD+)
to concurrently deacetylate histones and non-histone from proteins
involved in metabolic processes and stress responses (17,37). It is widely expressed in mammalian
cells in a number of organs, including the brain, adipose tissue,
kidneys, pancreas, endothelium, spleen, skeletal muscle and liver.
Furthermore, its expression is known to be involved in several
diseases, including metabolic diseases and age-related diseases, as
well as CVD (38). SIRT1 is a
protein that regulates metabolism, including fat cell accumulation
and maturation, lipid metabolism in the liver, systemic
inflammation, nutrition sensing and circadian rhythms (39). Previous studies have demonstrated
that SIRT1 inhibits DNL enzymes, as well as their key regulator
proteins, SREBP1c and ChREBP, culminating in abolishing
perturbation of hepatic lipid metabolism (19,40).
The primary function of SIRT1 is to deacetylate the
acetyl-lysine residue of histone substrate or non-histone proteins,
including transcription factors, co-regulators and enzymes
(41,42). Therefore, SIRT1 has multiple
physiological functions, particularly in metabolism. It has been
characterized as the ‘master of metabolic regulators’, due to its
pivotal role in maintaining the homeostasis of lipid metabolism by
affecting several proteins involved. SREBP1c is a critical
transcription factor that initiates several lipogenic genes,
inducing lipogenesis within the cell. SIRT1 inhibits SREBP1c
activity and decreases lipogenesis in mouse liver (19). Another lipogenesis inducer aside
SREBP1c and ChREBP is SIRT1 (43). Furthermore, AMPK, which is the
natural regulator of ChREBP and SREBP1c, is also affected by SIRT1
activity through an indirect mechanism by deacetylating the
upstream kinase of AMPK, LKB1 (18,44). This demonstrates that SIRT1 plays
a prominent role in the development of lipid-related diseases,
including non-alcoholic liver disease. This is in line with several
studies demonstrating that SIRT1 activator alleviates fatty liver
in rodent models and NAFLD patients (45–49).
A previous in silico study revealed that the
crystal structure of SIRT1 is composed of the following three major
domains: the catalytic, N-terminal, and C-terminal (50). The catalytic region consists of
the binding site of substrate and NAD+ that promotes the
deacetylation of lysine, whereas the N- and C-terminals bind to
several compounds such as resveratrol, suramin, or EX-527 and
regulate SIRT1 deacetylase activity (51). Inside the cell, SIRT1 is localized
in the cytoplasm and affects other proteins, including NF-κB,
peroxisome proliferator-activated receptor γ, peroxisome
proliferator-activated receptor-γ coactivator, AMPK and p-53
(52,53), while in the nucleus, it affects
the translocation of proteins, including FOXO3a and several
antioxidant genes such as SOD2/3, HO-1, and NQO-1 (54).
AMPK activators in NAFLD clinical
studies
Evidence supports the role of AMPK in
metabolism-related diseases, such as NALFD (55). AMPK regulates other proteins and
provides homeostasis within the cell through several mechanisms
involved in lipid metabolism, glucose metabolism, protein
metabolism, autopaghy, and mitochondrial biogenesis (27,55,56). It has been well-established that
AMPK is involved in the prevention of hepatic steatosis. Metformin,
an indirect activator of AMPK, has been widely studied for its
effects on NAFLD. Several clinical trials have reported the
beneficial effects of metformin on certain features of NAFLD. A
previous randomized control trial on children diagnosed with NAFLD
and treated with metformin at 500 mg twice per day for 24 months,
reported an improvement in steatosis grade and lipid profiles
(57). Moreover, an open-label,
multi-centred, randomized trial, reported that metformin in
combination with acetylcysteine administered for 12 months led to
the significant improvement in the NAFLD Activity Score measured by
liver biopsies of adult patients with NAFLD (58). Acetylcysteine provides a potent
antioxidant effect on the liver, thereby protecting the liver from
oxidative stress (59,60). AMPK activity also affects the
antioxidant defense system in cells (61,62). A combination of AMPK activator and
antioxidant such as acetylcysteine yielded a positive impact
against hepatic steatosis (58).
