Aberrant epigenetic modifications serve pivotal
roles in the pathogenesis of a wide range of diseases. A key player
in this process is methylation, a biochemical modification
catalyzed by methyltransferases that add methyl groups to various
targets, including DNA, RNA, histones and other proteins. The
specificity and outcome of these methylation events are finely
regulated by a complex network of proteins known as
methyltransferases (writers), demethylases (erasers) and specific
binding proteins (readers) (1).
Recent multiomics studies have clarified that DNA and RNA
methylation should not be viewed as isolated layers of regulation;
integrated analyses have shown that compared with DNA methylation,
RNA methylation often explains a comparable or even larger fraction
of transcriptional variance (2,3). RNA
modifications, such as N6-methyladenosine (m6A),
5-methylcytosine (m5C) and 7-methylguanylate
(m7G), are crucial for the epigenetic regulation of gene
expression and cellular functions, and markedly impact the
development of numerous human diseases (4,5).
Advances in single-nucleotide resolution sequencing and
m5C antibody-based sequencing technologies have
reignited interest in the role of m5C modifications in
human diseases (6).
The present review aimed to provide an in-depth
overview of the distribution and regulation of m5C
modifications in both coding and ncRNAs and to explore their roles
in cancer progression. By examining how m5C modification
influences RNA metabolism and function, the present study aimed to
deepen the understanding of the critical role of RNA methylation in
epigenetic regulation and offer new insights into the regulatory
networks contributing to cancer.
RNA methyltransferases use S-adenosylmethionine
(SAM) as a methyl donor to transfer a methyl group to cytosine,
forming m5C, which occurs at position 5 of cytidine
residues in RNA (15,16). Key enzymes involved in this process
include members of the NOL1/NOP2/SUN domain (NSUN) family, the DNA
methyltransferase (DNMT) homolog DNMT2 and members of the
tRNA-specific methyltransferase family. NSUN proteins contain an
RNA recognition motif and a Rossmann fold catalytic core that
accommodates the SAM cofactor; NSUN proteins use two catalytic
cysteines in the active site, whereas DNMT2 uses a single active
site cysteine (17,18). A covalent intermediate is formed
between the protein cysteine and the cytosine in RNA, which
activates the electron-deficient pyrimidine ring, allowing the
nucleophilic attack of carbon 5 on the methyl group of SAM. NSUN
family proteins use the cysteine in motif VI to perform a
nucleophilic attack on carbon 6 of the target cytosine in RNA,
whereas DNMT family proteins use the cysteine in motif IV (19,20).
The NSUN family, which spans NSUN1 to NSUN7, serves extensive and
potentially overlapping roles in regulating early embryogenesis,
tumorigenesis and cell proliferation (21). DNMT2, traditionally known as a
cytosine DNMT, also functions as an RNA methyltransferase because
of its unique sequence and structure in the catalytic motif,
notably impacting cell proliferation and migration (22–24).
Additionally, the α-ketoglutarate-dependent
dioxygenase ABH1 (ALKBH1) functions as a demethylase and serves a
well-documented role in the mitochondria (32). ALKBH1 is involved in the biogenesis
of 5-hydroxymethyl-2′-O-methylcytidine (hm5Cm) and
5-formyl-2′-O-methylcytidine (f5Cm) at position 34 of
cytoplasmic tRNA-leucine (ct-tRNALeu) and f5C
in mitochondrial (mt)-tRNAMet, which are essential for
efficient translation (33).
ALKBH1 exhibits enzymatic activity against several substrates,
including m6A, N1-methyladenosine, N3-methylcytidine,
m5C and histone H2A (34); this substrate diversity and
specificity are key to its association with tumors.
Proteins that specifically bind to methylation
sites, such as RNA export factor binding protein 2 [Aly/REF export
factor (ALYREF)] and Y-box binding protein 1 (YBX1), are crucial
for determining the biological functions of RNA modifications
(35,36). ALYREF is responsible for exporting
mRNA from the nucleus, specifically binding to
m5C-modified mRNA to form mRNA ribonucleoprotein
complexes. YBX1 can recognize and bind to m5C-modified
mRNA through the Trp45 residue in its cold shock domain, thereby
maintaining its stability; however, a mutation in the indole ring
of Trp45 disrupts this interaction and abolishes its binding
capability (37–40). Another notable binding protein for
m5C is methyl-CpG binding protein 2 (MeCP2), which
interacts with the m5C-modified long non-coding RNA
(lncRNA) lnRncr3 to maintain the neural progenitor pool, thereby
limiting neuronal differentiation. Bioinformatic analyses also
suggest that MeCP2 may bind various cytosine-methylated RNAs in the
brain (41). Moreover, while the
YTH domain family protein 2 (YTHDF2) is a reader for RNA
m6A modification, studies have indicated that it can
also directly bind to m5C in RNA, although with a lower
affinity compared with m6A (42).
The detection of target genes with RNA methylation
modifications is essential for studying m5C
modifications. Current technologies and methods enable both
qualitative and quantitative analysis at the single-nucleotide
level, offering new approaches for research. The primary methods
for analyzing m5C methylation are discussed in the
present section.
MS uses nucleases to catalytically degrade RNA
sequences into nucleosides, which are then separated and identified
based on their mass-to-charge ratio; this approach enables the
detection of modified nucleosides. MS is widely used for detecting
various RNA modifications; however, it requires markedly purified
and concentrated target RNA and has relatively low sensitivity
(43). LC-MS/MS partially
addresses these limitations, as it can determine the overall
abundance of RNA m5C modifications in a sample; however,
its resolution does not reach the single-nucleotide level (44,45).
RNA-BisSeq is a widely applicable and cost-effective
method for the specific analysis of m5C RNA. Bisulfite
treatment converts unmethylated cytosines in single-stranded RNA to
uracil, whereas methylated cytosines remain unchanged. When
combined with high-throughput sequencing, this technique can
generate a transcriptome-wide m5C map with
single-nucleotide resolution. However, this method can be
influenced by factors such as RNA structure, experimental
temperature and other variables, which may lead to false-positive
results (16,46).
TAWO sequencing combines peroxotungstate oxidation
with TET enzyme oxidation, using peroxotungstate to oxidize
f5C to trihydroxylated thymine (thT), and thermostable
group II intron reverse transcriptase to convert thT to T during
cDNA synthesis. TAWO sequencing allows for the direct detection of
modified cytosines without affecting unmodified cytosines, thus
overcoming the false-positive issue associated with BisSeq
(47). However, because TAWO
sequencing relies on the conversion of m5C, the
conversion efficiency in mRNA samples requires further
optimization.
