Rheumatoid arthritis (RA) is a chronic autoimmune
disease with chronic polyarticular inflammation as the main
clinical manifestation. The pathogenesis of RA is associated with
the production of autoantibodies such as anti-immunoglobulin G
(IgG) and citrullinated proteins (1). The main pathological features of RA
are synovial hyperplasia of the joint cavity, thickening of the
lining layer, formation of vascular opacities and infiltration of a
variety of autoimmune cells and activated inflammatory cells, which
in turn cause the destruction of cartilage and bone tissues,
ultimately leading to joint deformity and loss of function
(2,3).
The pathogenesis of RA involves dysregulation of
both intrinsic and adaptive immunity. In adaptive immunity, T cells
and B cells are involved to varying degrees in the pathogenesis of
RA. B cells accumulate in the synovium of inflamed RA and secrete
rheumatoid factor (RF) and anti-citrullinated protein antibodies
(ACPA), which are involved in the inflammatory process (4). Helper T (Th) cells are a diverse
group of CD4+ T cell subsets that play a crucial role in the immune
system. Intrinsic defects in the naive CD4+ T cells cause cellular
mis-differentiation, leading to irreversible tissue-tolerant
destruction as a key pathogenesis of RA (2).
A systematic analysis of global, regional and
national burden studies of RA from 1990–2017 showed that the global
age-standardized point prevalence and annual incidence of RA in
2017 were 246.6 [95% uncertainty interval (UI) of 222.4–270.8] and
14.9 (95% UI of 13.3–16.4), respectively, which increased by 7.4%
(95% UI 5.3–9.4) and 8.2% (95% UI 5.9–10.5) from 1990, respectively
(all estimates presented as counts and age-standardized rates per
100,000 population) (5). The data
in this systematic analysis also showed that the prevalence of RA
was highest in developed countries, followed by India and South
America, and appeared to be lower in rural areas compared with
urban settings, suggesting that population characteristics,
socioeconomic or environmental risk factors influence the
prevalence of RA (5). Women make
up the majority of patients with RA, with a ratio of up to 4:1 or
more to men (6–8). The influence of specific events such
as menopause, the number of parturiency and breastfeeding on the
risk of developing RA, as well as the sex ratio of patients with
RA, all contribute to the role of sex hormones in RA. This role has
been explored from as early as the last century and clinical trials
have confirmed the anti-inflammatory effects of sex hormones in RA
and the significant improvement of symptoms (9). However, hormone replacement therapy
(HRT) has since declined substantially due to potentially serious
side effects such as increased risk of coronary heart disease,
breast cancer and stroke (10,11).
Currently glucocorticoids are widely used with the treatment of RA,
but sex hormones have not been included in the treatment regimen
for RA. A recent study of female patients with RA treated with
tocilizumab, an IL-6 receptor antibody and/or traditional
disease-modifying anti-rheumatic drugs found that exogenous sex
hormone use appeared to be associated with higher remission rates
(12). Although epidemiologic
investigations and mechanistic studies remain contradictory, the
effect of sex hormones on RA is evident. Few articles have looked
at Th cells to illustrate the role of sex hormones in RA and
repairing Th cell defects during asymptomatic autoimmunity may be
the next cutting-edge intervention in RA. Exploring the effect of
sex hormones on RA from Th cells may be another research proposal
for using sex hormones to treat RA.
The present study used ‘rheumatoid arthritis’, ‘T
cells’ and ‘sex hormone’ as the key words and collected relevant
information from different databases, including PubMed (https://pubmed.ncbi.nlm.nih.gov), Web of Science
(https://ras.cdutcm.edu.cn:7080/s/cn/clarivate/webofscience/G.https/wos/woscc/basic-search),
Springer (https://link.springer.com), Science
Direct (https://www.sciencedirect.com), ACS
(https://pubs.acs.org/?locale=zh_CN),
Wiley (https://onlinelibrary.wiley.com) and CNKI (https://www.cnki.net). Inclusion criteria were:
Clinical studies, laboratory studies (including purely experimental
animal studies or in vitro cellular studies, combined animal
and in vitro cellular studies), meta-analyses and
bioinformatics studies validated with clinical or laboratory data,
regardless of the languages of publication. Exclusion criteria
were: Studies that were repeatedly published, clinical trials from
which no relevant data could be extracted, newspapers, conferences,
comments and other information without experimental data, studies
that were low quality or had incorrect assertions or conclusions
and different reports on the same issue (these have been retained
for comparison and discussion purposes).
The immunopathogenesis of RA spans decades and is
characterized by defective immune responses, primarily involving
pro-inflammatory cytokines and alterations in peripheral immune
tolerance, especially Th cells (13,14).
Genetic and environmental factors are major risk factors for RA and
shared epitope-positive HLA-DRB1 alleles and PTPN22 variants are
associated with the development of RF and ACPA (15,16).
Although these studies have taken less account of the effect that
other factors have on this process, they have revealed the
relationship between these genes and autoantibodies and the effect
of this process on RA. T cells recognize citrullinated antigens in
the context of HLA-DRB1*04 and B cells respond by producing large
amounts of citrullinated proteins (17,18).
B cells can also play an important role as antigen-presenting cells
(APCs) to self-reactive T cells (19,20).
There are several important stages in the transition from a healthy
state to clinical RA, which a high-quality review from Nature
Immunology categorized as the systemic breakdown of
self-tolerance, the transition from asymptomatic autoimmunity to
tissue inflammation and the transition from acute synovitis to
prolonged chronic synovitis (2).
Specifically, individuals with genetic and
environmental risks begin with the recognition of modified protein
antigens and the appearance of autoantibodies (phase I). Following
a prolonged period of asymptomatic autoimmunity and immune system
remodeling, cell-intrinsic changes in metabolic networks and DNA
instability drive T cell differentiation to tissue-invasive
short-term effector T cells and protective macrophage failure,
followed by disruption of tissue tolerance and the development of
early synovitis (phase II). The transformation of synovial stromal
cells to self-invasive effector cells transforms synovitis from
acute to chronic destructive, leading to destruction of articular
cartilage and bone (phase III). In the vast majority of cases, the
destruction of tissue tolerance is irreversible (2).
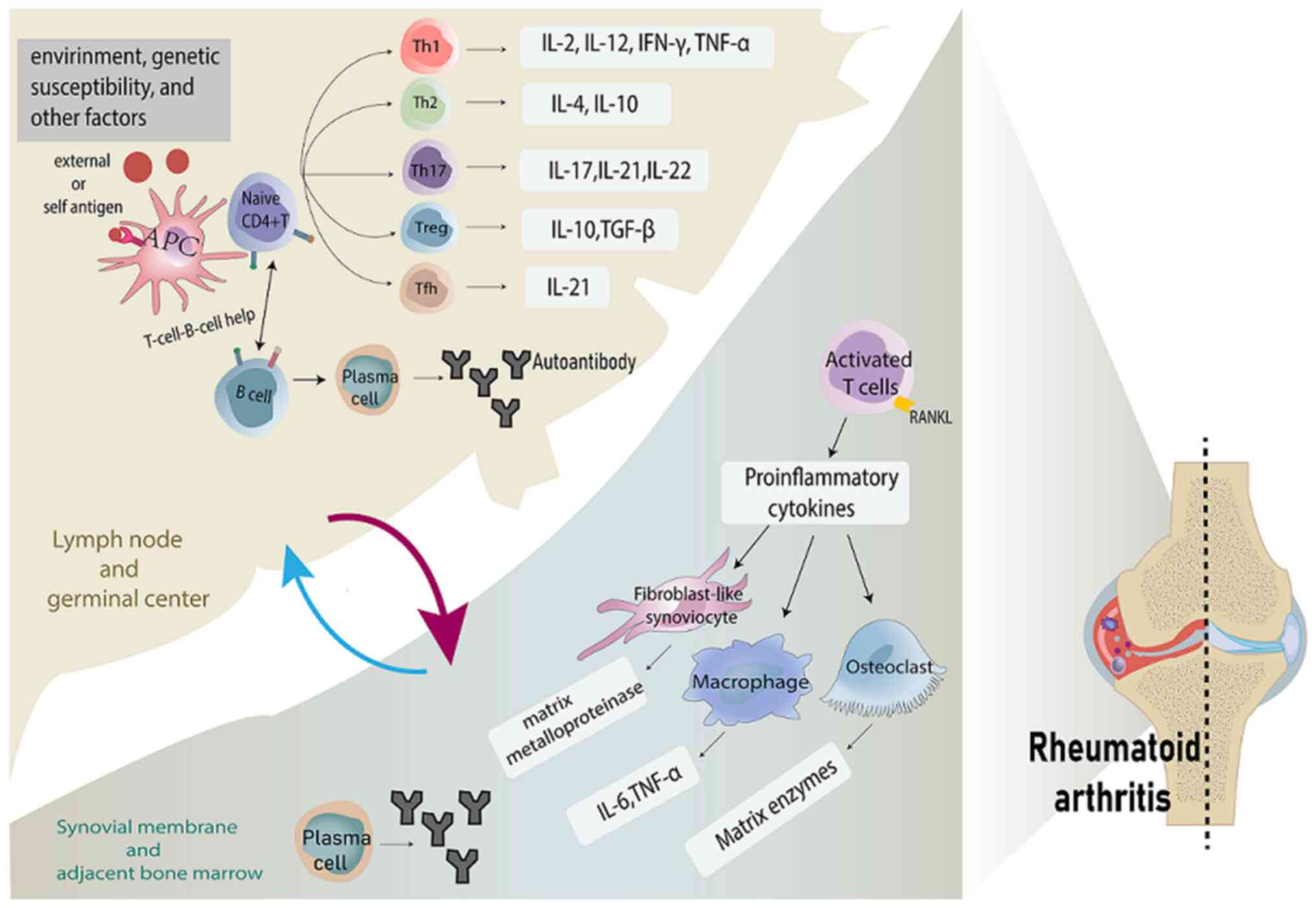
The second phase of RA is clinically marked by
synovial inflammation, which is closely related to cell-intrinsic
defects in CD4+ T cells, caused by mis-differentiation during the
conversion of initially resting CD4+ T cells into memory T cells
and effector T cell (21–23). Specifically, naive CD4+ T cells are
transformed into highly proliferative, tissue-invasive and
pro-inflammatory T cells rather than relatively quiescent memory T
cells; initial self-tolerance is disrupted, the disease process
shifts localization and a variety of immune cells infiltrate the
synovium (Fig. 1). Activated CD4+
T cells make up a large proportion of inflammatory cells in
synovial tissue and are involved in the pathologic process of RA. A
reduced naive CD4+ T cell frequency is the strongest predictor of
the development of synovitis in patients with ACPA-positive
populations (24).
Naive CD4+ T cells are activated and differentiated
into various Th cell subpopulations in response to antigenic
stimulation and cytokine signaling and their differentiation
process is dependent on the expression of specific transcription
factors induced by specific cytokines. These Th cells exert varying
degrees of disease-promoting or protective effects.
Initially, Th1 cells were hypothesized to play a
dominant role in the development of RA due to the high expression
levels of interleukin-12 (IL-12) and interferon-gamma (IFN-γ) found
at sites of inflammation and the positive correlation of IL-12
levels with disease activity observed in the serum and synovial
fluid of a significant proportion of patients with RA (25,26).