AMPK activation through metformin exerts a beneficial effect by
reducing hepatic steatosis in patients with NAFLD (57,58). Another study with a direct AMPK
activator, PXL770, supports this statement. The mechanisms of
action of metformin and PXL770 as activators of AMPK are summarized
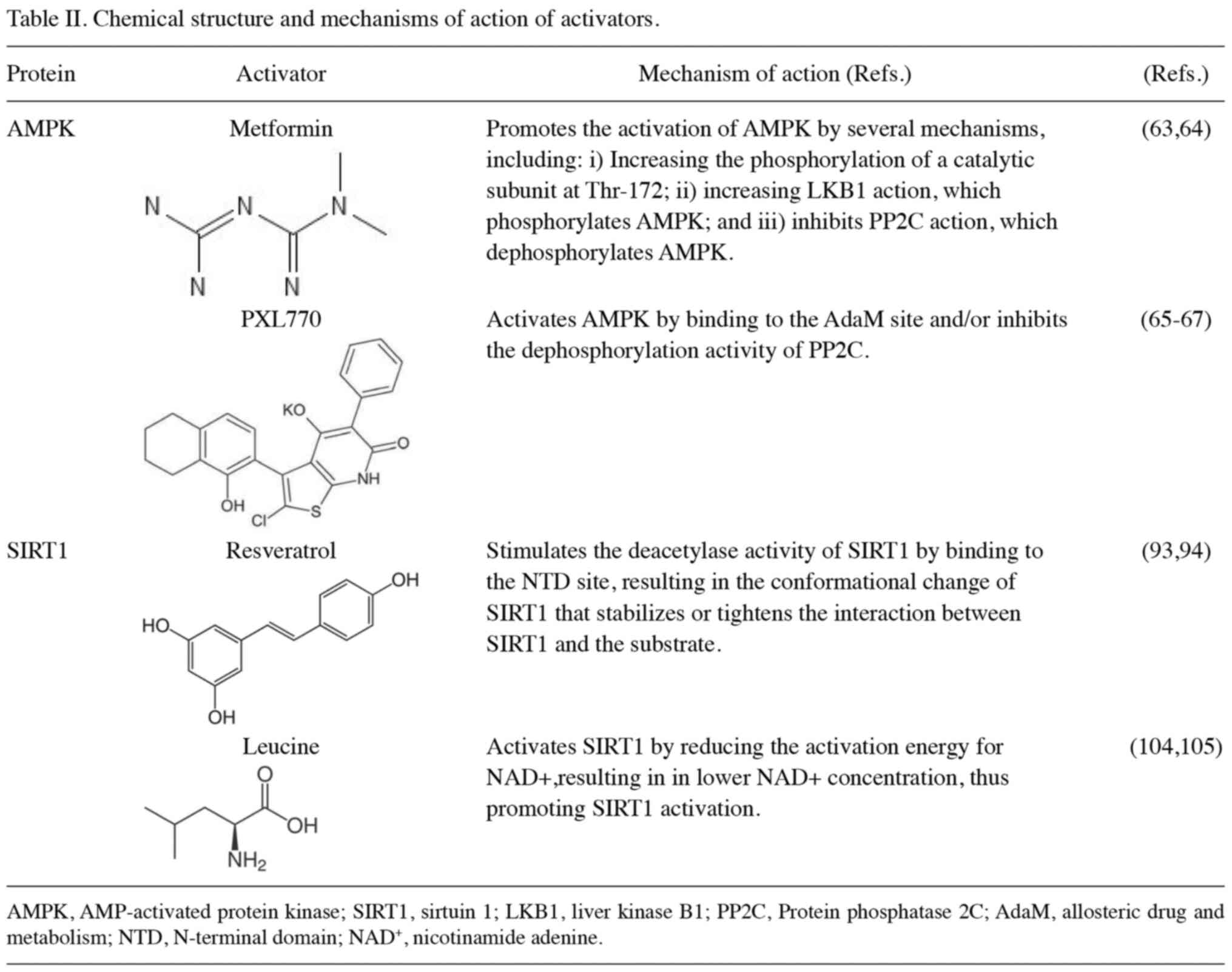
in Table II (58–67). A randomized, double-blind,
placebo-controlled trial reported that treatment using PXL770 for
12 weeks decreased DNL percentage and improved glucose metabolism.
Lipid profiles concerning triglycerides and very-low-density
lipoprotein (VLDL) decreased in the PXL770 group compared to the
placebo group (65). Furthermore,
AMPK activation, direct or indirect, has a beneficial effect by
reducing steatosis in patients with NAFLD.
Metformin also has a beneficial effect on the lipid
profiles of patients with NAFLD, according to a previous trial,
where 500 mg metformin administered for 4 months significantly
decreased liver enzyme and triglyceride levels, and increased
high-density lipoprotein (HDL)-cholesterol levels in patients
(68). This is in line with
another study which revealed that 500 mg metformin administered for
3 months decreased VLDL and triglyceride levels in 10 patients who
were at a risk of developing NAFLD (69). Another study similarly reported
that the daily administration of 850 mg metformin for 6 months
reduced liver enzyme, total cholesterol and triglyceride levels,
and increased HDL-cholesterol levels (70). Furthermore, in children diagnosed
with NAFLD, treatment with metformin 500 mg twice per day, for 24
months, led to a beneficial effect in the form of improvement in
lipid profiles (57). Different
doses of metformin, including 250 mg three times per day, 500 mg
three times per day, and 1,000 mg twice per day, administered for 6
months, have been shown to produce similar results, namely an
improvement in liver enzyme levels and lipid profiles in patients
with T2DM and NAFLD (71). Lipid
profiles are greatly influenced by metformin at various doses in
children and adult patients.
AMPK activation is involved in several mechanisms in
lipid metabolism. A previous study revealed that the activation of
AMPK decreased SREBP1c activity in mice fed a high-fat diet,
thereby attenuating hepatic steatosis (14). SREB1c regulates the protein that
is crucial for lipid and glucose metabolism. AMPK activation has
been reported to inhibit fat-forming enzymes, including ACC, FAS
and SCD1 through SREBP1 inhibition, leading to decreased
intracellular fat accumulation (15). Another possible mechanism is
through the inhibition of 6-phosphogluconate dehydrogenase (6PGD),
which is an enzyme involved in glycolysis. A previous in
vitro study demonstrated that the inhibition of 6PGD activated
the AMPK pathway and reduced ACC1 activity, thereby inhibiting
lipid biosynthesis (72). 6PGD is
the third enzyme in the pentose phosphate pathway (PPP) which is
responsible for converting the 6-phosphogluconate into ribulose
5-phosphate (R-5-P). The upregulation of R-5-P frequently
antagonizes the LKB1 complex, resulting in the decrease of AMPK
activity. Another protein involved in this mechanism is mammalian
target of rapamycin complex 1 (mTORC), which is the upstream
protein target of SREBP1c. In the cancer cell, activation of mTORC
may upregulate the PPP through SREBP1c (73). It is well known that AMPK activity
inhibits mTORC; therefore, it may also alter the PPP, resulting in
the reduction of the lipogenesis. PPP may be a critical pathway in
lipogenesis. In a recent study on cancer cells, metformin was
reported to interfere with several enzymes related to PPP and
decreased the effect of PPP via modulation of mTORC (74). However, the information about the
association between metformin and 6PGD remains unclear (74). Briefly, in vitro, in vivo,
or clinical trials have provided evidence that AMPK activation may
be a critical step in improving lipid metabolism.