Methylated RNA immunoprecipitation sequencing
(MeRIP-seq) identifies markedly methylated RNA fragments by using
an RNA m5C antibody to bind m5C-modified RNA,
combined with high-throughput sequencing. This method can detect
low-abundance m5C modification sites and avoids
interference from other RNA modifications, although it is notably
dependent on the specificity of the m5C antibody
(48,49). Methylation individual-nucleotide
resolution crosslinking and immunoprecipitation (miCLIP) uses an
NSUN2 antibody instead of an m5C antibody to target the
NSUN2 m5C transcriptome landscape (50,51).
Moreover, 5-azacytidine-mediated RNA immunoprecipitation (5-azaIP)
utilizes 5-azacytidine, a cytidine analogue that forms a reversible
covalent bond with RNA m5C methyltransferases;
methyltransferase antibodies are then used to pull down the target
RNA. 5-azaIP reduces the non-specific RNA background and can
identify enzyme-specific methylation sites with single-nucleotide
resolution; however, 5-azacytidine is markedly toxic, and its
incorporation efficiency into RNA can influence the results
(52,53).
Nanopore direct RNA sequencing (DRS) enables direct
interrogation of RNA molecules without the need for cDNA
conversion, allowing for the capture of transcript isoforms and the
preservation of epitranscriptomic modifications (54,55).
As RNA passes through the nanopore, the bases cause characteristic
changes in the electrical current, which are then used to infer the
corresponding bases from the signal data (56). DRS provides a powerful platform for
parallel profiling of various RNA modifications, revealing the
complexity of the epitranscriptome (57). However, its implementation poses
challenges due to high costs and a reliance on machine
learning-based analysis, where methodological variability can limit
reproducibility (55); therefore,
the development of more robust and integrative analytical tools
remains a critical hurdle.
Experimental detection methods provide precise
information about RNA modifications; however, they are not without
limitations, including operational complexity and high costs. As a
result, several models have been developed to predict RNA
m5C modifications. These include PEA-m5C
(58), m5C-PseDNC
(59) and m5C-HPCR
(60). Additionally, some models,
such as m5C-Pred-SVM (61) and DeepMRMP (62), are designed to predict specific
m5C sites. However, the predictive accuracies of these
models vary notably across different species, highlighting the need
to improve the accuracy and specificity of traditional prediction
models. The integration of experimental detection tools with
predictive models can lead to more accurate experimental data,
ultimately enhancing the precision of research outcomes.
NSUN6 also modifies mRNA, primarily by targeting the
3′UTR within the consensus sequence motif CTCCA present in hairpin
loop structures, which marks the translation termination site.
NSUN6 expression varies across human tissues, being most prevalent
in the testes and least prevalent in the blood (68). The role of NSUN6 in tumorigenesis
remains a subject of ongoing scientific debate; in cancers such as
pancreatic cancer, glioblastoma and esophageal squamous cell
carcinoma, NSUN6 acts as a tumor suppressor, potentially inhibiting
malignant progression or modulating therapeutic responsiveness
through m5C-mediated mRNA methylation (Table I) (69–71).
Conversely, NSUN6 is frequently upregulated in cervical cancer,
gastric cancer and triple-negative breast cancer, where it
facilitates tumor initiation and aggressiveness (Table I) (72–74).
This functional dichotomy may be linked to the complexity of the
tumor microenvironment (TME), where extracellular matrix
interactions and metabolic heterogeneity collectively influence
oncogenesis and metastatic behavior (75,76).
Immune infiltration patterns also differ markedly between NSUN2 and
NSUN6: NSUN2 expression predominates in monocytes/macrophages and
proliferative T cells, whereas NSUN6 is most abundant in regulatory
T cells (74,77). Such distinctions may help explain
the context-dependent roles of NSUN6 across cancer types and
disease stages. Future studies should aim to elucidate the
integrated expression dynamics of NSUN6 within both the tumor
parenchyma and stromal compartments to advance personalized
oncology strategies.
TET2, a recognized tumor suppressor, inhibits
myeloid tumors in an enzyme activity-dependent manner and serves a
marked role in inflammatory responses (78); however, the involvement of TET2 in
m5C methylation modification remains controversial. A
study by Li et al (79)
demonstrated that TET2 functions as an m5C demethylase
and its deficiency leads to the accumulation of TSPAN13 mRNA,
promoting leukemogenesis and enhancing the self-renewal capacity of
leukemia stem cells. Zou et al (31) proposed that TET2 acts as a DNA 5mC
(abbreviated as ‘5mC’ in DNA) oxidase in leukemia; additionally,
TET2-mediated oxidation also serves a role in glioma (80). However, TET2 promotes M2 macrophage
polarization in allergic rhinitis by regulating m5C
oxidation in mRNA (81). In
patients with diabetic nephropathy (DN), TET2 expression is
associated with renal impairment, and TET2-mediated m5C
modification regulates mitophagy, suggesting its potential as a
therapeutic strategy for DN (82).
Due to the multifaceted biological functions of TET2, the specific
molecular mechanisms by which it acts as an RNA m5C
demethylase in tumors have not yet been fully elucidated. This lack
of clarity may arise from its broad range of actions and
involvement in multiple signaling pathways, which could contribute
to a non-specific role in RNA methylation modification. Future
research should integrate both the oxidative and demethylating
functions of TET2 to further explore its impact on the TME, thus
providing directions for the development of novel targeted or
immunotherapeutic strategies.
Among the tRNA methyltransferases, DNMT2 and NSUN
enzymes have been extensively studied (23). The deletion of DNMT2 in mouse cells
increases the proportion of uncharged tRNAAsp, resulting
in decreased translation efficiency of proteins containing
polyaspartic acid (86); this
deletion also triggers tRNA fragmentation and decreases
steady-state levels (87,88). Additionally, the m5C
modification safeguards tRNAs against stress-induced
endonuclease-mediated fragmentation, thereby ensuring the accurate
translation of near-cognate codons. For example, DNMT2-mediated
m5C modification in tRNAAsp helps to
distinguish it from tRNAGlu, preventing amino acid
misincorporation (89). Studies
have also shown that a reduction in DNMT2 methyltransferase
activity due to somatic cancer mutations strongly associated with
decreased tRNA levels (90,91).
Furthermore, the TET2-mediated oxidation of m5C in tRNA
disrupts the binding of readers to RNA and produces
5-hydroxymethylcytosine (hm5C), altering tRNA
methylation and thus impacting translation (25). In anaplastic thyroid cancer (ATC),
NSUN2 knockdown markedly reduces tRNA m5C modification,
suggesting it has a role in stabilizing tRNA in ATC, facilitating
the transport of amino acids such as leucine and enhancing
translation efficiency. Moreover, NSUN2 also contributes to the
formation, proliferation and drug resistance of ATC cells (Table I) (92). In addition to influencing codon
translation, NSUN2 also regulates the cleavage site of
tRNAArg in colorectal cancer in an
m5C-dependent manner under hypoxic conditions and serves
a critical role in tumor metastasis (Table I) (93).