In addition, most CD4+ T cells infiltrating the synovium express
IFN-γ, which subsequently activates macrophages and induces tumor
necrosis factor-alpha (TNF-α) production (25). However, the Th1 phenotype does not
explain the full mechanism of RA development because of the higher
susceptibility to collagen-induced arthritis (CIA) in
IFN-γ-deficient mice and IFN-γ receptor-deficient mice (27,28),
as well as the lack of efficacy of IFN-γ monoclonal antibodies in
the majority of patients with RA (29).
The proposed Th1/Th2 model was used to explain RA
pathology in early studies. In arthritis dominated by Th1 cells,
IL-4 secreted by Th2 cells prevents disease and induces a switch
from a Th1-type to a Th2-type response. IL-4 deficiency is
essential for disease induction in animal models of CIA (30). Treatment of CIA mice using
mesenchymal stem cells (MSC) in combination with IL-4 restored
synovitis symptoms to the level of healthy controls (31). Later, the hypothesis that the
Th1/Th2 model is dominant began to change due to the emergence of
anti-inflammatory regulatory T (Treg) cells that produce growth
transformation factor-beta (TGF-β) and pro-inflammatory Th17 cells
that produce IL-17 (32).
Multiple molecules are involved in the
differentiation process of Th17 cells and the combination of these
molecules with each other generates pathogenic and non-pathogenic
Th17 cell subpopulations with varying degrees of pro- and
anti-inflammatory properties (33,34).
In patients with RA, both in vivo and in vitro
anti-TNF-α treatment induced the production of the
anti-inflammatory factor IL-10 by Th17 cells, suggesting that TNF-α
may have an inhibitory effect on IL-10 production by Th17 cells
(35). Although Th17 cells and
their effector molecules play a positive role in the maintenance of
immune homeostasis in the body, they are also involved in the
pathology of a variety of autoimmune diseases. Peripheral blood
mononuclear cells (PBMC) from patients with RA had a large number
of Th17 cells and their proportion correlated with the progression
and activity of RA (36,37). IL-17-deficient and blocked mice
exhibited resistance to CIA while ameliorating disease severity in
a mouse model of CIA (38,39). IL-22 is an important cytokine
produced by Th17 cells and IL-22(−/-) mice had severely reduced
splenic germinal centers and their CIA severity was also
significantly reduced (40).
Th17 cells expressing the receptor activator of
NF-κB ligand (RANKL), an osteoclast-activating factor, stimulate
localized bone resorption by regulating the migratory state and
functional changes of mature osteoclasts through cell-to-cell
contacts and this may be the mechanism by which Th17 cells mediate
inflammatory bone destruction (41). As IL-17 blockade has therapeutic
potential for RA and clinical trials have been conducted to explore
the feasibility of this treatment approach. For instance, a study
has shown that IL-17 blocking drugs such as secukinumab and
ixekizumab are effective in the treatment of RA, but their efficacy
appears to be inferior to that of already existing biologics such
as anti-TNF-α and anti-IL-6 (42).
Treg cells play a crucial role in maintaining
autoimmune tolerance by secreting the anti-inflammatory cytokine
IL-10 and the immunosuppressive molecule TGF-β (43,44).
However, the number of Treg cells in the synovial fluid of patients
with RA with persistent joint inflammation is significantly
increased, suggesting that Treg cell function is altered in
patients with RA (45). This is
supported by other studies which have found that Treg cell
regulation is reduced in both peripheral blood and synovium in
patients with RA (46–48). Forkhead box protein 3 (Foxp3) is a
key transcription factor for Treg cells and TNF-α dephosphorylation
inhibits the transcriptional activity of Foxp3, disabling Foxp3+
Treg cells in patients with RA (46). In arthritic conditions,
CD25(lo)Foxp3(+) T cells lose Foxp3 expression and
transdifferentiate into inflammatory Th17 cells (49). The regulatory capacity of Treg
cells in an inflammatory environment is reduced or lost, so that
Treg cells isolated from active patients with RA cannot prevent
effector T cells from secreting pro-inflammatory cytokines such as
IFN-γ and TNF-α (47,48), which may be a vicious cycle. In
addition, studies have shown that MSC has a favorable effect on the
regulatory function of Treg cells in the context of RA (50,51).
Tfh cells are characterized by high expression of
the chemokine receptor C-X-C motif chemokine receptor 5, the
transcription factor Bcl6, inducible co-stimulatory molecule and
the co-inhibitory molecule programmed cell death protein 1 (PD-1).
Once the naive CD4+ T cells are activated by APCs, IL-6 and IL-21,
they differentiate into Tfh cells (52). Tfh cells help B cells produce
relevant antibodies in patients with RA. Transplantation of MSC
into CIA mice prevented the progression of arthritis by suppressing
the number and function of Tfh cells (53). patients with RA express high levels
of IL-21 in serum (54,55) and the percentage of Tfh cells is
also positively correlated with disease activity (55). After 1 month of drug treatment, the
percentage of PD-1+ Tfh cells was significantly decreased in
drug-responsive patients with RA (55). In the synovium of IL-21R-deficient
RA model mice, fewer Tfh cells and more Th17 cells were found in
comparison with the control, as well as low levels of RANKL
expression, suggesting that IL-21 plays an important role in the
disease process of RA through Tfh cell proliferation and RANKL
induction rather than Th17 cell function (56,57).
The thymus is a key site for the production of a
number of types of T cells and the effects of sex hormones on the
thymus were recognized as early as the last century when it was
observed that the thymus is enlarged in male castrated mice and
shrinks following the administration of androgens (58). Compared with men, women have a
higher absolute number of CD4+ T cells and show stronger T-cell
responses and B-cell responses to antigens, which contributes to
autoimmunity (59). Women are more
inclined to develop T-cell-mediated autoimmune diseases, whereas
men are more susceptible to cancer and infectious diseases
(60), which has been commonly
described as an immunostimulatory effect of estrogens and an
immunosuppressive effect of androgens, which is clearly an
oversimplified description. The effect of sex hormones on the
immune system and immune disorders is complex and can be influenced
by a combination of factors such as genetics, the relative
expression of hormone receptor subtypes, hormone concentrations,
the tissues in which they are found and the stage of life (Table I).
Women have three main natural estrogens, estrone,
estradiol (E2) and estriol, with E2 being its most potent form. In
studying the role of estrogen in autoimmune diseases, Cutolo et
al (61) conclude that
estrogen may promote B-cell-driven diseases, while T-cell-driven
diseases may be inhibited by estrogen. Estrogen is mainly mediated
by two specific intracellular receptors namely estrogen receptors
(ER) α and β and a membrane G-protein coupled receptor. There are
significant differences in the distribution of ERα and ERβ in
immune cells and tissues and the complex effects of estrogen on the
immune system are partly due to the fact that the expression of one
receptor relative to the other may alter the action of estrogen
(62). For example, in a mouse
model of systemic lupus erythematosus (SLE), ERα has significant
pro-inflammatory effects, whereas ERβ exhibits anti-inflammatory
and immunosuppressive effects (63–65).
ERβ expression is also significantly reduced in T cells from
patients with SLE and inflammatory bowel disease (66,67).
In addition, it has been found that ERα controls the production of
autoantibodies to prevent autoimmunity by inhibiting the reaction
of Tfh cells (68), while ERβ
enhances immunosuppression by promoting the differentiation of Treg
cells (69).
In peripheral T cells, low concentrations of E2
induce T cells to express T-box transcription factor expressed in T
cells (T-bet) and IFN-γ to promote the Th1 response and ERα, but
not ERβ, is required for this process (70). E2 levels are consistently elevated
during pregnancy and high concentrations of E2 affect CD4+ T cell
polarization by enhancing the expression of Th2-related genes
(GATA3 and IL-4) and Treg-related genes (Foxp3, IL-10 and TGF-β),
while inhibiting the expression of Th1-related genes (T-bet, IL-2,
TNF-α and IFN-γ) and Th17-related genes (ROR-γ T, IL-6, IL-17 and
IL-23) expression (71–74). By contrast, the decline in ovarian
function and rapid decline in circulating estrogen during menopause
are associated with an increase in pro-inflammatory cytokines such
as IL-6, TNF-α and IL-1β (75,76).
The addition of E2 at ovulation level to PBMC of postmenopausal
women in vitro can inhibit the release of these
pro-inflammatory factors (77).
Androgens mainly include testosterone,
dihydrotestosterone and dehydroepiandrosterone (DHEA) and the
biological actions of androgens are mediated by the androgen
receptor (AR) (78). Androgens
have recognized immunosuppressive effects, in large part because of
their ability to inhibit cytokine production and impair T cell
effector activity (79,80).
The autoimmune regulatory gene (AIRE) provides
strong protection against autoimmune disease and the effect of
androgens on them is stronger in males than in females (81). Androgens recruit AR to the AIRE
promoter region, enhancing AIRE transcription, improving immune
tolerance and producing more effective negative selection of
self-reactive T cells (82). By
contrast, estrogen promotes autoimmunity by decreasing AIRE
expression through ERα upregulation of AIRE methylation (83). The number of Treg cells is
significantly higher in men than in women (84) and the androgen/AR pathway can
stabilize the inhibitory function of Treg cells by enhancing Foxp3
expression (85). In addition,
androgens regulate Th2 cytokine production and mRNA levels of IL-4
and IL-10 are reduced in gonadectomized male mice (86), but in Th2 cell-mediated allergic
asthma, the androgen/AR pathway restricts Th2 cytokine release
(87,88). Androgens/AR may inhibit the
differentiation of CD4+ T cells into Th1 cells through a direct
increase in protein tyrosine phosphatase non-receptor type 1, a
phosphatase that inhibits Th1 differentiation, thus effectively
preventing male autoimmunity (89). Androgen deficient men also showed
higher levels of IL-1β, IL-2, TNF-α and CD4+/CD8+ T cell ratios
(90). In turn, TNF-α has been
shown to have an inhibitory effect on testosterone synthesis in the
testis by inhibiting important enzymatic steps in the adrenal and
gonads (91).
Progesterone (Pg) is a female sex hormone that plays
a key role in establishing and maintaining the pregnancy process
and has powerful immunomodulatory properties (92). T cells express Pg receptors (PR-a,
PR-b and PR-c) and membrane Pg receptors (mPRa, mPRb and mPRg)
(93). Pg reduces T cell
proliferation by impairing the ability of dendritic cells to
stimulate T cell proliferation (94). In mouse experiments, administration
of a comparable dose of Pg in midgestation promotes CD4+CD25+ Treg
cell proliferation and enhances the inhibitory function of
CD4+CD25+Treg cell by increasing IL-10 expression (95). High levels of Pg during pregnancy
selectively induce conventional CD4+ T cell death and CD4 + Treg
cell enrichment (96). In human
studies, Pg enhances anti-inflammatory responses and immune
tolerance by inducing Th2 and Treg cell subsets during pregnancy
(97). In addition, Pg inhibits
the differentiation of naive CD4+ T cells to Th1 and Th17 cells
(98,99).
Prolactin (PRL) is also involved in immunomodulation
and has been shown to favor the survival and differentiation of
T-cell progenitors (100). PRL
reduces the inhibitory function of Treg cells (101) and SLE patients have elevated PRL
receptor expression in Treg cells but decreased Treg cell
percentage and function (100).