The SIRT1 activator, resveratrol, in NAFLD
clinical studies
In recent years, the use of resveratrol as a therapy
for certain diseases has attracted increasing attention, due to its
beneficial effects in reducing insulin resistance, the risk of CVD,
hyperlipidemia, obesity and fatty liver-related diseases, such as
NAFLD. Several clinical studies have demonstrated the beneficial
effects of resveratrol in patients with NAFLD (Table I). A previous randomized,
placebo-controlled, double-blinded study on 50 patients with NAFLD
treated with 500 mg resveratrol daily for 3 months indicated an
improvement in anthropometric measurements (weight, body mass
index, waist circumference), liver enzyme levels, inflammatory
marker levels and liver steatosis compared to the placebo group. It
was proven that liver steatosis and inflammatory cytokines,
including TNF-α, IL-6 and NF-κB were reduced by resveratrol for the
activation of SIRT1 (75).
Another randomized trial also reported that a lower dose of
resveratrol (150 mg/day) for 3 months reduced the TNF-α level in
patients (76). SIRT1 activation
in hepatocytes in steatosis is associated with the inflammation
system, preventing further hepatocyte damage (77).
Inflammation and oxidative stress have been widely
reported in hepatic steatosis, due to elevated lipid peroxidation
and free radical production, eventually leading to cell damage or
dysfunction (78–80). A previous study on mice revealed
that resveratrol inhibited the activity of NF-κB and TNF-α
(81). The inhibition of SIRT1
expression can lead to an increase in inflammatory cytokine levels.
Moreover, SIRT1 activation induces nuclear factor erythroid
2-related factor 2 activity, thereby providing a protective effect
through the antioxidant defense system of the cell (82,83). Other pre-clinical studies have
reported that resveratrol ameliorates high-fat diet induced fatty
liver disease, culminating in decreased triglyceride levels
(84,85). These studies generally confirm
that SIRT1 activation may inhibit fatty liver and improve the
inflammation condition in hepatic steatosis both in animals and
humans.
In several clinical trials, SIRT1 activation by
resveratrol at different doses has been shown to lead to a decrease
in lipid content. A double-blind, randomized, placebo-controlled
trial with 60 participants with NAFLD treated with 150 mg
resveratrol, twice per day, for 3 months, reported a significant
decrease in liver enzyme, total cholesterol and low density
lipoprotein (LDL)-cholesterol levels, and homeostatic model
assessment for insulin resistance (HOMA-IR) compared to the placebo
group (76). According to a
previous study, lower doses of resveratrol, such as 150 mg reduced
the intrahepatic lipid content (47). By contrast, a randomized control
trial failed to show the beneficial role of resveratrol in glucose
metabolism and lipid profile in higher doses, but not in the
steatosis level (48). The lipid
profile comprising triglycerides, LDL-cholesterol, total
cholesterol and HDL, as well as HOMA-IR did not differ not
significantly between the cohort treated with 500 mg resveratrol
for 3 months and the placebo group. However, this trial
demonstrated a significant reduction in hepatic steatosis grade and
also liver enzyme, indicating the beneficial effect of resveratrol
for steatosis patients (48).
Another randomized control trial reported a 3.8% lipid content
reduction in patients with NAFLD treated with high doses of
resveratrol 1.5 g daily, for 6 months (49). Concerning lower daily doses of 50
and 200 mg for 6 months, a lower triglyceride and LDL level in
patients with NAFLD has also been observed (86).
In a previous animal study, resveratrol demonstrated
an undoubtedly beneficial effect on lipid metabolism (87). Lipid levels, including
triglycerides, LDL-cholesterol and total cholesterol are
significantly depleted in mice with hepatic steatosis treated with
resveratrol (86–88). Additionally, it may also improve
glucose metabolism (81,84,89) and reduce the hepatic steatosis
score in high-fat/carbohydrate-induced NAFLD rats (90,91). Moreover, a previous study revealed
that the overexpression of SIRT1 culminated in the alleviation of
high-fat diet-induced hepatic steatosis and glucose intolerance in
mice (42). Another pre-clinical
study reported that mice lacking SIRT1 expression in the liver had
hepatic steatosis accompanied by elevated AST levels (92). These studies prove that SIRT1
activity improves lipid and glucose metabolism in NAFLD animal
models and in vitro study.
Several mechanisms have been proposed for
resveratrol in the treatment of NAFLD. The proposed mechanisms of
SIRT1 activators are summarized in Table II (93,94). As a direct SIRT1 activator,
resveratrol is crucial for lipid metabolism (95,96). The activation of SIRT1 inhibits
SREBP1c activity, thereby preventing lipogenesis (19). SIRT1 also inhibits the activity of
lipogenesis enzymes, including ACC and FAS (81). In an indirect mechanism, it
activates the AMPK pathway to amplify the effect of AMPK on
maintaining the homeostasis of lipid metabolism (97,98). In general, SIRT1 has been proven,
in clinical investigations except from in vitro and in
vivo studies, to possess a crucial role in improving fatty
liver conditions.
The combination of AMPK and SIRT1
activation
AMPK and SIRT1 interact with each other, affecting
lipid metabolism. It has been previously reviewed that AMPK and
SIRT1 simultaneously function, in order to regulate other proteins
(99). A combination of
resveratrol and metformin decreased glucose and triglyceride levels
as well as improved liver function in diabetic mice (100). A previous study also reported
that a similar combination reduced liver weight and visceral fat in
mice (100). Furthermore, the
concurrent activation of AMPK and SIRT1 pathways contributes to
decreasing lipogenesis, thereby alleviating hepatic steatosis in
mice with NAFLD (101,102).
A previous randomized control trial of 91
participants with NAFLD reported that a combination of leucine and
metformin given daily for 16 weeks culminated in decreased hepatic
fat and a significantly increased fatty acid oxidation compared to
the placebo group (103).