A growing body of evidence links alterations in
rRNA modification levels and defects in components of the rRNA
modification machinery to tumors. rRNAs are extensively modified
during their transcription and subsequent maturation in the
nucleolus, nucleus and cytoplasm (102,103). The m5C modification in
rRNA is associated with ribosome synthesis and protein translation;
the m5C modification stabilizes the RNA structure by
promoting base stacking and increasing the thermal stability of
hydrogen bonding with guanine. In functionally crucial areas of the
ribosome, m5C modification helps stabilize rRNA folding,
which is essential for efficient ribosome function (42,104,105). Studies have shown that NSUN4 is
crucial for the biosynthesis of mitochondrial rRNA; the knockout of
NSUN4 results in the loss of methylation at position C911 of
mitochondrial rRNA in mouse hearts, suggesting its key role in
coordinating mitochondrial ribosome biosynthesis (106,107). Similarly, NSUN5 is necessary for
the specific methylation of 28S rRNA in humans and mice, and its
absence reduces protein synthesis, underscoring the importance of
NSUN5 in ribosome function and translation regulation (108). In hepatocellular carcinoma,
overexpression of NSUN5 upregulates the m5C level of 28S
rRNA, which in turn promotes growth and metastasis (Table I) (109). Additionally, the depletion of
YTHDF2, a multifunctional reader, results in a marked increase in
m5C levels at various sites within rRNA. This alteration
may influence translation fidelity, suggesting a potential link
between the role of YTHDF2 in recognizing m5C and its
impact on translation accuracy (42). A number of studies have shown that
YTHDF2 is dysregulated in malignant tumors such as bladder cancer,
liver cancer, gastric cancer, osteosarcoma and blood system by
m6A methylation modification (110–115), while, to the best of our
knowledge, no studies have investigated the specific mechanism of
m5C methylation modification in malignant tumors.
Further research is expected to uncover the potential biological
functions of YTHDF2.
The sncRNAs, which include microRNAs (miRNAs),
PIWI-interacting RNAs, small interfering RNAs and tRNA-derived
small RNAs (tsRNAs), play vital roles within cells by binding to
proteins, regulating transcription factors and participating in
genome stability (24).
High-throughput next-generation sequencing-based methods, such as
bisulphite miRNA sequencing, and an analysis pipeline such as
methylation assessment of miRNAs after bisulphite analysis, have
revealed widespread m5C modifications in miRNAs
(116). The miRNAs regulate gene
transcription and posttranscriptional regulation through partial
base pairing with sequences mainly in the 3′UTRs of target mRNAs,
guiding the RNA-induced silencing complex to suppress mRNA
translation. The m5C modification in miRNA affects its
pairing with mRNA, potentially disrupting its gene-silencing
activity (117). To the best of
our knowledge, few studies have investigated the role of RNA
m5C methylation in miRNAs in cancer, although some
studies have explored DNA methylation. In glioblastoma, miRNA is
methylated at cytosine residues through complexes such as
DNMT3A/AGO4, which abrogates its inhibitory effect on gene
expression and is associated with poor prognosis (118). Additionally, a pivotal study has
successfully developed a serum diagnostic signature using
m5C-modified miRNAs (119). Due to the potential crosstalk
between DNA methylation and RNA methylation, studying the role of
RNA methylation in miRNAs may lead to notable breakthroughs in
tumor treatment and diagnosis (119).
tsRNAs, derived from tRNA, interact with
cytoplasmic ribonucleoproteins (RNPs) to form tsRNA-RNP complexes.
These complexes can bind to Argonaute family proteins, effectively
mediating posttranscriptional gene silencing through mechanisms
such as RNA interference or direct inhibition of mRNA translation
(120). DNMT2-mediated
m5C modifications in mice markedly affect sperm tsRNA
levels, with the loss of DNMT2 disrupting the transmission of
high-fat diet-induced metabolic disorders mediated by sperm sncRNA
to offspring (24). It has been
demonstrated that tsRNAs can influence translation rates through
YBX1. In breast cancer cells, tsRNAs can induce tumor-suppressive
effects by replacing YBX1 with oncogenic mRNAs, thereby promoting
the degradation of such transcripts (121–123). Thus, the m5C
modifications in tRNA notably influence tsRNA functionality, and
the regulation of protein translation mechanisms by tsRNA
represents a promising new avenue of investigation in cancer
studies.
The utilization of innovative detection
technologies has notably advanced research into m5C
modifications of sncRNAs. These advancements enhance the
understanding of intracellular signaling mechanisms associated with
various diseases. These findings open new avenues for targeted
sncRNA therapies, offering more precise and effective treatment
options.
lncRNAs, a category of ncRNAs >200 nucleotides
in length, constitute a substantial part of the mammalian
transcriptome (124). lncRNAs
participate in various physiological and pathological processes in
normal tissues and tumors by interacting with RNA-binding proteins.
These processes include chromatin modification, transcriptional
activation, transcriptional interference and nuclear transport
(125,126). One of the enzymes involved in
modifying lncRNAs is NSUN2, which has been linked to the
development of various cancers. In gastric cancer, NSUN2 modifies
the lncRNA NR_033928, which in turn affects the stability of
glutaminase mRNA and participates in metabolic reprogramming
(127). Similarly, in
cholangiocarcinoma, NSUN2 interacts with the lncRNA NKILA,
promoting disease progression (Table
I) (128). In the nervous
system, lncRNAs are involved in ischemic stroke through mechanisms
such as calcium overload, oxidative stress, hypoxia and
inflammatory responses (129).
Notably, MeRIP-Seq analysis revealed increased methylation levels
and frequency in lncRNAs in the middle cerebral artery occlusion
model, which simulates human ischemic stroke. These differentially
methylated lncRNAs are associated with several notable pathways,
including mTOR signaling, Rap1 signal transduction, pyrimidine
metabolism, dopamine receptor binding and phosphatidylinositol
phosphate kinase activity (130).
circRNAs are a class of covalently closed RNA
molecules lacking free 5′ and 3′ ends, and most circRNAs are
non-coding; however, growing evidence indicates that some can
encode functional proteins (132). Although circRNAs play marked
roles in the pathogenesis of various diseases, the functional
implications of m5C methylation on circRNAs remain
largely unexplored. The emerging role of m5C-modified
circRNAs in lung cancer pathogenesis has attracted increasing
attention in recent studies. Chen et al (133) reported that NSUN2-mediated
m5C modification of circFAM190B enhances its stability,
suppresses autophagy and ultimately drives tumorigenesis. The study
by Wu et al (134)
revealed that NSUN4-catalyzed m5C modification of
circERI3 facilitates its nuclear export, which in turn promotes the
development and progression of lung cancer by enhancing
mitochondrial energy metabolism. Cai et al (135) identified circRREB1 as an
m5C-modified circRNA in lung cancer. This modification,
mediated by the methyltransferase NSUN2 and recognized by the
reader protein ALYREF, activates mitophagy to promote tumor
progression. The functional relevance of circRNA m5C
modification has also been implicated in other malignancies,
including esophageal squamous cell carcinoma and breast cancer
(Table I) (136,137). These findings demonstrate that
m5C methylation can regulate broader processes such as
autophagy and mitochondrial metabolism through the modification of
circRNAs. Given the fundamental role of RNA modifications in
physiology, research on m5C methylation in ncRNAs has
been increasing, extending beyond mRNA, tRNA and rRNA to include
insights into the mechanisms of lncRNA and circRNA modification.
