Introduction
Hypertrophic scars (HS) are fibroproliferative
disorders resulting from abnormal wound healing processes,
characterized by excessive extracellular matrix (ECM) accumulation
and ongoing inflammation. These scars develop in 30–90% of
individuals, with higher incidences reported among adolescents and
pregnant women and a markedly high prevalence following deep dermal
injuries such as burns (1).
Particularly in patients with full thickness burns, the prevalence
of HS can be as high as 70%, highlighting a substantial clinical
challenge due to the significant functional and aesthetic
disruptions and the psychological distress these scars often cause
(2). The presence of HS
substantially affects the quality of life of patients, causing
physical symptoms such as severe itching, tenderness and pain, as
well as psychological effects such as sleep disturbances, anxiety
and depression (3). These symptoms
are known to markedly contribute to morbidity, particularly
following burn injuries, leading to disruptions in daily activities
and psychosocial impairments (4).
Despite the variety of treatments available for HS,
such as corticosteroid injections, laser therapy, pressure therapy,
silicone applications, botulinum toxin, surgical interventions and
radiation therapy, there remains a significant demand for more
specialized and efficacious therapeutic choices (5). TGF-β1 inhibitors, as one of the
emerging treatments, target a crucial pathological element of HS by
blocking the activity of TGF-β, a cytokine known to contribute to
fibrosis by stimulating fibroblast proliferation and collagen
production (6). This advance
toward targeted therapies emphasizes the necessity of continuing
research dedicated to improving the precision and effectiveness of
treatments for HS and to lessen the extensive effect these scars
have on individuals. Within this research frontier, non-coding RNAs
(ncRNAs) emerge as crucial regulators in the post-transcriptional
network, influencing a wide array of biological pathways. Although
ncRNAs hold substantial promise for HS therapy due to their
regulatory capabilities, the development of ncRNA-based treatments
is challenging. The complexity of the ncRNA-mediated regulatory
networks, coupled with an incomplete understanding of their
mechanistic roles in scar formation and remodeling, poses
significant scientific challenges. Practical issues related to the
stability, delivery and specificity of ncRNA-based interventions
further complicate their clinical translation (7).
Adipose-derived stem cells (ADSCs) have gathered
attention in regenerative medicine for their versatility and
therapeutic potential. Derived from human adipose tissue, these
mesenchymal stem cells offer multipotency, abundant sources and
paracrine effects conducive to regenerative applications ranging
from muscle repair to cardiac rejuvenation (8). ADSCs have also shown potential in
differentiating into chondrocyte and osteocyte analogues, hinting
at new possibilities for bone and cartilage restoration,
particularly when paired with scaffold technologies (9). Refining the efficacy of ADSCs for
clinical application is an area full of potential for innovation
(10). Exosomes derived from ADSCs
(ADSCs-Exos) have been identified as powerful agents in enhancing
wound healing. They promote regenerative processes, such as cell
proliferation and collagen synthesis, thus accelerating wound
closure and improving healing quality, which are crucial to both
clinical and cosmetic skin repair (11,12).
However, as promising as ADSCs-Exos are, challenges remain,
particularly in understanding their precise mechanistic roles in
healing and in optimizing their use with biomaterials for enhanced
therapy (13).
The present study used a rabbit ear model to conduct
an in-depth examination of the dynamics of ncRNAs during
hypertrophic scar formation. By analyzing the differential
expression of ncRNAs, their interactions with mRNAs and the
enrichment of specific pathways, the aim of the present study was
to uncover potential therapeutic targets for HS, with particular
attention to micro (mi)RNA 194 as a notable candidate.
Materials and methods
Establishment of rabbit ear
hypertrophic scar model
A total of six female New Zealand white rabbits,
aged 3–4 months and weighing 2.5–3.0 kg, were sourced from Shanghai
Xinova Medical Research Co., Ltd. The rabbits were acclimated for 1
week under controlled conditions: Temperature was maintained at
22±2°C, relative humidity was kept at 50–60%, and the rabbits were
housed under a 12-h light/dark cycle. No abnormalities in the
morphology of either ear and no breaks in the soft tissues of the
skin were observed. Anesthesia was administered intravenously using
sodium pentobarbital (30 mg/kg). Following aseptic preparation, six
full-thickness wounds were created on the ventral ear surface,
avoiding the perichondrium and major blood vessels. Following
surgery, the rabbits were administered antibiotics and analgesics.
Each rabbit was housed individually post-operatively. Rabbits were
randomly assigned to either the control group or the ADSCs-Exos
treatment group, with three rabbits in each group. The health and
behavior of the rabbits were monitored every other day throughout
the duration of the experiment.
Procaine penicillin (cat. no. 6130-64-9; Merck KGaA)
was administered subcutaneously at a dose of 60,000 IU/kg every 48
h for 7 days (14). Concurrently,
buprenorphine (cat. no. 52485-79-7; Merck KGaA) was administered
subcutaneously at a dose of 0.05 mg/kg every 8 h for 2 days
(15,16). The wound care involved the
application of either saline or ADSCs-Exos (100 µg/2 ml) over the
first 3 days from injury. The micrON hsa-miR-194 agomir (micrON
miR-194) or micrON NC agomir (micrON NC) was administrated
subcutaneously at a dose of 0.2 µM. Scar progression was evaluated
at specified intervals, with tissue collection for histology on
days 3, 7, 14 and 21.
Following the administration of anesthesia using
sodium pentobarbital at a dose of 30 mg/kg, potassium chloride was
rapidly administered intravenously at a concentration of 150 mg/kg
(17). The rabbits were
subsequently assessed for death based on the following criteria: i)
Absence of respiration, pulse and heartbeat for >5 min, verified
both by auscultation with a stethoscope and palpation of the
cardiac region of the thorax; ii) loss of corneal reflexes; iii)
pupil dilation; and iv) absence of neural reflexes. The presence of
these conditions confirmed the animal's death. Ethical approval was
obtained from the Ethics Committee of The First Affiliated Hospital
of Harbin Medical University (approval no.,
XNM-YX-20220728-01).
ADSCs-Exos were obtained from Shanghai Xinova
Medical Research Co., Ltd. The negative control, micrON agomir NC
(cat. no. miR4N0000001-4-5) and micrON hsa-miR-194 agomir (cat. no.
miR40000460-4-5) were purchased from Guangzhou RiboBio Co.,
Ltd.
Hematoxylin and eosin (H&E) and
Masson's trichrome staining
Histological analyses were conducted on scar tissue
samples from both groups. H&E staining was performed to observe
cellular structures, while Masson's trichrome staining provided
insights into collagen distribution. The experiments were performed
according to the established protocols (18).
Briefly, slides were placed in staining can and
deparaffinized by immersion in three sequential baths of absolute
xylene for 4 min each. This was followed by a series of ethanol
washes at concentrations of 100, 100, 95, 90 and 70%, with each
wash lasting 4 min. Subsequently, the slides were rinsed under
running tap water for 2 min. They were then stained with
hematoxylin (cat. no. C0105, Beyotime) for 2 min, followed by
another 2-min rinse under tap water. To decolorize, the slides were
briefly dipped into 1% acid alcohol three times and rinsed again
for 2 min. After decolorization, the slides were immersed in 2%
potassium acetate for 3 min and rinsed once more for 2 min. The
staining process was completed by submerging the slides in eosin
for 2 min, followed by a final 2-min wash under running tap water.
All of the aforementioned procedures were carried out at room
temperature. The slides were then air-dried for 24 h at 38°C. Prior
to observation, the slides were dipped in absolute xylene for 1 min
and permanently mounted with a cover slip using DPX mounting medium
(cat. no. MM1410; Shanghai Maokang Biotechnology Co., Ltd.).
Slides were deparaffinized in staining cans using
three 4-min immersions in absolute xylene, followed by a descending
ethanol series from 100–70% in 5% decrements, each for 4 min. They
were then submerged in 60°C Bouin's solution (cat. no. HT101128;
Merck KGaA) for 45 min and rinsed until clear under running tap
water. Nuclei were differentiated with 8 min in modified Weigert's
hematoxylin, then rinsed for 2 min. Cytoplasm and erythrocytes were
stained with acid fuchsin (cat. no. HT15; Merck KGaA) for 5 min,
rinsed for 2 min, treated with phosphomolybdic acid (cat. no.
51429-74-4; Merck KGaA) for 10 min, then stained with methyl blue
(cat. no. 28983-56-4; Merck KGaA) for 5 min and rinsed. The slides
were treated with 1% acetic acid for 1 min, dehydrated in an
alcohol series from 100–70%, each for 1 min, dipped in xylene for 1
min and mounted with a cover slip using DPX (Shanghai Maokang
Biotechnology Co., Ltd.). All of the aforementioned procedures were
carried out at room temperature.
Immunohistochemical (IHC)
staining
Initially, these 4-µm paraffin-embedded tissue
sections were incubated at 60°C for 20 min, followed by a
deparaffinization process. The antigen retrieval process entailed
immersing the slides in a Tris/EDTA buffer (pH 9.0), followed by
heating at 100°C for 40 min using a microwave oven. The slides were
allowed to cool at room temperature for 30 min before being washed
twice with PBS with Tween 20 (PBST) buffer (0.1% Tween 20 in PBS;
pH 7.4) for 5 min per wash. The sections were treated with a
hydrogen peroxide blocking reagent (cat. no. ab64218; Abcam) for 10
min, followed by two 5-min washes using PBST buffer. The sections
were further incubated at room temperature for 1 h in a blocking
buffer containing 1% DMSO and 1% bovine serum albumin (cat. no.
ab64009; Abcam) in PBS buffer. The TGF-β1 (cat. no. ab215715;
1:500; Abcam) or VEGFA (cat. no. ab52917; Abcam) antibody in
blocking buffer were used for primary staining. The sections were
incubated with the primary antibodies overnight at 4°C in a
humidified chamber. The sections underwent six PBST washes (5 min
each) and were then incubated with a secondary goat anti-rabbit HRP
antibody (cat. no. ab6112; 1:750; Abcam) in blocking buffer for 1 h
at room temperature, followed by three washes in PBST (5 min each).
Visualization of the staining was achieved by incubating the
sections with 3,3′-diaminobenzidine (cat. no. ab209101; Abcam) for
10 min, according to the manufacturer's instructions. Excess DAB
was removed by washing under running deionized water for 5 min.
Counterstaining was performed with hematoxylin for 5 min at room
temperature, followed by a rinse in distilled water and pat-drying
prior to mounting. A Nikon H600L (Nikon Corporation) microscope in
normal brightfield mode was used to obtain images. The images of
H&E staining, Masson's trichrome staining and IHC staining were
captured at a ×100 magnification using a Nikon H600L light
microscope (Nikon Corporation).
RNA library preparation and
sequencing
The RNA sequencing library from rabbit ear scar
tissue on post-operative day 21 was constructed and sequenced by
Wuhan HealthCare Biotechnology Co., Ltd. Briefly, total RNA was
extracted from samples with an achieved yield of 1 µg per sample.
Integrity and purity were confirmed using agarose gel
electrophoresis (28S:18S ≥1.5) and Nanodrop spectrophotometry (OD
260/280 ratio between 1.8 and 2.2), respectively. RNA concentration
was quantified using a Qubit 4 Fluorometer (Thermo Fisher
Scientific, Inc.), with minimum readings of 500 ng/µl.
For small RNA libraries (18–30 nt), adaptors were
added to both ends of small RNAs and cDNA was synthesized through
reverse transcription, followed by purification using QIAseq miRNA
NGS Beads (cat. no. 333923; Qiagen GmbH). For long ncRNAs (lncRNAs)
and circular RNAs (circRNAs), RNA was fragmented into ~200 bp
segments after rRNA removal. Strand-specific cDNA was synthesized,
end-repaired, A-tailed and purified. Single-stranded cDNA libraries
were prepared by digesting double-stranded cDNA with USER enzyme
(cat. no. 5508; New England BioLabs, Inc.) and enriched by PCR for
15 cycles.