L-leucine directly activates SIRT1 through allosteric interaction
in an in vitro study. Its mechanism of action as an
activator of SIRT1 is also summarized in Table II (104,105). Furthermore, leucine also affects
AMPK activity (106,107). The combination of leucine and
metformin produced a beneficial effect related to NAFLD features.
Several studies have also demonstrated that the activation of AMPK
and SIRT1 plays a principal role to improve NAFLD features
(15,46–48,57,67). However, clinical trials that adopt
the combination of AMPK and SIRT1 activators are still lacking;
hence, further research on the combinatory use of these activators
is required, in order to elucidate a strong correlation between
AMPK and SIRT1 in lipid metabolism.
Conclusions
The existing data indicated that SIRT1 and AMPK
might have a pivotal role in the pathogenesis of NAFLD. Both of
Activators of SIRT1 and activators of AMPK, produce a benefit in
preventing lipogenesis, thus reduce the impact of fatty liver.
Several randomized control trials have proven that treatment using
SIRT1 and AMPK activators in patients with NAFLD can improve
hepatic steatosis, prevent inflammation, and inhibit lipogenesis.
However, further studies are warranted for the confirmation of the
effects of SIRT1 and AMPK activator alone or in combination for the
treatment of fatty liver-related diseases. The present review
demonstrates that SIRT1 and AMPK activators are promising
therapeutics for treating NAFLD.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Doctoral Research Grant
(PDD) 2022 provided by the Indonesian Ministry of Education and
Culture (Grant nos. 094/E5/PG.02.00.PT/2022 and
044/E5/RA.02.00.PM/2022).
Availability of data and materials
Not applicable.
Authors' contributions
SAS, HK and JL were involved in the conception and
design of the study. PA principally collected previously published
studies and wrote the original draft of the manuscript. HK and SAS
were responsible for the acquisition of the collected articles. JL
critically revised the article for intellectual content. All
authors have read and approved the final manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tiniakos DG, Anstee QM and Burt AD: Fatty
liver disease. MacSween's Pathol Liver. 7th edition. Elsevier;
Philadephia, PA: pp. 308–371. 2018, View Article : Google Scholar
|
|
2
|
Iqbal U, Perumpail B, Akhtar D, Kim D and
Ahmed A: The epidemiology, risk profiling and diagnostic challenges
of nonalcoholic fatty liver disease. Medicines (Basel). 6:412019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sayiner M, Koenig A, Henry L and Younossi
ZM: Epidemiology of nonalcoholic fatty liver disease and
nonalcoholic steatohepatitis in the United States and the rest of
the world. Clin Liver Dis. 20:205–214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Byrne CD and Targher G: NAFLD: A
multisystem disease. J Hepatol. 62:S47–S64. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Buzzetti E, Pinzani M and Tsochatzis EA:
The multiple-hit pathogenesis of non-alcoholic fatty liver disease
(NAFLD). Metabolism. 65:1038–1048. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ter Horst KW and Serlie MJ: Fructose
consumption, lipogenesis, and non-alcoholic fatty liver disease.
Nutrients. 9:9812017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Knebel B, Fahlbusch P, Dille M, Wahlers N,
Hartwig S, Jacob S, Kettel U, Schiller M, Herebian D, Koellmer C,
et al: Fatty liver due to increased de novo lipogenesis:
Alterations in the hepatic peroxisomal proteome. Front Cell Dev
Biol. 7:2482019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ferré P and Foufelle F: Hepatic steatosis:
A role for de novo lipogenesis and the transcription factor
SREBP-1c. Diabetes Obes Metab. 12:83–92. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Witte N, Muenzner M, Rietscher J, Knauer
M, Heidenreich S, Nuotio-Antar AM, Graef FA, Fedders R, Tolkachov
A, Goehring I and Schupp M: The glucose sensor ChREBP links de novo
lipogenesis to PPARγactivity and adipocyte differentiation.
Endocrinol. 156:4008–4019. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vijayakumar A, Aryal P, Wen J, Syed I,
Vazirani RP, Moraes-Vieira PM, Camporez JP, Gallop MR, Perry RJ,
Peroni OD, et al: Absence of carbohydrate response element binding
protein in adipocytes causes systemic insulin resistance and
impairs glucose transport. Cell Rep. 21:1021–1035. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stoeckman AK and Towle HC: The role of
SREBP-1c in nutritional regulation of lipogenic enzyme gene
expression. J Biol Chem. 277:27029–27035. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
von Loeffelholz C, Coldewey SM and
Birkenfeld AL: A narrative review on the role of ampk on de novo
lipogenesis in non-alcoholic fatty liver disease: Evidence from
human studies. Cells. 10:18222021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Viollet B, Foretz M, Guigas B, Horman S,
Dentin R, Bertrand L, Hue L and Andreelli F: Activation of
AMP-activated protein kinase in the liver: A new strategy for the
management of metabolic hepatic disorders. J Physiol. 574:41–53.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Xu S, Mihaylova MM, Zheng B, Hou X,
Jiang B, Park O, Luo Z, Lefai E, Shyy JYJ, et al: AMPK
phosphorylates and inhibits SREBP activity to attenuate hepatic
steatosis and atherosclerosis in diet-induced insulin-resistant
mice. Cell Metab. 13:376–388. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ha JH, Jang J, Chung SI and Yoon Y: AMPK
and SREBP-1c mediate the anti-adipogenic effect of
β-hydroxyisovalerylshikonin. Int J Mol Med. 37:816–824. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liangpunsakul S, Ross RA and Crabb DW:
Activation of carbohydrate response element binding protein by
ethanol. J Investig Med. 61:270–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cantó C and Auwerx J: Targeting sirtuin 1
to improve metabolism: All you need is NAD +? Pharmacol Rev.