Future studies should further expand the scope from cancer to other
pathological and functional fields, providing new perspectives for
diagnosing diseases through non-invasive examinations.
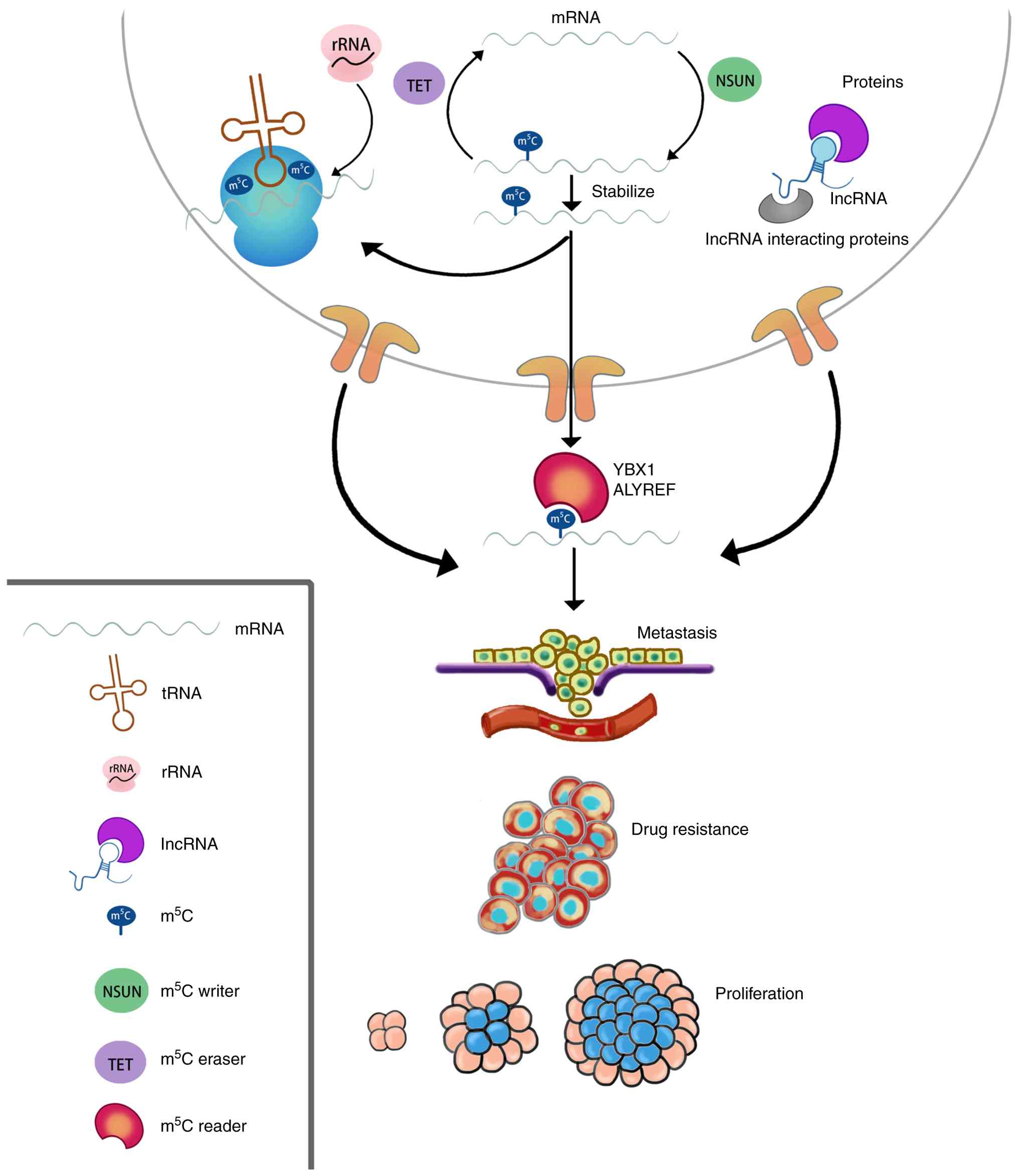
RNA modifications, particularly RNA methylation,
represent a rapidly evolving field and play an indispensable role
in regulating gene transcription, expression, editing, stability
and degradation. A notable number of tumors exploit RNA
modifications as a mechanism to enhance or suppress gene
expression, ultimately leading to tumor development (Fig. 1). Methylation is critically
involved in diverse physiological and pathological processes;
notably, its reversible nature underscores the considerable
therapeutic potential of targeting specific methylation sites in
cancer, positioning it as a promising strategy for future
early-stage interventions. Pioneering this approach, the DNA
methyltransferase inhibitor 5-azacytidine was developed to reverse
aberrant methylation and is now a standard treatment for high-risk
myelodysplastic syndromes (138).
This success has spurred interest in targeting RNA modifications as
a novel strategy for treating various diseases. Key research
directions in precision medicine now include developing
small-molecule inhibitors against specific modifying enzymes and
utilizing serum methylation patterns to predict disease
progression. However, both DNA- and RNA-methylation-targeted
therapies still face notable challenges, such as selectively
targeting the oncogenic functions of these proteins without
inducing widespread epigenetic disruption. Inhibiting methylation
or demethylation at specific loci often results in limited
efficacy, likely due to functional redundancy and crosstalk among
different methyltransferases. Therefore, future efforts must
prioritize understanding these interactions within specific tumor
contexts and developmental stages, while rigorously evaluating the
safety profiles of these emerging therapies (139).
Not applicable.
The present study was supported by Provincial Natural Science
Foundation of Hunan (grant no. 2023JJ30769, 2022JJ30806), Health
Research Project of Hunan Provincial Health Commission (grant no.
W20243103) and Natural Science Foundation of Changsha (grant no.
kq2208345).
Not applicable.