Sequencing was performed on the NovaSeq6000
(Illumina, Inc.) using 2×150 bp paired-end reads (sequencing by
synthesis). Post-sequencing, libraries underwent quality control
and alignment for differential expression analysis.
Analysis of genetic variability
In the analysis of genetic variability, several key
steps were meticulously followed. Initially, read normalization was
conducted to ensure uniformity in the number of reads.
Subsequently, a statistical model was employed to calculate both
the P-value and the fold change for hypothesis testing. To enhance
the robustness of the results, a multiple testing correction method
was used to obtain the adjusted P-value (adjP). Differentially
expressed RNAs were then identified based on specific criteria: A
log2 (Fold Change) >1 and a adjP<0.05. This rigorous approach
allowed for the identification of miRNAs with statistically
significant differences in expression levels among the samples,
categorizing them as either upregulated or downregulated.
Corresponding P-values and fold changes were obtained and a volcano
plot was created to visualize these findings.
Functional enrichment analysis
The functional enrichment analysis of selected
differential RNA target genes was conducted using Gene Ontology
(GO) (https://geneontology.org/) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) (release 109.0)
(https://www.genome.jp/kegg/). The
analysis was conducted using the clusterProfiler R package (version
4.4.4) in R (version 4.2.1) (https://www.r-project.org/), which facilitated the
identification of the primary functions and biological processes
(BP) associated with these genes (19). This led to the identification of
enriched GO terms and pathways, indicating the significant roles
and impacts of differential miRNA target genes on various cellular
mechanisms. The top 30 influenced pathways were visualized for
detailed insights. GO terms and KEGG pathways were considered
statistically significant with P-values <0.05. The gene count
thresholds were set to ≥20 for GO terms and ≥4 for KEGG pathways
according to the established protocol.
Protein-protein interaction (PPI)
analysis
The construction of the PPI network was facilitated
using the STRING database (version 12.0, http://string-db.org/), while visualization of the
network was performed using Cytoscape (version 3.10.1; National
Resource for Network Biology) (20). Interactions among proteins encoded
by differentially expressed genes were identified based on
predefined confidence criteria, with the network's significant
modules being elucidated using analytical tools. This methodology
underscored the complex interactions and potential functional
pathways influenced by treatment.
Prediction of miRNA-mRNA
interaction
TargetScanHuman (Release 7.2, http://www.targetscan.org/vert_72/) (21). miRDB (Version 6.0, http://mirdb.org/) (22) and STARBASE (https://rnasysu.com/encori/) (23) were used to predict interactions
between miRNA and mRNA.
Cell culture and transfection
Human keloid fibroblasts (cat. no. CP-H235) were
purchased from Procell Life Science & Technology Co., Ltd.
CP-H235 cells were cultured in DMEM/F12 culture medium (cat. no.
D8437; Merck KGaA), which was supplemented with 10% FBS (cat. no.
12006C; Merck KGaA), 100 U/ml penicillin (cat. no. P3032; Merck
KGaA) and 100 µg/ml streptomycin (cat. no. S9137; Merck KGaA).
Cells were maintained at a temperature of 37°C in an environment
containing 5% CO2. The absence of mycoplasma
contamination was confirmed by the supplier.
Cell seeding was conducted in 6-well plates 24 h
prior to the commencement of the experiment, ensuring that the
cultivation proceeded until the cell density approached ~75% of
each well's capacity. In preparation for transfection, 2.75 µl of
the pre-diluted liposome reagent (Lipofectamine®; cat.
no. L3000001; Thermo Fisher Scientific, Inc.) was mixed with miRNAs
at a final concentration of 20 nM, including negative control (NC),
miR-194 mimic, inhibitor NC or inhibitor, alongside untreated cells
for comparison. This mixture was then allowed a 20-min incubation
period to ensure adequate preparation. Following incubation, the
mixture was gently introduced to the growth medium and evenly
distributed across the wells. The plates were subsequently
incubated at 37°C in a CO2-enriched environment for 4–6
h.
The oligonucleotide sequences for the miR-194 mimic,
its NC, miR-194 inhibitor and the miR-194 inhibitor NC were
produced by Guangzhou RiboBio Co., Ltd. The sequences are as
follows: miR-194 mimic, sense strand sequence:
5′-CCAGUGGAGCUGCUGUUACUUC-3′; antisense strand sequence:
5′-GAAGUAACAGCAGCUCCACUGG-3′. miR-194 mimic NC, sense strand
sequence: 5′-UUCUCCGAACGUGUCACGU-3′, antisense strand sequence:
5′-ACGUGACACGUUCGGAGAA-3′. miR-194 inhibitor, sequence:
5′-CAGAUAACAGCAGCCCCACUGG-3′, miR-194 inhibitor NC, sequence:
5′-CAGUACUUUUGUGUAGUACAA-3′. Subsequent experiments were conducted
48 h post-transfection.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
RNA was extracted from rabbit scar tissue on
post-operative day 21 using 1 ml TRIzol® reagent (cat.
no. 15596026; Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocols and homogenized with a
rotor-stator. For RT, 2 µg total RNA, components of the reaction
mix and water were combined to a total volume of 20 µl. The mixture
was incubated through specific temperature cycles to synthesize
cDNA, according to the manufacturer's instructions (QuantiTect Rev.
Transcription Kit, cat. no. 205311; Qiagen GmbH). The reaction mix
of qPCR included 50 ng template DNA/cDNA, 0.5 µl of each primer, 2
µl PCR buffer, 1 U of Taq DNA polymerase and 0.3 µl PowerTrack SYBR
Green Master Mix (cat. no. A46109; Thermo Fisher Scientific, Inc.),
adjusted to 20 µl with deionized water. Thermal cycling conditions
comprised an initial denaturation at 95°C for 5 min, followed by
35–40 cycles of denaturation at 95°C for 30 sec, primer-specific
annealing and extension at 72°C for 60 sec, concluding with a final
extension at 72°C for 5 min. qPCR was performed with Sybr Green on
an Exicycler 96 instrument (Bioneer Corporation). RT-qPCR was
conducted in triplicate to ensure reproducibility. The primers for
the genes of interest are listed in Table I. Gene expression was normalized to
human or rabbit β-actin to analyze the relative expression using
the previously described 2−ΔΔCq method (24).
 | Table I.Primers for quantitative PCR assays
and corresponding targeted genes. |
Table I.
Primers for quantitative PCR assays
and corresponding targeted genes.
| Gene | Accession
number | Forward primers
(5′-3′) | Reverse primers
(5′-3′) |
|---|
| Human TGF-β1 | XM_054321897.1 |
TACCTGAACCCGTGTTGCTCTC |
GTTGCTGAGGTATCGCCAGGAA |
| Rabbit TGF-β1 | XM_051836380.1 |
GCCTGCAGAGGCTCAAGTTA |
CAACCACTCTGCTGTGTTGC |
| Human β-actin | NM_001017992.4 |
CATGATAGGGCGTCCTCGAC |
TGAGGAGGATGGGATGCTCA |
| Rabbit β-actin | NM_001101683.1 |
TCTCGACGAAACCTAACGGC |
GTCACCTTCACCGTTCCAGT |
| Hsa-miR-194-3p | MIMAT0004671 |
AAGAATCCAGTGGGGCTGC |
GTCGTATCCAGTGCAGGGT |
| Human U6 | NR_004394.1 |
CTCGCTTCGGCAGCACA |
AACGCTTCACGAATTTGCGT |
| Rabbit U6 | XM_008267384.3 |
CGGACGACCAGTTGTGGTAA |
CAGGGTCTTCACATTCGCCT |
The stem-loop RT-PCR method was used as outlined by
the sRNAPrimerDB protocol from Huazhong Agricultural University
(http://www.srnaprimerdb.com/protocolC) (25). Initially, the stem-loop RT primer
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCAGATA-3′ was
specifically designed to hybridize with hsa-miR-194-3p,
facilitating its reverse transcription. The cDNA product was then
amplified in real-time assays using the forward primer
5′-AAGAATCCAGTGGGGCTGC-3′ and the reverse primer
5′-GTCGTATCCAGTGCAGGGT-3′. The reverse-transcription, cDNA
synthesis and qPCR assays were performed on an Exicycler 96
instrument (Bioneer Corporation) following the manufacturers'
instructions.
Western blotting
Whole-cell protein extracts were obtained using 100
µl NP-40 lysis buffer (comprising 150 mM sodium chloride, 1.0%
NP-40 and 50 mM Tris, pH 8.0) supplemented with
ProteoGuard™ protease inhibitor cocktail (cat. no.
635672; Takara Bio, Inc.) in accordance with the manufacturer's
instructions. Pierce™ BCA Protein Assay Kit (cat. no.
23225; Thermo Fisher Scientific, Inc.) was used for protein
quantification according to the manufacturer's instructions. A
total of 25 µg protein from each sample was then separated by
SDS-PAGE on 10% gels and was subsequently transferred to
nitrocellulose membranes (cat. no. 88018; Thermo Fisher Scientific,
Inc.) by electroblotting. The membranes were blocked for 1 h with a
1% bovine serum albumin solution (cat. no. J61089.AP; Thermo Fisher
Scientific, Inc.) at room temperature before being incubated
overnight at 4°C with the following primary antibodies: Anti-TGF-β1
(cat. no. ab215715; 1:1,000; Abcam), anti-VEGFA (cat. no. ab52917;
1:1,000; Abcam), anti-collagen I (cat. no. ab138492; 1:500; Abcam)
and anti-GAPDH (cat. no. ab8245; 1:2,000; Abcam). The membranes
were then washed with PBST three times (10 min each time), before
being incubated for 1 h at room temperature with the corresponding
HRP-conjugated secondary antibodies, goat anti-rabbit IgG H&L
(cat. no. ab6721; 1:2,500; Abcam) and goat anti-mouse IgG H&L
(cat. no. ab205719; 1:2,500; Abcam). Visualization of the proteins
was achieved using the Gel Doc EZ Gel Documentation System (Bio-Rad
Laboratories, Inc.). The densitometric analysis of the protein
bands was performed using ImageJ software (version 1.4; National
Institutes of Health). The expression levels of the target proteins
were normalized to GAPDH expression to ensure accurate quantitative
analysis.
Luciferase reporter assay
To generate the wild-type and mutant variants of the
target nucleotides, a segment spanning nucleotides 40262 to 40504
in Homo sapiens TGF-β1 was selected, including the predicted
sponging region of hsa-miR-194-3p and TGF-β1 3′UTR. Initially, PCR
was performed using the initial forward primer pairs. Mutagenic
primers for the 3′UTR incorporating ‘GGTGAG’ were designed using
PrimerX (https://www.bioinformatics.org/primerx/index.htm).
These primers were then used to generate mutants by employing the
Q5 Site-Directed Mutagenesis Kit (cat. no. E0554; New England
Biolabs, Inc.) and using the Exicycler 96 instrument (Bioneer
Corporation) for the PCR amplification. The mutants were created
following the manufacturer's detailed instructions. To connect the
multiple cloning site to both the wild-type and mutant target
sequences, the forward primer with an EcoRI site and the
reverse primer with an XhoI site were used. Next, the
amplified products were inserted into the pCMV-Green Renilla
Luc Vector (cat. no. 16153; Thermo Fisher Scientific, Inc.) for
cloning. The primers and corresponding products are summarized in
Table II. In the transfection
process, a mixture of 0.5 µg Renilla Luc-TGF-β1 3′ UTR [or
TGF-β1 mutant (MUT)], 20 nM miR-194 (or NC) and 2.5 µl of the
liposome reagent (cat. no. L3000001; Thermo Fisher Scientific,
Inc.) was diluted in 2 ml DMEM/F12 culture medium. The transfection
was then performed in accordance with the manufacturer's
instructions. The detection of luciferase activity was conducted 48
h after transfection.
| Table II.Primers and the corresponding
products in establishing the wild-type and mutant TGF-β1 3′UTR. |
Table II.
