Introduction
Obesity has rapidly become a widespread public
health concern, the incidence of which has gradually increased over
numerous decades. By 2025, the World Health Organization predicts
that one in five adults will be obese globally (1). Obesity is a highly heterogeneous and
complicated disorder caused by unbalanced energy metabolism. In
addition, obesity is also closely associated with the pathogenesis
of various metabolic diseases, such as type 2 diabetes mellitus
(T2D), dyslipidemia, and cardiovascular diseases (CVDs), including
hypertension and stroke, neurological disorders, musculoskeletal
disease, and certain types of cancer (for example, breast, liver,
ovarian, kidney, prostate and colon cancer) (2). Furthermore, several studies have
found that obese patients with associated comorbidities are more
susceptible to SARS-CoV-2, exhibiting a higher risk of death
(3–5).
Adipose tissues (ATs) are complex tissues that
primarily exhibit a regulatory function. In mammals, ATs are
primarily classified into white AT (WATs) and brown AT (BATs). In
addition to metabolizing fat into energy, ATs also serve as a
critical endocrine organ that can regulate energy metabolism,
immunological responses, and cardiovascular balance by secreting a
variety of adipokines, peptide hormones, and cytokines (6). Over the years, ~100 adipokines have
been identified (7). Adipokines
have a wide range of physiological effects on tissues and organs in
different systems, such as the nervous system, immune system, and
vascular system (8). For example,
ATs secrete adiponectin that can promote insulin sensitivity and
the antiatherosclerotic properties of cells by binding to
adiponectin receptor (AdipoR) 1 and AdipoR2 (9–11).
An inverse association exists between circulating adiponectin and
obesity-related cancer incidence (11). Furthermore, adiponectin acts as an
essential metabolic reprogramming factor by promoting the
interaction between adaptor protein, phosphotyrosine interacting
with PH domain, and leucine zipper 1 (APPL1) and AMP-activated
protein kinase (AMPK), promoting glucose uptake through glucose
transporter 4 (GLUT4) (12). The
release of leptin by adipocytes was correlated with alterations in
cell metabolism, such as the switch from mitochondrial β-oxidation
to aerobic glycolysis (13).
Chronic inflammation is another well-established characteristic of
obesity (14). In obesity, there
is an increase in oxidative stress and inflammation, leading to an
increased release of proinflammatory adipokines, which can
contribute to insulin resistance in the liver, muscles, and ATs,
resulting in metabolic abnormalities (15). Studies have shown a positive
correlation between obesity, insulin levels, insulin resistance,
and increased tumor necrosis factor (TNF)-α production in human
adipocytes (16).
In addition to the classical polypeptide adipokines
and cytokines, ATs can also produce and secrete extracellular
vesicles (EVs) (17). The
composition of all EVs is similar to that of the parent cells,
packed with bioactive molecules such as lipids, proteins, and DNA
delivered to cells within ATs or in distant organs, mediating
intercellular and interorgan communication (14). In this context, AT-derived
extracellular vesicles (ADEVs) have been identified as crucial
players in the cellular communication of immune and metabolic
responses, regulating cellular processes in local and distant
tissues (18,19). This review will focus on the
compositions and functions of ADEVs from different cellular sources
in ATs and their contribution to AT homeostasis and the development
of metabolic complications, such as metabolic diseases, CVDs,
several types of cancer, and neurological disorders.
Introduction to EVs
Initially, EVs were viewed as a quality control
system to eliminate harmful or unnecessary molecules from the cell
(20). EVs are now identified as a
group of submicron-sized membrane-bound organelles secreted by
almost all cells, carrying several biological cargoes, such as
lipids, fatty acids (FAs), and nucleic acids, capable of targeting
and transferring their contents to various receptor cells within
the tissue or distal tissues (Fig.
1). EVs primarily consist of exosomes, microvesicles (MVs) and
apoptotic bodies (21). Apoptotic
bodies are known to be produced during apoptotic cell death and
have a diameter >5 µm (22).
MVs are small vesicles (100–1,000 nm size range) formed by plasma
membrane fusion and budding. Although the exact process by which
MVs are formed is not fully understood, cytoskeletal elements such
as actin and microtubules, coat proteins, and fusion machinery,
such as SNAREs, are hypothesized to be necessary. Specifically,
coat proteins such as clathrin and cytoplasmic coat protein
complex, are drawn to the membrane to reshape the flat membranes
into rounded buds, cargo, and vesicle-SNAREs (v-SNAREs, primarily
including VAMP) are integrated into the budding vesicle by
attaching to coat subunits, for example, adaptor protein (AP)
complexes (23). In addition, the
molecular composition of MVs primarily consists of cytoplasmic and
plasma membrane-associated proteins since MVs are formed by the
outward budding of the membrane, and they may vary greatly
depending on the cell type (24).
In addition, MVs were first described as subcellular material
originating from platelets and were demonstrated to play a role in
blood coagulation (25,26). More recently, they have been
reported to transfer cargo to target cells, thus playing an
essential role in cell communication (27). Exosomes are vesicles 30–150 nm in
diameter secreted by the endosome pathway. During the biogenesis of
exosomes, endocytosis-mediated invagination of the plasma membrane
(PM) forms early endosomes. Endosomal membranes bud inward into the
lumen to create intraluminal vesicles (ILVs). These late endosomes
contain ILVs called multivesicular bodies (MVBs). MVBs can fuse
with lysosomes to be degraded or fuse with the PM to release ILVs
as exosomes into the extracellular environment (28,29).
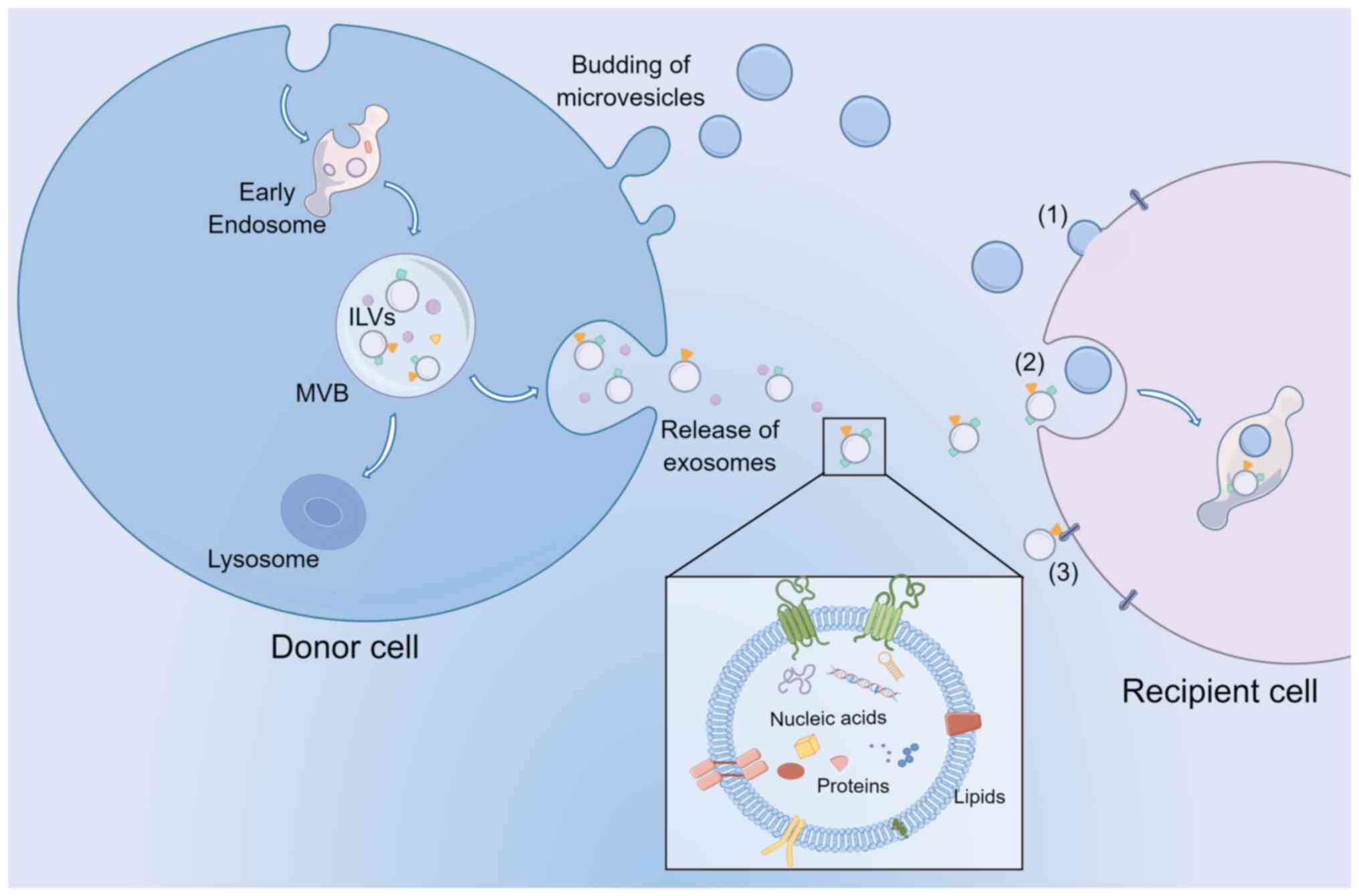 | Figure 1.EV biogenesis, release, and
communication with recipient cells. Exosomes and MVs released from
cells are primarily stratified based on size. In addition, MVs can
bud from the plasma membrane, while exosomes are derived from ILVs
within the lumen of MVBs. MVBs then fuse with the plasma membrane
to release the exosomes. EVs transport nucleic acids, proteins, and
lipids to target cells by (1)
membrane fusion, (2) endocytosis,
and (3) ligand binding mechanisms
to exert their biological functions. EV, extracellular vesicle;
MVs, microvesicles; ILVs, intraluminal vesicles; MVBs,
multivesicular bodies. |
The nature and abundance of EV cargoes are specific
to the cell type. They are frequently affected by the state of
donor cells and the molecular processes that result in their
biogenesis (30,31). EVs are loaded with various
biomolecular components, such as nucleic acids and proteins,
contributing to their functional diversity, heterogeneity, and
complexity. Proteins commonly found in EVs are those associated
with biogenetic mechanisms, including those related to endosomal
pathways. Several membrane proteins and transcription factors can
also be found in EVs (32,33). EVs are rich in sphingomyelin,
cholesterol, desaturated lipids, phosphatidylserine, and ceramide
(34). In addition, a range of
genetic material is found in EVs, such as DNA, mRNAs, microRNAs
(miRNAs), and several noncoding RNAs (ncRNAs). As soon as EVs bind
to target cells, they may remain in the PM or be ingested through
endocytosis, direct membrane fusion, and ligand binding mechanisms
(35–37).