The present review was designed by QJ and YG. QJ
and YG wrote the manuscript, and SL contributed to the drafting,
editing, and critical revision of the manuscript. MZ and TT
participated in revising the manuscript for important intellectual
content. All authors read and approved the final manuscript. Data
authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Dai X, Ren T, Zhang Y and Nan N:
Methylation multiplicity and its clinical values in cancer. Expert
Rev Mol Med. 23:e22021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Orji OC, Stones J, Rajani S, Markus R, Öz
MD and Knight HM: Global Co-regulatory Cross Talk Between
m6A and m5C RNA methylation systems
coordinate cellular responses and brain disease pathways. Mol
Neurobiol. 62:5006–5021. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xie S, Hagen D, Becker GM, Davenport KM,
Shira KA, Stegemiller MR, Thorne JW, Khilji S, Konetchy D,
Villamediana P, et al: Analyzing the relationship of RNA and DNA
methylation with gene expression. Genome Biol. 26:1402025.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Han X, Wang M, Zhao YL, Yang Y and Yang
YG: RNA methylations in human cancers. Semin Cancer Biol.
75:97–115. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roundtree IA, Evans ME, Pan T and He C:
Dynamic RNA modifications in gene expression regulation. Cell.
169:1187–1200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nombela P, Miguel-López B and Blanco S:
The role of m6A, m5C and Ψ RNA modifications
in cancer: Novel therapeutic opportunities. Mol Cancer. 20:182021.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Y, Wei J, Feng L, Li O, Huang L, Zhou
S, Xu Y, An K, Zhang Y, Chen R, et al: Aberrant m5C
hypermethylation mediates intrinsic resistance to gefitinib through
NSUN2/YBX1/QSOX1 axis in EGFR-mutant non-small-cell lung cancer.
Mol Cancer. 22:812023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu L, Chen Y, Zhang T, Cui G, Wang W,
Zhang G, Li J, Zhang Y, Wang Y, Zou Y, et al: YBX1 promotes
esophageal squamous cell carcinoma progression via m5C-dependent
SMOX mRNA stabilization. Adv Sci (Weinh). 11:e23023792024.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang JZ, Zhu W, Han J, Yang X, Zhou R, Lu
HC, Yu H, Yuan WB, Li PC, Tao J, et al: The role of the
HIF-1α/ALYREF/PKM2 axis in glycolysis and tumorigenesis of bladder
cancer. Cancer Commun (Lond). 41:560–575. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cui L, Ma R, Cai J, Guo C, Chen Z, Yao L,
Wang Y, Fan R, Wang X and Shi Y: RNA modifications: Importance in
immune cell biology and related diseases. Signal Transduct Target
Ther. 7:3342022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qiu L, Jing Q, Li Y and Han J: RNA
modification: Mechanisms and therapeutic targets. Mol Biomed.
4:252023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang L, Tian H, Min Q, You H, Yin M, Yang
L, Zhao Y, Wu X, Li M, Du F, et al: Decoding the epitranscriptome:
A new frontier for cancer therapy and drug resistance. Cell Commun
Signal. 22:5132024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kong Y, Yu J, Ge S and Fan X: Novel
insight into RNA modifications in tumor immunity: Promising targets
to prevent tumor immune escape. Innovation (Camb).
4:1004522023.PubMed/NCBI
|
|
14
|
Chen B, Hong Y, Zhai X, Deng Y, Hu H, Tian
S, Zhang Y, Ren X, Zhao J and Jiang C: m6A and m5C modification of
GPX4 facilitates anticancer immunity via STING activation. Cell
Death Dis. 14:8092023. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Helm M: Post-transcriptional nucleotide
modification and alternative folding of RNA. Nucleic Acids Res.
34:721–733. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schaefer M, Pollex T, Hanna K and Lyko F:
RNA cytosine methylation analysis by bisulfite sequencing. Nucleic
Acids Res. 37:e122009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Y and Santi DV: m5C RNA and m5C DNA
methyl transferases use different cysteine residues as catalysts.
Proc Natl Acad Sci USA. 97:8263–8265. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
King MY and Redman KL: RNA
methyltransferases utilize two cysteine residues in the formation
of 5-methylcytosine. Biochemistry. 41:11218–11225. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bohnsack KE, Höbartner C and Bohnsack MT:
Eukaryotic 5-methylcytosine (m5C) RNA methyltransferases:
Mechanisms, cellular functions, and links to disease. Genes
(Basel). 10:1022019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jeltsch A: Beyond watson and crick: DNA
methylation and molecular enzymology of DNA methyltransferases.
Chembiochem. 3:274–293. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Moon J, Lee H, Jang Y and Kim SK:
NSUN-mediated m5C RNA modification in stem cell regulation. Cells.
14:16092025. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang X, Yang Y, Sun BF, Chen YS, Xu JW,
Lai WY, Li A, Wang X, Bhattarai DP, Xiao W, et al: 5-methylcytosine
promotes mRNA export-NSUN2 as the methyltransferase and ALYREF as
an m5C reader. Cell Res. 27:606–625. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jeltsch A, Ehrenhofer-Murray A, Jurkowski
TP, Lyko F, Reuter G, Ankri S, Nellen W, Schaefer M and Helm M:
Mechanism and biological role of Dnmt2 in nucleic acid methylation.
RNA Biol. 14:1108–1123. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Zhang X, Shi J, Tuorto F, Li X,
Liu Y, Liebers R, Zhang L, Qu Y, Qian J, et al: Dnmt2 mediates
intergenerational transmission of paternally acquired metabolic
disorders through sperm small non-coding RNAs. Nat Cell Biol.
20:535–540. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shen H, Ontiveros RJ, Owens MC, Liu MY,
Ghanty U, Kohli RM and Liu KF: TET-mediated 5-methylcytosine
oxidation in tRNA promotes translation. J Biol Chem.
296:1000872021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ito S, Shen L, Dai Q, Wu SC, Collins LB,
Swenberg JA, He C and Zhang Y: Tet proteins can convert
5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine.
Science. 333:1300–1303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shen Q, Zhang Q, Shi Y, Shi Q, Jiang Y, Gu
Y, Li Z, Li X, Zhao K, Wang C, et al: Tet2 promotes pathogen
infection-induced myelopoiesis through mRNA oxidation. Nature.
554:123–127. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang W, Lan MD, Qi CB, Zheng SJ, Wei SZ,
Yuan BF and Feng YQ: Formation and determination of the oxidation
products of 5-methylcytosine in RNA. Chem Sci. 7:5495–5502. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moen EL, Mariani CJ, Zullow H, Jeff-Eke M,
Litwin E, Nikitas JN and Godley LA: New themes in the biological
functions of 5-methylcytosine and 5-hydroxymethylcytosine. Immunol
Rev. 263:36–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fu L, Guerrero CR, Zhong N, Amato NJ, Liu
Y, Liu S, Cai Q, Ji D, Jin SG, Niedernhofer LJ, et al: Tet-mediated
formation of 5-hydroxymethylcytosine in RNA. J Am Chem Soc.