Primers and the corresponding
products in establishing the wild-type and mutant TGF-β1 3′UTR.
| Primer name | Sequence
(5′-3′) | Products |
|---|
| Initial forward
primer |
GTGGTTGCCAGCATAATCC | Used in the first
round PCR |
| Initial reverse
primer |
AGAGTCCTGAGGATATTCTAGA | Used in the first
round PCR |
| Wild-type forward
primer |
GAATTCGTGGTTGCCAGCATAATCC | Contains
EcoRI site for the second round PCR |
| Wild-type reverse
primer |
CTCGAGAGAGTCCTGAGGATATTCTAGA | Contains
XhoI site for the second round PCR |
| Mutant forward
primer |
GAATTCGTGGTTGCCAGCATAATCC | Contains
EcoRI site for the second round PCR |
| Mutant reverse
primer |
CTCGAGGTTGTTGGTGAGGTGGAGAGTCCTGAGGATATTCTAGA | Contains
XhoI site and mutation ‘GGTGAG’ |
For the purpose of initiating cell lysis, 5X passive
lysis buffer was diluted to a 1X concentration using distilled
water. This diluted solution was then dispensed into 96-well
plates, at 100 µl per well. The disrupted cell solution was
subsequently transferred into 1.5 ml centrifuge tubes and
centrifuged at 13,200 × g for 10 min at a temperature of 4°C. The
supernatant was decanted into a fresh tube, Next, Luciferase Assay
Reagent II (cat. no. E1910; Promega Corporation) and the cell
lysate were thoroughly equilibrated to room temperature before
mixing. A volume of 100 µl Luciferase Assay Reagent II was combined
with 20 µl prepared cell lysate in a new 96-well plate.
Subsequently, 100 µl Dual-Glo Stop & Glo Buffer (cat. no.
E314B-C; Promega Corporation) was added to the mixture, which was
gently mixed 2–3 times, with a 2-sec wait before analysis. This
procedure enabled the measurement of Renilla luciferase
activity, providing a quantifiable luminescence readout of reporter
gene expression. The reaction products of the Dual Luciferase Assay
were quantified by measuring the absorbances at 560 nm (A560) and
480 nm (A480) on a 25-344S BioTek Fluorescence Microplate Reader
(Lonza Group Ltd.).
Enzyme-linked immunosorbent assay
(ELISA)
TGF-β1 Human ELISA Kit (cat. no. BMS249-4;
Invitrogen; Thermo Fisher Scientific, Inc.), IL-1β Human ELISA Kit
(cat. no. BMS224-2; Invitrogen; Thermo Fisher Scientific, Inc.) and
TNF-α Human ELISA Kit (cat. no. BMS223-2HS; Invitrogen; Thermo
Fisher Scientific, Inc.) were used to measure the concentrations of
cytokines in both groups. Antigen dilution was performed according
to the manufacturer's instructions, followed by sealing the wells
with closure solution and incubating at 37°C for 40 min. Wells were
washed three times with the provided washing solution, each for 3
min. Diluted samples were dispensed into enzyme-coated wells, with
a minimum of two wells per sample, each receiving 100 µl of the
sample. Incubation was performed at 37°C for 60 min, followed by a
washing step identical to the first. Development involved adding
100 µl of MB-hydrogen peroxide urea solution to each well,
incubating for 5 min at 37°C in a light-protected environment and
stopping the reaction with 50 µl of termination solution.
Concentrations of IL-1β, TGF-β1 and TNF-α were determined by
comparing optical density values to a standard curve provided with
each kit. The results of ELISA were detected by measuring the
absorbances at 450 nm (A450) and 620 nm (A620) on a 25-344S BioTek
Fluorescence Microplate Reader (Lonza Group Ltd.).
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical evaluations were conducted using GraphPad Prism
software version 8.0 (Dotmatics). Experiments were conducted a
minimum of three times to ensure robustness and reproducibility of
the findings. For the analysis of differences between two distinct
groups, a two-tailed Student's t-test, either paired or unpaired
depending on the data structure, was employed. When comparing three
or more groups, a two-way ANOVA followed by Tukey's multiple
comparisons test was used to identify significant differences among
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Establishment of rabbit ear
hypertrophic scar model
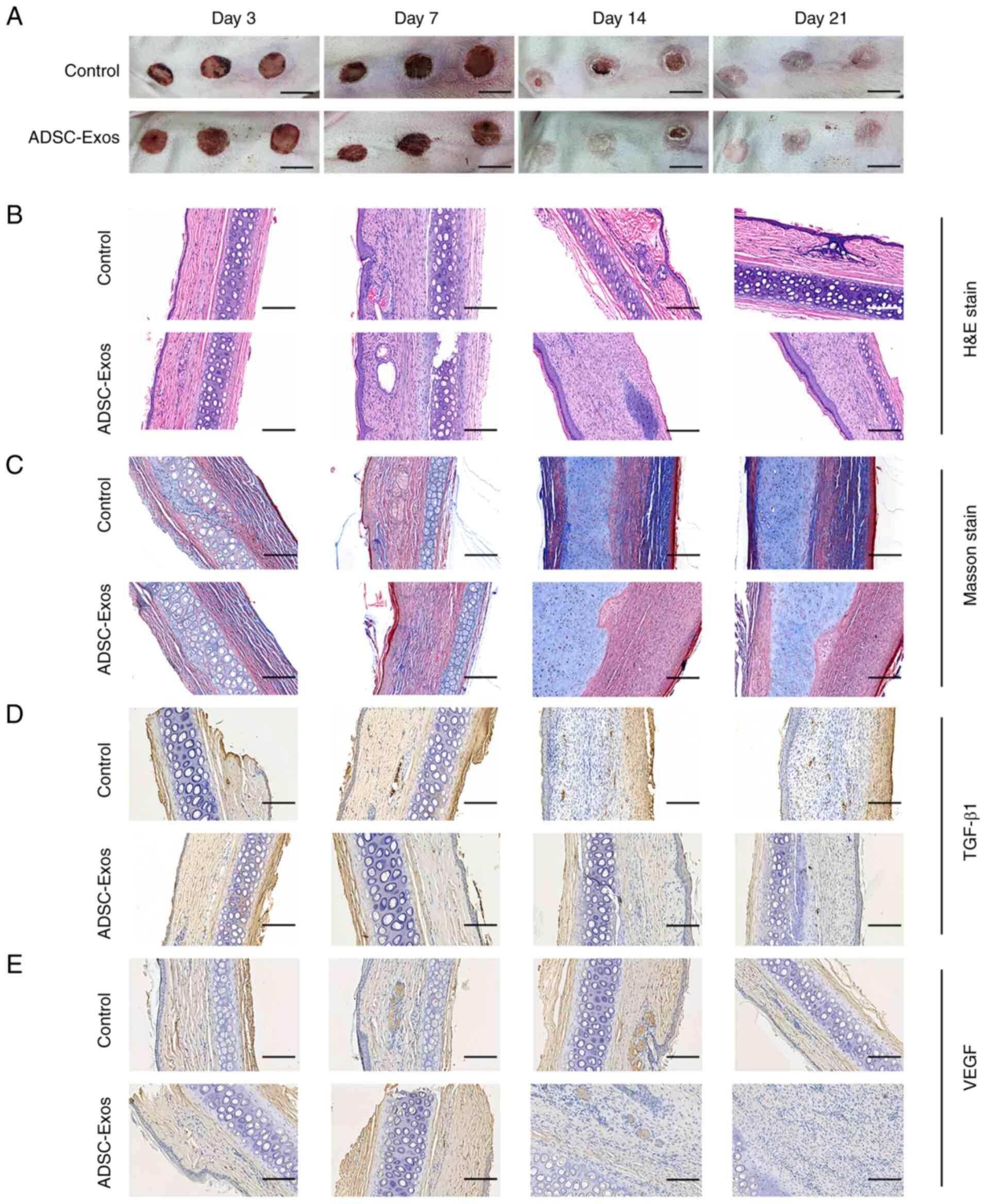
To assess the hypertrophic scar formation in rabbit
ears, gross examination and histological measurement were
performed. In the comparative analysis of hypertrophic scar
development over 21 days, the ADSCs-Exos-treated group demonstrated
a markedly moderated inflammatory response relative to the control
group. This was evidenced by a consistently lower degree of
inflammation, redness and swelling. The wound margins in the
ADSCs-Exos-treated group healed more quickly compared with the
control group, resulting in a faster decrease in scar size.
Furthermore, the coloration of scar tissue in the treatment group
transitioned rapidly from red to pink, ultimately achieving a
similar color to the surrounding skin, signifying an accelerated
reduction in blood supply and a more rapid progression through the
scar maturation phases (Fig.
1A).
H&E staining was conducted to elucidate the
cellular and structural changes within the scar tissue. Throughout
the experiment, the treatment group demonstrated a more regulated
arrangement of collagen fibers and less vascular proliferation,
indicating a neater and less aggressive healing process compared
with the control. The treatment appeared to modulate the wound
healing curve, leading to a more structured and less fibrotic scar
formation, as evidenced by the more orderly ECM and reduced
vascular proliferation seen in the H&E-stained sections
(Fig. 1B). Throughout the 21-day
study period, Masson's trichrome staining revealed a temporal
progression of tissue healing characterized by changes in the
inflammatory response and collagen deposition. Across all stages,
the treatment group consistently showed fewer and more regularly
arranged collagen fibers, suggesting a more controlled healing
process and a potential modification of the typical fibrotic
response, resulting in the formation of a less dense scar (Fig. 1C). IHC staining showed that TGF-β
expression levels in the control group remained high over the
21-day period. By contrast, the ADSCs-Exos-treated group exhibited
a marked reduction in TGF-β1 expression (Fig. 1D). In addition, scars in the
control group had consistently high levels of VEGF expression. By
contrast, the ADSCs-Exos treatment was associated with a
significant decrease in VEGF staining, suggesting a potential
regulatory effect on angiogenic signaling within the scar tissue
(Fig. 1E).
In brief, the evidence in Fig. 1 indicated that adipose-derived stem
cell exosomes significantly inhibited the formation of hypertrophic
scarring in rabbit ears.
lncRNA expression profiles and
regulatory networks in ADSCs-Exos-treated hypertrophic scar
formation
High-throughput sequencing of lncRNA from tissues
uncovered a broad spectrum of lncRNAs in both the
ADSCs-Exos-treated and control groups. The lncRNAs expression
profiles are summarized in Table
SI. These were effectively visualized in a volcano plot
(Fig. 2A), which segregated the
upregulated and downregulated lncRNAs, presenting a clear
disagreement in the response to treatment. This visualization was
complemented by a heatmap (Fig.
2B), which offered a detailed perspective of the expression
levels, accentuating the differences in lncRNA profiles between the
treatment and control groups. The advanced analysis (Fig. 2C and D) built on the previous
expression patterns, revealing that the complex regulatory networks
were controlled by these differentially expressed lncRNAs.