Proteins of the tetraspanin family, such as CD81 and
CD9, are enriched in EVs and considered unique markers of EVs,
including exosomes and MVs (38).
However, researchers have demonstrated that CD81, CD63, and CD9 are
exosome markers in a recent study on the difference between
exosomes and MVs (39). At the
same time, they emphasized that Annexin A1 is present in MVs, not
exosomes. Furthermore, MVs also contain several biological
molecules, such as integrins, selectins, and CD40 ligands, which
may facilitate the formation of MVs (40,41).
Therefore, more research is required to distinguish them from each
other. In the present review, ‘EVs’ is used to refer to exosomes
and MVs only.
AT and ADEVs
AT
ATs are complex metabolic organs with profound
effects on regulating systemic metabolism, energy storage, and
homeostasis. The primary characteristic of BATs is the presence of
multilocular lipid droplets and several mitochondria expressing
high levels of uncoupling protein 1 (UCP1), responsible for
nonshivering thermogenesis, leading to increased energy expenditure
(42). WATs primarily consist of
white adipocytes, which carry large lipid droplets and fewer
mitochondria, making them the primary site for storing and
releasing energy (42). In
addition, studies have shown that WATs can undergo a process called
‘browning’, during which part of the white adipose tissue can be
transformed into beige adipose tissue (BeATs), morphologically
distinct from WATs and BATs (43,44).
Browning occurs under certain circumstances, such as in the cold
and as a result of exercise. Moreover, medicines, such as
β-adrenergic receptor and peroxisome proliferator-activated
receptor (PPAR)γ agonists, can also trigger browning by promoting
the decomposition of triglyceride and glucose in ATs or by inducing
the expression of thermogenesis-related genes, respectively, which
ultimately encourages lipolysis and thermogenesis (45–47).
Characteristically, beige adipocytes contain several small lipid
droplets, are typically larger than brown adipocytes, have more
mitochondria than white adipocytes, and express UCP1 (48). Additionally, these types of ATs
differ in critical ways that include aspects of their gene
expression profile and secretome. WAT, known as the active
endocrine organ, can release cytokines and adipokines such as
leptin and adiponectin (6). In
contrast, BAT/BeAT have fewer secretory functions than WAT. In
addition to UCP1, Cell death-inducing DFFA like effector a,
Cytochrome c oxidase subunit 7A1, and ELOVL fatty acid elongase 3
are specifically expressed in BAT, while BeAT expresses T-box
transcription factor 1, Solute carrier family 27 member 1,
Cbp/p300-interacting transactivator with Glu/Asp-rich
carboxy-terminal domain 1, CD40 and CD137 (49,50).
Since the UCP1 expression of beige adipocytes is considerably lower
than those of classical brown adipocytes, BeATs were once
considered unimportant in whole-body energy expenditure (51). However, in adults with a minimal
classic BAT reserve, BeATs are the primary energy source for
nonshivering thermogenesis (52).
Additionally, BeATs regulate whole-body energy metabolism and
glucose homeostasis through an UCP1 independent mechanism (51). According to the report, activation
of BeATs with β-adrenergic agonist CL316243 enhanced the selective
uptake of fatty acids from triglyceride-rich lipoproteins by ATs,
and reduced plasma TG and cholesterol levels, thereby alleviating
hypercholesterolemia and atherosclerosis (53). Another study showed that prolonged
maintenance of thermogenically active BeATs enhanced whole-body
energy expenditure and protected mice from diet-induced obesity and
insulin resistance (54). As the
most abundant form of AT, WAT is distributed throughout the body
and are active endocrine organs. They release free fatty acids and
adipokines, such as leptin, adiponectin, TNF-α, and IL-6, which act
on distal tissues, including the brain, liver, and muscle tissue,
to regulate food intake, energy homeostasis, and insulin
sensitivity (55). The ATs found
in the hypodermis layer are called subcutaneous ATs (SCATs), and
form a connective tissue in the dermis between the aponeurosis and
the muscle fascia, insulating and storing energy. SCATs are
primarily distributed in the abdominal and gluteofemoral regions of
the human body, storing >80% of total body fat (56). Dermal WATs (DATs), located directly
below the reticular dermis (primarily above the SCATs), have been
reported to be involved in insulation, hair regeneration, wound
healing, and the prevention of skin infections (57,58).
In addition, WATs that accumulate around internal organs are
visceral ATs (VATs), which are primarily found in the intrathoracic
region and abdominal cavity, such as epicardial and pericardial fat
and perigonadal, mesenteric, perirenal, and retroperitoneal fat,
protecting the internal organs of rodents and humans and storing
5–20% of total body fat (59,60).
Cellular composition of ATs
ATs are connective tissues primarily made up of
lipid-rich cells known as adipocytes. Adipocytes, the parenchymal
cells of ATs, are critical regulators of energy metabolism and
endocrine modulators engaged in numerous physiological or
pathological processes, such as appetite regulation and
immunological response (61,62).
In addition to adipocytes, there are several nonadipocyte
compartments termed the stromal vascular fraction (SVF), composed
of AT-derived stem cells (ADSCs), preadipocytes, endothelial cells,
and a broad spectrum of adaptive and innate immune cells (63–66).
Preadipocytes can differentiate into mature adipocytes to maintain
adipogenesis and homeostasis in adipose tissue (67). The ADSCs in ATs are mesenchymal
stem cells (MSCs) of mesodermal origin, serving as progenitors
responsible for adipocyte regeneration and replenishment. ADSCs
also have potent self-renewal capacity and a high capacity for
classical adipogenic, osteogenic, and chondrogenic differentiation.
In addition to mesenchymal cells, ADSCs can differentiate into
nonmesenchymal cell lineages, such as endothelial cells, myocytes,
and neuronal lineages (68).
Endothelial cells and pericytes provide vasculature to ATs by
forming capillaries (69–71). The immune cell types and functions
of ATs have been widely discussed, primarily in the context of
obesity. Various immune cells form a dynamic immunological
microenvironment with variable metabolic status, including
macrophages, eosinophils, dendritic cells (DCs), invariant natural
killer cells (iNKT cells), T cells, and B cells (72–74).
For example, under conditions of obesity or chronic metabolic
stress, increased infiltration and activation of proinflammatory
immune cells can accelerate WAT inflammation, thus influencing the
effect of insulin and other metabolic hormones on parenchymal
cells, thus further damaging the glucose and lipid metabolism
process of metabolic organs (75–77).
Working model and the source of
ADEVs
Although EVs are physiologically released from
cells, pathophysiological stimuli can regulate their biogenesis and
release. Furthermore, certain proteins and mRNAs can be selectively
packaged into EVs during physiological changes or pathological
injuries. Similar to normal EVs, ADEVs exert their biological
functions by transporting bioactive cargos such as miRNAs, ncRNAs,
proteins, and lipids to receptor cells. Studies have shown that
ADEVs can not only modulate the immune responses of local ATs
through cellular communication but can also regulate systemic
insulin sensitivity and glycolipid metabolic processes through
their remote effects on other metabolic organs (for example, the
brain and liver) (78–80) (Fig.
2). Research has shown that ADEVs directly modulate glucose
tolerance and insulin sensitivity in adipocytes, myocytes, and
hepatocytes through modulation of PPARγ and perhaps fibroblast
growth factor 21 (81,82). In the brain, ADEVs derived from
adipocytes have been shown to carry the long noncoding RNA (lncRNA)
metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) and
to activate the mTOR signaling pathway through miR-181b and miR-144
in hypothalamic anorexigenic pro-opiomelanocortin (POMC) neurons,
thereby leading to increased appetite and body weight (83). In addition, analysis of the protein
profiles of EVs derived from adipocytes confirmed that EVs from
obese mice are enriched in proteins and enzymes involved in the
metabolism and transport of lipids, such as caveolin 1, lipoprotein
lipase, and aquaporin 7, which may be associated with ectopic lipid
accumulation and lead to mitochondrial energy metabolism
disturbance, and systemic insulin resistance (84,85).
In particular, quantitative proteomic analysis of EVs released by
3T3-L1 adipocytes showed that enzymes related to de novo
lipogenesis, including glucose-6-phosphate dehydrogenase and fatty
acid synthase, were selectively enriched in EVs from adipocytes,
promoting lipid accumulation in recipient adipocytes and
preadipocytes (85).