64:166–187. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hou X, Xu S, Maitland-Toolan KA, Sato K,
Jiang B, Ido Y, Lan F, Walsh K, Wierzbicki M, Verbeuren TJ, et al:
SIRT1 regulates hepatocyte lipid metabolism through activating
AMP-activated protein kinase. J Biol Chem. 283:20015–20026. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ponugoti B, Kim DH, Xiao Z, Smith Z, Miao
J, Zang M, Wu SY, Chiang CM, Veenstra TD and Kemper JK: SIRT1
deacetylates and inhibits SREBP-1C activity in regulation of
hepatic lipid metabolism. J Biol Chem. 285:33959–33970. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paglialunga S and Dehn CA: Clinical
assessment of hepatic de novo lipogenesis in non-alcoholic fatty
liver disease. Lipids Health Dis. 15:1592016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sanders FWB and Griffin JL: De novo
lipogenesis in the liver in health and disease: More than just a
shunting yard for glucose. Biol Rev. 91:452–468. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sato S, Jung H, Nakagawa T, Pawlosky R,
Takeshima T, Lee WR, Sakiyama H, Laxman S, Wynn RM, Tu BP, et al:
Metabolite regulation of nuclear localization of
carbohydrate-response element-binding protein (ChREBP): Role of amp
as an allosteric inhibitor. J Biol Chem. 291:10515–10527. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Viscarra J, Kim SJ and Sul HS:
Transcriptional regulation of hepatic lipogenesis. Nat Rev Mol Cell
Biol. 16:678–689. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dentin R, Benhamed F, Hainault I, Fauveau
V, Foufelle F, Dyck JRB, Girard J and Postic C: Liver-specific
inhibition of ChREBP improves hepatic steatosis and insulin
resistance in ob/ob mice. Diabetes. 55:2159–2170. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao X, Xiaoli, Zong H, Abdulla A, Yang
EST, Wang Q, Ji JY, Pessin JE, Das BC and Yang F: Inhibition of
SREBP transcriptional activity by a boron-containing compound
improves lipid homeostasis in diet-induced obesity. Diabetes.
63:2464–2473. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nguyen LT, Mak CH, Chen H, Zaky AA, Wong
MG, Pollock CA and Saad S: SIRT1 attenuates kidney disorders in
male offspring due to maternal high-fat diet. Nutrients.
11:1462019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Herzig S and Shaw RJ: AMPK: Guardian of
metabolism and mitochondrial homeostasis. Nat Rev Mol Cell Biol.
19:121–135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jeon SM: Regulation and function of AMPK
in physiology and diseases. Exp Mol Med. 48:e2452016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiao B, Sanders MJ, Carmena D, Bright NJ,
Haire LF, Underwood E, Patel BR, Heath RB, Walker PA, Hallen S, et
al: Structural basis of AMPK regulation by small molecule
activators. Nat Commun. 4:30172013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Suter M, Riek U, Tuerk R, Schlattner U,
Wallimann T and Neumann D: Dissecting the role of 5′-AMP for
allosteric stimulation, activation, and deactivation of
AMP-activated protein kinase. J Biol Chem. 281:32207–32216. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Oakhill JS, Steel R, Chen ZP, Scott JW,
Ling N, Tam S and Kemp BE: AMPK is a direct adenylate
charge-regulated protein kinase. Science. 332:1433–1435. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gormand A, Henriksson E, Ström K, Jensen
TE, Sakamoto K and Göransson O: Regulation of AMP-activated protein
kinase by LKB1 and CaMKK in adipocytes. J Cell Biochem.
112:1364–1375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shackelford DB and Shaw RJ: The LKB1-AMPK
pathway: Metabolism and growth control in tumour suppression. Nat
Rev Cancer. 9:563–575. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hawley SA, Boudeau J, Reid JL, Mustard KJ,
Udd L, Mäkelä TP, Alessi DR and Hardie DG: Complexes between the
LKB1 tumor suppressor, STRAD alpha/beta and MO25 alpha/beta are
upstream kinases in the AMP-activated protein kinase cascade. J
Biol. 2:282003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hawley SA, Pan DA, Mustard KJ, Ross L,
Bain J, Edelman AM, Frenguelli BG and Hardie DG:
Calmodulin-dependent protein kinase kinase-beta is an alternative
upstream kinase for AMP-activated protein kinase. Cell Metab.
2:9–19. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee GH, Peng C, Jeong SY, Park SA, Lee HY,
Hoang TH, Kim J and Chae HJ: Ginger extract controls mTOR-SREBP1-ER
stress-mitochondria dysfunction through AMPK activation in obesity
model. J Funct Foods. 87:1046282021. View Article : Google Scholar
|
|
37
|
Rahman S and Islam R: Mammalian Sirt1:
Insights on its biological functions. Cell Commun Signal. 9:112011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Elibol B and Kilic U: High levels of SIRT1
expression as a protective mechanism against disease-related
conditions. Front Endocrinol (Lausanne). 9:6142018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schug TT and Li X: Sirtuin 1 in lipid
metabolism and obesity. Ann Med. 43:198–211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang RH, Li C and Deng CX: Liver steatosis
and increased ChREBP expression in mice carrying a liver specific
SIRT1 null mutation under a normal feeding condition. Int J Biol
Sci. 6:682–690. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cantó C and Auwerx J: PGC-1alpha, SIRT1
and AMPK, an energy sensing network that controls energy
expenditure. Curr Opin Lipidol. 20:98–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Banks AS, Kon N, Knight C, Matsumoto M,
Gutiérrez-Juárez R, Rossetti L, Gu W and Accili D: SirT1 gain of
function increases energy efficiency and prevents diabetes in mice.