It was observed that co-culturing T cells with PRL led to more
production of Th1-related cytokines such as TNF-α, IFN-γ and IL-2
(102).
Human chorionic gonadotropin (HCG) peaks in early
pregnancy. Studies demonstrate that HCG regulates Th1/Th2 balance
(103), interferes with Th17
differentiation and induces its anti-inflammatory profile,
increases the frequency of Treg cells (104) and enhances the inhibitory
capacity of Tregs by increasing the secretion of IL-10 and TGF-β
(105–107). Follicle stimulating hormone (FSH)
is closely related to female ovulation and its level directly
reflects the ovarian function status. Studies have found that FSH
promotes the high expression of pro-inflammatory cytokines IL-1β,
IL-6 and TNF-α (108,109).
Regardless of sex, decreased androgen levels and
increased estrogen levels are found in the synovial fluid of
patients with RA (112,113). This phenomenon may be due to the
activation of peripheral tissue aromatase stimulated by elevated
inflammatory factors (TNF-α, IL-6 and IL-1) in the synovium and
promotes the peripheral conversion of androgens to estrogens
(114,115). Injection of glucocorticoids into
the joints suppresses their serum estrogen androgen levels compared
with baseline before injection therapy, but this tends to be
reversible (116). Other studies
have shown that in synoviocytes from patients with RA, estrogen
hydroxylated metabolites have pro-inflammatory activity and
pathogenic effects that cause synovial hyperplasia (117,118). Although these reports show a
pro-RA effect of estrogen, a growing body of research suggests that
the protective effect of estrogen on RA is dose-dependent and that
estrogen deficiency is more likely to bring about joint
inflammation and cause bone erosion. For example, estrogen
deficiency induces T cell proliferation and prolongs active T-cell
lifespan via IFN-γ, which leads to bone loss (119). Ovariectomized (OVX) mice have
normal T cell numbers in the bone marrow, but have more
RANKL-expressing CD3+ T cells and B cells (120). In MRL/lpr mice (with a genetic
susceptibility similar to human RA), estrogen deficiency may induce
the activation of RANKL-carrying CD4+ T cells, leading to
osteoclastogenesis and bone resorption (121). Treatment of CIA mice with E2 led
to a decrease in the number of Th17 cells and IL-17(+) γδ T cells
in the joints but an increase in the number of Th17 cells and
IL-17(+) γδ T cells in the draining lymph nodes, suggesting that E2
mediates the prevention of migration of these two types of cells
from the lymph nodes to the joints (122). In addition, estrogen antagonizes
acid-sensing ion channel 1α-induced mitochondrial stress and
protects against cartilage damage in OVX rats with adjuvant
arthritis (123).
As aforementioned, the immunizing effects of
estrogen are closely related to the relative expression of ER. The
expression of ER mRNA in synovial tissues of patients with RA was
significantly higher than that in healthy non-inflammatory synovial
tissues and the relative expression ratio of ERα/ERβ mRNA was
significantly lower (124).
Treatment with E2 improved synovial inflammation and joint
destruction in mice with arthritis and reduced the number of Th17
cells in the joints, both of which are dependent on Erα (125,126). Recent studies have also revealed
the effects of ERβ on bone. Xu et al (127) show that bone mass decreases in
male mice with osteoblast ERβ deletion, but has no effect on bone
mass in female mice, suggesting that the mechanism by which
osteoblast ERβ regulates bone modeling varies by sex.
Androgens are generally considered to be a natural
anti-inflammatory agent with immunosuppressive effects (79,80).
Serum androgen levels and synovial androgen metabolism levels are
decreased in patients with RA regardless of sex (113,128) and serum androgen levels are
negatively correlated with RA disease activity (129,130). Therefore, the deficiency of
androgen regulating the immune system is considered to be related
to the onset of RA. Androgen deprivation has been found to increase
the risk of RA when androgen deprivation is administered to
patients with prostate cancer (131,132) and the longer the duration of
treatment deprivation, the higher the risk (131). In terms of improving clinical and
chemical indicators of immune response in men with RA, several
studies have demonstrated the beneficial effects of testosterone
therapy (129,133). Animal studies have also shown
that both physiologic and pharmacologic concentrations of
testosterone produce anti-inflammatory effects on joint
inflammation in rats (134,135). In addition, polarization of RA
synovial macrophages activates intracellular androgens, which
contribute to the suppression of local inflammation (136). Anti-TNF-α and anti-IL-6 treatment
of synoviocytes from patients with RA both attenuate androgen
suppression (137,138) and methotrexate (MTX) treatment
also improves testosterone levels in rats with adjuvant-induced
arthritis (139).
Other studies have also provided evidence that
androgens ameliorate RA, such as the study by Stark et al
(140) which found that the
cytochrome B5 type A (CYB5A) single nucleotide polymorphism (SNP)
increases androgens in women and is associated with a reduced
genetic risk of RA in women, but the CYB5A SNP is not associated
with RA risk in men. Overexpression of CRF6-interacting factor 1, a
nuclear protein that interacts with the AR, was found by Park et
al (141) to attenuate
activation of Th17 cells and osteoclast differentiation and reduce
arthritic symptoms and histological manifestations in CIA mice. By
inhibiting cellular immunity and autoantibody formation, exogenous
DHEA ameliorated the severity of acute and chronic antigen-induced
arthritis in mice (142).
However, DHEA treatment at 50 mg/day for 12 consecutive weeks did
not show any greater improvement in patients with RA compared with
the placebo group (143).
Studies using next-generation sequencing show that
Pg-induced transcriptomic changes are significantly enriched in
genes associated with pregnancy-regulated diseases (for example,
multiple sclerosis, RA and psoriasis), suggesting a potential role
for Pg in the immunomodulation of pregnancy-induced diseases
(144). Experimental studies have
demonstrated the inhibitory effect of Pg on matrix
metalloproteinase activity produced by fibroblast-like synoviocytes
(145). M2000 is a novel
nonsteroidal anti-inflammatory drug with immunosuppressive effects
that improves clinical symptoms and increases serum Pg levels in
patients with RA (146).
Luteinizing hormone (LH) levels are significantly
lower in both male and postmenopausal female patients with RA
compared with healthy controls (147–149), but are not associated with
disease activity (148). In a
study using the gonadotropin-releasing hormone antagonist ASP1707
in combination with MTX for the treatment of RA, a 90% decrease in
plasma LH concentrations was found in 90% of patients treated with
ASP1707, but no clinical benefit was demonstrated and there were no
significant changes in levels of TNF-α, matrix metalloproteinase 3
and IL-6 (150).
FSH levels increase during the perimenopausal period
due to negative feedback in the gonadal axis (151) and FSH secretion promotes the
production of TNF-α, which increases the number of osteoclast
precursors in the bone marrow (103), but this does not affected
osteoclastogenesis (152). High
FSH levels have been found to be associated with an increased risk
of RA and are positively correlated with RA disease activity
(153). However, earlier studies
have shown that serum FSH is significantly lower in postmenopausal
women with RA compared with healthy controls (149).
Following menopause, estrogen levels plummet and
testosterone levels begin to decline and postmenopausal women have
a higher pro-inflammatory immune status (157). The peak incidence of RA in women
is roughly during their menopausal years (158) and menopausal status is associated
with the progression of joint function decline and deterioration
(159). Early age at menopause
has long been recognized to be associated with an increased risk of
RA (160,161) and women with menopausal age in
their 40s have more than double the risk of RA (160). Postmenopausal women are at
increased risk of ACPA-positive RA, especially in early menopause
when estrogen plummets (162,163). Treatment with E2 in
postmenopausal women with RA significantly increased the salivation
of crystallizable fragments of IgG and induced its capacity for
anti-inflammatory effects (164).
Pregnancy has been found to have a protective effect
on the development and disease activity of RA, with any number of
births being significantly associated with a reduced risk of RA,
but no such protective effect was found in nulliparous women
(165), which, as previously
described, is perhaps associated with high estrogen and Pg
concentrations (166,167). A large population-based Swedish
study showed that ACPA-positive patients are unlikely to experience
disease improvement from pregnancy (168), possibly because pregnancy in
ACPA-positive patients failed to cause elevated ACPA-IgG
galactosylation (169). CD4+CD25+
Treg cell levels are significantly higher in patients with RA
during pregnancy than at 8 weeks postpartum and CD4+CD25+ Treg cell
frequency is negatively correlated with RA disease activity in both
periods (170). Experimental
studies demonstrate that pregnancy protects mice from CIA, with
Treg cells playing a considerable role and that transfer of Treg
cells from pregnant ‘protected’ mice is sufficient to confer
protection to non-pregnant mice (171). Fertility is affected in patients
with RA (172,173) and anti-Müllerian hormone is
currently the most reliable biomarker of ovarian reserve. A number
of studies have demonstrated that serum anti-Müllerian hormone
levels are not reduced in patients with RA compared with healthy
controls (174,175), suggesting that reduced fertility
in patients with RA may not be caused by decreased ovarian reserve
function.
Studies on the role of breastfeeding in RA show
conflicting results. The incidence of RA is significantly higher in
postpartum breastfeeding women, with ~90% of patients experiencing
an onset in the first 3 months postpartum and a decrease in the
following 9 months, which also suggests that elevated PRL is
associated with episodes and recurrences of RA (154,176). However, prolonged breastfeeding
(>12 months) has been found to be associated with a reduced risk
of RA (177,178). A subsequent systematic review and
meta-analysis covering 1,672 RA cases from six studies show that
breastfeeding is associated with a lower risk of RA regardless of
whether breastfeeding is longer or shorter than 12 months (179). Hormone levels fluctuate greatly
in postpartum women, with a sudden drop in estrogen and Pg and an
increase in PRL levels and coupled with the fact that PRL has been
little studied in RA, there is still a large gap to be filled in
terms of the effects of breastfeeding on RA and its causes.
A study based on an epidemiologic survey of RA
evaluating the relationship between postmenopausal HRT use and RA
risk showed a reduced risk of ACPA-positive RA but no association
with ACPA-negative RA in postmenopausal women who used HRT compared
with those who did not (180). OC
has also shown a protective effect against ACPA-positive RA only
and the earlier the first exposure to OC, the lower the odds ratio
for RA (180,181). No correlation with the risk of
ACPA-negative RA is found for either HRT or OC (180,182) and these differences suggest the
existence of different hormone-related etiologic pathologies for
ACPA-positive and ACPA-negative RA. However, these results have not
been fully harmonized, as a meta-analysis of 17 studies showed that
OC did not provide protection against the risk of RA in women
(183) and a number of large
cohort studies have failed to identify an association between OC or
HRT use and the risk of RA (177,184,185).
The present study found that androgens and Pg had a
more definite inhibitory effect on immune response and RA compared
with other sex hormones, whereas the protective effect of estrogen
on RA appears to be dose-dependent, but the sex differences and
specific mechanisms of action of these hormones remain to be
investigated. These hormones regulate immune and inflammatory
responses by modulating CD4+ T-cell differentiation to promote the
balance between Th1/Th2 and Th17/Treg cells and altering Th-cell
function and alleviate the joint symptoms of RA by modulating the
balance between osteoclasts and osteoblasts to ameliorate bone
destruction. From epidemiologic investigations of female-specific
events on the risk of RA and mechanistic studies of RA-hormones, it
is not difficult to find a common feature that an acute decline in
ovarian function and estrogen drives RA progression. However, the
role of estrogen combined with Pg in RA is controversial, as not
all studies have shown favorable results (9,181,186). Finally, little is known about the
role of hormones such as LH and FSH in the RA disease process; are
their effects and mechanisms of action on RA worthy of
attention?