136:11582–11585. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zou Z, Dou X, Li Y, Zhang Z, Wang J, Gao
B, Xiao Y, Wang Y, Zhao L, Sun C, et al: RNA m5C
oxidation by TET2 regulates chromatin state and leukaemogenesis.
Nature. 634:986–994. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Haag S, Sloan KE, Ranjan N, Warda AS,
Kretschmer J, Blessing C, Hübner B, Seikowski J, Dennerlein S,
Rehling P, et al: NSUN3 and ABH1 modify the wobble position of
mt-tRNAMet to expand codon recognition in mitochondrial
translation. EMBO J. 35:2104–2119. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Evke S, Lin Q, Melendez JA and Begley TJ:
Epitranscriptomic reprogramming is required to prevent stress and
damage from acetaminophen. Genes (Basel). 13:4212022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhong J, Xu Z, Ding N, Wang Y and Chen W:
The biological function of demethylase ALKBH1 and its role in human
diseases. Heliyon. 10:e334892024. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen YS, Yang WL, Zhao YL and Yang YG:
Dynamic transcriptomic m5 C and its regulatory role in
RNA processing. Wiley Interdiscip Rev RNA. 12:e16392021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li M, Tao Z, Zhao Y, Li L, Zheng J, Li Z
and Chen X: 5-methylcytosine RNA methyltransferases and their
potential roles in cancer. J Transl Med. 20:2142022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dominissini D and Rechavi G:
5-methylcytosine mediates nuclear export of mRNA. Cell Res.
27:717–719. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang Y, Wang L, Han X, Yang WL, Zhang M,
Ma HL, Sun BF, Li A, Xia J, Chen J, et al: RNA 5-methylcytosine
facilitates the Maternal-to-Zygotic transition by preventing
maternal mRNA decay. Mol Cell. 75:1188–1202.e11. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chang CT, Hautbergue GM, Walsh MJ,
Viphakone N, van Dijk TB, Philipsen S and Wilson SA: Chtop is a
component of the dynamic TREX mRNA export complex. EMBO J.
32:473–486. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lyabin DN, Eliseeva IA and Ovchinnikov LP:
YB-1 protein: Functions and regulation. Wiley Interdiscip Rev RNA.
5:95–110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang J, Li H and Niswander LA:
m5C methylated lncRncr3-MeCP2 interaction restricts
miR124a-initiated neurogenesis. Nat Commun. 15:51362024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dai X, Gonzalez G, Li L, Li J, You C, Miao
W, Hu J, Fu L, Zhao Y, Li R, et al: YTHDF2 Binds to
5-methylcytosine in RNA and modulates the maturation of ribosomal
RNA. Anal Chem. 92:1346–1354. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gaston KW and Limbach PA: The
identification and characterization of non-coding and coding RNAs
and their modified nucleosides by mass spectrometry. RNA Biol.
11:1568–1585. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Thüring K, Schmid K, Keller P and Helm M:
Analysis of RNA modifications by liquid chromatography-tandem mass
spectrometry. Methods. 107:48–56. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang N, Shi S, Jia TZ, Ziegler A, Yoo B,
Yuan X, Li W and Zhang S: A general LC-MS-based RNA sequencing
method for direct analysis of multiple-base modifications in RNA
mixtures. Nucleic Acids Res. 47:e1252019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gao Y and Fang J: RNA 5-methylcytosine
modification and its emerging role as an epitranscriptomic mark.
RNA Biol. 18:117–127. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yuan F, Bi Y, Siejka-Zielinska P, Zhou YL,
Zhang XX and Song CX: Bisulfite-free and base-resolution analysis
of 5-methylcytidine and 5-hydroxymethylcytidine in RNA with
peroxotungstate. Chem Commun (Camb). 55:2328–2331. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang L, Perrera V, Saplaoura E, Apelt F,
Bahin M, Kramdi A, Olas J, Mueller-Roeber B, Sokolowska E, Zhang W,
et al: m5C methylation guides systemic transport of
messenger RNA over graft junctions in plants. Curr Biol.
29:2465–2476.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cui X, Liang Z, Shen L, Zhang Q, Bao S,
Geng Y, Zhang B, Leo V, Vardy LA, Lu T, et al: 5-methylcytosine RNA
methylation in arabidopsis thaliana. Mol Plant. 10:1387–1399. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hussain S, Benavente SB, Nascimento E,
Dragoni I, Kurowski A, Gillich A, Humphreys P and Frye M: The
nucleolar RNA methyltransferase Misu (NSun2) is required for
mitotic spindle stability. J Cell Biol. 186:27–40. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Trixl L and Lusser A: The dynamic RNA
modification 5-methylcytosine and its emerging role as an
epitranscriptomic mark. Wiley Interdiscip Rev RNA. 10:e15102019.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Khoddami V and Cairns BR: Identification
of direct targets and modified bases of RNA cytosine
methyltransferases. Nat Biotechnol. 31:458–464. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Khoddami V and Cairns BR:
Transcriptome-wide target profiling of RNA cytosine
methyltransferases using the mechanism-based enrichment procedure
Aza-IP. Nat Protoc. 9:337–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Garalde DR, Snell EA, Jachimowicz D, Sipos
B, Lloyd JH, Bruce M, Pantic N, Admassu T, James P, Warland A, et
al: Highly parallel direct RNA sequencing on an array of nanopores.
Nat Methods. 15:201–206. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sun K, Li J, Chen C, Zhou X, Ma G, Mao L,
Tang Q, Ma B, Li D, Chen Z, et al: Advances in nanopore direct RNA
sequencing and its impact on biological research. Biotechnol Adv.
85:1087102025. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhao L, Zhang H, Kohnen MV, Prasad K, Gu L
and Reddy ASN: Analysis of transcriptome and epitranscriptome in
plants using PacBio Iso-Seq and nanopore-based direct RNA
sequencing. Front Genet. 10:2532019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wu Y, Shao W, Liu S, Wang L, Xu P, Zhang
X, Song H, Li X, Wang J and Yu X: Simultaneous profiling of
ac4C and m5C modifications from nanopore
direct RNA sequencing. Int J Biol Macromol. 305:1408632025.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Song J, Zhai J, Bian E, Song Y, Yu J and
Ma C: Transcriptome-Wide annotation of m5C RNA
modifications using machine learning. Front Plant Sci. 9:5192018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lv H, Zhang ZM, Li SH, Tan JX, Chen W and
Lin H: Evaluation of different computational methods on
5-methylcytosine sites identification. Brief Bioinform. 21:982–995.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhang M, Xu Y, Li L, Liu Z, Yang X and Yu
DJ: Accurate RNA 5-methylcytosine site prediction based on
heuristic physical-chemical properties reduction and classifier
ensemble. Anal Biochem. 550:41–48. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen X, Xiong Y, Liu Y, Chen Y, Bi S and
Zhu X: m5CPred-SVM: A novel method for predicting m5C sites of RNA.