Cell Metab. 8:333–341. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Noriega LG, Feige JN, Canto C, Yamamoto H,
Yu J, Herman MA, Mataki C, Kahn BB and Auwerx J: CREB and ChREBP
oppositely regulate SIRT1 expression in response to energy
availability. EMBO Rep. 12:1069–1076. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lan F, Cacicedo JM, Ruderman N and Ido Y:
SIRT1 modulation of the acetylation status, cytosolic localization,
and activity of LKB1: Possible role in AMP-activated protein kinase
activation. J Biol Chem. 283:27628–27635. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gao M and Liu D: Resveratrol suppresses
T0901317-induced hepatic fat accumulation in mice. AAPS J.
15:744–752. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ajmo JM, Liang X, Rogers CQ, Pennock B and
You M: Resveratrol alleviates alcoholic fatty liver in mice. Am J
Physiol Gastrointest Liver Physiol. 295:G833–G842. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Timmers S, Konings E, Bilet L, Houtkooper
RH, van de Weijer T, Goossens GH, Hoeks J, van der Krieken S, Ryu
D, Kersten S, et al: Calorie restriction-like effects of 30 days of
resveratrol supplementation on energy metabolism and metabolic
profile in obese humans. Cell Metab. 14:612–622. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Faghihzadeh F, Adibi P and Hekmatdoost A:
The effects of resveratrol supplementation on cardiovascular risk
factors in patients with non-alcoholic fatty liver disease: A
randomised, double-blind, placebo-controlled study. Br J Nutr.
114:796–803. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Heebøll S, Kreuzfeldt M, Hamilton-Dutoit
S, Poulsen MK, Stødkilde-Jørgensen H, Møller HJ, Jessen N, Thorsen
K, Hellberg YK, Pedersen SB and Grønbæk H: Placebo-controlled,
randomised clinical trial: High-dose resveratrol treatment for
non-alcoholic fatty liver disease. Scand J Gastroenterol.
51:456–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Davenport AM, Huber FM and Hoelz A:
Structural and functional analysis of human SIRT1. J Mol Biol.
426:526–541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pan M, Yuan H, Brent M, Ding EC and
Marmorsteins R: SIRT1 contains N- and C-terminal regions that
potentiate deacetylase activity. J Biol Chem. 287:2468–2476. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
McBurney MW, Clark-Knowles KV, Caron AZ
and Gray DA: SIRT1 is a highly networked protein that mediates the
adaptation to chronic physiological stress. Genes Cancer.
4:125–134. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Olmos Y, Brosens JJ and Lam EWF: Interplay
between SIRT proteins and tumour suppressor transcription factors
in chemotherapeutic resistance of cancer. Drug Resist Updat.
14:35–44. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yanagisawa S, Baker JR, Vuppusetty C, Koga
T, Colley T, Fenwick P, Donnelly LE, Barnes PJ and Ito K: The
dynamic shuttling of SIRT1 between cytoplasm and nuclei in
bronchial epithelial cells by single and repeated cigarette smoke
exposure. PLoS One. 13:e01939212018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Smith BK, Marcinko K, Desjardins EM, Lally
JS, Ford RJ and Steinberg GR: Treatment of nonalcoholic fatty liver
disease: Role of AMPK. Am J Physiol Endocrinol Metab.
311:E730–E740. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hardie DG, Ross FA and Hawley SA: AMPK: A
nutrient and energy sensor that maintains energy homeostasis. Nat
Rev Mol Cell Biol. 13:251–262. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Corey KE, Vuppalanchi R, Vos M, Kohli R,
Molleston JP, Wilson L, Unalp-Arida A, Cummings OW, Lavine JE,
Chalasani N, et al: Improvement in liver histology is associated
with reduction in dyslipidemia in children with nonalcoholic fatty
liver disease. J Pediatr Gastroenterol Nutr. 60:360–367. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
de Oliveira CP, Cotrim HP, Stefano JT,
Siqueira ACG, Salgado ALA and Parise ER: N-acetylcysteine and/or
ursodeoxycholic acid associated with metformin in non-alcoholic
steatohepatitis: An open-label multicenter randomized controlled
trial. Arq Gastroenterol. 56:184–190. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Cai Z, Lou Q, Wang F, Li E, Sun J, Fang H,
Xi J and Ju L: N-acetylcysteine protects against liver injure
induced by carbon tetrachloride via activation of the Nrf2/HO-1
pathway. Int J Clin Exp Pathol. 8:8655–8662. 2015.PubMed/NCBI
|
|
60
|
Bauerlein DK, Akbar HN, von Rosenvinge EC,
Loughry ND and John PR: Benefit of N-acetylcysteine in
postoperative hepatic dysfunction: Case report and review of
literature. Case Reports Hepatol. 2019:47303812019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Jansen T, Kvandová M, Daiber A, Stamm P,
Frenis K, Schulz E, Münzel T and Kröller-Schön S: The AMP-activated
protein kinase plays a role in antioxidant defense and regulation
of vascular inflammation. Antioxidants. 9:5252020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang M, Yang D, Gong X, Ge P, Dai J, Lin
L and Zhang L: Protective benefits of AMP-activated protein kinase
in hepatic ischemia-reperfusion injury. Am J Transl Res. 9:823–829.
2017.PubMed/NCBI
|
|
63
|
Meng S, Cao J, He Q, Xiong L, Chang E,
Radovick S, Wondisford FE and He L: Metformin activates
AMP-activated protein kinase by promoting formation of the
αβγheterotrimeric complex. J Biol Chem. 290:3393–3802. 2015.
View Article : Google Scholar
|
|
64
|
Ouyang J, Parakhia RA and Ochs RS:
Metformin activates AMP kinase through inhibition of AMP deaminase.