In addition, studies of hormone therapy for RA are
quite limited and the findings are inconclusive. Regardless of sex,
androgen-assisted treatment of RA has a limited effect on disease
activity, but brings improvements in quality of life (143,187–189). The addition of estrogen to
pre-existing therapy in postmenopausal women with RA seems to bring
some relief from the disease, but this is associated with serum
estrogen levels (9,190) and more studies show that estrogen
is well suited to ameliorate bone loss and increase bone density in
postmenopausal women with RA (9,191–193). In addition, two studies of the
gonadotropin-releasing hormone antagonist, cetrorelix, for the
short-term treatment of RA found that cetrorelix has a rapid
anti-inflammatory effect, but it is only in patients with RA and
with high gonadotropin levels that adjunctive treatment with
cetrorelix can show a rapid improvement in the disease (194,195).
Differences in hormone levels and regulation
between men and women, as well as multiple life stages and specific
events in women, have made the study of sex hormones in disease
difficult and complicated the findings, with high or low levels of
a particular hormone appearing to be associated with disease. In
addition, the level of expression of the hormone receptor proteins
may also cause the hormones to act differently and it needs to be
verified whether these differences in action have an effect on RA
pathogenesis or, conversely, whether RA activity causes changes in
the expression of the hormone receptor proteins.
The failure of T cell tolerance is attributed to
endogenous cellular abnormalities already present in the naive T
cells that shift the differentiation program to favor the
production of short-term effector T cells over long-lived memory T
cells. Current therapeutic strategies for RA are focused on
controlling inflammation and by recognizing that RA undergoes a
period of relatively stable impaired immune tolerance prior to the
onset of inflammation and the associated molecular features of this
period, it is possible to identify upstream therapeutic targets
that can abort the disease process before irreversible tissue
damage occurs. The role of sex hormones in the immune system was
recognized at an early stage and it is now clear that Th cell
responses are regulated by sex hormone levels and that sex hormones
not only directly affect T cell transcriptional profiles, but also
influence T cell responses and alter CD4+ cell differentiation by
controlling gene expression in thymic epithelial cells and
regulating innate immune cells (196). However, despite the a number of
advances in the study of the effects of sex hormones on T cells,
there are still a number of unanswered questions, especially in RA,
where epidemiologic investigations are still contradictory and
controversial and the study of the mechanisms of sex hormone
effects on Th cells has not been carried out extensively or
intensively.
Nonetheless, existing research has also revealed
new areas for combating RA. Future studies may be able to devise a
way to limit the targets of action of sex hormones to make T cell
differentiation more stable or to correct mis-differentiation,
which would be an important step in the effective use of sex
hormones as an immunotherapy, but also a lengthy process that needs
to be precisely controlled.
Not applicable.
Funding: No funding was received.
Not applicable.
QN wrote and revised the manuscript; JH reviewed
and edited the manuscript; ZL reviewed and edited the manuscript;
HZ conceptualized the article and reviewed the manuscript. All
authors have read and approved the final version of the manuscript.
Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Malmstrom V, Catrina AI and Klareskog L:
The immunopathogenesis of seropositive rheumatoid arthritis: From
triggering to targeting. Nat Rev Immunol. 17:60–75. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weyand CM and Goronzy JJ: The immunology
of rheumatoid arthritis. Nat Immunol. 22:10–18. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scherer HU, Haupl T and Burmester GR: The
etiology of rheumatoid arthritis. J Autoimmun. 110:1024002020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu F, Gao J, Kang J, Wang X, Niu Q, Liu J
and Zhang L: B Cells in Rheumatoid Arthritis: Pathogenic mechanisms
and treatment prospects. Front Immunol. 12:7507532021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Safiri S, Kolahi AA, Hoy D, Smith E,
Bettampadi D, Mansournia MA, Almasi-Hashiani A, Ashrafi-Asgarabad
A, Moradi-Lakeh M, Qorbani M, et al: Global, regional and national
burden of rheumatoid arthritis 1990–2017: A systematic analysis of
the Global Burden of Disease study 2017. Ann Rheum Dis.
78:1463–1471. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Galarza-Delgado DA, Azpiri-Lopez JR,
Colunga-Pedraza IJ, Cárdenas-de la Garza JA, Vera-Pineda R,
Wah-Suárez M, Arvizu-Rivera RI, Martínez-Moreno A, Ramos-Cázares
RE, Torres-Quintanilla FJ, et al: Prevalence of comorbidities in
Mexican mestizo patients with rheumatoid arthritis. Rheumatol Int.
37:1507–1511. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cardiel MH, Pons-Estel BA, Sacnun MP,
Wojdyla D, Saurit V, Marcos JC, Pinto MR, Cordeiro de Azevedo AB,
da Silveira IG, Radominski SC, et al: Treatment of early rheumatoid
arthritis in a multinational inception cohort of Latin American
patients: The GLADAR experience. J Clin Rheumatol. 18:327–335.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Castillo-Canon JC, Trujillo-Caceres SJ,
Bautista-Molano W, Valbuena-García AM, Fernández-Ávila DG and
Acuña-Merchán L: Rheumatoid arthritis in Colombia: A clinical
profile and prevalence from a national registry. Clin Rheumatol.
40:3565–3573. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
D'elia HF, Larsen A, Mattsson LA,
Waltbrand E, Kvist G, Mellström D, Saxne T, Ohlsson C, Nordborg E
and Carlsten H: Influence of hormone replacement therapy on disease
progression and bone mineral density in rheumatoid arthritis. J
Rheumatol. 30:1456–1463. 2003.PubMed/NCBI
|
|
10
|
Rossouw JE, Anderson GL, Prentice RL,
LaCroix AZ, Kooperberg C, Stefanick ML, Jackson RD, Beresford SA,
Howard BV, Johnson KC, et al: Risks and benefits of estrogen plus
progestin in healthy postmenopausal women: Principal results From
the Women's Health Initiative randomized controlled trial. JAMA.
288:321–333. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anderson GL, Limacher M, Assaf AR,
Bassford T, Beresford SA, Black H, Bonds D, Brunner R, Brzyski R,
Caan B, et al: Effects of conjugated equine estrogen in
postmenopausal women with hysterectomy: The Women's Health
Initiative randomized controlled trial. JAMA. 291:1701–1712. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Daraghmeh DN, Hopkins AM, King C, Abuhelwa
AY, Wechalekar MD, Proudman SM, Sorich MJ and Wiese MD: Female
reproductive status and exogenous sex hormone use in rheumatoid
arthritis patients treated with tocilizumab and csDMARDs.
Rheumatology (Oxford). 62:583–595. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gomez-Puerta JA, Celis R, Hernandez MV,
Ruiz-Esquide V, Ramírez J, Haro I, Cañete JD and Sanmartí R:
Differences in synovial fluid cytokine levels but not in synovial
tissue cell infiltrate between anti-citrullinated peptide/protein
antibody-positive and -negative rheumatoid arthritis patients.
Arthritis Res Ther. 15:R1822013. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Van Den Broek M, Dirven L, Klarenbeek NB,
Molenaar TH, Han KH, Kerstens PJ, Huizinga TW, Dijkmans BA and
Allaart CF: The association of treatment response and joint damage
with ACPA-status in recent-onset RA: A subanalysis of the 8-year
follow-up of the BeSt study. Ann Rheum Dis. 71:245–248. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Berglin E, Padyukov L, Sundin U, Hallmans
G, Stenlund H, Van Venrooij WJ, Klareskog L and Dahlqvist SR: A
combination of autoantibodies to cyclic citrullinated peptide (CCP)
and HLA-DRB1 locus antigens is strongly associated with future
onset of rheumatoid arthritis. Arthritis Res Ther. 6:R303–R308.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Johansson M, Arlestig L, Hallmans G and
Rantapää-Dahlqvist S: PTPN22 polymorphism and anti-cyclic
citrullinated peptide antibodies in combination strongly predicts
future onset of rheumatoid arthritis and has a specificity of 100%
for the disease. Arthritis Res Ther. 8:R192006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ge C and Holmdahl R: The structure,
specificity and function of anti-citrullinated protein antibodies.
Nat Rev Rheumatol. 15:503–508. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kissel T, Reijm S, Slot LM, Cavallari M,
Wortel CM, Vergroesen RD, Stoeken-Rijsbergen G, Kwekkeboom JC,
Kampstra A, Levarht E, et al: Antibodies and B cells recognising
citrullinated proteins display a broad cross-reactivity towards
other post-translational modifications. Ann Rheum Dis. 79:472–480.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shlomchik MJ, Craft JE and Mamula MJ: From
T to B and back again: Positive feedback in systemic autoimmune
disease. Nat Rev Immunol. 1:147–153. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan J, Harvey BP, Gee RJ, Shlomchik MJ and
Mamula MJ: B cells drive early T cell autoimmunity in vivo prior to
dendritic cell-mediated autoantigen presentation. J Immunol.
177:4481–4487. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Weyand CM and Goronzy JJ: Immunometabolism
in early and late stages of rheumatoid arthritis. Nat Rev
Rheumatol. 13:291–301. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weyand CM, Shen Y and Goronzy JJ:
Redox-sensitive signaling in inflammatory T cells and in autoimmune
disease. Free Radic Biol Med. 125:36–43. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu B, Goronzy JJ and Weyand CM: Metabolic
fitness of t cells in autoimmune disease. Immunometabolism.
2:e2000172020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ponchel F, Burska AN, Hunt L, Gul H, Rabin
T, Parmar R, Buch MH, Conaghan PG and Emery P: T-cell subset
abnormalities predict progression along the Inflammatory Arthritis
disease continuum: Implications for management. Sci Rep.
10:36692020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Paunovic V, Carroll HP, Vandenbroeck K and
Gadina M: Signalling, inflammation and arthritis: crossed signals:
The role of interleukin (IL)-12, −17, −23 and −27 in autoimmunity.
Rheumatology (Oxford). 47:771–776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim W, Min S, Cho M, Youn J, Min J, Lee S,
Park S, Cho C and Kim H: The role of IL-12 in inflammatory activity
of patients with rheumatoid arthritis (RA). Clin Exp Immunol.
119:175–181. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chu CQ, Swart D, Alcorn D, Tocker J and
Elkon KB: Interferon-gamma regulates susceptibility to
collagen-induced arthritis through suppression of interleukin-17.
Arthritis Rheum. 56:1145–1151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Geboes L, De Klerck B, Van Balen M,
Kelchtermans H, Mitera T, Boon L, De Wolf-Peeters C and Matthys P:
Freund's complete adjuvant induces arthritis in mice lacking a
functional interferon-gamma receptor by triggering tumor necrosis
factor alpha-driven osteoclastogenesis. Arthritis Rheum.