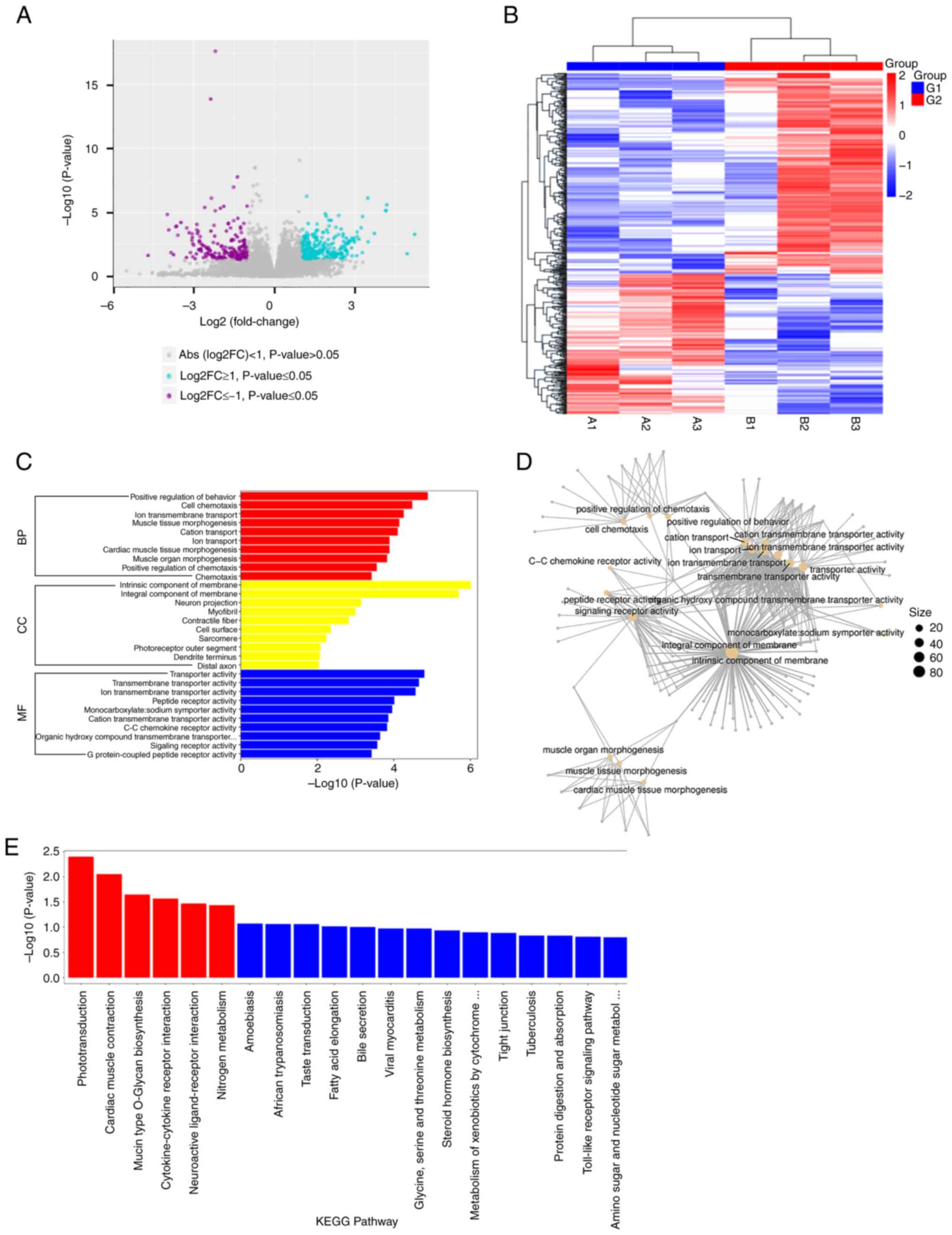 | Figure 2.Integrative analysis of lncRNA
expression profiles, regulatory networks and functional pathways in
ADSCs-Exos treated hypertrophic scar formation. (A) Volcano plot
representing the differential expression of lncRNAs between Control
Group (G1) and ADSCs-Exos treated Group (G2). Significantly
upregulated lncRNAs are indicated in cyan, downregulated lncRNAs in
purple and non-significant changes in grey. The x-axis represents
the log2 fold change and the y-axis shows the-log10 (P-value),
highlighting the magnitude and statistical significance of
expression differences. (B) Heatmap illustrating the expression
patterns of lncRNAs in samples from G1 and G2. A z-score normalized
color gradient indicates expression levels, with red for
upregulated and blue for downregulated lncRNAs. The hierarchical
clustering on the top dendrogram groups samples based on the
similarity of their lncRNA expression profiles. (C) Bar graph
showing the top 10 significantly enriched GO terms within the BP,
CC and MF categories that are associated with differentially
expressed lncRNAs. Only GO terms with a P-value ≤0.05 are included.
The horizontal axis indicates the statistical significance of the
enrichment, represented as-log10 P-value, while the vertical axis
lists the names of the significantly enriched GO terms. (D) Network
diagram of the top 20 enriched GO terms among genes associated with
differentially expressed lncRNAs, organized by the statistical
significance-log10 (P-value). The size of each node corresponds to
the count of associated candidate genes. This visualization
demonstrates the interconnectedness of significant BPs, and MFs,
with node size directly relating to the gene count for each GO
category. (E) Histogram of KEGG pathways significantly enriched in
connection with genes linked to differentially expressed lncRNAs.
The horizontal axis lists the names of the significantly enriched
pathways, while the vertical axis represents the-log10 (P-value),
indicating the level of enrichment significance. Pathways with a
P-value ≤0.05 are marked with red bars, signaling considerable
enrichment, while blue bars indicate pathways with lesser
significance. lncRNAs, long non-coding RNAs; ADSCs-Exos,
Adipose-derived stem cell exosomes; GO, Gene Ontology; BP,
biological process; CC, cellular component; MF, molecular function;
KEGG, Kyoto Encyclopedia of Genes and Genomes. |
GO functional analysis systematically categorized
the roles of genes influenced by the lncRNAs. A reference set of
genes was analyzed to determine statistically significant
enrichments, using Fisher's exact test for initial significance
testing. The resulting P-values were subsequently adjusted for
multiple comparisons, applying a controlled false discovery rate to
enhance the robustness of the findings. This analysis revealed
significant enrichments across BP, cellular components (CC) and
molecular functions (MF), highlighting the extensive networks and
pathways at play. To explore the broader implications of these
lncRNA-mediated changes, KEGG pathway analysis was conducted. This
annotated the modulated genes within known metabolic pathways,
linking them to established KEGG pathways and merged this with
differential expression data for a comprehensive understanding of
the metabolic pathways involved (Fig.
2E).
As well as the lncRNA expression profiles, the
differentially expressed circRNAs were examined (Table SII). The results were visualized
using both heatmap and volcano plot representations (Fig. S1). Although distinct expression
patterns were evident in the heatmap (Fig. S1A), the corresponding volcano plot
demonstrated that the observed differences in circRNA expression
between the treatment and control groups did not reach statistical
significance (Fig. S1B). These
findings indicated that, under the given experimental conditions,
the alterations in circRNA expression were not statistically
significant.
Collectively, these insights suggested that the
application of ADSCs-Exos exerted a modulatory effect on lncRNA
expression, which in turn may play a pivotal role during HS
formation. The identified GO terms and KEGG pathways provided a
deeper grasp of the BP and MF associated with the treatment, laying
the groundwork for uncovering potential therapeutic targets.
miRNA regulation in ADSCs-Exos-treated
scar formation
An integrative analysis was performed on the miRNA
expression profiles in HS treated with ADSCs-Exos (Table SIII). The volcano plot (Fig. 3A) outlined the significantly
modulated miRNAs, separating those upregulated from those
downregulated. Furthermore, the heat map (Fig. 3B) detailed the expression levels
across samples, exhibiting distinctive patterns. Functional
enrichment analysis was conducted using GO, highlighting the
involvement of the miRNAs in BP, CC and MF (Fig. 3C). Regulatory networks were also
visualized to display the complex interactions between miRNAs and
their target genes (Fig. 3D).
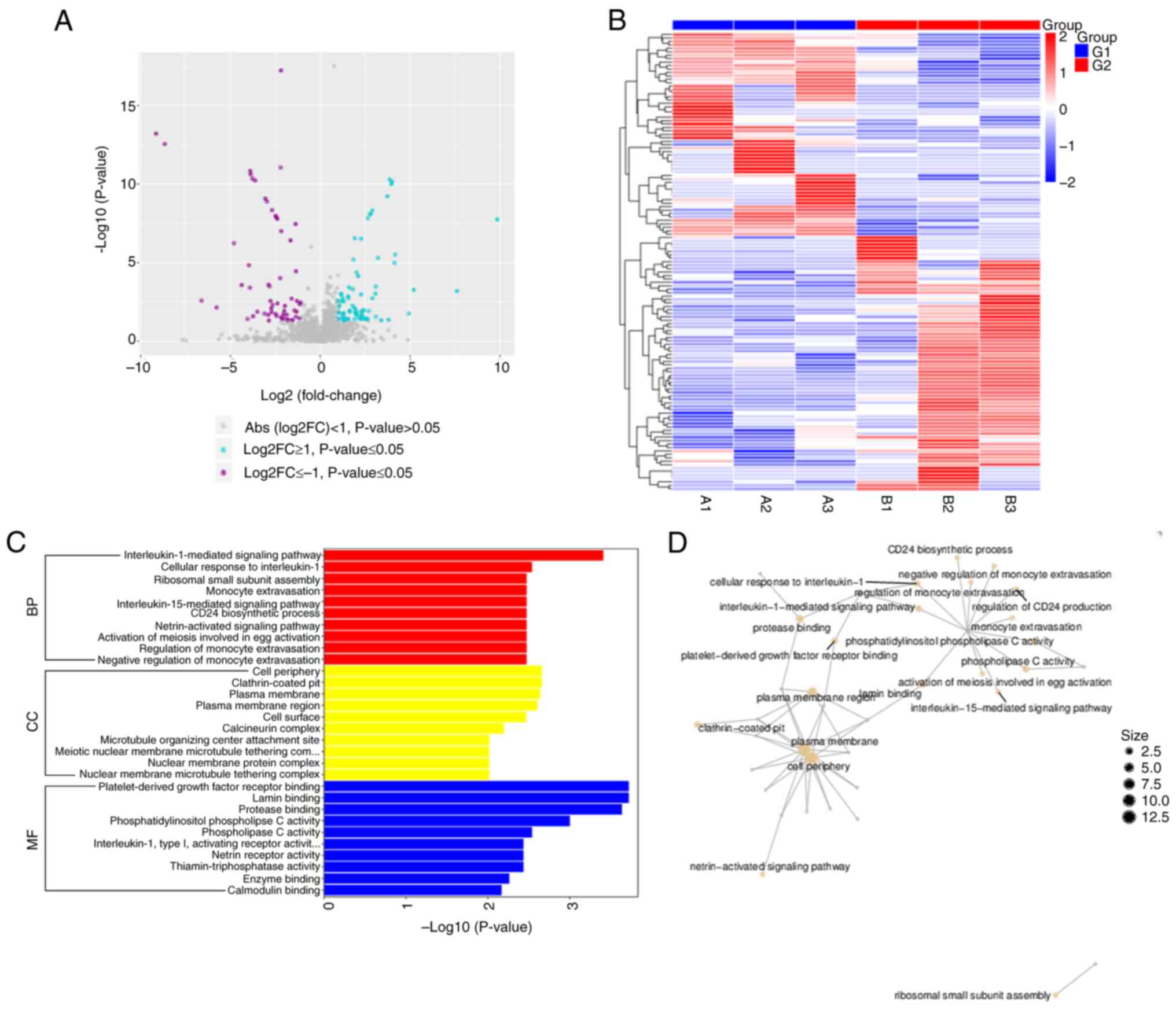 | Figure 3.Integrative analysis of miRNA
expression profiles, functional enrichment and regulatory networks
in ADSCs-Exos treated hypertrophic scar formation. (A) Volcano plot
visualizing the differential expression of miRNAs between the
control group (G1) and the ADSCs-Exos treated group (G2). Purple
points indicate miRNAs significantly downregulated in the
ADSCs-Exos treated group (G2) compared to the control (G1) and cyan
points represent miRNAs significantly upregulated in G2 relative to
G1. Grey points correspond to miRNAs with no significant difference
in expression between the two groups. (B) Heatmap detailing the
expression levels of miRNAs across individual samples in both
groups. Red indicates higher expression and blue denotes lower
expression levels. The hierarchical clustering on the y-axis groups
miRNAs with similar expression patterns, while the x-axis separates
the samples into control (G1) and treated (G2) groups. (C) GO
enrichment in BP, CC and MF associated with the differentially
expressed miRNAs. The length of each bar corresponds to the-log
(P-value), signifying the enrichment significance of the GO terms.