J Biol Chem. 286:1–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Fouqueray P, Bolze S, Dubourg J,
Hallakou-Bozec S, Theurey P, Grouin JM, Chevalier C, Gluais-Dagorn
P, Moller DE and Cusi K: Pharmacodynamic effects of direct AMP
kinase activation in humans with insulin resistance and
non-alcoholic fatty liver disease: A phase 1b study. Cell Reports
Med. 2:1004742021. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gluais-Dagorn P, Foretz M, Steinberg GR,
Batchuluun B, Zawistowska-Deniziak A, Lambooij JM, Guigas B,
Carling D, Monternier PA, Moller DE, et al: Direct AMPK activation
corrects NASH in rodents through metabolic effects and direct
action on inflammation and fibrogenesis. Hepatol Commun. 6:101–119.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Monternier PA, Parasar P, Theurey P,
Dagorn PG, Kaur N, Nagaraja TN, Fouqueray P, Bolze S, Moller DE,
Singh J and Hallakou-Bozec S: Beneficial effects of the direct
AMP-kinase activator PXL770 in in vitro and in vivo models of
X-linked adrenoleukodystrophy. J Pharmacol Exp Ther. 382:208–222.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Shargorodsky M, Omelchenko E, Matas Z,
Boaz M and Gavish D: Relation between augmentation index and
adiponectin during one-year metformin treatment for nonalcoholic
steatohepatosis: Effects beyond glucose lowering? Cardiovasc
Diabetol. 11:612012. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Green CJ, Marjot T, Walsby-Tickle J,
Charlton C, Cornfield T, Westcott F, Pinnick KE, Moolla A,
Hazlehurst JM, McCullagh J, et al: Metformin maintains intrahepatic
triglyceride content through increased hepatic de novo lipogenesis.
Eur J Endocrinol. 186:367–377. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Resuli B, Demiraj V, Babameto A, Sema K
and Malaj V: Metformin superior to low-fat diet for the treatment
of patients with nonalcoholic fatty liver disease and/or
steatohepatitis. Pol Arch Med Wewn. 122:68–71. 2012.PubMed/NCBI
|
|
71
|
Feng WH, Bi Y, Li P, Yin TT, Gao CX, Shen
SM, Gao LJ, Yang DH and Zhu DL: Effects of liraglutide, metformin
and gliclazide on body composition in patients with both type 2
diabetes and non-alcoholic fatty liver disease: A randomized trial.
J Diabetes Investig. 10:399–407. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Yang X, Peng X and Huang J: Inhibiting
6-phosphogluconate dehydrogenase selectively targets breast cancer
through AMPK activation. Clin Transl Oncol. 20:1145–1152. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Sarfraz I, Rasul A, Hussain G, Shah MA,
Zahoor AF, Asrar M, Selamoglu Z, Ji XY, Adem S and Sarker SD:
6-Phosphogluconate dehydrogenase fuels multiple aspects of cancer
cells: From cancer initiation to metastasis and chemoresistance.
Biofactors. 46:550–562. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Marini C, Cossu V, Bauckneht M, Lanfranchi
F, Raffa S, Orengo AM, Ravera S, Bruno S and Sambuceti G: Metformin
and cancer glucose metabolism: At the bench or at the bedside?
Biomolecules. 11:12312021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Faghihzadeh F, Adibi P, Rafiei R and
Hekmatdoost A: Resveratrol supplementation improves inflammatory
biomarkers in patients with nonalcoholic fatty liver disease. Nutr
Res. 34:837–843. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chen S, Zhao X, Ran L, Wan J, Wang X, Qin
Y, Shu F, Gao Y, Yuan L, Zhang Q and Mi M: Resveratrol improves
insulin resistance, glucose and lipid metabolism in patients with
non-alcoholic fatty liver disease: A randomized controlled trial.
Dig Liver Dis. 47:226–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Purushotham A, Schug TT, Xu Q, Surapureddi
S, Guo X and Li X: Hepatocyte-specific deletion of SIRT1 alters
fatty acid metabolism and results in hepatic steatosis and
inflammation. Cell Metab. 9:327–338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Rezzani R and Franco C: Liver, oxidative
stress and metabolic syndromes. Nutrients. 13:3012021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cichoz-Lach H and Michalak A: Oxidative
stress as a crucial factor in liver diseases. World J
Gastroenterol. 20:8082–8091. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Furman D, Campisi J, Verdin E,
Carrera-Bastos P, Targ S, Franceschi C, Ferrucci L, Gilroy DW,
Fasano A, Miller GW, et al: Chronic inflammation in the etiology of
disease across the life span. Nat Med. 25:1822–1832. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Andrade JMO, Paraíso AF, de Oliveira MVM,
Martins AME, Neto JF, Guimarães ALS, de Paula AM, Qureshi M and
Santos SHS: Resveratrol attenuates hepatic steatosis in high-fat
fed mice by decreasing lipogenesis and inflammation. Nutrition.
30:915–919. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Chai D, Zhang L, Xi S, Cheng Y, Jiang H
and Hu R: Nrf2 activation induced by Sirt1 ameliorates acute lung
injury after intestinal ischemia/reperfusion through NOX4-mediated
gene regulation. Cell Physiol Biochem. 46:781–792. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ren Z, He H, Zuo Z, Xu Z, Wei Z and Deng
J: The role of different SIRT1-mediated signaling pathways in toxic
injury. Cell Mol Biol Lett. 24:362019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Du F, Huang R, Lin D, Wang Y, Yang X,
Huang X, Zheng B, Chen Z, Huang Y, Wang X and Chen F: Resveratrol
improves liver steatosis and insulin resistance in non-alcoholic
fatty liver disease in association with the gut microbiota. Front
Microbiol. 12:6113232021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wardani HA, Rahmadi M, Ardianto C, Balan
SS, Kamaruddin NS and Khotib J: Development of nonalcoholic fatty
liver disease model by high-fat diet in rats. J Basic Clin Physiol
Pharmacol. 30:1–7. 2020.PubMed/NCBI
|
|
86
|
Theodotou M, Fokianos K, Moniatis D,
Kadlenic R, Chrysikou A, Aristotelous A, Mouzouridou A, Diakides J
and Stavrou E: Effect of resveratrol on non-alcoholic fatty liver
disease. Exp Ther Med. 559–565. 2019.PubMed/NCBI
|
|
87
|
Zhou Q, Wang Y, Han X, Fu S, Zhu C and
Chen Q: Efficacy of resveratrol supplementation on glucose and
lipid metabolism: A meta-analysis and systematic review. Front
Physiol. 13:7959802022. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhao H, Zhang Y, Shu L, Song G and Ma H:
Resveratrol reduces liver endoplasmic reticulum stress and improves
insulin sensitivity in vivo and in vitro. Drug Des Devel Ther.