56:2595–2607. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sigidin YA, Loukina GV, Skurkovich B and
Skurkovich S: Randomized, double-blind trial of
anti-interferon-gamma antibodies in rheumatoid arthritis. Scand J
Rheumatol. 30:203–207. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ortmann RA and Shevach EM: Susceptibility
to collagen-induced arthritis: Cytokine-mediated regulation. Clin
Immunol. 98:109–118. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Haikal SM, Abdeltawab NF, Rashed LA, Abd
El-Galil TI, Elmalt HA and Amin MA: Combination Therapy of
Mesenchymal Stromal Cells and Interleukin-4 Attenuates Rheumatoid
Arthritis in a Collagen-Induced Murine Model. Cells. 8:8232019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Steinman L: A brief history of T(H)17, the
first major revision in the T(H)1/T(H)2 hypothesis of T
cell-mediated tissue damage. Nat Med. 13:139–145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Krebs CF and Steinmetz OM: CD4(+) T Cell
Fate in Glomerulonephritis: A Tale of Th1, Th17, and Novel Treg
Subtypes. Mediators Inflamm. 2016:53938942016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zielinski CE, Mele F, Aschenbrenner D,
Jarrossay D, Ronchi F, Gattorno M, Monticelli S, Lanzavecchia A and
Sallusto F: Pathogen-induced human TH17 cells produce IFN-ү or
IL-10 and are regulated by IL-1β. Nature. 484:514–518. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Evans HG, Roostalu U, Walter GJ, Gullick
NJ, Frederiksen KS, Roberts CA, Sumner J, Baeten DL, Gerwien JG,
Cope AP, et al: TNF-α blockade induces IL-10 expression in human
CD4+ T cells. Nat Commun. 5:31992014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Penatti A, Facciotti F, De Matteis R,
Larghi P, Paroni M, Murgo A, De Lucia O, Pagani M, Pierannunzii L,
Truzzi M, et al: Differences in serum and synovial CD4+ T cells and
cytokine profiles to stratify patients with inflammatory
osteoarthritis and rheumatoid arthritis. Arthritis Res Ther.
19:1032017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Van Hamburg JP, Asmawidjaja PS, Davelaar
N, Mus AM, Colin EM, Hazes JM, Dolhain RJ and Lubberts E: Th17
cells, but not Th1 cells, from patients with early rheumatoid
arthritis are potent inducers of matrix metalloproteinases and
proinflammatory cytokines upon synovial fibroblast interaction,
including autocrine interleukin-17A production. Arthritis Rheum.
63:73–83. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nakae S, Nambu A, Sudo K and Iwakura Y:
Suppression of immune induction of collagen-induced arthritis in
IL-17-deficient mice. J Immunol. 171:6173–6177. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lubberts E, Koenders MI, Oppers-Walgreen
B, van den Bersselaar L, Coenen-de Roo CJ, Joosten LA and van den
Berg WB: Treatment with a neutralizing anti-murine interleukin-17
antibody after the onset of collagen-induced arthritis reduces
joint inflammation, cartilage destruction, and bone erosion.
Arthritis Rheum. 50:650–659. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Corneth OB, Reijmers RM, Mus AM,
Asmawidjaja PS, van Hamburg JP, Papazian N, Siegers JY, Mourcin F,
Amin R, Tarte K, et al: Loss of IL-22 inhibits autoantibody
formation in collagen-induced arthritis in mice. Eur J Immunol.
46:1404–1414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kikuta J, Wada Y, Kowada T, Wang Z,
Sun-Wada GH, Nishiyama I, Mizukami S, Maiya N, Yasuda H, Kumanogoh
A, et al: Dynamic visualization of RANKL and Th17-mediated
osteoclast function. J Clin Invest. 123:866–873. 2013.PubMed/NCBI
|
|
42
|
Tlustochowicz W, Rahman P, Seriolo B,
Krammer G, Porter B, Widmer A and Richards HB: Efficacy and safety
of subcutaneous and intravenous loading dose regimens of
secukinumab in patients with active rheumatoid arthritis: Results
from a randomized phase II study. J Rheumatol. 43:495–503. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Corthay A: How do regulatory T cells work?
Scand J Immunol. 70:326–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rudensky AY: Regulatory T cells and Foxp3.
Immunol Rev. 241:260–268. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang R, Miao J, Zhang K, Zhang B, Luo X,
Sun H, Zheng Z and Zhu P: Th1-like treg cells are increased but
deficient in function in rheumatoid arthritis. Front Immunol.
13:8637532022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nie H, Zheng Y, Li R, Guo TB, He D, Fang
L, Liu X, Xiao L, Chen X, Wan B, et al: Phosphorylation of FOXP3
controls regulatory T cell function and is inhibited by TNF-α in
rheumatoid arthritis. Nat Med. 19:322–328. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Walter GJ, Fleskens V, Frederiksen KS,
Rajasekhar M, Menon B, Gerwien JG, Evans HG and Taams LS:
Phenotypic, functional, and gene expression profiling of peripheral
CD45RA+ and CD45RO+ CD4+CD25+CD127(low) Treg cells in patients with
chronic rheumatoid arthritis. Arthritis Rheumatol. 68:103–116.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kommoju V, Mariaselvam CM, Bulusu SN,
Ganapathy CK, Narasimhan PB, Thabah MM and Negi VS: Conventional
Tregs in treatment-naive rheumatoid arthritis are deficient in
suppressive function with an increase in percentage of CXCR3 and
CCR6 expressing Tregs. Immunol Res. 72:396–408. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Komatsu N, Okamoto K, Sawa S, Nakashima T,
Oh-hora M, Kodama T, Tanaka S, Bluestone JA and Takayanagi H:
Pathogenic conversion of Foxp3+ T cells into TH17 cells in
autoimmune arthritis. Nat Med. 20:62–68. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gonzalez-Rey E, Gonzalez MA, Varela N,
O'Valle F, Hernandez-Cortes P, Rico L, Büscher D and Delgado M:
Human adipose-derived mesenchymal stem cells reduce inflammatory
and T cell responses and induce regulatory T cells in vitro in
rheumatoid arthritis. Ann Rheum Dis. 69:241–248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pedrosa M, Gomes J, Laranjeira P, Duarte
C, Pedreiro S, Antunes B, Ribeiro T, Santos F, Martinho A, Fardilha
M, et al: Immunomodulatory effect of human bone marrow-derived
mesenchymal stromal/stem cells on peripheral blood T cells from
rheumatoid arthritis patients. J Tissue Eng Regen Med. 14:16–28.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ji LS, Sun XH, Zhang X, Zhou ZH, Yu Z, Zhu
XJ, Huang LY, Fang M, Gao YT, Li M and Gao YQ: Mechanism of
follicular helper T cell differentiation regulated by transcription
factors. J Immunol Res. 2020:18265872020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu R, Li X, Zhang Z, Zhou M, Sun Y, Su D,
Feng X, Gao X, Shi S, Chen W and Sun L: Allogeneic mesenchymal stem
cells inhibited T follicular helper cell generation in rheumatoid
arthritis. Sci Rep. 5:127772015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ma J, Zhu C, Ma B, Tian J, Baidoo SE, Mao
C, Wu W, Chen J, Tong J, Yang M, et al: Increased frequency of
circulating follicular helper T cells in patients with rheumatoid
arthritis. Clin Dev Immunol. 2012:8274802012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang J, Shan Y, Jiang Z, Feng J, Li C, Ma
L and Jiang Y: High frequencies of activated B cells and T
follicular helper cells are correlated with disease activity in
patients with new-onset rheumatoid arthritis. Clin Exp Immunol.
174:212–220. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kwok SK, Cho ML, Park MK, Oh HJ, Park JS,
Her YM, Lee SY, Youn J, Ju JH, Park KS, et al: Interleukin-21
promotes osteoclastogenesis in humans with rheumatoid arthritis and
in mice with collagen-induced arthritis. Arthritis Rheum.
64:740–751. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jang E, Cho SH, Park H, Paik DJ, Kim JM
and Youn J: A positive feedback loop of IL-21 signaling provoked by
homeostatic CD4+CD25-T cell expansion is essential for the
development of arthritis in autoimmune K/BxN mice. J Immunol.
182:4649–4656. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Olsen NJ, Watson MB, Henderson GS and
Kovacs WJ: Androgen deprivation induces phenotypic and functional
changes in the thymus of adult male mice. Endocrinology.
129:2471–2476. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Xu D, Wu Y, Gao C, Qin Y, Zhao X, Liang Z,
Wang Y, Feng M, Zhang C, Liu G and Luo J: Characteristics of and
reference ranges for peripheral blood lymphocytes and CD4+ T cell
subsets in healthy adults in Shanxi Province, North China. J Int
Med Res. 48:3000605209131492020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wilkinson NM, Chen HC, Lechner MG and Su
MA: Sex differences in immunity. Annu Rev Immunol. 40:75–94. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cutolo M, Sulli A and Straub RH: Estrogen
metabolism and autoimmunity. Autoimmun Rev. 11:A460–A464. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chakraborty B, Byemerwa J, Krebs T, Lim F,
Chang CY and McDonnell DP: Estrogen receptor signaling in the
immune system. Endocr Rev. 44:117–141. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Bynote KK, Hackenberg JM, Korach KS,
Lubahn DB, Lane PH and Gould KA: Estrogen receptor-alpha deficiency
attenuates autoimmune disease in (NZB × NZW)F1 mice. Genes Immun.
9:137–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Li J and Mcmurray RW: Effects of estrogen
receptor subtype-selective agonists on autoimmune disease in
lupus-prone NZB/NZW F1 mouse model. Clin Immunol. 123:219–226.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Svenson JL, Eudaly J, Ruiz P, Korach KS
and Gilkeson GS: Impact of estrogen receptor deficiency on disease
expression in the NZM2410 lupus prone mouse. Clin Immunol.
128:259–268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Maselli A, Conti F, Alessandri C,
Colasanti T, Barbati C, Vomero M, Ciarlo L, Patrizio M, Spinelli
FR, Ortona E, et al: Low expression of estrogen receptor beta in T
lymphocytes and high serum levels of anti-estrogen receptor alpha
antibodies impact disease activity in female patients with systemic
lupus erythematosus. Biol Sex Differ. 7:32016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Pierdominici M, Maselli A, Varano B,
Barbati C, Cesaro P, Spada C, Zullo A, Lorenzetti R, Rosati M,
Rainaldi G, et al: Linking estrogen receptor β expression with
inflammatory bowel disease activity. Oncotarget. 6:40443–40451.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kim DH, Park HJ, Park HS, Lee JU, Ko C,
Gye MC and Choi JM: Estrogen receptor α in T cells suppresses
follicular helper T cell responses and prevents autoimmunity. Exp
Mol Med. 51:1–9. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Guo D, Liu X, Zeng C, Cheng L, Song G, Hou
X, Zhu L and Zou K: Estrogen receptor β activation ameliorates
DSS-induced chronic colitis by inhibiting inflammation and
promoting Treg differentiation. Int Immunopharmacol. 77:1059712019.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Maret A, Coudert JD, Garidou L, Foucras G,
Gourdy P, Krust A, Dupont S, Chambon P, Druet P, Bayard F and Guéry
JC: Estradiol enhances primary antigen-specific CD4 T cell
responses and Th1 development in vivo. Essential role of estrogen
receptor alpha expression in hematopoietic cells. Eur J Immunol.