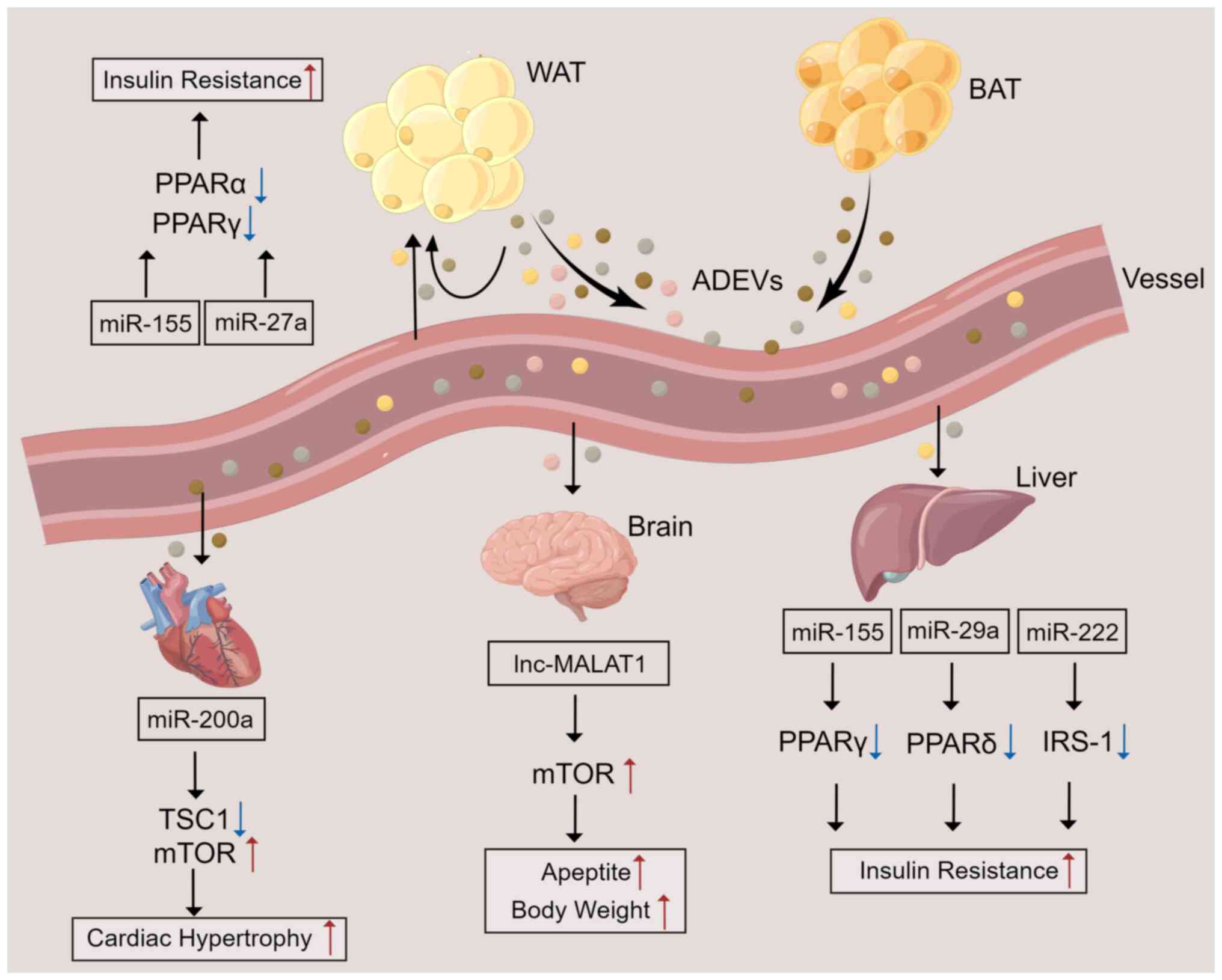 | Figure 2.ADEV-mediated crosstalk between ATs
and other organs. ATs secrete EVs containing various components
into the blood circulation to affect ATs and distant organs. In ATs
and the liver, miRNAs contained in ADEVs result in insulin
resistance by inhibiting the activation or expression of PPARα,
PPARγ, PPARδ, or IRS-1. In the brain, ADEVs contain lncRNA MALAT1,
which enhances mTOR signaling in POMC neurons, leading to increased
appetite and a gain in body gain. miR-200a contained in ADEVs can
be delivered to cardiomyocytes to inhibit TSC1 and activate the
mTOR pathway, leading to cardiac hypertrophy. ATs, adipose tissues;
ADEVs, AT-derived extracellular vesicles; WAT, white AT; BAT, brown
AT; miR, microRNA; lnc, long non-coding RNA; TSC1, tuberous
sclerosis complex 1; PPAR, peroxisome proliferator-activated
receptor; IRS-1, insulin receptor substrate 1; MALAT1,
metastasis-associated lung adenocarcinoma transcript 1. |
Most of the current body of knowledge on ADEVs comes
from studies using 3T3-L1 cells. These studies found that ADEVs are
highly adipocyte-specific and can be identified from complicated
heterogeneous origins, such as plasma (86,87).
Currently, multiple types of cells in ATs, such as adipocytes,
ADSCs, and macrophages, have been shown to release EVs, mediating
intercellular and interorgan crosstalk and regulating ATs and
systemic homeostasis (88,89). EVs derived from adipocytes have
been reported to carry proteins or enzymes involved in fatty acid
oxidation (FAO), which can induce metabolic reprogramming and
stimulate the migration and invasion of melanoma cells when EVs are
taken up by tumor cells, thus amplifying the deleterious dialog
between cancer cells and adipocytes (78,90).
Furthermore, EVs derived from AT macrophages (ATMs) can modulate
mouse glucose tolerance and insulin sensitivity (81,91).
ADSCs have a high capacity for differentiation into multiple cell
types and play an essential role in immune regulation (92–94).
EVs released from ADSCs may at least be partially responsible for
some of these functions. ADSC-derived EVs (ADSC-EVs) obtained from
patients with or without cancer show equivalent miRNA content,
which suggests that ADSC-EVs have the same therapeutic paracrine
effects regardless of the health status of the donor (95). Previous studies have found that
delivering ADSC-EVs from lean mice to obese mice showed desirable
effects on alleviating obesity and IR (96). Additionally, ADSC-EVs obtained from
human WATs and BATs can induce ADSCs to differentiate into WATs and
BATs, respectively, attenuating diet-induced obesity, glucose
tolerance, and liver steatosis (97). These findings suggest that ADEVs
can be used as cell-free therapeutics for AT regeneration and
remodeling.
ADEVs in disease
ADEVs regulate metabolic
disorders
AT dysfunction is accompanied by chronic low-grade
inflammation, in which excessive adipokine synthesis and secretion
are closely related to cardiovascular, chronic liver, and kidney
diseases as well as other systemic metabolic disorders (98,99).
The inflammation of WATs caused by obesity is characterized by the
accumulation of macrophages, including monocyte-derived macrophages
(moMacs). Moreover, obese WAT monocytes can differentiate locally
into moMacs and contribute to ATM pools (100). During the development of obesity,
macrophages transition from an anti-inflammatory phenotype to a
proinflammatory phenotype, producing proinflammatory factors and
exacerbating adipose inflammation (101). Initially, ADEVs were shown to be
taken up by peripheral monocytes, which then differentiated into
macrophages that increased TNF-α and IL-6 secretion, leading to
insulin resistance and glucose intolerance through the Toll-like
receptor 4/TIR domain-containing adaptor molecule 1 pathway
(102). Notably, a recent study
revealed a mechanism by which adipocytes regulate ATM polarization
through EVs. ADEVs derived from mature adipocytes containing
miR-34a inhibited macrophage M2 polarization by downregulating
Krüppel-like factor 4, a crucial transcription factor for
maintaining an adipose M2 macrophage phenotype, thereby leading to
adipose inflammation (103).
Several immunomodulatory proteins, such as macrophage migration
inhibitory factor, retinol-binding protein 4, and soluble
adiponectin, have been identified in EVs from cultured human
adipocytes and human AT explants, which may induce monocytes to
differentiate into M1-phenotype macrophages, exacerbating adipose
inflammation and insulin resistance (86).
Insulin resistance (IR) is a disordered biological
response in which the body cannot respond to high insulin levels or
absorb and utilize glucose normally, causing the body to produce
more insulin in response. It is the critical cause of several
metabolic syndromes, such as T2D and obesity (104). As mentioned above, ADEVs from the
VATs of obese mice were enriched in fatty acid binding protein 4
(FABP4) and induced the differentiation of macrophages to the M1
phenotype. In addition, those ADEVs were shown to be taken up by
monocytes and to induce IR (102). Ying et al (91) found that injecting ATM-derived EVs
from obese mice into lean mice inhibited the expression of PPARγ,
which can promote whole-body lipid metabolism and insulin
sensitivity, and GLUT4 (a PPARγ target gene), thereby reducing
adipocyte sensitivity to insulin. Specifically, EVs containing
miR-155 derived from ATMs target PPARγ in adipocytes, the liver,
and the muscle, regulating insulin activity (91). In addition, adipocytes secrete EVs
containing miR-27a, a negative regulator of PPARγ, leading to
PPARγ-dependent obesity-induced IR by directly binding to the 3′UTR
of PPARγ (105). Subsequently,
ATM-derived EVs from obese mice were confirmed to possess miR-29a
and to cause IR by binding to the 3′UTR of PPARδ (a potent
modulator of insulin sensitivity) in adipocytes, myocytes, and
hepatocytes (81). Another study
showed that miR-222 from gonadal WAT-derived EVs promoted IR in the
liver and skeletal muscle by suppressing IRS-1 expression via
binding to the 3′UTR of IRS-1 (106). In contrast, ADSC-EVs facilitated
metabolic homeostasis and improved insulin sensitivity in obese
mice by alternatively activating M2 macrophage polarization and
reducing inflammation by activating arginase-1 through STAT3
carried by EVs (96). In addition,
miR-27a was enriched in adipocyte-derived EVs isolated from the
VATs of obese individuals, and those EVs were shown to contribute
to IR in the liver and skeletal muscle by inhibiting
insulin-induced Akt phosphorylation and PPARα expression (107).
Given that obesity is a risk factor for the
development of T2D, research has focused on the relationship
between ATs and diabetes/diabetic complications. As no definitive
marker of ADEVs has yet been identified, it is challenging to
elucidate the detailed role of ADEVs in obesity and metabolic
syndromes such as T2D. The production and specific cargo of ADEVs
are altered under metabolic stresses (108). Perilipin A levels were higher in
circulating adipocyte-derived EVs from obese mice and humans with
metabolic diseases (109).