13:1473–1485. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
León D, Uribe E, Zambrano A and Salas M:
Implications of resveratrol on glucose uptake and metabolism.
Molecules. 22:3982017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Abd El-Haleim EA, Bahgat AK and Saleh S:
Resveratrol and fenofibrate ameliorate fructose-induced
nonalcoholic steatohepatitis by modulation of genes expression.
World J Gastroenterol. 22:2931–2948. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ding S, Jiang J, Zhang G, Bu Y, Zhang G
and Zhao X: Resveratrol and caloric restriction prevent hepatic
steatosis by regulating SIRT1-autophagy pathway and alleviating
endoplasmic reticulum stress in high-fat diet-fed rats. PLoS One.
12:e01835412017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Yang H, Liu Y, Wang Y, Xu S and Su D:
Knockdown of Sirt1 gene in mice results in lipid accumulation in
the liver mediated via PGC-1α-induced mitochondrial dysfunction and
oxidative stress. Bull Exp Biol Med. 172:180–186. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Hou X, Rooklin D, Fang H and Zhang Y:
Resveratrol serves as a protein-substrate interaction stabilizer in
human SIRT1 activation. Sci Rep. 6:381862016. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cao D, Wang M, Qiu X, Liu D, Jiang H, Yang
N and Xu RM: Structural basis for allosteric, substratedependent
stimulation of SIRT1 activity by resveratrol. Genes Dev.
29:1316–1325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Gertz M, Nguyen GTT, Fischer F, Suenkel B,
Schlicker C, Fränzel B, Tomaschewski J, Aladini F, Becker C,
Wolters D and Steegborn C: A molecular mechanism for direct sirtuin
activation by resveratrol. PLoS One. 7:e497612012. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Schug TT and Li X: Sirtuin 1 in lipid
metabolism and obesity. Ann Med. 43:198–211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Price NL, Gomes AP, Ling AJY, Duarte FV,
Martin-Montalvo A, North BJ, Agarwal B, Ye L, Ramadori G, Teodoro
JS, et al: SIRT1 is required for AMPK activation and the beneficial
effects of resveratrol on mitochondrial function. Cell Metab.
15:675–690. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Ford RJ, Desjardins EM and Steinberg GR:
Are SIRT1 activators another indirect method to increase AMPK for
beneficial effects on aging and the metabolic syndrome?
EBioMedicine. 19:16–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ruderman NB, Xu XJ, Nelson L, Cacicedo JM,
Saha AK, Lan F and Ido Y: AMPK and SIRT1: A long-standing
partnership? Am J Physiol Endocrinol Metab. 298:E751–E760. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Duarte-Vázquez MA, Gómez-Solis A,
Gómez-Cansino R, Reyes-Esparza J, Luis Rosado J and
Rodriguez-Fragoso L: Effect of combined resveratrol plus metformin
therapy in db/db diabetic mice. FASEB J. 31:1001.8. 2017.
|
|
101
|
Li S, Qian Q, Ying N, Lai J, Feng L, Zheng
S, Jiang F, Song Q, Chai H and Dou X: Activation of the AMPK-SIRT1
pathway contributes to protective effects of Salvianolic acid A
against lipotoxicity in hepatocytes and NAFLD in mice. Front
Pharmacol. 11:5609052020. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Chen XY, Cai CZ, Yu ML, Feng ZM, Zhang YW,
Liu PH, Zeng H and Yu CH: LB100 ameliorates nonalcoholic fatty
liver disease via the AMPK/Sirt1 pathway. World J Gastroenterol.
25:6607–6618. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Chalasani N, Vuppalanchi R, Rinella M,
Middleton MS, Siddiqui MS, Barritt AS IV, Kolterman O, Flores O,
Alonso C, Iruarrizaga-Lejarreta M, et al: Randomised clinical
trial: A leucine-metformin-sildenafil combination (NS-0200) vs
placebo in patients with non-alcoholic fatty liver disease. Aliment
Pharmacol Ther. 47:1639–1651. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Banerjee J, Bruckbauer A and Zemel MB:
Activation of the AMPK/Sirt1 pathway by a leucine-metformin
combination increases insulin sensitivity in skeletal muscle, and
stimulates glucose and lipid metabolism and increases life span in
Caenorhabditis elegans. Metabolism. 65:1679–1691. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Bruckbauer A and Zemel MB: Synergistic
effects of polyphenols and methylxanthines with leucine on
AMPK/Sirtuin-mediated metabolism in muscle cells and adipocytes.
PLoS One. 9:e891662014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Liang C, Curry BJ, Brown PL and Zemel MB:
Leucine modulates mitochondrial biogenesis and SIRT1-AMPK signaling
in C2C12 myotubes. J Nutr Metab. 2014:2397502014. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Bruckbauer A and Zemel MB: Effects of
dairy consumption on SIRT1 and mitochondrial biogenesis in
adipocytes and muscle cells. Nutr Metab (Lond). 8:912011.
View Article : Google Scholar : PubMed/NCBI
|