33:512–521. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Lambert KC, Curran EM, Judy BM, Milligan
GN, Lubahn DB and Estes DM: Estrogen receptor alpha (ERalpha)
deficiency in macrophages results in increased stimulation of CD4+
T cells while 17beta-estradiol acts through ERalpha to increase
IL-4 and GATA-3 expression in CD4+ T cells independent of antigen
presentation. J Immunol. 175:5716–5723. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Polanczyk MJ, Hopke C, Huan J, Vandenbark
AA and Offner H: Enhanced FoxP3 expression and Treg cell function
in pregnant and estrogen-treated mice. J Neuroimmunol. 170:85–92.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Tai P, Wang J, Jin H, Song X, Yan J, Kang
Y, Zhao L, An X, Du X, Chen X, et al: Induction of regulatory T
cells by physiological level estrogen. J Cell Physiol. 214:456–464.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Haghmorad D, Amini AA, Mahmoudi MB, Rastin
M, Hosseini M and Mahmoudi M: Pregnancy level of estrogen
attenuates experimental autoimmune encephalomyelitis in both
ovariectomized and pregnant C57BL/6 mice through expansion of Treg
and Th2 cells. J Neuroimmunol. 277:85–95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Pfeilschifter J, Koditz R, Pfohl M and
Schatz H: Changes in proinflammatory cytokine activity after
menopause. Endocr Rev. 23:90–119. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Sinatora RV, Chagas EFB, Mattera FOP,
Mellem LJ, Santos AROD, Pereira LP, Aranão ALC, Guiguer EL, Araújo
AC, Haber JFDS, et al: Relationship of inflammatory markers and
metabolic syndrome in postmenopausal women. Metabolites. 12:732022.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ralston SH, Russell RG and Gowen M:
Estrogen inhibits release of tumor necrosis factor from peripheral
blood mononuclear cells in postmenopausal women. J Bone Miner Res.
5:983–988. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lucas-Herald AK and Touyz RM: Androgens
and androgen receptors as determinants of vascular sex differences
across the lifespan. Can J Cardiol. 38:1854–1864. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Kwon H, Schafer JM, Song NJ, Kaneko S, Li
A, Xiao T, Ma A, Allen C, Das K, Zhou L, et al: Androgen conspires
with the CD8(+) T cell exhaustion program and contributes to sex
bias in cancer. Sci Immunol. 7:eabq26302022. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yang C, Jin J, Yang Y, Sun H, Wu L, Shen
M, Hong X, Li W, Lu L, Cao D, et al: Androgen receptor-mediated
CD8(+) T cell stemness programs drive sex differences in antitumor
immunity. Immunity. 55:1268–1283. e92022. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Berrih-Aknin S, Panse RL and Dragin N:
AIRE: A missing link to explain female susceptibility to autoimmune
diseases. Ann N Y Acad Sci. 1412:21–32. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhu ML, Bakhru P, Conley B, Nelson JS,
Free M, Martin A, Starmer J, Wilson EM and Su MA: Sex bias in CNS
autoimmune disease mediated by androgen control of autoimmune
regulator. Nat Commun. 7:113502016. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Dragin N, Bismuth J, Cizeron-Clairac G,
Biferi MG, Berthault C, Serraf A, Nottin R, Klatzmann D, Cumano A,
Barkats M, et al: Estrogen-mediated downregulation of AIRE
influences sexual dimorphism in autoimmune diseases. J Clin Invest.
126:1525–1537. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Afshan G, Afzal N and Qureshi S:
CD4+CD25(hi) regulatory T cells in healthy males and females
mediate gender difference in the prevalence of autoimmune diseases.
Clin Lab. 58:567–571. 2012.PubMed/NCBI
|
|
85
|
Gandhi VD, Cephus JY, Norlander AE,
Chowdhury NU, Zhang J, Ceneviva ZJ, Tannous E, Polosukhin VV, Putz
ND, Wickersham N, et al: Androgen receptor signaling promotes Treg
suppressive function during allergic airway inflammation. J Clin
Invest. 132:e1533972022. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Vargas-Villavicencio JA, Larralde C and
Morales-Montor J: Gonadectomy and progesterone treatment induce
protection in murine cysticercosis. Parasite Immunol. 28:667–674.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Xia T, Ma J and Sun Y and Sun Y: Androgen
receptor suppresses inflammatory response of airway epithelial
cells in allergic asthma through MAPK1 and MAPK14. Hum Exp Toxicol.
41:96032712211213202022. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Kalidhindi RSR, Ambhore NS, Balraj P,
Schmidt T, Khan MN and Sathish V: Androgen receptor activation
alleviates airway hyperresponsiveness, inflammation, and remodeling
in a murine model of asthma. Am J Physiol Lung Cell Mol Physiol.
320:L803–L818. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Kissick HT, Sanda MG, Dunn LK, Pellegrini
KL, On ST, Noel JK and Arredouani MS: Androgens alter T-cell
immunity by inhibiting T-helper 1 differentiation. Proc Natl Acad
Sci USA. 111:9887–9892. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Klein SL, Jedlicka A and Pekosz A: The Xs
and Y of immune responses to viral vaccines. Lancet Infect Dis.
10:338–349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Hong CY, Park JH, Ahn RS, Im SY, Choi HS,
Soh J, Mellon SH and Lee K: Molecular mechanism of suppression of
testicular steroidogenesis by proinflammatory cytokine tumor
necrosis factor alpha. Mol Cell Biol. 24:2593–2604. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Druckmann R and Druckmann MA: Progesterone
and the immunology of pregnancy. J Steroid Biochem Mol Biol.
97:389–396. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Scarpin KM, Graham JD, Mote PA and Clarke
CL: Progesterone action in human tissues: Regulation by
progesterone receptor (PR) isoform expression, nuclear positioning
and coregulator expression. Nucl Recept Signal. 7:e0092009.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Quispe Calla NE, Ghonime MG, Cherpes TL
and Vicetti Miguel RD: Medroxyprogesterone acetate impairs human
dendritic cell activation and function. Hum Reprod. 30:1169–1177.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Mao G, Wang J, Kang Y, Tai P, Wen J, Zou
Q, Li G, Ouyang H, Xia G and Wang B: Progesterone increases
systemic and local uterine proportions of CD4+CD25+ Treg cells
during midterm pregnancy in mice. Endocrinology. 151:5477–5488.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Hierweger AM, Engler JB, Friese MA,
Reichardt HM, Lydon J, DeMayo F, Mittrücker HW and Arck PC:
Progesterone modulates the T-cell response via glucocorticoid
receptor-dependent pathways. Am J Reprod Immunol. 81:e130842019.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Robinson DP and Klein SL: Pregnancy and
pregnancy-associated hormones alter immune responses and disease
pathogenesis. Horm Behav. 62:263–271. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hughes GC, Martin D, Zhang K, Hudkins KL,
Alpers CE, Clark EA and Elkon KB: Decrease in glomerulonephritis
and Th1-associated autoantibody production after progesterone
treatment in NZB/NZW mice. Arthritis Rheum. 60:1775–1784. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Hughes GC and Choubey D: Modulation of
autoimmune rheumatic diseases by oestrogen and progesterone. Nat
Rev Rheumatol. 10:740–751. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Carreno PC, Sacedon R, Jimenez E, Vicente
A and Zapata AG: Prolactin affects both survival and
differentiation of T-cell progenitors. J Neuroimmunol. 160:135–145.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Legorreta-Haquet MV, Chavez-Rueda K,
Chavez-Sanchez L, Cervera-Castillo H, Zenteno-Galindo E,
Barile-Fabris L, Burgos-Vargas R, Álvarez-Hernández E and
Blanco-Favela F: Function of treg cells decreased in patients with
systemic lupus erythematosus due to the effect of prolactin.
Medicine (Baltimore). 95:e23842016. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Dimitrov S, Lange T, Fehm HL and Born J: A
regulatory role of prolactin, growth hormone, and corticosteroids
for human T-cell production of cytokines. Brain Behav Immun.
18:368–374. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Tsampalas M, Gridelet V, Berndt S, Foidart
JM, Geenen V and Perrier d'Hauterive S: Human chorionic
gonadotropin: A hormone with immunological and angiogenic
properties. J Reprod Immunol. 85:93–98. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Lentz LS, Stutz AJ, Meyer N, Schubert K,
Karkossa I, von Bergen M, Zenclussen AC and Schumacher A: Human
chorionic gonadotropin promotes murine Treg cells and restricts
pregnancy-harmful proinflammatory Th17 responses. Front Immunol.
13:9892472022. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Sha J, Liu F, Zhai J, Liu X, Zhang Q and
Zhang B: Alteration of Th17 and Foxp3(+) regulatory T cells in
patients with unexplained recurrent spontaneous abortion before and
after the therapy of hCG combined with immunoglobulin. Exp Ther
Med. 14:1114–1118. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Diao LH, Li GG, Zhu YC, Tu WW, Huang CY,
Lian RC, Chen X, Li YY, Zhang T, Huang Y and Zeng Y: Human
chorionic gonadotropin potentially affects pregnancy outcome in
women with recurrent implantation failure by regulating the homing
preference of regulatory T cells. Am J Reprod Immunol. 77:2017.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Schumacher A, Heinze K, Witte J, Poloski
E, Linzke N, Woidacki K and Zenclussen AC: Human chorionic
gonadotropin as a central regulator of pregnancy immune tolerance.
J Immunol. 190:2650–2658. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Iqbal J, Sun L, Kumar TR, Blair HC and
Zaidi M: Follicle-stimulating hormone stimulates TNF production
from immune cells to enhance osteoblast and osteoclast formation.
Proc Natl Acad Sci USA. 103:14925–14930. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Qian H, Jia J, Yang Y, Bian Z and Ji Y: A
follicle-stimulating hormone exacerbates the progression of
periapical inflammation through modulating the cytokine release in
periodontal tissue. Inflammation. 43:1572–1585. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Kvien TK, Uhlig T, Odegard S and Heiberg
MS: Epidemiological aspects of rheumatoid arthritis: The sex ratio.
Ann N Y Acad Sci. 1069:212–222. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Sokka T, Toloza S, Cutolo M, Kautiainen H,
Makinen H, Gogus F, Skakic V, Badsha H, Peets T, Baranauskaite A,
et al: Women, men, and rheumatoid arthritis: Analyses of disease
activity, disease characteristics, and treatments in the QUEST-RA
study. Arthritis Res Ther. 11:R72009.PubMed/NCBI
|
|
112
|
Castagnetta LA, Carruba G, Granata OM,
Stefano R, Miele M, Schmidt M, Cutolo M and Straub RH: Increased
estrogen formation and estrogen to androgen ratio in the synovial
fluid of patients with rheumatoid arthritis. J Rheumatol.
30:2597–2605. 2003.PubMed/NCBI
|
|
113
|
Nieminen P, Hamalainen W, Savinainen J,
Lehtonen M, Lehtiniemi S, Rinta-Paavola J, Lehenkari P, Kääriäinen
T, Joukainen A, Kröger H, et al: Metabolomics of synovial fluid and
infrapatellar fat pad in patients with osteoarthritis or rheumatoid
arthritis. Inflammation. 45:1101–1117. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Schmidt M, Weidler C, Naumann H, Anders S,
Schölmerich J and Straub RH: Androgen conversion in osteoarthritis
and rheumatoid arthritis synoviocytes-androstenedione and
testosterone inhibit estrogen formation and favor production of
more potent 5alpha-reduced androgens. Arthritis Res Ther.