(D) Network diagram of the most significant GO terms connected with
the differentially expressed miRNAs. The size of the nodes reflects
the number of miRNA-related genes within each GO term and the
layout of the network illustrates the interconnectedness and
relevance of these terms to the ADSCs-Exos treatment effects.
miRNA. microRNA; ADSCs-Exos, Adipose-derived stem cell exosomes;
GO, Gene Ontology; BP, biological process; CC, cellular component;
MF, molecular function; KEGG, Kyoto Encyclopedia of Genes and
Genomes. |
Briefly, the results demonstrated significant
changes in miRNA expression profiles and related biological
pathways due to ADSCs-Exos treatment of hypertrophic scar
formation.
mRNA expression and pathway analysis
in ADSCs-Exos-treated HS
An extensive analysis was undertaken to understand
the modulation of mRNA expression in HS post-ADSCs-Exos
intervention. The mRNA expression profiles are summarized in
Table SIV. Statistical analysis
was performed, as evidenced by the volcano plot (Fig. 4A), which delineated differentially
expressed mRNAs with statistical rigor. mRNAs with a two-fold
increase and P<0.05 were identified, where cyan and purple
points represented statistically significantly upregulated and
downregulated genes, respectively. A heatmap (Fig. 4B) further illustrated these
differences, with gene expression variations between two groups,
the control and the treated group, evident through distinctive red
(upregulated) and blue (downregulated) color gradients. The GO
categorization and KEGG pathway analysis (Fig. 4C and D) revealed patterns and
themes within the data. Notably, ‘cardiac muscle tissue
morphogenesis’ and ‘sarcomere structures’ emerged as significantly
enriched terms within the BP and CC categories. The scatter plot
within the enrichment analysis visually underscored these themes,
with the size of the markers correlating to gene counts and hues
representing categorical significance. Contrasts between the
observed patterns and the expected outcomes highlighted the effect
of ADSCs-Exos treatment on the association between mRNA expression
and hypertrophic scar pathophysiology.
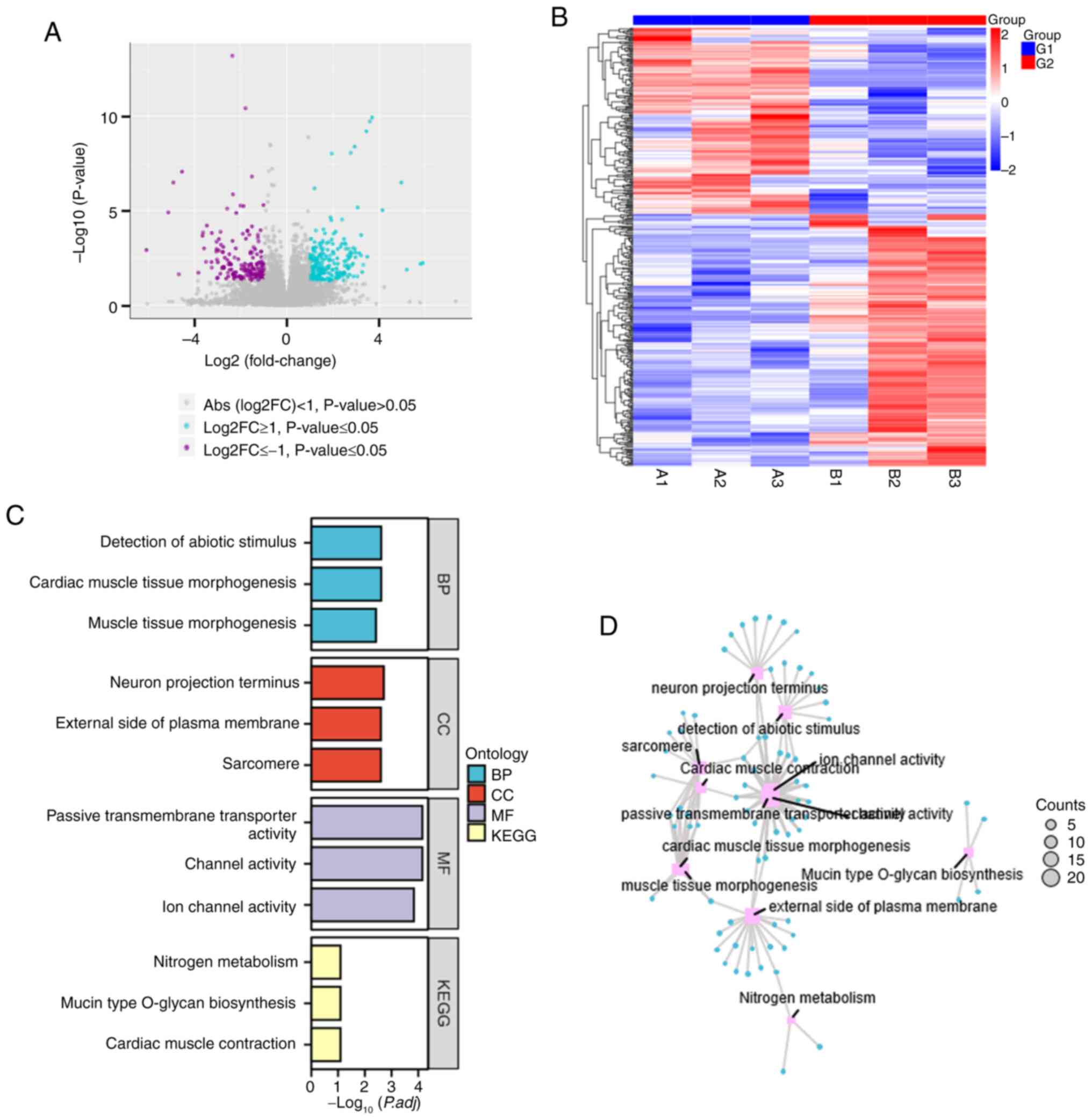 | Figure 4.Integrative analysis of mRNA
expression profiles, enrichment pathways and regulatory networks in
ADSCs-Exos treated hypertrophic scar formation. (A) Volcano plot
illustrating the differential expression of mRNAs in rabbit ear
scars, comparing the control group (G1) and the ADSCs-Exos treated
group (G2). Purple points denote mRNAs significantly downregulated
in G2, while cyan points represent mRNAs significantly upregulated
in G2, relative to G1. Grey points indicate mRNAs with no
significant expression changes. (B) Heatmap showing the mRNA
expression levels across samples, with red indicating upregulation
and blue signifying downregulation. The expression data are
hierarchically clustered to group similar expression patterns in G1
and G2. (C) Stacked bar chart of pathway enrichment based on
differentially expressed mRNAs. GO categories include BP, CC, MF
and KEGG pathways, with the significance of enrichment represented
by the-log10 (P.adj) value. (D) Network diagram of the top enriched
pathways associated with the differentially expressed mRNAs. The
size of each node reflects the number of mRNAs involved in that
particular pathway and the diagram highlights the most
significantly altered pathways in the ADSCs-Exos treated group.
ADSCs-Exos, Adipose-derived stem cell exosomes; GO, Gene Ontology;
BP, biological process; CC, cellular component; MF, molecular
function; KEGG, Kyoto Encyclopedia of Genes and Genomes. |
miR-194 orchestrates an extensive
regulatory network affecting gene expression
The analysis of the present study began by mapping
the potential targets of significantly changed miRNAs following
ADSCs-Exos treatment (Table
III). The Venn diagram (Fig.
5A) provided a visual summary of the extensive regulatory
capacity of these miRNAs. The genes found in the intersecting
region all bind to the three miRNAs, suggesting that they
collectively play a role in regulating scar formation. This subset
was determined through the intersection of predictions and
experimentally validated miRNA interactions, providing a strong
foundation for subsequent analyses. In addition, GO enrichment
analysis (Fig. 5B) was performed
on the intersected gene set, revealing significant BP, CC and KEGG
categories that the mRNAs candidates may influence. In the BP
category, ‘protein localization to cell’ was markedly enriched,
whereas ‘postsynaptic specialization’ was the primary CC affected.
The KEGG analysis indicated significant enrichment in ‘TGF-β
signaling pathway’. Furthermore, the PPI network diagram (Fig. 5C) showed the putative PPIs among
the targets shared by the miRNAs. The network described numerous
connections between the target genes, with nodes representing genes
and edges denoting the predicted regulatory associations.
Particularly, TGFB1, EGFR and BMPR2 emerged as focal nodes within
the network, each exhibiting a high degree of connectivity,
suggesting their significance as influential hubs within the PPI
landscape.
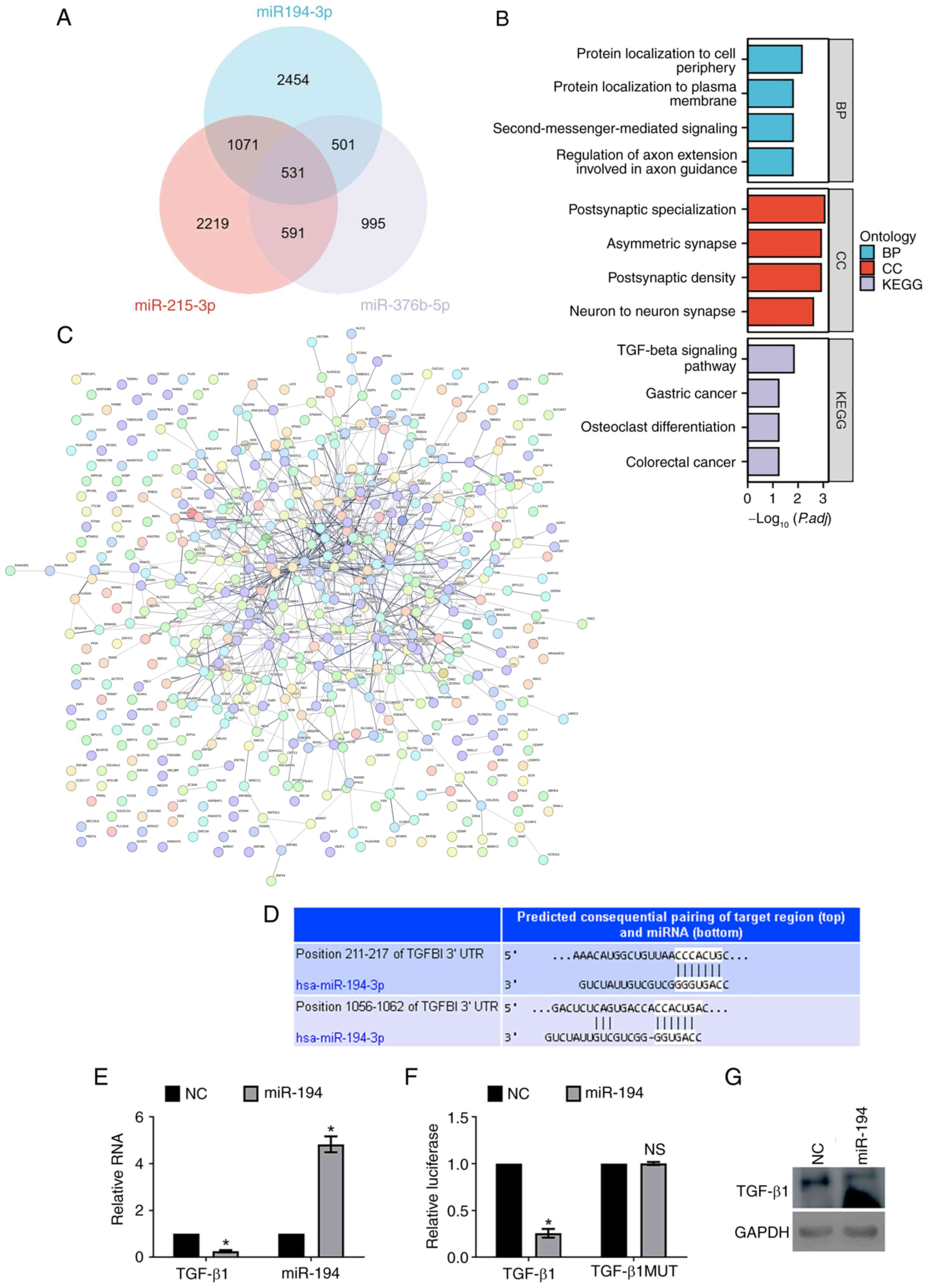 | Figure 5.miR-194-mediated regulatory networks
on gene expression. (A) Venn diagram depicting the shared target
genes of the indicated miRNAs, illustrating the extensive
regulatory reach of these miRNAs within the gene expression
landscape. The diagram highlights the number of unique and
overlapping target genes predicted by multiple databases. (B) Bar
graph of GO and KEGG pathway enrichment analysis among the
intersecting targets. The categories of BP, CC and significant KEGG
pathways are represented, with the significance of enrichment
indicated by-log10(P.adj) values. (C) Network of protein-protein
interactions among the intersecting targets, revealing potential
regulatory pathways that miRNAs may modulate. (D) Schematic
representation of the predicted miR-194 seed regions within the 3′
UTR of the TGF-β1 gene. This alignment suggests a direct regulatory
mechanism by which miR-194 may post-transcriptionally silence
TGF-β1 expression. (E) The expression of TGF-β1 and miR-194 in
CP-H235 cells were measured by quantitative PCR. (F) The relative
dual luciferase activity of TGF-β1 3′ UTR (TGF-β1) and TGF-β1 3′
UTR mutant (TGF-β1 MUT) was detected by luciferase reporter assay.