BMC Bioinformatics. 21:4892020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Sun PP, Chen YB, Liu B, Gao YX, Han Y, He
F and Ji JC: DeepMRMP: A new predictor for multiple types of RNA
modification sites using deep learning. Math Biosci Eng.
16:6231–6241. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hussain S, Sajini AA, Blanco S, Dietmann
S, Lombard P, Sugimoto Y, Paramor M, Gleeson JG, Odom DT, Ule J and
Frye M: NSun2-mediated cytosine-5 methylation of vault noncoding
RNA determines its processing into regulatory small RNAs. Cell Rep.
4:255–261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
George H, Ule J and Hussain S:
Illustrating the epitranscriptome at nucleotide resolution using
methylation-iCLIP (miCLIP). Methods Mol Biol. 1562:91–106. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Amort T, Rieder D, Wille A,
Khokhlova-Cubberley D, Riml C, Trixl L, Jia XY, Micura R and Lusser
A: Distinct 5-methylcytosine profiles in poly(A) RNA from mouse
embryonic stem cells and brain. Genome Biol. 18:12017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Popis MC, Blanco S and Frye M:
Posttranscriptional methylation of transfer and ribosomal RNA in
stress response pathways, cell differentiation, and cancer. Curr
Opin Oncol. 28:65–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chen SJ, Zhang J, Zhou T, Rao SS, Li Q,
Xiao LY, Wei ST and Zhang HF: Epigenetically upregulated NSUN2
confers ferroptosis resistance in endometrial cancer via
m5C modification of SLC7A11 mRNA. Redox Biol.
69:1029752024. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Selmi T, Hussain S, Dietmann S, Heiß M,
Borland K, Flad S, Carter JM, Dennison R, Huang YL, Kellner S, et
al: Sequence- and structure-specific cytosine-5 mRNA methylation by
NSUN6. Nucleic Acids Res. 49:1006–1022. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Awah CU, Winter J, Mazdoom CM and Ogunwobi
OO: NSUN6, an RNA methyltransferase of 5-mC controls glioblastoma
response to temozolomide (TMZ) via NELFB and RPS6KB2 interaction.
Cancer Biol Ther. 22:587–597. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yang R, Liang X, Wang H, Guo M, Shen H,
Shi Y, Liu Q, Sun Y, Yang L and Zhan M: The RNA methyltransferase
NSUN6 suppresses pancreatic cancer development by regulating cell
proliferation. EBioMedicine. 63:1031952021. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Han H, Sun Y, Wei W, Huang Z, Cheng M, Qiu
H, Wang J, Zheng S, Liu L, Zhang Q, et al: RNA modification-related
genes illuminate prognostic signature and mechanism in esophageal
squamous cell carcinoma. iScience. 27:1093272024. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Guo J, Wu B, Wang S, Huang D and Hu Y:
NSUN6 promotes gastric cancer progression by stabilizing CEBPZ mRNA
in a m5C-dependent manner. Appl Biochem Biotechnol.
197:7296–7313. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Yu M, Ni M, Xu F, Liu C, Chen L, Li J, Xia
S, Diao Y, Chen J, Zhu J, et al: NSUN6-mediated 5-methylcytosine
modification of NDRG1 mRNA promotes radioresistance in cervical
cancer. Mol Cancer. 23:1392024. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Huang Z, Pan J, Wang H, Du X, Xu Y, Wang Z
and Chen D: Prognostic significance and tumor immune
microenvironment heterogenicity of m5C RNA methylation regulators
in triple-negative breast cancer. Front Cell Dev Biol.
9:6575472021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Nasrollahzadeh E, Razi S, Keshavarz-Fathi
M, Mazzone M and Rezaei N: Pro-tumorigenic functions of macrophages
at the primary, invasive and metastatic tumor site. Cancer Immunol
Immunother. 69:1673–1697. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Pernot S, Evrard S and Khatib AM: The
Give-and-Take interaction between the tumor microenvironment and
immune cells regulating tumor progression and repression. Front
Immunol. 13:8508562022. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Yan D, Xie Y, Huang L, Zhang Y, Gu R, Xie
H, Huang X and Luo H: RNA m5C methylation orchestrates BLCA
progression via macrophage reprogramming. J Cell Mol Med.
27:2398–2411. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhang Q, Zhao K, Shen Q, Han Y, Gu Y, Li
X, Zhao D, Liu Y, Wang C, Zhang X, et al: Tet2 is required to
resolve inflammation by recruiting Hdac2 to specifically repress
IL-6. Nature. 525:389–393. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Li Y, Xue M, Deng X, Dong L, Nguyen LXT,
Ren L, Han L, Li C, Xue J, Zhao Z, et al: TET2-mediated mRNA
demethylation regulates leukemia stem cell homing and self-renewal.
Cell Stem Cell. 30:1072–1090.e10. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Wu R, Sun C, Chen X, Yang R, Luan Y, Zhao
X, Yu P, Luo R, Hou Y, Tian R, et al: NSUN5/TET2-directed
chromatin-associated RNA modification of 5-methylcytosine to
5-hydroxymethylcytosine governs glioma immune evasion. Proc Natl
Acad Sci USA. 121:e23216111212024. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Fan W, Liu P, Tan L, Lv H, Zhou H, Tao Z
and Xu Y: Tet2 modulates M2 macrophage polarization via mRNA
5-methylcytosine in allergic rhinitis. Int Immunopharmacol.
143:1134952024. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Ma XH, Hu ZY, Wang YK, Ma Y, Liu GP and
Liu XQ: TET2 deficiency exacerbates podocyte injury and mitophagy
disorder in diabetic nephropathy by regulating M5C methylation of
Bcas3. FASEB J. 39:e709662025. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Chen X, Li A, Sun BF, Yang Y, Han YN, Yuan
X, Chen RX, Wei WS, Liu Y, Gao CC, et al: 5-methylcytosine promotes
pathogenesis of bladder cancer through stabilizing mRNAs. Nat Cell
Biol. 21:978–990. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Yang Q, Wang M, Xu J, Yu D, Li Y, Chen Y
and Zhang X, Zhang J, Gu J and Zhang X: LINC02159 promotes
non-small cell lung cancer progression via ALYREF/YAP1 signaling.