7:R938–R948. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
115
|
Capellino S, Straub RH and Cutolo M:
Aromatase and regulation of the estrogen-to-androgen ratio in
synovial tissue inflammation: Common pathway in both sexes. Ann N Y
Acad Sci. 1317:24–31. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Weitoft T, Larsson A and Ronnblom L: Serum
levels of sex steroid hormones and matrix metalloproteinases after
intra-articular glucocorticoid treatment in female patients with
rheumatoid arthritis. Ann Rheum Dis. 67:422–424. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Capellino S, Montagna P, Villaggio B,
Soldano S, Straub RH and Cutolo M: Hydroxylated estrogen
metabolites influence the proliferation of cultured human
monocytes: possible role in synovial tissue hyperplasia. Clin Exp
Rheumatol. 26:903–909. 2008.PubMed/NCBI
|
|
118
|
Schmidt M, Hartung R, Capellino S, Cutolo
M, Pfeifer-Leeg A and Straub RH: Estrone/17beta-estradiol
conversion to, and tumor necrosis factor inhibition by, estrogen
metabolites in synovial cells of patients with rheumatoid arthritis
and patients with osteoarthritis. Arthritis Rheum. 60:2913–2922.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Cenci S, Toraldo G, Weitzmann MN, Roggia
C, Gao Y, Qian WP, Sierra O and Pacifici R: Estrogen deficiency
induces bone loss by increasing T cell proliferation and lifespan
through IFN-gamma-induced class II transactivator. Proc Natl Acad
Sci USA. 100:10405–10410. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Garcia-Perez MA, Noguera I, Hermenegildo
C, Martínez-Romero A, Tarín JJ and Cano A: Alterations in the
phenotype and function of immune cells in ovariectomy-induced
osteopenic mice. Hum Reprod. 21:880–887. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Yoneda T, Ishimaru N, Arakaki R, Kobayashi
M, Izawa T, Moriyama K and Hayashi Y: Estrogen deficiency
accelerates murine autoimmune arthritis associated with receptor
activator of nuclear factor-kappa B ligand-mediated
osteoclastogenesis. Endocrinology. 145:2384–2391. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Andersson A, Grahnemo L, Engdahl C,
Stubelius A, Lagerquist MK, Carlsten H and Islander U:
IL-17-producing үδT cells are regulated by estrogen during
development of experimental arthritis. Clin Immunol. 161:324–332.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Zai Z, Xu Y, Qian X, Li Z, Ou Z, Zhang T,
Wang L, Ling Y, Peng X, Zhang Y and Chen F: Estrogen antagonizes
ASIC1a-induced chondrocyte mitochondrial stress in rheumatoid
arthritis. J Transl Med. 20:5612022. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Ishizuka M, Hatori M, Suzuki T, Miki Y,
Darnel AD, Tazawa C, Sawai T, Uzuki M, Tanaka Y, Kokubun S and
Sasano H: Sex steroid receptors in rheumatoid arthritis. Clin Sci
(Lond). 106:293–300. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Engdahl C, Borjesson AE, Forsman HF,
Andersson A, Stubelius A, Krust A, Chambon P, Islander U, Ohlsson
C, Carlsten H and Lagerquist MK: The role of total and
cartilage-specific estrogen receptor alpha expression for the
ameliorating effect of estrogen treatment on arthritis. Arthritis
Res Ther. 16:R1502014. View
Article : Google Scholar : PubMed/NCBI
|
|
126
|
Andersson A, Stubelius A, Karlsson MN,
Engdahl C, Erlandsson M, Grahnemo L, Lagerquist MK and Islander U:
Estrogen regulates T helper 17 phenotype and localization in
experimental autoimmune arthritis. Arthritis Res Ther. 17:322015.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Xu X, Yang H, Bullock WA, Gallant MA,
Ohlsson C, Bellido TM and Main RP: Osteocyte estrogen receptor β
(Ot-ERβ) regulates bone turnover and skeletal adaptive response to
mechanical loading differently in male and female growing and adult
mice. J Bone Miner Res. 38:186–197. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Sims NA, Dupont S, Krust A,
Clement-Lacroix P, Minet D, Resche-Rigon M, Gaillard-Kelly M and
Baron R: Deletion of estrogen receptors reveals a regulatory role
for estrogen receptors-beta in bone remodeling in females but not
in males. Bone. 30:18–25. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Bove R: Autoimmune diseases and
reproductive aging. Clin Immunol. 149:251–264. 2015. View Article : Google Scholar
|
|
130
|
Cutolo M, Sulli A, Pizzorni C, Craviotto C
and Straub RH: Hypothalamic-pituitary-adrenocortical and gonadal
functions in rheumatoid arthritis. Ann N Y Acad Sci. 992:107–117.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Yang DD, Krasnova A, Nead KT, Choueiri TK,
Hu JC, Hoffman KE, Yu JB, Spratt DE, Feng FY, Trinh QD and Nguyen
PL: Androgen deprivation therapy and risk of rheumatoid arthritis
in patients with localized prostate cancer. Ann Oncol. 29:386–391.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Drevinskaite M, Dadoniene J, Miltiniene D,
Patasius A and Smailyte G: Association between androgen deprivation
therapy and the risk of inflammatory rheumatic diseases in men with
prostate cancer: Nationwide Cohort Study in Lithuania. J Clin Med.
11:20392007. View Article : Google Scholar
|
|
133
|
Cutolo M, Seriolo B, Villaggio B, Pizzorni
C, Craviotto C and Sulli A: Androgens and estrogens modulate the
immune and inflammatory responses in rheumatoid arthritis. Ann N Y
Acad Sci. 966:131–142. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Ganesan K, Selvam R, Abhirami R, Raju KV,
Manohar BM and Puvanakrishnan R: Gender differences and protective
effects of testosterone in collagen induced arthritis in rats.
Rheumatol Int. 28:345–353. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Ganesan K, Balachandran C, Manohar BM and
Puvanakrishnan R: Effects of testosterone, estrogen and
progesterone on TNF-α mediated cellular damage in rat arthritic
synovial fibroblasts. Rheumatol Int. 32:3181–3188. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Martin CS, Crastin A, Sagmeister MS,
Turner JD, MacDonald L, Kurowska-Stolarska M, Scheel-Toellner D,
Taylor AE, Gilligan LC, Storbeck K, et al: Inflammation dynamically
regulates steroid hormone metabolism and action within macrophages
in rheumatoid arthritis. J Autoimmun. 147:1032632024. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Weidler C, Struharova S, Schmidt M, Ugele
B, Schölmerich J and Straub RH: Tumor necrosis factor inhibits
conversion of dehydroepiandrosterone sulfate (DHEAS) to DHEA in
rheumatoid arthritis synovial cells: A prerequisite for local
androgen deficiency. Arthritis Rheum. 52:1721–1729. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Straub RH, Harle P, Yamana S, Matsuda T,
Takasugi K, Kishimoto T and Nishimoto N: Anti-interleukin-6
receptor antibody therapy favors adrenal androgen secretion in
patients with rheumatoid arthritis: A randomized, double-blind,
placebo-controlled study. Arthritis Rheum. 54:1778–1785. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Jurcovicova J, Svik K, Scsukova S,
Bauerova K, Rovensky J and Stancikova M: Methotrexate treatment
ameliorated testicular suppression and anorexia related leptin
reduction in rats with adjuvant arthritis. Rheumatol Int.
29:1187–1191. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Stark K, Straub RH, Rovensky J, Blažičková
S, Eiselt G and Schmidt M: CYB5A polymorphism increases androgens
and reduces risk of rheumatoid arthritis in women. Arthritis Res
Ther. 17:562015. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Park JS, Choi SY, Hwang SH, Kim SM, Choi
J, Jung KA, Kwon JY, Kong YY, Cho ML and Park SH: CR6-interacting
factor 1 controls autoimmune arthritis by regulation of signal
transducer and activator of transcription 3 pathway and T helper
type 17 cells. Immunology. 156:413–421. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Rontzsch A, Thoss K, Petrow PK, Henzgen S
and Bräuer R: Amelioration of murine antigen-induced arthritis by
dehydroepiandrosterone (DHEA). Inflamm Res. 53:189–198. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Sandoughi M, Kaykhaei MA, Langarizadeh E
and Dashipour A: Effects of dehydroepiandrosterone on quality of
life in premenopausal women with rheumatoid arthritis: A
preliminary randomized clinical trial. Int J Rheum Dis.
23:1692–1697. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Hellberg S, Raffetseder J, Rundquist O,
Magnusson R, Papapavlou G, Jenmalm MC, Ernerudh J and Gustafsson M:
Progesterone dampens immune responses in in vitro activated CD4(+)
T cells and affects genes associated with autoimmune diseases that
improve during pregnancy. Front Immunol. 12:6721682021. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Khalkhali-Ellis Z, Seftor EA, Nieva DR,
Handa RJ, Price RH Jr, Kirschmann DA, Baragi VM, Sharma RV, Bhalla
RC, Moore TL and Hendrix MJ: Estrogen and progesterone regulation
of human fibroblast-like synoviocyte function in vitro:
implications in rheumatoid arthritis. J Rheumatol. 27:1622–1631.
2000.PubMed/NCBI
|
|
146
|
Jahanbakhshi M, Babaloo Z,
Mortazavi-Jahromi SS, Shokri MM, Ahmadi H and Mirshafiey A:
Modification of sexual hormones in rheumatoid arthritis patients by
M2000 (β-D-mannuronic Acid) as a Novel NSAID with immunosuppressive
property. Endocr Metab Immune Disord Drug Targets. 18:530–536.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Tengstrand B, Carlstrom K and Hafstrom I:
Bioavailable testosterone in men with rheumatoid arthritis-high
frequency of hypogonadism. Rheumatology (Oxford). 41:285–289. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Tengstrand B, Carlstrom K and Hafstrom I:
Gonadal hormones in men with rheumatoid arthritis-from onset
through 2 years. J Rheumatol. 36:887–892. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Cevik R, Em S, Gur A, Nas K, Sarac AJ and
Colpan L: Sex and thyroid hormone status in women with rheumatoid
arthritis: Are there any effects of menopausal state and disease
activity on these hormones? Int J Clin Pract. 58:327–332. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Takeuchi T, Tanaka Y, Higashitani C, Iwai
M, Komatsu K, Akazawa R and Lademacher C: A phase 2a, randomized,
double-blind, placebo-controlled trial of the efficacy and safety
of the oral gonadotropin-releasing hormone antagonist, ASP1707, in
postmenopausal female patients with rheumatoid arthritis taking
methotrexate. Mod Rheumatol. 31:53–60. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Weitzmann MN and Pacifici R: Estrogen
deficiency and bone loss: An inflammatory tale. J Clin Invest.
116:1186–1194. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Belenska-Todorova L, Zhivkova R, Markova M
and Ivanovska N: Follicle stimulating hormone and estradiol alter
immune response in osteoarthritic mice in an opposite manner. Int J
Immunopathol Pharmacol. 35:205873842110161982021. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Zhang X, Qiao P, Guo Q, Liang Z, Pan J, Wu
F, Wang X and Zhang L: High follicle-stimulating hormone level
associated with risk of rheumatoid arthritis and disease activity.