AT-derived miRNAs are the main circulating EV miRNAs (18). A study showed that miR-20b-5p was
abundant in serum EVs of T2D patients, and miR-20b-5p modulated
insulin action in skeletal muscle by downregulating Akt interacting
protein (AKTIP) and STAT3 expression (110). In addition, certain ADEVs exert
beneficial effects on T2D. A previous study assessed the
relationship between metabolic syndrome and adipose tissue-derived
EV markers, and revealed that individuals with CD14-positive EVs
had a 16% lower risk of developing T2D after 6.5 years of follow-up
(111). EVs derived from
activated beige adipocytes contain diabetes-preventing factors.
When administered to primary white adipocytes, these EVs improved
insulin sensitivity and insulin-stimulated glucose uptake (112). Together, based on these studies,
adipose-derived EVs are viewed as a novel cellular communication
tool within ATs and perhaps between ATs and distant organs to
regulate T2D.
ADEVs and CVD
Dysfunctional ATs in obese individuals can lead to
an increased risk of CVD, which remains one of the principal causes
of death worldwide, despite advances in risk factor management
(113–115). To date, efficient treatments for
CVD are lacking. Recently, ADEVs have emerged as critical actors in
the crosstalk between obesity and CVD progression.
Aside from polarization, macrophage foaming also
plays a vital role in the progression of atherosclerotic lesions.
ADEVs from VATs in obese mice facilitate macrophage foam cell
generation by downregulating ATP binding cassette subfamily A
member 1 (ABCA1) and ATP binding cassette subfamily G member 1
(ABCG1)-mediated cholesterol efflux and exacerbated atherosclerosis
(116). ADEVs from adipocytes and
their miRNA contents were confirmed to reduce macrophage
cholesterol efflux by targeting ABCA1, thus promoting the
development of atherosclerosis (117). In contrast, another study
identified the beneficial effects of EVs derived from ADSCs in
cardiac recovery, highlighting their potential in regenerative
therapy (118). In addition,
studies on 3T3-L1 models showed that adipocyte-derived EVs
containing miR-802-5p promoted IR in cardiomyocytes by
downregulating heat shock protein 60, which has been proven to
prevent inflammation, mitochondrial dysfunction, and even insulin
resistance (119).
Coronary artery disease and atherosclerosis are
caused by endothelial dysfunction during the early stages of the
disease (120). It is not well
understood how EVs are exchanged between adipocytes and endothelial
cells during obesity despite extensive evidence of proinflammatory
crosstalk between ATMs and ADEVs. It was found that hypertrophic
and dysfunctional adipocytes release EVs that impair vascular
endothelial cell function, potentially contributing to
obesity-related atherosclerosis (121). In addition, research has
demonstrated that hypoxia and inflammation promote synergistic EV
production from adipocytes. These ADEVs promote the expression of
endothelial vascular cell adhesion molecule 1 (VCAM-1), which
increases the subsequent attachment of leukocytes to endothelial
cells and exacerbates vascular disease in obesity (122).
One of the causes of heart failure is cardiac
hypertrophy (CH). Research has shown that miR-200a in ADEVs derived
from adipocytes can be transferred into cardiomyocytes to inhibit
TSC1 and activate the mTOR pathway, leading to CH. It was also
shown that inhibition of miR-200a could abrogate CH (123). In addition, Gan et al
(124) demonstrated that ADEVs
derived from diabetic adipocytes were delivered to cardiomyocytes
where they facilitated the pathogenic interaction between the heart
and defective ATs, aggravating ischemic heart damage in
obese/diabetic individuals. miR-130b-3p was found to be a vital
agent mediating this proapoptotic effect of diabetic
adipocyte-derived EVs and identified AMPK as a novel target of
miR-130b-3p, in which miR-130b-3p was shown to impair the
expression of AMPK, the latter of which is a crucial regulator of
metabolic disorder-induced cellular malfunction and cell death
(124). However, the precise role
of ADEVs is still poorly understood in the context of CVD and
requires further investigation.
ADEVs as major actors in cancer
An increasing body of data from animal and human
studies indicates that obesity increases the risk of developing
cancer, cancer-associated mortality, and cancer recurrence
following treatment (125).
Previously, studies on the communication between adipocytes and
tumor cells have been limited to cytokines such as endorphin,
leptin, or chemokines. Then, the discovery of cancer-associated
adipocytes (CAAs) revealed a vicious cycle in which tumors activate
CAAs, and CAAs can further contribute to tumor progression by
secreting adipokines, inflammatory cytokines, and metabolites
(126,127). More recently, the role of ADEVs
in tumor–adipose tissue communication has also been confirmed, and
obesity modifies ADEV secretion quantitatively and qualitatively,
thus amplifying their effect on tumor aggressiveness.
The first line of evidence linking ADEVs with cancer
showed that ADEVs from different adipocyte models enhanced melanoma
cell migration, invasion, and lung metastases in the context of
obesity (90). Subsequently, Wang
et al (128) showed that
EVs derived from 3T3-L1 adipocytes induced lung tumor metastasis by
increasing MMP9 activity of 3LL lung cancer cells, in which MMP9
has been shown to promote tumor invasion and metastasis. Another
study demonstrated that ADEVs from AT-derived mesenchymal stem
cells (ADMSCs) could foster the invasion, migration, and
proliferation of osteosarcoma cells by increasing galactosyl
transferase 2 and MMP2/9 expression (129). Gangadaran et al (130) demonstrated that ADSC-EVs contain
angiogenic proteins such as IL-8, CCL2, TIMP-1, TIMP-2, and VEGF-D.
Following internalization of ADSC-EVs, endothelial cells undergo
differentiation, develop a tube-like formation and promote
angiogenesis in vitro and in vivo. Khanh et al
(131) showed that in patients
with T2D, ADEVs derived from ADMSCs can promote breast cancer
metastasis by targeting the JAK/STAT3 pathway. In conditions of
obesity, ADEVs from AT macrophages are rich in miR-155, which is
not only involved in IR but also plays an oncogenic/antiapoptotic
role through caspase-3 and Bcl-2 in breast cancer cells (132).
By contributing local FAs to the process of FAO
within tumor cells, a novel beneficial metabolic route is activated
that increases tumor aggressiveness and proliferation, and
adipocytes also aid in the evolution of tumors through metabolic
collaboration (90,133,134). The proteins, including the
enzymes needed for FAO, can be transferred to cancer cells or
across farther distances by ADEVs released by adipocytes. Lazar
et al (90) demonstrated
that proteins implicated in FAO were enriched in ADEVs, which were
then taken up by melanoma cells, leading to increased lung
metastases and an increase in FAO in tumor cells. Clement et
al (78) revealed the role of
ADEVs in the crosstalk between melanoma cells and adipocytes, which
triggers metabolic remodeling and ultimately facilitates the FAO
process and tumor aggressiveness. In addition to FAO enzyme
transfer, ADEVs also deliver FAs to tumor cells to enhance the FAO
process, reinforcing the effect of ADEVs on obesity. Together, this
research revealed that ADEVs are involved in guiding the growth,
invasion, metabolic reprogramming, and metastasis of cancer cells
by modulating the acquisition and maintenance of cancer
markers.
Effects of ADEVs on neurological
disorders
Neurological disorders, including neurodegenerative
diseases such as Alzheimer's disease (AD) and Parkinson's disease
(PD), as well as ischemic stroke, are often characterized by
neuroinflammation. Treating neurological disorders has long been
challenging as many therapeutics do not cross the blood-brain
barrier (BBB). To date, EVs carrying biological molecules are
recognized as an excellent tool for treating central nervous system
(CNS)-related ailments since they are prospective drug delivery
systems that can cross the BBB (135).
The deposition of β-amyloid peptide (Aβ) in the
brain and neurofibrillary tangles (NFTs) formed by
hyperphosphorylated Tau protein plays a critical role in the
pathogenesis of AD (136,137). According to Katsuda et al
(138), ADSC-derived EVs carried
enzymatically active neprilysin (NEP), the brain's most important
Aβ-degrading enzyme. Furthermore, Aβ levels were decreased when
ADSC-derived EVs were transfused into N2a neuroblastoma cells in
vitro. Another study demonstrated that ADSC-derived EVs
inhibited inflammatory polarization of activated human microglia,
which has been shown to mediate several CNS inflammatory processes,
such as AD (139). In another rat
model of ischemic stroke, ADSC-derived EVs containing miR-126 were
shown to prevent ischemic stroke, promote neurogenesis, and
vasculogenesis following ischemic stroke, and inhibit ischemic
stroke-induced microglial activation and inflammatory responses
(140). Jiang et al
(141) showed that EVs derived
from ADSCs suppress autophagy by inhibiting the expression of
Beclin-1 and Atg5 via miR-30d-5p, thereby promoting microglial M2
polarization and ultimately preventing acute ischemic stroke. These
characteristics make ADSC-derived EVs promising candidates for
therapeutic relevance in neurodegenerative diseases.
Conclusions and future directions
In addition to the classical polypeptide adipokines
and cytokines, the critical role of ADEVs in communication between
ATs and other organs has been gradually deciphered. More recently,
growing evidence from both epidemiologic and preclinical studies
further highlights the effects of ADEVs on mediating cell-to-cell
communication within ATs, and between ATs and other peripheral
organs. A comprehensive understanding of the interaction patterns
between ATs and other tissues and the molecular changes in AT
dysfunction during obesity, such as ADEVs and their cargoes, may
provide novel avenues for developing new therapeutic interventions
in obesity-related metabolic diseases.