The relative luciferase activity of TGF-β1 is reduced in the
presence of miR-194, while that of TGF-β1 MUT alters little.
*P<0.05; NS vs. NC group. (G) Results of western blotting show
the TGF-β1 is reduced in the miR-194 group vs. NC group. miRNA/miR.
microRNA; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and
Genomes; BP, biological process; CC, cellular component; MUT,
mutant; NS, not significant; NC, negative control. |
| Table III.Differential expression of miRNAs
after ADSCs-Exos treatments. |
Table III.
Differential expression of miRNAs
after ADSCs-Exos treatments.
| ID |
FoldChange_Log2 | P-value |
|---|
| ocu-miR-144-3p | −1.309525013 |
6.407×10−4 |
| ocu-miR-144-5p | −1.347413066 |
1.976×10−4 |
|
ocu-miR-181c-3p | −1.201633861 |
1.003×10−4 |
| ocu-miR-206-3p | −5.247927513 |
6.750×10−4 |
| ocu-miR-215-3p | 1.378511623 |
2.821×10−4 |
| ocu-miR-215-5p | 3.790431619 |
5.560×10−4 |
|
ocu-miR-376b-5p | 1 |
2.713×10−4 |
| ocu-miR-451-5p | −1.470746932 |
4.324×10−4 |
| ocu-miR-486-5p | −1.168227713 |
3.874×10−4 |
| ocu-miR-194-3p | 7.48762561 |
8.040×10−4 |
According to the aforementioned results and the fact
that TGF-β-mediated signaling pathways dominate the proliferation
of wound healing, TGF-β1 was selected for further investigations.
Schematic evidence of the action of miR-194 on the TGF-β1 gene
(Fig. 5D) was provided. The
diagram displays the predicted binding of miR-194 to the seed
regions within the 3′ UTR of TGF-β1. This suggested a direct
post-transcriptional regulatory mechanism, where miR-194 can
silence TGF-β1 expression, a critical gene in fibrogenesis.
The Renilla Luc-TGF-β1 3′UTR or
Renilla Luc-TGF-β1 3′UTR MUT, along with NC or miR-194 mimic
were introduced into cells before the luciferase reporter assay.
The expression of miR-194 was validated by PCR assays (Fig. 5E). Finally, the results of the
luciferase reporter assay (Fig.
5F) confirmed the functional effect of miR-194 on TGF-β1
translation. When miR-194 was present, the luciferase activity
linked to the TGF-β1 3′ UTR was significantly reduced, which was
not observed with the MUT promoter. This reduction was consistent
with the findings of western blotting, which showed a decrease in
TGF-β1 protein levels upon the introduction of miR-194, validating
the repression effect of miR-194 on TGF-β1 expression (Fig. 5G).
In conclusion, miR-194 strongly influenced gene
regulation networks, with a specific suppression of TGF-β1,
suggesting its potential as a therapeutic target for diseases
characterized by aberrant fibrogenesis.
miR-194 modulates hypertrophic scar
formation by targeting TGF-β1
The efficacy of miR-194 intervention on hypertrophic
scar formation was evaluated by in vitro and in vivo
experiments. The transfection efficacy was confirmed by qPCR
assays, demonstrating that a significant increase in miR-194
expression was observed in the mimic group, whereas a decrease was
noted in the inhibitor group compared to their respective NCs. In
the untreated, NC and inhibitor NC groups, the miR-194 expression
remained relatively stable (Fig.
6A). Consequently, the miR-194 mimic and inhibitor were
employed in subsequent experiments. A significant reduction in
TGF-β1 RNA expression was demonstrated by the miR-194 mimics,
similar to that observed with ADSC-Exos treatment (Fig. 6B). Concurrently, the expression of
pro-inflammatory cytokines IL-1β, TNF-α and IL-10 was diminished,
as measured by ELISA (Fig. 6C). A
reduction in TGF-β1 protein levels was also observed (Fig. 6D). Conversely, an increase in
TGF-β1 expression and cytokine levels was observed with miR-194
inhibitor treatment (Fig.
6E-G).
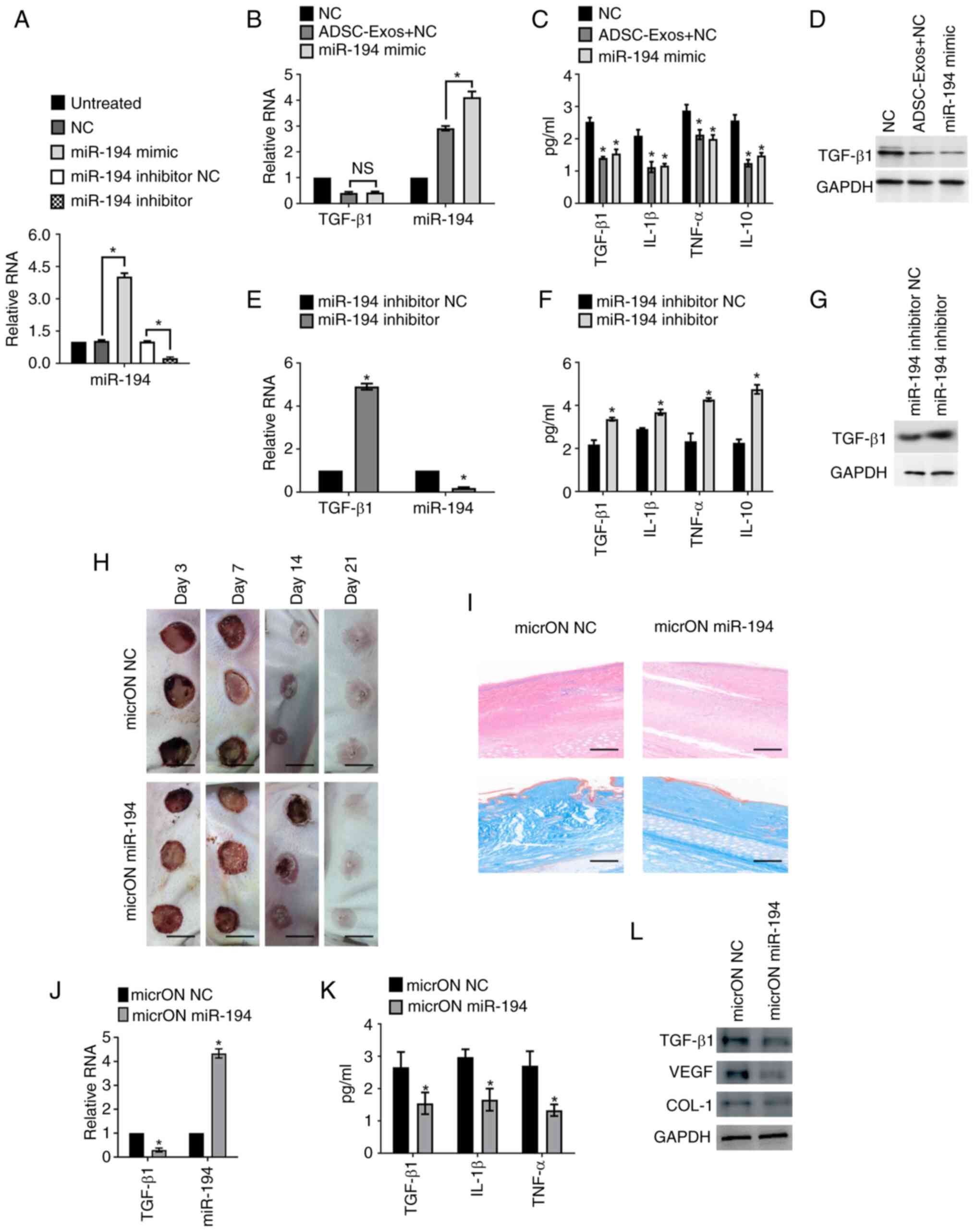 | Figure 6.miR-194 inhibits hypertrophic scar
formation via suppression of TGF-β1. (A) Expression of TGF-β1 and
miR-194 at the RNA level measured by quantitative PCR. *P<0.05
vs. NC group. (B) The relative RNA expression of TGF-β1 and miR-194
are determined by quantitative PCR. *P<0.05; NS vs. ADSC-Exos+NC
group. (C) Concentrations of specified cytokines measured by ELISA
analysis. *P<0.05; NS vs. NCADSC-Exos group. (D) The protein
levels of TGF-β1 detected by western blotting. (E) RNA levels of
TGF-β1 and miR-194 are determined by quantitative PCR. *P<0.05
vs. miR-194 inhibitor NC group. (F) Cytokine concentrations
detected by ELISA analysis. *P<0.05 vs. miR-194 inhibitor NC
group. (G) The protein levels of TGF-β1 detected by western
blotting. (H) Rabbit ear scars are observed grossly over a period
of 21 days, control miRON NC vs. miR-194 overexpressed tissues. A
reduction in scar erythema, flattening of scar borders and
diminution of scar size are noted with miR-194 overexpression,
indicating a mitigated hypertrophic response. (I) Top: Significant
differences between groups are shown by H&E staining.
Disorganized collagen fibers and abundant inflammatory cells are
displayed by the control miRON NC group. More organized collagen,
fewer inflammatory cells and improved cellular architecture result
from miRON miR-194 treatment. Bottom: Masson's trichrome staining
suggests that dense collagen, abundant fibroblasts and active
angiogenesis are exhibited by the control miRON NC group. Reduced
collagen density, fewer fibroblasts and decreased angiogenesis are
displayed by the miR-194 group, indicating attenuated scar
formation. (J) Expression of miRON miR-194 and TGF-β1 in scar
tissues is validated by quantitative PCR. *P<0.05 vs. miRON NC
group. A graph illustrating TGF-β1 and miR-194 RNA levels between
groups confirms the inhibition of miR-194 overexpression on TGF-β1
expression. (K) Cytokine concentrations in scar tissues are
measured by ELISA. (L) The expressions of TGF-β1, VEGF and COL-1
determined by western blotting, with GAPDH serving as a loading
control. The attenuation of key inflammatory factors involved in
scar formation by miR-194 overexpression is shown. *P<0.05 vs.
miRON NC group. Untreated, cells that have not received treatment;
NC, the negative control for the miR-194 mimic; miR-194 inhibitor
NC, the negative control for the miR-194 inhibitor; H&E. |
The effect of miR-194 on hypertrophic scar formation
was further investigated in vivo over a 21-day period.