Front Endocrinol (Lausanne). 13:8628492022. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Vieira Borba V and Shoenfeld Y: Prolactin,
autoimmunity, and motherhood: when should women avoid
breastfeeding? Clin Rheumatol. 38:1263–1270. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Ram S, Blumberg D, Newton P, Anderson NR
and Gama R: Raised serum prolactin in rheumatoid arthritis: genuine
or laboratory artefact? Rheumatology (Oxford). 43:1272–1274. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Zoli A, Lizzio MM, Ferlisi EM, Massafra V,
Mirone L, Barini A, Scuderi F, Bartolozzi F and Magaró M: ACTH,
cortisol and prolactin in active rheumatoid arthritis. Clin
Rheumatol. 21:289–293. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Han A, Kim JY, Kwak-Kim J and Lee SK:
Menopause is an inflection point of age-related immune changes in
women. J Reprod Immunol. 146:1033462021. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Humphreys JH, Verstappen SM, Hyrich KL,
Chipping JR, Marshall T and Symmons DP: The incidence of rheumatoid
arthritis in the UK: Comparisons using the 2010 ACR/EULAR
classification criteria and the 1987 ACR classification criteria.
Results from the Norfolk Arthritis Register. Ann Rheum Dis.
72:1315–1320. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Mollard E, Pedro S, Chakravarty E, Clowse
M, Schumacher R and Michaud K: The impact of menopause on
functional status in women with rheumatoid arthritis. Rheumatology
(Oxford). 57:798–802. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Beydoun HA, El-Amin R, McNeal M, Perry C
and Archer DF: Reproductive history and postmenopausal rheumatoid
arthritis among women 60 years or older: Third national health and
nutrition examination survey. Menopause. 20:930–935. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Salliot C, Nguyen Y, Gusto G, Gelot A,
Gambaretti J, Mariette X, Boutron-Ruault MC and Seror R: Female
hormonal exposures and risk of rheumatoid arthritis in the French
E3N-EPIC cohort study. Rheumatology (Oxford). 60:4790–4800. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Alpizar-Rodriguez D, Mueller RB, Moller B,
Dudler J, Ciurea A, Zufferey P, Kyburz D, Walker UA, von Mühlenen
I, Roux-Lombard P, et al: Female hormonal factors and the
development of anti-citrullinated protein antibodies in women at
risk of rheumatoid arthritis. Rheumatology (Oxford). 56:1579–1585.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Harlow SD, Gass M, Hall JE, Lobo R, Maki
P, Rebar RW, Sherman S, Sluss PM and de Villiers TJ; STRAW + 10
Collaborative Group, : Executive summary of the Stages of
Reproductive Aging Workshop + 10: Addressing the unfinished agenda
of staging reproductive aging. J Clin Endocrinol Metab.
97:1159–1168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Engdahl C, Bondt A, Harre U, Raufer J,
Pfeifle R, Camponeschi A, Wuhrer M, Seeling M, Mårtensson IL,
Nimmerjahn F, et al: Estrogen induces St6gal1 expression and
increases IgG sialylation in mice and patients with rheumatoid
arthritis: A potential explanation for the increased risk of
rheumatoid arthritis in postmenopausal women. Arthritis Res Ther.
20:842018. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Guthrie KA, Dugowson CE, Voigt LF,
Koepsell TD and Nelson JL: Does pregnancy provide vaccine-like
protection against rheumatoid arthritis? Arthritis Rheum.
62:1842–1848. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Straub RH: The complex role of estrogens
in inflammation. Endocr Rev. 28:521–574. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Hodis HN, Mack WJ, Henderson VW, Shoupe D,
Budoff MJ, Hwang-Levine J, Li Y, Feng M, Dustin L, Kono N, et al:
Vascular effects of early versus late postmenopausal treatment with
estradiol. N Engl J Med. 374:1221–1231. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Orellana C, Wedren S, Kallberg H,
Holmqvist M, Karlson EW, Alfredsson L and Bengtsson C; EIRA Study
Group, : Parity and the risk of developing rheumatoid arthritis:
Results from the Swedish Epidemiological Investigation of
Rheumatoid Arthritis study. Ann Rheum Dis. 73:752–755. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Bondt A, Hafkenscheid L, Falck D, Kuijper
TM, Rombouts Y, Hazes JMW, Wuhrer M and Dolhain RJEM: ACPA IgG
galactosylation associates with disease activity in pregnant
patients with rheumatoid arthritis. Ann Rheum Dis. 77:1130–1136.
2018.PubMed/NCBI
|
|
170
|
Forger F, Marcoli N, Gadola S, Möller B,
Villiger PM and Østensen M: Pregnancy induces numerical and
functional changes of CD4+CD25 high regulatory T cells in patients
with rheumatoid arthritis. Ann Rheum Dis. 67:984–990. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Munoz-Suano A, Kallikourdis M, Sarris M
and Betz AG: Regulatory T cells protect from autoimmune arthritis
during pregnancy. J Autoimmun. 38:J103–J108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Clowse ME, Chakravarty E, Costenbader KH,
Chambers C and Michaud K: Effects of infertility, pregnancy loss,
and patient concerns on family size of women with rheumatoid
arthritis and systemic lupus erythematosus. Arthritis Care Res
(Hoboken). 64:668–674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Littlejohn EA: Pregnancy and rheumatoid
arthritis. Best Pract Res Clin Obstet Gynaecol. 64:52–58. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Lopez-Corbeto M, Martinez-Mateu S, Pluma
A, Ferrer R, López-Lasanta M, De Agustín JJ, Barceló M, Julià A and
Marsal S: The ovarian reserve as measured by the anti-Mullerian
hormone is not diminished in patients with rheumatoid arthritis
compared to the healthy population. Clin Exp Rheumatol. 39:337–343.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Brouwer J, Laven JS, Hazes JM, Schipper I
and Dolhain RJ: Levels of serum anti-Mullerian hormone, a marker
for ovarian reserve, in women with rheumatoid arthritis. Arthritis
Care Res (Hoboken). 65:1534–1538. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Hampl JS and Papa DJ:
Breastfeeding-related onset, flare, and relapse of rheumatoid
arthritis. Nutr Rev. 59((8 Pt 1)): 264–268. 2001.PubMed/NCBI
|
|
177
|
Karlson EW, Mandl LA, Hankinson SE and
Grodstein F: Do breast-feeding and other reproductive factors
influence future risk of rheumatoid arthritis? Results from the
Nurses' Health Study. Arthritis Rheum. 50:3458–3467. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Adab P, Jiang C Q, Rankin E, Tsang YW, Lam
TH, Barlow J, Thomas GN, Zhang WS and Cheng KK: Breastfeeding
practice, oral contraceptive use and risk of rheumatoid arthritis
among Chinese women: The guangzhou biobank cohort study.
Rheumatology (Oxford). 53:860–866. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Chen H, Wang J, Zhou W, Yin H and Wang M:
Breastfeeding and risk of rheumatoid arthritis: A systematic review
and metaanalysis. J Rheumatol. 42:1563–1569. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Orellana C, Saevarsdottir S, Klareskog L,
Karlson EW, Alfredsson L and Bengtsson C: Postmenopausal hormone
therapy and the risk of rheumatoid arthritis: Results from the
Swedish EIRA population-based case-control study. Eur J Epidemiol.
30:449–457. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Doran MF, Crowson CS, O'fallon WM and
Gabriel SE: The effect of oral contraceptives and estrogen
replacement therapy on the risk of rheumatoid arthritis: A
population based study. J Rheumatol. 31:207–213. 2004.PubMed/NCBI
|
|
182
|
Pedersen M, Jacobsen S, Garred P, Madsen
HO, Klarlund M, Svejgaard A, Pedersen BV, Wohlfahrt J and Frisch M:
Strong combined gene-environment effects in anti-cyclic
citrullinated peptide-positive rheumatoid arthritis: A nationwide
case-control study in Denmark. Arthritis Rheum. 56:1446–1453. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Qi S, Xin R, Guo W and Liu Y:
Meta-analysis of oral contraceptives and rheumatoid arthritis risk
in women. Ther Clin Risk Manag. 10:915–923. 2014.PubMed/NCBI
|
|
184
|
Walitt B, Pettinger M, Weinstein A, Katz
J, Torner J, Wasko MC and Howard BV; Women's Health Initiative
Investigators, : Effects of postmenopausal hormone therapy on
rheumatoid arthritis: The women's health initiative randomized
controlled trials. Arthritis Rheum. 59:302–310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Yuk JS, Seo YS, Im YH and Kim JH:
Menopausal hormone therapy and risk of seropositive rheumatoid
arthritis: A nationwide cohort study in Korea. Semin Arthritis
Rheum. 63:1522802023. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
D'elia HF and Carlsten H: The impact of
hormone replacement therapy on humoral and cell-mediated immune
responses in vivo in post-menopausal women with rheumatoid
arthritis. Scand J Immunol. 68:661–667. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Cutolo M, Balleari E, Giusti M, Intra E
and Accardo S: Androgen replacement therapy in male patients with
rheumatoid arthritis. Arthritis Rheum. 34:1–5. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Booji A, Biewenga-Booji CM, Huber-Bruning
O, Cornelis C, Jacobs JW and Bijlsma JW: Androgens as adjuvant
treatment in postmenopausal female patients with rheumatoid
arthritis. Ann Rheum Dis. 55:811–815. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Hall GM, Larbre JP, Spector TD, Perry LA
and Da Silva JA: A randomized trial of testosterone therapy in
males with rheumatoid arthritis. Br J Rheumatol. 35:568–573. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Hall GM, Daniels M, Huskisson EC and
Spector TD: A randomised controlled trial of the effect of hormone
replacement therapy on disease activity in postmenopausal
rheumatoid arthritis. Ann Rheum Dis. 53:112–116. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Hall GM, Daniels M, Doyle DV and Spector
TD: Effect of hormone replacement therapy on bone mass in
rheumatoid arthritis patients treated with and without steroids.
Arthritis Rheum. 37:1499–505. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Forsblad-D'elia H and Carlsten H: Hormone
replacement therapy in postmenopausal women with rheumatoid
arthritis stabilises bone mineral density by digital x-ray
radiogrammetry in a randomised controlled trial. Ann Rheum Dis.
70:1167–1168. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Pullerits R, D'elia HF, Tarkowski A and
Carlsten H: The decrease of soluble RAGE levels in rheumatoid
arthritis patients following hormone replacement therapy is
associated with increased bone mineral density and diminished
bone/cartilage turnover: A randomized controlled trial.
Rheumatology (Oxford). 48:785–790. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Kass A, Hollan I, Fagerland MW, Gulseth
HC, Torjesen PA and Førre ØT: Rapid anti-inflammatory effects of
gonadotropin-releasing hormone antagonism in rheumatoid arthritis
patients with high gonadotropin levels in the AGRA Trial. PLoS One.
10:e01394392015. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Kass AS, Forre OT, Fagerland MW, Gulseth
HC, Torjesen PA and Hollan I: Short-term treatment with a
gonadotropin-releasing hormone antagonist, cetrorelix, in
rheumatoid arthritis (AGRA): A randomized, double-blind,
placebo-controlled study. Scand J Rheumatol. 43:22–27. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Brown MA and Su MA: An inconvenient
variable: Sex hormones and their impact on T cell responses. J
Immunol. 202:1927–1933. 2019. View Article : Google Scholar : PubMed/NCBI
|