EVs have long been recognized as crucial
intercellular communication tools. In addition, EV-mediated
cellular communication is implicated in various diseases and
biological events, including certain immune responses such as
inflammation. Therefore, EVs are also considered a therapeutic
target for multiple diseases (142). In addition, due to their
endogenous origin, EVs have been widely explored as next-generation
nanoscale drug delivery systems, allowing them to circumvent
certain drawbacks associated with existing therapies (143). The impact of obesity on the
biological components of ADEVs from different cell origins has been
described previously (144).
Therefore, the critical function of ADEVs and the role of ADEVs and
their cargoes in multiple diseases were emphasized here.
The present review summarizes the composition and
function of ADEVs derived from different cell sources in ATs, such
as adipocytes, preadipocytes, macrophages, and MSCs. In addition,
ADEVs participate in developing pathologies associated with
metabolic diseases, CVDs, and several types of cancer (Table I). Understanding the mechanisms
behind the effects of ADEVs on obesity or metabolic disorders and
CVDs may contribute to the development of novel therapeutic
strategies. However, the vast majority of current research is
currently in the early stages, and no definitive marker of ADEV has
yet been identified, complicating the isolation of ADEVs from ATs
with high purity. In current research models, ADEVs from different
sources are frequently derived from in vitro cell cultures.
Therefore, further investigation is required to reveal the detailed
characteristics of ADEVs.
 | Table I.Summary of ADEV cargos and their
functions in recipient cells. |
Table I.
Summary of ADEV cargos and their
functions in recipient cells.
| A, ADEVs in
metabolic disorders |
|---|
|
|---|
| Origin | Cargo | Functions | (Refs.) |
|---|
| Adipose
tissue-EVs | N.D. | Promote M1
polarization of macrophages; Induce IR | (102) |
| Adipocyte-EVs | miR-34a | Inhibit M2
polarization of macrophages | (103) |
| Human
adipocyte-EVs | MIF, M-CSF,
TNF-α | Promote M1
polarization of macrophages | (86) |
| ATM-EVs | miR-155 | Reduce the insulin
sensitivity in adipocytes, the liver, and the muscle | (91) |
| Adipocyte-EVs | miR-27a | Induce hepatic and
skeletal muscle IR | (105,107) |
| ATM-EVs | miR-29a | Induce IR in
adipocytes, myocytes, and hepatocytes | (81) |
| WAT-EVs | miR-222 | Promote IR in the
liver and skeletal muscle | (106) |
| Human
adipocyte-EVs | miR-20b-5p | Impair insulin
action in skeletal muscle | (110) |
|
| B, ADEVs and
CVDs |
|
| Origin | Cargo |
Functions | (Refs.) |
|
| VAT-EVs | N.D. | Facilitate
macrophage foam cell generation and exacerbate atherosclerosis | (116) |
| Adipocyte-EVs | miRNAs | Increase macrophage
cholesterol efflux | (117) |
| 3T3-L1-EVs | miR-802-5p | Promote insulin
resistance in cardiomyocytes | (119) |
| Adipocyte-EVs | N.D. | Impair the function
of vascular endothelial cells | (121) |
| Adipocyte-EVs | N.D. | Promote the
attachment of leukocytes to endothelial cells | (122) |
| Adipocyte-EVs | miR-200a | Impair the function
of cardiomyocytes, and promote the process of cardiac
hypertrophy | (123) |
| Adipocyte-EVs | miR-130b-3p | Exacerbate ischemic
heart injury | (124) |
|
| C, ADEVs in
cancer |
|
| Origin | Cargo |
Functions | (Refs.) |
|
| Adipocyte-EVs | MMP3 | Promote lung tumor
metastasis | (128) |
| ADMSC-EVs | N.D. | Promote the
invasion, migration, and proliferation of osteosarcoma cells | (129) |
| ADSC-EVs | Angiogenic
proteins | Promote the
tube-like formation of endothelial cells | (130) |
| ADMSC-EVs | N.D. | Promote the
metastasis of breast cancer cells | (131) |
| Adipocyte-EVs | FA substrates,
proteins implicated in FAO | Promote lung tumor
metastasis | (90) |
| Adipocyte-EVs | FAO enzyme, FA
substrates | Trigger metabolic
remodeling, facilitate FAO and tumor aggressiveness | (78) |
Acknowledgments
Not applicable.
Funding
This work was supported by the National Natural Science
Foundation of China (grant no. 82000003), the Natural Science
Foundation of Zhejiang Province, China (grant no. LY23HO60009), the
Natural Science Foundation of Zhejiang Province, China (grant no.
LGF20H040009), and the China Postdoctoral Science Foundation (grant
no. 2020M671748).
Availability of data and materials
Not applicable.
Authors' contributions
XBY, JYH, and JL wrote the manuscript. XHK and XLL
conceived the subject of review and edited the manuscript. XHK
designed and created the schematic representations. Data
authentication is not applicable. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
EVs
|
extracellular vesicles
|
|
ADEVs
|
adipose tissue-derived extracellular
vesicles
|
|
T2D
|
type 2 diabetes
|
|
CVD
|
cardiovascular disease
|
|
AD
|
Alzheimer's disease
|
|
CH
|
cardiac hypertrophy
|
|
WATs
|
white adipose tissues
|
|
BATs
|
brown adipose tissues
|
|
BeATs
|
beige adipose tissues
|
|
MVs
|
microvesicles
|
|
ILVs
|
intraluminal vesicles
|
|
MVBs
|
multivesicular bodies
|
|
ncRNAs
|
noncoding RNAs
|
|
miRNAs
|
microRNAs
|
|
SCATs
|
subcutaneous ATs
|
|
DATs
|
dermal WATs
|
|
VAT
|
visceral ATs
|
|
ADSCs
|
adipose tissue-derived stem cells
|
|
ATMs
|
adipose tissue macrophages
|
|
IR
|
insulin resistance
|
|
GLUT4
|
glucose transporter 4
|
|
PPAR
|
peroxisome proliferator-activated
receptor
|
|
FAs
|
fatty acids
|
|
FAO
|
fatty acid oxidation
|
References
|
1
|
Mohammed MS, Sendra S, Lloret J and Bosch
I: Systems and WBANs for Controlling Obesity. J Healthc Eng.
2018:15647482018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kusminski CM, Bickel PE and Scherer PE:
Targeting adipose tissue in the treatment of obesity-associated
diabetes. Nat Rev Drug Discov. 15:639–660. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Abu-Farha M, Al-Mulla F, Thanaraj TA,
Kavalakatt S, Ali H, Abdul Ghani M and Abubaker J: Impact of
Diabetes in Patients Diagnosed With COVID-19. Front Immunol.
11:5768182020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Goodman KE, Magder LS, Baghdadi JD,
Pineles L, Levine AR, Perencevich EN and Harris AD: Impact of sex
and metabolic comorbidities on coronavirus disease 2019 (COVID-19)
mortality risk across age groups: 66 646 inpatients across 613 U.S.
Hospitals. Clin Infect Dis. 73:e4113–e4123. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Piroth L, Cottenet J, Mariet AS, Bonniaud
P, Blot M, Tubert-Bitter P and Quantin C: Comparison of the
characteristics, morbidity, and mortality of COVID-19 and seasonal
influenza: A nationwide, population-based retrospective cohort
study. Lancet Respir Med. 9:251–259. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ottaviani E, Malagoli D and Franceschi C:
The evolution of the adipose tissue: A neglected enigma. Gen Comp
Endocrinol. 174:1–4. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Unamuno X, Gomez-Ambrosi J, Rodriguez A,
Becerril S, Fruhbeck G and Catalan V: Adipokine dysregulation and
adipose tissue inflammation in human obesity. Eur J Clin Invest.
48:e129972018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Burhans MS, Hagman DK, Kuzma JN, Schmidt
KA and Kratz M: Contribution of adipose tissue inflammation to the
development of type 2 diabetes Mellitus. Compr Physiol. 9:1–58.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Berg AH, Combs TP, Du X, Brownlee M and
Scherer PE: The adipocyte-secreted protein Acrp30 enhances hepatic
insulin action. Nat Med. 7:947–953. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yamauchi T, Kamon J, Ito Y, Tsuchida A,
Yokomizo T, Kita S, Sugiyama T, Miyagishi M, Hara K, Tsunoda M, et
al: Cloning of adiponectin receptors that mediate antidiabetic
metabolic effects. Nature. 423:762–769. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hefetz-Sela S and Scherer PE: Adipocytes:
Impact on tumor growth and potential sites for therapeutic
intervention. Pharmacol Ther. 138:197–210. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Igata M, Motoshima H, Tsuruzoe K, Kojima
K, Matsumura T, Kondo T, Taguchi T, Nakamaru K, Yano M, Kukidome D,
et al: Adenosine monophosphate-activated protein kinase suppresses
vascular smooth muscle cell proliferation through the inhibition of
cell cycle progression. Circ Res. 97:837–844. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Douros JD, Baltzegar DA, Reading BJ, Seale
AP, Lerner DT, Grau EG and Borski RJ: Leptin stimulates cellular
glycolysis through a STAT3 dependent mechanism in Tilapia. Front
Endocrinol (Lausanne). 9:4652018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang Z and Xu A: Adipose extracellular
vesicles in intercellular and inter-organ crosstalk in metabolic
health and diseases. Front Immunol. 12:6086802021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Padilla J, Vieira-Potter VJ, Jia G and
Sowers JR: Role of perivascular adipose tissue on vascular reactive
oxygen species in type 2 diabetes: A give-and-take relationship.
Diabetes. 64:1904–1906. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kern PA, Ranganathan S, Li C, Wood L and
Ranganathan G: Adipose tissue tumor necrosis factor and
interleukin-6 expression in human obesity and insulin resistance.