Wounds treated with miR-194 mimics exhibited faster closure
compared with the control group, with notable diminutions in scar
erythema, scar borders and scar size being noted, indicating a
mitigated hypertrophic response (Fig.
6H). By H&E staining, a lowering in the scar elevation
index in the miR-194-treated group was revealed, suggesting
lessened scar tissue formation (Fig.
6I, top). Complementary Masson's trichrome staining supported
these findings, showing a significant cutback in collagen
deposition in the miR-194-treated samples (Fig. 6I, bottom). Quantitative assessments
of post-intervention gene expression confirmed a marked downturn in
TGF-β1 mRNA in the miR-194-treated group (Fig. 6J). This was accompanied by a
decrease in the levels of TGF-β1, IL-1β and TNF-α, as indicated by
ELISA (Fig. 6K). By western
blotting, a decrease in TGF-β1 protein levels and a decline in the
expression of fibrotic markers VEGF and type I collagen (COL-1) in
the miR-194 group were validated, confirming the inhibitory effects
on key inflammatory factors involved in scar formation (Fig. 6L).
Collectively, a significant inhibitory on
hypertrophic scar development was exerted by miR-194, primarily
through the suppression of TGF-β1.
Discussion
In the present study, rabbit ear models were used to
investigate the effects of adipose-derived stem cell exosomes
(ADSCs-Exos) on HS, a common and particularly challenging type of
proliferative scarring. HS is characterized by raised, often
discolored scar tissue that extends beyond the boundaries of the
original wound, significantly affecting the patient's self-image,
causing physical discomfort and diminishing overall quality of
life. Current HS management strategies, including surgical
excision, laser therapy and various pharmacological interventions
such as glucocorticoids, are associated with a number of drawbacks,
ranging from the risk of further scar formation following surgery
to inconsistent outcomes and the potential for adverse effects,
such as skin atrophy and hyperpigmentation. Such treatments, while
partly effective, do not provide a definitive solution and are
frequently associated with a high rate of scar recurrence (4). This highlights the pressing need for
novel and more effective treatments for HS, which was the
motivation for the present exploration of the therapeutic promise
of ADSCs-Exos in this challenging clinical area.
In exploring the intricate regulatory landscape of
ncRNAs within the context of proliferative scarring, the present
study delved into the roles of lncRNAs, miRNAs and circRNAs. These
ncRNAs, which do not encode proteins, are nonetheless pivotal in
regulating gene expression and consequently have a profound effect
on disease development, including scarring processes (26). LncRNAs such as H19 are implicated
in exacerbating scarring by promoting fibroblast proliferation and
collagen synthesis, highlighting their effect on scar development
through numerous signaling pathways (27). Concurrently, adipose-derived stem
cell exosomes (ADSCs-Exos) have shown promise in reducing scarring
by modulating these BP. They transport proteins, lipids and RNAs,
effectively attenuating inflammation, enhancing fibroblast activity
and accelerating wound healing and scar remodeling (28,29)
miRNAs (miRs), short single-stranded ncRNAs, are critical in
post-transcriptional gene regulation by targeting mRNAs for
degradation or translational repression. For example, miR-21 is
overexpressed in proliferative scarring, activating the PI3K/AKT
pathway by suppressing phosphatase and tensin homolog expression,
thus promoting fibroblast proliferation and collagen synthesis
(30). The sequencing data from
scar model tissues and post-treatment samples revealed an array of
differentially expressed miRNAs, implicating their regulatory
capacity across various BP and signaling pathways. Of note, the
expression of TGF-β1, a protein significantly implicated in scar
formation, was markedly influenced by ADSCs-Exos via ncRNA
mediation. Furthermore, miRNAs within ADSCs-Exos, such as miR-29,
target fibroblast activities crucial to scarring, affecting
collagen production and fibroblast differentiation (31). For instance, miR-124, reduces
inflammation by modulating macrophage activity and cytokine
release, which are crucial for mitigating the inflammatory response
integral to scar formation (32).
CircRNAs also contribute by functioning as miRNA ‘sponges’,
exemplified by circRNA CDR1as, which absorbs miR-7 to promote
collagen synthesis and fibroblast proliferation (33). However, the present findings from
rabbit ear scar model tissues showed that, while circRNAs were
abundantly expressed, they lacked statistical significance.
Overall, the present study underscored the complex interactions of
ncRNAs in ADSCs-Exos and their promising role in the innovative
treatment of proliferative scarring.
The present study revealed that ADSCs-Exos
significantly reduced proliferative scarring in rabbit ear models.
This was accompanied by a marked change in the expression of miRNAs
and mRNAs, particularly the post-treatment upregulation of miR-194,
which is implicated in regulating proteins including TGF-β1, a
protein downregulated following exosome treatment. The
overexpression of TGF-β1, crucial in proliferative scar formation,
enhances fibroblast proliferation and ECM deposition, thereby
exacerbating scarring (34). The
regulatory effect of miR-194 on TGF-β1 suggested the therapeutic
potential of adipose-derived stem cell exosomes in scarring, as
evidenced by predictive software and validated through luciferase
reporter assays showing the ability of miR-194 to downregulate
TGF-β1 activity. Previous studies have shown that several miRs,
including miR-124-3p, miR-26a, miR-29b, miR-145-5p and miR-485-5p,
contribute to the suppression of HS formation by downregulating
TGF-β1 (35–38). By contrast, miR-15a-5p enhances
hypertrophic scar development by boosting the TGF-β1 signaling
pathways (39). The verification
of miR-15a-5p and TGF-β1 sponge would be necessary to investigate
the hyperactivity of TGF-β1. The differences between the findings
of previous studies suggested that the mechanisms underlying
miRNA-regulated TGF-β1 on HS initiation and development need
further exploration. Subsequent experiments further confirmed the
role of miR-194 in the modulation of TGF-β1 expression, indicating
a promising therapeutic avenue in proliferative scarring management
through miR-194 and TGF-β1 regulation.
While TGF-β1 primarily promotes fibroblast
proliferation and collagen synthesis, VEGF-mediated angiogenesis is
beneficial for normal wound healing. The balance of VEGF-mediated
signaling pathways is critical for effective wound healing
(40). The present study provided
evidence that VEGF expression was decreased in the miR-194-treated
group. Similarly, Zhou et al (41) noted that miR-188-5p suppresses VEGF
expression, thereby restraining keloid formation. By contrast, Chen
et al (42) demonstrated
that ADSC treatments improved diabetic wound closure by activating
the VEGF-ERK pathway, accompanied by the suppression of
inflammatory factors. Qi et al (37) found that the inhibition of
miR-145-5p enhances TGF-β2 expression and VEGF secretion, reducing
HS development. Liu et al (31) showed that ADSCs-promote miR-21 and
miR-29b releases VEGF by activating the PI3K/AKT pathway,
facilitating wound healing. The discrepancies between studies may
be caused by various interactions, including the interplay of
lncRNA-miRNA-mRNA, or by different conditions, such as hypertrophic
scarring vs. keloid fibrosis. These controversial results suggest
that the global networks of miRNA-mediated VEGF expression require
further investigation.
In addition, the crosstalk between TGF-β1 and VEGF
cannot be ignored. Although specific research directly connecting
TGF-β1 and VEGF in hypertrophic scarring has not been addressed,
several studies have emphasized the potential for crosstalk between
these factors. Consistent with the current findings, Zhang et
al (43) demonstrate that VEGF
and collagen fibrosis are significantly increased in HS compared
with adjacent skin. Kinashi et al (44) illustrated that TGF-β1 regulates
VEGF expression in peritoneal fibrosis, promoting
lymphangiogenesis. Komi et al (45) highlight that mast cell-derived
VEGF, TGF-β and other mediators trigger scar formation, providing a
promising therapeutic target to improve wound healing. However, the
specific relationship between TGF-β1 and VEGF in HS has not been
fully elucidated. Further investigations into the interplay of
TGF-β1 and VEGF in hypertrophic scarring would help clarify the
mechanisms involved.
The development of proliferative scars is closely
associated with the activation and infiltration of inflammatory
factors, which are critical to the post-traumatic inflammatory
response, fibrosis and scar tissue remodeling. Key inflammatory
cytokines such as IL-1 and TNF-α play instrumental roles in these
processes. TNF-α, as an early mediator of inflammation, enhances
the recruitment and activation of immune cells, increases vascular
permeability and initiates the inflammatory response.
Simultaneously, IL-1 stimulates fibroblast proliferation and
collagen synthesis, directly linking inflammation to the fibrotic
changes observed in proliferative scars (46). The present study highlighted the
significant regulatory effect of miR-194 on these inflammatory
pathways. It was found that miR-194 inhibitors significantly
increased the levels of inflammatory cytokines, whereas miR-194
mimics decreased them. This suggested that miR-194 not only has the
ability to suppress TGF-β1 expression, which is known to drive
fibroblast proliferation and collagen synthesis, but also plays a
crucial role in modulating the inflammatory response itself.
In addition, the balance between collagen synthesis
and degradation, which is critical during normal wound healing, is
disrupted in proliferative scars. Here, there is a marked increase
in type I collagen synthesis coupled with reduced degradation,
leading to collagen accumulation within the scar tissue (47). This imbalance is largely driven by
the abnormal activation of fibroblasts into myofibroblasts,
potentiated by growth factors such as TGF-β and platelet-derived
growth factor. These myofibroblasts exhibit enhanced synthetic
capabilities, significantly contributing to the excessive
deposition of type I collagen seen in HS (45). TGF-β, in particular, plays a
pivotal role by promoting the increased production of type I
collagen, further exacerbating the fibrotic component of scarring
(48). The findings that miR-194
can modulate both TGF-β1 levels and the inflammatory milieu offer
promising therapeutic insights into managing the complex
pathophysiology of proliferative scarring.
ADSC-Exos have garnered considerable interest for
their ready availability, straightforward isolation techniques and
cost-effectiveness. In the present study, Exo-miR-194 showed
significant potential in modulating inflammatory responses,
maintaining angiogenic balance and reducing the overactivation of
TGF-β-mediated signaling pathways. These properties highlight its
promising role in the management of HS formation. A comparative
analysis of Exo-miR-194 with standard HS therapies could provide
deeper insights into its efficacy and utility, potentially leading
to more effective treatment strategies for HS. Further research is
warranted to delineate the specific mechanisms through which
Exo-miR-194 influences these pathways and to compare its effects
with those of established treatments.
While the present study demonstrated the potential
of exosomal miR-194 from ADSCs in attenuating hypertrophic scar
formation, it is important to acknowledge the limitations of the
phenotype verification. The current focus on histological and
molecular markers, although indicative, may not fully capture the
complexity of hypertrophic scar phenotypes. Despite observing
significant reductions in collagen density, fibroblast abundance
and angiogenesis in the miR-194 treated group, a more comprehensive
phenotypic analysis is required. Future directions include
implementing functional assays to evaluate mechanical properties of
scar tissue, such as tensile strength and elasticity, over 6–12
months. Extending the observation period will help assess long-term
effects on scar maturation and remodeling. Detailed molecular
pathway analyses using RNA sequencing and proteomics will explore
how miR-194 modulates scar formation. Employing additional animal
models with different wound healing profiles will enhance the
generalizability of the findings of the present study.
Investigating various delivery methods for ADSCs-Exos, such as
topical application, local injection, or systemic administration
and optimizing dosage and frequency will determine the most
effective strategies. These steps will validate the therapeutic
potential of miR-194 and ADSCs-Exos in treating hypertrophic scars
and facilitate clinical application.