Am J Physiol Endocrinol Metab. 280:E745–E751. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Keller S, Sanderson MP, Stoeck A and
Altevogt P: Exosomes: From biogenesis and secretion to biological
function. Immunol Lett. 107:102–108. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thomou T, Mori MA, Dreyfuss JM, Konishi M,
Sakaguchi M, Wolfrum C, Rao TN, Winnay JN, Garcia-Martin R,
Grinspoon SK, et al: Adipose-derived circulating miRNAs regulate
gene expression in other tissues. Nature. 542:450–455. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rome S, Blandin A and Le Lay S:
Adipocyte-Derived extracellular vesicles: State of the art. Int J
Mol Sci. 22:17882021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vidal M: Exosomes: Revisiting their role
as ‘garbage bags’. Traffic. 20:815–828. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
van der Pol E, Boing AN, Harrison P, Sturk
A and Nieuwland R: Classification, functions, and clinical
relevance of extracellular vesicles. Pharmacol Rev. 64:676–705.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tricarico C, Clancy J and D'Souza-Schorey
C: Biology and biogenesis of shed microvesicles. Small GTPases.
8:220–232. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cai H, Reinisch K and Ferro-Novick S:
Coats, tethers, Rabs, and SNAREs work together to mediate the
intracellular destination of a transport vesicle. Dev Cell.
12:671–682. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Escola JM, Kleijmeer MJ, Stoorvogel W,
Griffith JM, Yoshie O and Geuze HJ: Selective enrichment of
tetraspan proteins on the internal vesicles of multivesicular
endosomes and on exosomes secreted by human B-lymphocytes. J Biol
Chem. 273:20121–20127. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Giebel B and Helmbrecht C: Methods to
Analyze EVs. Methods Mol Biol. 1545:1–20. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wolf P: The nature and significance of
platelet products in human plasma. Br J Haematol. 13:269–288. 1967.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Al-Nedawi K, Meehan B, Micallef J, Lhotak
V, May L, Guha A and Rak J: Intercellular transfer of the oncogenic
receptor EGFRvIII by microvesicles derived from tumour cells. Nat
Cell Biol. 10:619–624. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Harding C, Heuser J and Stahl P:
Endocytosis and intracellular processing of transferrin and
colloidal gold-transferrin in rat reticulocytes: Demonstration of a
pathway for receptor shedding. Eur J Cell Biol. 35:256–263.
1984.PubMed/NCBI
|
|
29
|
Pan BT, Teng K, Wu C, Adam M and Johnstone
RM: Electron microscopic evidence for externalization of the
transferrin receptor in vesicular form in sheep reticulocytes. J
Cell Biol. 101:942–948. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kalra H, Drummen GP and Mathivanan S:
Focus on extracellular vesicles: Introducing the next small big
thing. Int J Mol Sci. 17:1702016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Minciacchi VR, Freeman MR and Di Vizio D:
Extracellular vesicles in cancer: Exosomes, microvesicles and the
emerging role of large oncosomes. Semin Cell Dev Biol. 40:41–51.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Raposo G and Stoorvogel W: Extracellular
vesicles: Exosomes, microvesicles, and friends. J Cell Biol.
200:373–383. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mulcahy LA, Pink RC and Carter DR: Routes
and mechanisms of extracellular vesicle uptake. J Extracell
Vesicles. 3:2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Llorente A, Skotland T, Sylvanne T,
Kauhanen D, Rog T, Orlowski A, Vattulainen I, Ekroos K and Sandvig
K: Molecular lipidomics of exosomes released by PC-3 prostate
cancer cells. Biochim Biophys Acta. 1831:1302–1309. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Laulagnier K, Javalet C, Hemming FJ,
Chivet M, Lachenal G, Blot B, Chatellard C and Sadoul R: Amyloid
precursor protein products concentrate in a subset of exosomes
specifically endocytosed by neurons. Cell Mol Life Sci. 75:757–773.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vargas A, Zhou S, Ethier-Chiasson M, Flipo
D, Lafond J, Gilbert C and Barbeau B: Syncytin proteins
incorporated in placenta exosomes are important for cell uptake and
show variation in abundance in serum exosomes from patients with
preeclampsia. FASEB J. 28:3703–3719. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kamerkar S, LeBleu VS, Sugimoto H, Yang S,
Ruivo CF, Melo SA, Lee JJ and Kalluri R: Exosomes facilitate
therapeutic targeting of oncogenic KRAS in pancreatic cancer.
Nature. 546:498–503. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van Niel G, D'Angelo G and Raposo G:
Shedding light on the cell biology of extracellular vesicles. Nat
Rev Mol Cell Biol. 19:213–228. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jeppesen DK, Fenix AM, Franklin JL,
Higginbotham JN, Zhang Q, Zimmerman LJ, Liebler DC, Ping J, Liu Q,
Evans R, et al: Reassessment of exosome composition. Cell.
177:428–445. e182019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Corrado C, Raimondo S, Saieva L, Flugy AM,
De Leo G and Alessandro R: Exosome-mediated crosstalk between
chronic myelogenous leukemia cells and human bone marrow stromal
cells triggers an interleukin 8-dependent survival of leukemia
cells. Cancer Lett. 348:71–76. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ailawadi S, Wang X, Gu H and Fan GC:
Pathologic function and therapeutic potential of exosomes in
cardiovascular disease. Biochim Biophys Acta. 1852:1–11. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
van Marken Lichtenbelt W: Brown adipose
tissue and the regulation of nonshivering thermogenesis. Curr Opin
Clin Nutr Metab Care. 15:547–552. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lee YH, Kim SN, Kwon HJ and Granneman JG:
Metabolic heterogeneity of activated beige/brite adipocytes in
inguinal adipose tissue. Sci Rep. 7:397942017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Keipert S and Jastroch M: Brite/beige fat
and UCP1 - is it thermogenesis? Biochim Biophys Acta.
1837:1075–1082. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang Z, Ning T, Song A, Rutter J, Wang QA
and Jiang L: Chronic cold exposure enhances glucose oxidation in
brown adipose tissue. EMBO Rep. 21:e500852020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Shamsi BH, Ma C, Naqvi S and Xiao Y:
Effects of pioglitazone mediated activation of PPAR-ү on CIDEC and
obesity related changes in mice. PLoS One. 9:e1069922014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Giampietro L, Gallorini M, De Filippis B,
Amoroso R, Cataldi A and di Giacomo V: PPAR-ү agonist GL516 reduces
oxidative stress and apoptosis occurrence in a rat astrocyte cell
line. Neurochem Int. 126:239–245. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jung SM, Sanchez-Gurmaches J and Guertin
DA: Brown adipose tissue development and metabolism. Handb Exp
Pharmacol. 251:3–36. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lau P, Tuong ZK, Wang SC, Fitzsimmons RL,
Goode JM, Thomas GP, Cowin GJ, Pearen MA, Mardon K, Stow JL and
Muscat GE: Roralpha deficiency and decreased adiposity are
associated with induction of thermogenic gene expression in
subcutaneous white adipose and brown adipose tissue. Am J Physiol
Endocrinol Metab. 308:E159–E171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wu J, Bostrom P, Sparks LM, Ye L, Choi JH,
Giang AH, Khandekar M, Virtanen KA, Nuutila P, Schaart G, et al:
Beige adipocytes are a distinct type of thermogenic fat cell in
mouse and human. Cell. 150:366–376. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ikeda K, Maretich P and Kajimura S: The
common and distinct features of brown and beige adipocytes. Trends
Endocrinol Metab. 29:191–200. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pinckard KM and Stanford KI: The
heartwarming effect of brown adipose tissue. Mol Pharmacol.
102:460–471. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Berbee JF, Boon MR, Khedoe PP, Bartelt A,
Schlein C, Worthmann A, Kooijman S, Hoeke G, Mol IM, John C, et al:
Brown fat activation reduces hypercholesterolaemia and protects
from atherosclerosis development. Nat Commun. 6:63562015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Altshuler-Keylin S, Shinoda K, Hasegawa Y,
Ikeda K, Hong H, Kang Q, Yang Y, Perera RM, Debnath J and Kajimura
S: Beige adipocyte maintenance is regulated by autophagy-induced
mitochondrial clearance. Cell Metab. 24:402–419. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rosen ED and Spiegelman BM: Adipocytes as
regulators of energy balance and glucose homeostasis. Nature.
444:847–853. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Arner P: Regional adipocity in man. J
Endocrinol. 155:191–192. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen SX, Zhang LJ and Gallo RL: Dermal
white adipose tissue: A newly recognized layer of skin innate
defense. J Invest Dermatol. 139:1002–1009. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang LJ, Guerrero-Juarez CF, Hata T,
Bapat SP, Ramos R, Plikus MV and Gallo RL: Innate immunity. Dermal
adipocytes protect against invasive Staphylococcus aureus skin
infection. Science. 347:67–71. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Fruhbeck G: Overview of adipose tissue and
its role in obesity and metabolic disorders. Methods Mol Biol.
456:1–22. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Salvador J, Silva C, Pujante P and
Fruhbeck G: Abdominal obesity: An indicator of cardiometabolic
risk. Endocrinol Nutr. 55:420–432. 2008.(In English, Spanish).
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Scheja L and Heeren J: The endocrine
function of adipose tissues in health and cardiometabolic disease.
Nat Rev Endocrinol. 15:507–524. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Stern JH, Rutkowski JM and Scherer PE:
Adiponectin, leptin, and fatty acids in the maintenance of
metabolic homeostasis through adipose tissue crosstalk. Cell Metab.
23:770–784. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Eto H, Suga H, Matsumoto D, Inoue K, Aoi
N, Kato H, Araki J and Yoshimura K: Characterization of structure
and cellular components of aspirated and excised adipose tissue.
Plast Reconstr Surg. 124:1087–1097. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zuk PA, Zhu M, Mizuno H, Huang J, Futrell
JW, Katz AJ, Benhaim P, Lorenz HP and Hedrick MH: Multilineage
cells from human adipose tissue: Implications for cell-based
therapies. Tissue Eng. 7:211–228. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Brown JC, Shang H, Li Y, Yang N, Patel N
and Katz AJ: Isolation of adipose-derived stromal vascular fraction
cells using a novel point-of-care device: Cell characterization and
review of the literature. Tissue Eng Part C Methods. 23:125–135.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wu H and Ballantyne CM: Metabolic
inflammation and insulin resistance in obesity. Circ Res.
126:1549–1564. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hollenberg CH and Vost A: Regulation of
DNA synthesis in fat cells and stromal elements from rat adipose
tissue. J Clin Invest. 47:2485–2498. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Panina YA, Yakimov AS, Komleva YK, Morgun
AV, Lopatina OL, Malinovskaya NA, Shuvaev AN, Salmin VV,
Taranushenko TE and Salmina AB: Plasticity of adipose
tissue-derived stem cells and regulation of angiogenesis. Front
Physiol. 9:16562018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Cao Y: Adipose tissue angiogenesis as a
therapeutic target for obesity and metabolic diseases. Nat Rev Drug
Discov. 9:107–115. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Mahlakoiv T, Flamar AL, Johnston LK,
Moriyama S, Putzel GG, Bryce PJ and Artis D: Stromal cells maintain
immune cell homeostasis in adipose tissue via production of
interleukin-33. Sci Immunol. 4:eaax04162019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Sun C, Berry WL and Olson LE: PDGFRα
controls the balance of stromal and adipogenic cells during adipose
tissue organogenesis. Development. 144:83–94. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Mclaughlin T, Ackerman SE, Shen L and
Engleman E: Role of innate and adaptive immunity in
obesity-associated metabolic disease. J Clin Invest. 127:5–13.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Rochette L, Mazini L, Malka G, Zeller M,
Cottin Y and Vergely C: The crosstalk of adipose-derived stem cells
(ADSC), oxidative stress, and inflammation in protective and
adaptive responses. Int J Mol Sci. 21:92622020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Hui X, Zhang M, Gu P, Li K, Gao Y, Wu D,
Wang Y and Xu A: Adipocyte SIRT1 controls systemic insulin
sensitivity by modulating macrophages in adipose tissue. EMBO Rep.
18:645–657. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Hotamisligil GS: Inflammation,
metaflammation and immunometabolic disorders. Nature. 542:177–185.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hotamisligil GS: Foundations of
immunometabolism and implications for metabolic health and disease.
Immunity. 47:406–420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Man K, Kutyavin VI and Chawla A: Tissue
immunometabolism: Development, physiology, and pathobiology. Cell
Metab. 25:11–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Clement E, Lazar I, Attane C, Carrie L,
Dauvillier S, Ducoux-Petit M, Esteve D, Menneteau T, Moutahir M, Le
Gonidec S, et al: Adipocyte extracellular vesicles carry enzymes
and fatty acids that stimulate mitochondrial metabolism and
remodeling in tumor cells. EMBO J. 39:e1025252020. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hartwig S, De Filippo E, Goddeke S, Knebel
B, Kotzka J, Al-Hasani H, Roden M, Lehr S and Sell H: Exosomal
proteins constitute an essential part of the human adipose tissue
secretome. Biochim Biophys Acta Proteins Proteom. 1867:1401722019.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Valadi H, Ekstrom K, Bossios A, Sjostrand
M, Lee JJ and Lotvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Liu T, Sun YC, Cheng P and Shao HG:
Adipose tissue macrophage-derived exosomal miR-29a regulates
obesity-associated insulin resistance. Biochem Biophys Res Commun.
515:352–358. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Geng L, Lam K and Xu A: The therapeutic
potential of FGF21 in metabolic diseases: From bench to clinic. Nat
Rev Endocrinol. 16:654–667. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Gao J, Li X, Wang Y, Cao Y, Yao D, Sun L,
Qin L, Qiu H and Zhan X: Adipocyte-derived extracellular vesicles
modulate appetite and weight through mTOR signalling in the
hypothalamus. Acta Physiol (Oxf). 228:e133392020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Lee JE, Moon PG, Lee IK and Baek MC:
Proteomic Analysis of extracellular vesicles released by adipocytes
of otsuka long-evans tokushima fatty (OLETF) Rats. Protein J.
34:220–235. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Sano S, Izumi Y, Yamaguchi T, Yamazaki T,
Tanaka M, Shiota M, Osada-Oka M, Nakamura Y, Wei M, Wanibuchi H, et
al: Lipid synthesis is promoted by hypoxic adipocyte-derived
exosomes in 3T3-L1 cells. Biochem Biophys Res Commun. 445:327–333.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Kranendonk ME, Visseren FL, van Balkom BW,
Nolte-'t Hoen EN, van Herwaarden JA, de Jager W, Schipper HS,
Brenkman AB, Verhaar MC, Wauben MH and Kalkhoven E: Human adipocyte
extracellular vesicles in reciprocal signaling between adipocytes
and macrophages. Obesity (Silver Spring). 22:1296–1308. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Phoonsawat W, Aoki-Yoshida A, Tsuruta T
and Sonoyama K: Adiponectin is partially associated with exosomes
in mouse serum. Biochem Biophys Res Commun. 448:261–266. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Crewe C and Scherer PE: Intercellular and
interorgan crosstalk through adipocyte extracellular vesicles. Rev
Endocr Metab Disord. 23:61–69. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Connolly KD, Wadey RM, Mathew D, Johnson
E, Rees DA and James PE: Evidence for adipocyte-derived
extracellular vesicles in the human circulation. Endocrinology.
159:3259–3267. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Lazar I, Clement E, Dauvillier S, Milhas
D, Ducoux-Petit M, Legonidec S, Moro C, Soldan V, Dalle S, Balor S,
et al: Adipocyte Exosomes Promote Melanoma Aggressiveness through
Fatty Acid Oxidation: A Novel Mechanism Linking Obesity and Cancer.
Cancer Res. 76:4051–4057. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ying W, Riopel M, Bandyopadhyay G, Dong Y,
Birmingham A, Seo JB, Ofrecio JM, Wollam J, Hernandez-Carretero A,
Fu W, et al: Adipose tissue macrophage-derived exosomal miRNAs Can
modulate in vivo and in vitro insulin sensitivity. Cell.
171:372–384.e12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Bassi EJ, Moraes-Vieira PM, Moreira-Sa CS,
Almeida DC, Vieira LM, Cunha CS, Hiyane MI, Basso AS, Pacheco-Silva
A and Camara NO: Immune regulatory properties of allogeneic
adipose-derived mesenchymal stem cells in the treatment of
experimental autoimmune diabetes. Diabetes. 61:2534–2545. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Mizuno H, Tobita M and Uysal AC: Concise
review: Adipose-derived stem cells as a novel tool for future
regenerative medicine. Stem Cells. 30:804–810. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Gonzalez MA, Gonzalez-Rey E, Rico L,
Buscher D and Delgado M: Adipose-derived mesenchymal stem cells
alleviate experimental colitis by inhibiting inflammatory and
autoimmune responses. Gastroenterology. 136:978–989. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Garcia-Contreras M, Vera-Donoso CD,
Hernandez-Andreu JM, Garcia-Verdugo JM and Oltra E: Therapeutic
potential of human adipose-derived stem cells (ADSCs) from cancer
patients: A pilot study. PLoS One. 9:e1132882014. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Zhao H, Shang Q, Pan Z, Bai Y, Li Z, Zhang
H, Zhang Q, Guo C, Zhang L and Wang Q: Exosomes from
adipose-derived stem cells attenuate adipose inflammation and
obesity through polarizing M2 macrophages and beiging in white
adipose tissue. Diabetes. 67:235–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Jung YJ, Kim HK, Cho Y, Choi JS, Woo CH,
Lee KS, Sul JH, Lee CM, Han J, Park JH, et al: Cell reprogramming
using extracellular vesicles from differentiating stem cells into
white/beige adipocytes. Sci Adv. 6:eaay67212020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Fuster JJ, Ouchi N, Gokce N and Walsh K:
Obesity-Induced changes in adipose tissue microenvironment and
their impact on cardiovascular disease. Circ Res. 118:1786–1807.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhao S, Kusminski CM and Scherer PE:
Adiponectin, leptin and cardiovascular disorders. Circ Res.
128:136–149. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Weisberg SP, Hunter D, Huber R, Lemieux J,
Slaymaker S, Vaddi K, Charo I, Leibel RL and Ferrante AJ: CCR2
modulates inflammatory and metabolic effects of high-fat feeding. J
Clin Invest. 116:115–124. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Lumeng CN, Bodzin JL and Saltiel AR:
Obesity induces a phenotypic switch in adipose tissue macrophage
polarization. J Clin Invest. 117:175–184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Deng ZB, Poliakov A, Hardy RW, Clements R,
Liu C, Liu Y, Wang J, Xiang X, Zhang S, Zhuang X, et al: Adipose
tissue exosome-like vesicles mediate activation of
macrophage-induced insulin resistance. Diabetes. 58:2498–2505.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Pan Y, Hui X, Hoo RLC, Ye D, Chan CYC,
Feng T, Wang Y, Lam KSL and Xu A: Adipocyte-secreted exosomal
microRNA-34a inhibits M2 macrophage polarization to promote
obesity-induced adipose inflammation. J Clin Invest. 129:834–849.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
James DE, Stockli J and Birnbaum MJ: The
aetiology and molecular landscape of insulin resistance. Nat Rev
Mol Cell Biol. 22:751–771. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Yu Y, Du H, Wei S, Feng L, Li J, Yao F,
Zhang M, Hatch GM and Chen L: Adipocyte-Derived exosomal MiR-27a
induces insulin resistance in skeletal muscle through repression of
PPARү. Theranostics. 8:2171–2188. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Li D, Song H, Shuo L, Wang L, Xie P, Li W,
Liu J, Tong Y, Zhang CY, Jiang X, et al: Gonadal white adipose
tissue-derived exosomal MiR-222 promotes obesity-associated insulin
resistance. Aging (Albany NY). 12:22719–22743. 2020.PubMed/NCBI
|
|
107
|
Kranendonk ME, Visseren FL, van Herwaarden
JA, Nolte-'t Hoen EN, de Jager W, Wauben MH and Kalkhoven E: Effect
of extracellular vesicles of human adipose tissue on insulin
signaling in liver and muscle cells. Obesity (Silver Spring).