In conclusion, the present findings suggested that
ADSCs-Exos, can significantly inhibit scar formation and reduce the
expression of pro-inflammatory and fibrotic markers such as TGF-β1,
particularly through the modulation of miR-194. This positions
ADSC-Exos as a promising therapeutic approach for managing
proliferative scarring, offering a novel avenue for intervention in
conditions characterized by excessive fibrosis and
inflammation.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available in the GEO database under accession numbers
GSE271671 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE271671)
and GSE271672 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE271672).
Authors' contributions
Conceptualization was performed by ZX and LH. Formal
analysis was conducted by ZX and YT. Validation was performed by
YT. ZX wrote the original draft and LH was responsible for writing,
reviewing and editing the manuscript. ZX and LH confirm the
authenticity of all the raw data. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The animals were treated according to protocols
approved by the Ethics Committee of The First Affiliated Hospital
of Harbin Medical University (approval no.,
XNM-YX-20220728-01).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ADSCs-Exos
|
exosomes from adipose-derived stem
cells
|
|
TGF-β1
|
transforming growth factor β1
|
|
VEGF
|
vascular endothelial growth factor
|
|
IL-1β
|
interleukin-1β
|
|
IL-10
|
interleukin-10
|
References
|
1
|
Barone N, Safran T, Vorstenbosch J,
Davison PG, Cugno S and Murphy AM: Current advances in hypertrophic
scar and keloid management. Semin Plast Surg. 35:145–152. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Finnerty CC, Jeschke MG, Branski LK,
Barret JP, Dziewulski P and Herndon DN: Hypertrophic scarring: The
greatest unmet challenge after burn injury. Lancet. 388:1427–1436.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Edwards J: Hypertrophic scar management.
Br J Nurs. 31:S24–S31. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Frech FS, Hernandez L, Urbonas R, Zaken
GA, Dreyfuss I and Nouri K: Hypertrophic scars and keloids:
Advances in treatment and review of established therapies. Am J
Clin Dermatol. 24:225–245. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gauglitz GG, Korting HC, Pavicic T,
Ruzicka T and Jeschke MG: Hypertrophic scarring and keloids:
Pathomechanisms and current and emerging treatment strategies. Mol
Med. 17:113–125. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schmieder SJ and Ferrer-Bruker SJ:
Hypertrophic scarring. StatPearls [Internet]. StatPearls
Publishing; Treasure Island, FL: 2024
|
|
7
|
Su L and Han J: Non-coding RNAs in
hypertrophic scars and keloids: Current research and clinical
relevance: A review. Int J Biol Macromol. 256:1283342024.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jia Q, Zhao H, Wang Y, Cen Y and Zhang Z:
Mechanisms and applications of adipose-derived stem
cell-extracellular vesicles in the inflammation of wound healing.
Front Immunol. 1:1214757 4. 2023.
|
|
9
|
Romano IR, D'Angeli F, Vicario N, Russo C,
Genovese C, Lo Furno D, Mannino G, Tamburino S, Parenti R and
Giuffrida R: Adipose-derived mesenchymal stromal cells: A tool for
bone and cartilage repair. Biomedicines. 11:17812023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lopez-Yus M, García-Sobreviela MP, Del
Moral-Bergos R and Arbones-Mainar JM: Gene therapy based on
mesenchymal stem cells derived from adipose tissue for the
treatment of obesity and its metabolic complications. Int J Mol
Sci. 24:74682023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee JH, Won YJ, Kim H, Choi M, Lee E,
Ryoou B, Lee SG and Cho BS: Adipose tissue-derived mesenchymal stem
cell-derived exosomes promote wound healing and tissue
regeneration. Int J Mol Sci. 24:104342023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Y, Younis MR, He G, Zheng Z, Wang Y,
Xue K, Sun J, Liu K, Huang P and Wang X: Oxidative
stimuli-responsive ‘pollen-like’ exosomes from silver nanoflowers
remodeling diabetic wound microenvironment for accelerating wound
healing. Adv Healthc Mater. 12:e23004562023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jing S, Li H and Xu H: Mesenchymal stem
cell derived exosomes therapy in diabetic wound repair. Int J
Nanomedicine. 18:2707–2720. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Edis A, Lumbis RH and Hedley J: Nursing
management of a rabbit undergoing a rhinostomy. Vet Nurse. 7:18–24.
2016. View Article : Google Scholar
|
|
15
|
Feldman ER, Singh B, Mishkin NG,
Lachenauer ER, Martin-Flores M and Daugherity EK: Effects of
cisapride, buprenorphine, and their combination on gastrointestinal
transit in New Zealand white rabbits. J Am Assoc Lab Anim Sci.
60:221–228. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Askar R, Fredriksson E, Manell E, Hedeland
M, Bondesson U, Bate S, Olsén L and Hedenqvist P: Bioavailability
of subcutaneous and intramuscular administrated buprenorphine in
New Zealand white rabbits. BMC Vet Res. 16:4362020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Leary S and Johnson C: AVMA guidelines for
the euthanasia of animals: 2020 edition*. Members of the Panel on
Euthanasia AVMA Staff Consultants; 2020
|
|
18
|
Suvik A and Effendy A: The use of modified
Masson's trichrome staining in collagen evaluation in wound healing
study. Mal J Vet Res. 3:39–47. 2012.
|
|
19
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:e050052015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen Y and Wang X: miRDB: An online
database for prediction of functional microRNA targets. Nucleic
Acids Res. 48(D1): D127–D131. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang JH, Li JH, Shao P, Zhou H, Chen YQ
and Qu LH: starBase: A database for exploring microRNA-mRNA
interaction maps from Argonaute CLIP-Seq and Degradome-Seq data.
Nucleic Acids Res. 39((Database Issue)): D202–D209. 2010.PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xie S, Zhu Q, Qu W, Xu Z, Liu X, Li X, Li
S, Ma W, Miao Y, Zhang L, et al: sRNAPrimerDB: Comprehensive primer
design and search web service for small non-coding RNAs.
Bioinformatics. 35:1566–1572. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu Z, Hou Q, Li M and Fu X: Molecular
mechanism of myofibroblast formation and strategies for clinical
drugs treatments in hypertrophic scars. J Cell Physiol.
235:4109–4119. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu B, Lin L, Yu S, Xia R and Zheng L:
Long non-coding RNA H19 acts as a microRNA-194 sponge to inhibit
the apoptosis and promote the proliferation of hypertrophic scar
fibroblasts. Can J Physiol Pharmacol. 99:1288–1297. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen SH, Chen ZY, Lin YH, Chen SH, Chou
PY, Kao HK and Lin FH: Extracellular vesicles of adipose-derived
stem cells promote the healing of traumatized achilles tendons. Int
J Mol Sci. 22:123732021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Y and Holmes C: Tissue regeneration
capacity of extracellular vesicles isolated from bone
marrow-derived and adipose-derived mesenchymal stromal/stem cells.
Front Cell Dev Biol. 9:6480982021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu Y, Li Y, Li N, Teng W, Wang M, Zhang Y
and Xiao Z: TGF-β1 promotes scar fibroblasts proliferation and
transdifferentiation via up-regulating MicroRNA-21. Sci Rep.
6:322312016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu SC, Bamodu OA, Kuo KT, Fong IH, Lin
CC, Yeh CT and Chen SG: Adipose-derived stem cell induced-tissue
repair or wound healing is mediated by the concomitant upregulation
of miR-21 and miR-29b expression and activation of the AKT
signaling pathway. Arch Biochem Biophys. 705:1088952021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang S and Pan S: miR-124-3p targeting of
TGF-β1 inhibits the proliferation of hypertrophic scar fibroblasts.
Adv Clin Exp Med. 30:263–271. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yao W, Li Y, Han L, Ji X, Pan H, Liu Y,
Yuan J, Yan W and Ni C: The CDR1as/miR-7/TGFBR2 axis modulates EMT
in silica-induced pulmonary fibrosis. Toxicol Sci. 166:465–478.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Firmansyah Y, Sidharta VM, Wijaya L and
Tan ST: Unraveling the significance of growth factors (TGF-β, PDGF,
KGF, FGF, Pro Collagen, VEGF) in the dynamic of wound healing.
Asian J Med Health. 22:49–61. 2024. View Article : Google Scholar
|
|
35
|
Qi J, Wu Y, Zhang H and Liu Y: LncRNA
NORAD regulates scar hypertrophy via miRNA-26a mediating the
regulation of TGFβR1/2. Adv Clin Exp Med. 30:395–403. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu J, Zhang L, Zhang S, Xian G, Zhao Y and
Bu X: MiR-29b inhibits hypertrophic scar tissue inflammation after
burn through regulating TGF-β1/Smad signaling pathway. Ital J
Dermatol Venerol. 156:251–252. 2021.PubMed/NCBI
|
|
37
|
Qi X, Liu Y and Yang M: Circ_0057452
functions as a ceRNA in hypertrophic scar fibroblast proliferation
and VEGF expression by regulating TGF-β2 expression and adsorbing
miR-145-5p. Am J Transl Res. 13:6200–6210. 2021.PubMed/NCBI
|
|
38
|
Xu S, Dong W and Shi Y: LncRNA PICSAR
binds to miR-485-5p and activates TGF-β1/Smad to promote abnormal
proliferation of hypertrophic scar fibroblasts (HSFs) and excessive
deposition of extracellular matrix (ECM). Med Mol Morphol.
54:337–345. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pang J, Zhu L, Lv Y, Xu L, Li N, Guo P and
Feng Q: miR-15a-5p up-regulates TLR4 and induces the formation of
hypertrophic scars and keloids. Cell Mol Biol (Noisy-le-grand).
69:158–163. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lian N and Li T: Growth factor pathways in
hypertrophic scars: Molecular pathogenesis and therapeutic
implications. Biomed Pharmacother. 84:42–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhou X, Lu J, Wu B and Guo Z: HOXA11-AS
facilitates the proliferation, cell cycle process and migration of
keloid fibroblasts through sponging miR-188-5p to regulate VEGFA. J
Dermatol Sci. 106:111–118. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen L, Zheng Q, Liu Y, Li L, Chen X and
Wang L and Wang L: Adipose-derived stem cells promote diabetic
wound healing via the recruitment and differentiation of
endothelial progenitor cells into endothelial cells mediated by the
VEGF-PLCγ-ERK pathway. Arch Biochem Biophys. 692:1085312020.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang YF, Li HZ, Wang XY, Ma HC, Wu Y,
Yuan XH and Chu YH: Morphology of hypertrophic scar tissues and
expressions of vascular endothelial growth factor and transforming
growth factor beta activated kinase 1 in these tissues. Zhongguo Yi
Xue Ke Xue Yuan Xue Bao. 37:446–450. 2015.(In Chinese). PubMed/NCBI
|
|
44
|
Kinashi H, Ito Y, Mizuno M, Suzuki Y,
Terabayashi T, Nagura F, Hattori R, Matsukawa Y, Mizuno T, Noda Y,
et al: TGF-β1 promotes lymphangiogenesis during peritoneal
fibrosis. J Am Soc Nephrol. 24:1627–1642. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Komi DEA, Khomtchouk K and Santa Maria PL:
A review of the contribution of mast cells in wound healing:
Involved molecular and cellular mechanisms. Clin Rev Allergy
Immunol. 58:298–312. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ohnuma K, Kasagi S, Uto K, Noguchi Y,
Nakamachi Y, Saegusa J and Kawano S: MicroRNA-124 inhibits TNF-α-
and IL-6-induced osteoclastogenesis. Rheumatol Int. 39:689–695.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ma Y, Liu Z, Miao L, Jiang X, Ruan H, Xuan
R and Xu S: Mechanisms underlying pathological scarring by
fibroblasts during wound healing. Int Wound J. 20:2190–2206. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Eremenko EE, Kwan PO, Ding J, Ghosh S and
Tredget EE: The effects of TGF-β1 and IFN-α2b on decorin, decorin
isoforms and type I collagen in hypertrophic scar dermal
fibroblasts. Wound Repair Regen. 32:135–145. 2024. View Article : Google Scholar : PubMed/NCBI
|