22:2216–2223. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Gao X, Salomon C and Freeman DJ:
Extracellular vesicles from adipose tissue-A potential role in
obesity and type 2 diabetes? Front Endocrinol (Lausanne).
8:2022017. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Eguchi A, Lazic M, Armando AM, Phillips
SA, Katebian R, Maraka S, Quehenberger O, Sears DD and Feldstein
AE: Circulating adipocyte-derived extracellular vesicles are novel
markers of metabolic stress. J Mol Med (Berl). 94:1241–1253. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Katayama M, Wiklander OPB, Fritz T,
Caidahl K, El-Andaloussi S, Zierath JR and Krook A: Circulating
exosomal miR-20b-5p is elevated in type 2 diabetes and could impair
insulin action in human skeletal muscle. Diabetes. 68:515–526.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Kranendonk ME, de Kleijn DP, Kalkhoven E,
Kanhai DA, Uiterwaal CS, van der Graaf Y, Pasterkamp G and Visseren
FL; SMART Study Group, : Extracellular vesicle markers in relation
to obesity and metabolic complications in patients with manifest
cardiovascular disease. Cardiovasc Diabetol. 13:372014. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Su S, Guntur AR, Nguyen DC, Fakory SS,
Doucette CC, Leech C, Lotana H, Kelley M, Kohli J, Martino J, et
al: A renewable source of human beige adipocytes for development of
therapies to treat metabolic Syndrome. Cell Rep. 25:3215–3228,e9.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Connolly KD, Rees DA and James PE: Role of
adipocyte-derived extracellular vesicles in vascular inflammation.
Free Radic Biol Med. 172:58–64. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Dai W, Liu Z, Yang S and Kong J: Inflamed
adipose tissue: Therapeutic Targets for obesity-related endothelial
injury. Endocrinology. 164:bqad0942023. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Koenen M, Hill MA, Cohen P and Sowers JR:
Obesity, adipose tissue and vascular dysfunction. Circ Res.
128:951–968. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Xie Z, Wang X, Liu X, Du H, Sun C, Shao X,
Tian J, Gu X, Wang H, Tian J and Yu B: Adipose-Derived exosomes
exert proatherogenic effects by regulating macrophage foam cell
formation and polarization. J Am Heart Assoc. 7:e0074422018.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Barberio MD, Kasselman LJ, Playford MP,
Epstein SB, Renna HA, Goldberg M, Deleon J, Voloshyna I, Barlev A,
Salama M, et al: Cholesterol efflux alterations in adolescent
obesity: Role of adipose-derived extracellular vesical microRNAs. J
Transl Med. 17:2322019. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Fleury A, Martinez MC and Le Lay S:
Extracellular vesicles as therapeutic tools in cardiovascular
diseases. Front Immunol. 5:3702014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Wen Z, Li J, Fu Y, Zheng Y, Ma M and Wang
C: Hypertrophic adipocyte-derived exosomal miR-802-5p contributes
to insulin resistance in cardiac myocytes through targeting hSP60.
Obesity (Silver Spring). 28:1932–1940. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Monteiro JP, Bennett M, Rodor J,
Caudrillier A, Ulitsky I and Baker AH: Endothelial function and
dysfunction in the cardiovascular system: The long non-coding road.
Cardiovasc Res. 115:1692–1704. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Muller G: Microvesicles/exosomes as
potential novel biomarkers of metabolic diseases. Diabetes Metab
Syndr Obes. 5:247–282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Wadey RM, Connolly KD, Mathew D, Walters
G, Rees DA and James PE: Inflammatory adipocyte-derived
extracellular vesicles promote leukocyte attachment to vascular
endothelial cells. Atherosclerosis. 283:19–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Fang X, Stroud MJ, Ouyang K, Fang L, Zhang
J, Dalton ND, Gu Y, Wu T, Peterson KL, Huang HD, et al:
Adipocyte-specific loss of PPARү attenuates cardiac hypertrophy.
JCI Insight. 1:e899082016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Gan L, Xie D, Liu J, Bond LW, Christopher
TA, Lopez B, Zhang L, Gao E, Koch W, Ma XL and Wang Y: Small
extracellular microvesicles mediated pathological communications
between dysfunctional adipocytes and cardiomyocytes as a novel
mechanism exacerbating ischemia/reperfusion injury in diabetic
mice. Circulation. 141:968–983. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Parekh N, Chandran U and Bandera EV:
Obesity in cancer survival. Annu Rev Nutr. 32:311–342. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Dirat B, Bochet L, Dabek M, Daviaud D,
Dauvillier S, Majed B, Wang YY, Meulle A, Salles B, Le Gonidec S,
et al: Cancer-associated adipocytes exhibit an activated phenotype
and contribute to breast cancer invasion. Cancer Res. 71:2455–2465.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Nieman KM, Kenny HA, Penicka CV, Ladanyi
A, Buell-Gutbrod R, Zillhardt MR, Romero IL, Carey MS, Mills GB,
Hotamisligil GS, et al: Adipocytes promote ovarian cancer
metastasis and provide energy for rapid tumor growth. Nat Med.
17:1498–1503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Wang J, Wu Y, Guo J, Fei X, Yu L and Ma S:
Adipocyte-derived exosomes promote lung cancer metastasis by
increasing MMP9 activity via transferring MMP3 to lung cancer
cells. Oncotarget. 8:81880–81891. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Wang Y, Chu Y, Li K, Zhang G, Guo Z, Wu X,
Qiu C, Li Y, Wan X, Sui J, et al: Exosomes secreted by
adipose-derived mesenchymal stem cells foster metastasis and
osteosarcoma proliferation by increasing COLGALT2 expression. Front
Cell Dev Biol. 8:3532020. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Gangadaran P, Rajendran RL, Oh JM, Oh EJ,
Hong CM, Chung HY, Lee J and Ahn BC: Identification of angiogenic
cargo in extracellular vesicles secreted from human adipose
tissue-derived stem cells and induction of angiogenesis in vitro
and in vivo. Pharmaceutics. 13:4952021. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Khanh VC, Fukushige M, Moriguchi K,
Yamashita T, Osaka M, Hiramatsu Y and Ohneda O: Type 2 diabetes
mellitus induced paracrine effects on breast cancer metastasis
through extracellular vesicles derived from human mesenchymal stem
cells. Stem Cells Dev. 29:1382–1394. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Mattiske S, Suetani RJ, Neilsen PM and
Callen DF: The oncogenic role of miR-155 in breast cancer. Cancer
Epidemiol Biomarkers Prev. 21:1236–1243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Balaban S, Shearer RF, Lee LS, van
Geldermalsen M, Schreuder M, Shtein HC, Cairns R, Thomas KC,
Fazakerley DJ, Grewal T, et al: Adipocyte lipolysis links obesity
to breast cancer growth: Adipocyte-derived fatty acids drive breast
cancer cell proliferation and migration. Cancer Metab. 5:12017.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Kuo CY and Ann DK: When fats commit
crimes: Fatty acid metabolism, cancer stemness and therapeutic
resistance. Cancer Commun (Lond). 38:472018. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Zhuang X, Xiang X, Grizzle W, Sun D, Zhang
S, Axtell RC, Ju S, Mu J, Zhang L, Steinman L, et al: Treatment of
brain inflammatory diseases by delivering exosome encapsulated
anti-inflammatory drugs from the nasal region to the brain. Mol
Ther. 19:1769–1779. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Hardy J: Amyloid, the presenilins and
Alzheimer's disease. Trends Neurosci. 20:154–159. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Selkoe DJ: The cell biology of
beta-amyloid precursor protein and presenilin in Alzheimer's
disease. Trends Cell Biol. 8:447–453. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Katsuda T, Tsuchiya R, Kosaka N, Yoshioka
Y, Takagaki K, Oki K, Takeshita F, Sakai Y, Kuroda M and Ochiya T:
Human adipose tissue-derived mesenchymal stem cells secrete
functional neprilysin-bound exosomes. Sci Rep. 3:11972013.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Garcia-Contreras M and Thakor AS: Human
adipose tissue-derived mesenchymal stem cells and their
extracellular vesicles modulate lipopolysaccharide activated human
microglia. Cell Death Discov. 7:982021. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Geng W, Tang H, Luo S, Lv Y, Liang D, Kang
X and Hong W: Exosomes from miRNA-126-modified ADSCs promotes
functional recovery after stroke in rats by improving neurogenesis
and suppressing microglia activation. Am J Transl Res. 11:780–792.
2019.PubMed/NCBI
|
|
141
|
Jiang M, Wang H, Jin M, Yang X, Ji H,
Jiang Y, Zhang H, Wu F, Wu G, Lai X, et al: Exosomes from
MiR-30d-5p-ADSCs reverse acute ischemic stroke-induced,
autophagy-mediated brain injury by promoting M2
microglial/macrophage polarization. Cell Physiol Biochem.
47:864–878. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Takahashi Y and Takakura Y: Extracellular
vesicle-based therapeutics: Extracellular vesicles as therapeutic
targets and agents. Pharmacol Ther. 242:1083522023. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Greening DW, Xu R, Ale A, Hagemeyer CE and
Chen W: Extracellular vesicles as next generation
immunotherapeutics. Semin Cancer Biol. 90:73–100. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Kwan HY, Chen M, Xu K and Chen B: The
impact of obesity on adipocyte-derived extracellular vesicles. Cell
Mol Life Sci. 78:7275–7288. 2021. View Article : Google Scholar : PubMed/NCBI
|