Mol Cancer. 22:1222023. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Schumann U, Zhang HN, Sibbritt T, Pan A,
Horvath A, Gross S, Clark SJ, Yang L and Preiss T: Multiple links
between 5-methylcytosine content of mRNA and translation. BMC Biol.
18:402020. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Shanmugam R, Fierer J, Kaiser S, Helm M,
Jurkowski TP and Jeltsch A: Cytosine methylation of tRNA-Asp by
DNMT2 has a role in translation of proteins containing poly-Asp
sequences. Cell Discov. 1:150102015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Schaefer M, Pollex T, Hanna K, Tuorto F,
Meusburger M, Helm M and Lyko F: RNA methylation by Dnmt2 protects
transfer RNAs against stress-induced cleavage. Genes Dev.
24:1590–1595. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Tuorto F, Liebers R, Musch T, Schaefer M,
Hofmann S, Kellner S, Frye M, Helm M, Stoecklin G and Lyko F: RNA
cytosine methylation by Dnmt2 and NSun2 promotes tRNA stability and
protein synthesis. Nat Struct Mol Biol. 19:900–905. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Tuorto F, Herbst F, Alerasool N, Bender S,
Popp O, Federico G, Reitter S, Liebers R, Stoecklin G, Gröne HJ, et
al: The tRNA methyltransferase Dnmt2 is required for accurate
polypeptide synthesis during haematopoiesis. EMBO J. 34:2350–2362.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Elhardt W, Shanmugam R, Jurkowski TP and
Jeltsch A: Somatic cancer mutations in the DNMT2 tRNA
methyltransferase alter its catalytic properties. Biochimie.
112:66–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Li H, Liu H, Zhu D, Dou C, Gang B, Zhang M
and Wan Z: Biological function molecular pathways and druggability
of DNMT2/TRDMT1. Pharmacol Res. 205:1072222024. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Li P, Wang W, Zhou R, Ding Y and Li X: The
m5 C methyltransferase NSUN2 promotes codon-dependent
oncogenic translation by stabilising tRNA in anaplastic thyroid
cancer. Clin Transl Med. 13:e14662023. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Luan N, Cao Y, Sun J, Ye X, Zhou Q, Tang Z
and Wang J: Hypoxia-induced NSUN2 promotes the progression of colon
cancer by modulating the discrepant cleavage of tRNA-Arg. Int
Immunopharmacol. 161:1150232025. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Delaunay S, Pascual G, Feng B, Klann K,
Behm M, Hotz-Wagenblatt A, Richter K, Zaoui K, Herpel E, Münch C,
et al: Mitochondrial RNA modifications shape metabolic plasticity
in metastasis. Nature. 607:593–603. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Bilbille Y, Gustilo EM, Harris KA, Jones
CN, Lusic H, Kaiser RJ, Delaney MO, Spremulli LL, Deiters A and
Agris PF: The human mitochondrial tRNAMet: Structure/function
relationship of a unique modification in the decoding of
unconventional codons. J Mol Biol. 406:257–274. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Van Haute L, Powell CA and Minczuk M:
Dealing with an unconventional genetic code in mitochondria: The
biogenesis and pathogenic defects of the 5-formylcytosine
modification in mitochondrial tRNA(Met). Biomolecules. 7:242017.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kawarada L and Suzuki T, Ohira T, Hirata
S, Miyauchi K and Suzuki T: ALKBH1 is an RNA dioxygenase
responsible for cytoplasmic and mitochondrial tRNA modifications.
Nucleic Acids Res. 45:7401–7415. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Takemoto C, Spremulli LL, Benkowski LA,
Ueda T, Yokogawa T and Watanabe K: Unconventional decoding of the
AUA codon as methionine by mitochondrial tRNAMet with the anticodon
f5CAU as revealed with a mitochondrial in vitro translation system.
Nucleic Acids Res. 37:1616–1627. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Huber SM, van Delft P, Tanpure A, Miska EA
and Balasubramanian S: 2′-O-Methyl-5-hydroxymethylcytidine: A
second oxidative derivative of 5-methylcytidine in RNA. J Am Chem
Soc. 139:1766–1769. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Luu M and Visekruna A: Targeting metabolic
rewiring might decrease spread of tumor cells: Mitochondrial tRNA
modifications promote cancer metastasis. Signal Transduct Target
Ther. 7:3602022. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Nakano S and Suzuki T, Kawarada L, Iwata
H, Asano K and Suzuki T: NSUN3 methylase initiates 5-formylcytidine
biogenesis in human mitochondrial tRNA(Met). Nat Chem Biol.
12:546–551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Haruehanroengra P, Zheng YY, Zhou Y, Huang
Y and Sheng J: RNA modifications and cancer. RNA Biol.
17:1560–1575. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Sloan KE, Warda AS, Sharma S, Entian KD,
Lafontaine DLJ and Bohnsack MT: Tuning the ribosome: The influence
of rRNA modification on eukaryotic ribosome biogenesis and
function. RNA Biol. 14:1138–1152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Li Y, Sundquist K, Zhang N, Wang X,
Sundquist J and Memon AA: Mitochondrial related genome-wide
mendelian randomization identifies putatively causal genes for
multiple cancer types. EBioMedicine. 88:1044322023. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Sharma S, Yang J, Watzinger P, Kötter P
and Entian KD: Yeast Nop2 and Rcm1 methylate C2870 and C2278 of the
25S rRNA, respectively. Nucleic Acids Res. 41:9062–9076. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Metodiev MD, Spåhr H, Loguercio Polosa P,
Meharg C, Becker C, Altmueller J, Habermann B, Larsson NG and
Ruzzenente B: NSUN4 is a dual function mitochondrial protein
required for both methylation of 12S rRNA and coordination of
mitoribosomal assembly. PLoS Genet. 10:e10041102014. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Spåhr H, Habermann B, Gustafsson CM,
Larsson NG and Hallberg BM: Structure of the human MTERF4-NSUN4
protein complex that regulates mitochondrial ribosome biogenesis.
Proc Natl Acad Sci USA. 109:15253–15258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Heissenberger C, Liendl L, Nagelreiter F,
Gonskikh Y, Yang G, Stelzer EM, Krammer TL, Micutkova L, Vogt S,
Kreil DP, et al: Loss of the ribosomal RNA methyltransferase NSUN5
impairs global protein synthesis and normal growth. Nucleic Acids
Res. 47:11807–11825. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhang XW, Wu LY, Liu HR, Huang Y, Qi Q,
Zhong R, Zhu L, Gao CF, Zhou L, Yu J and Wu HG: NSUN5 promotes
progression and predicts poor prognosis in hepatocellular
carcinoma. Oncol Lett. 24:4392022. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Wang JY and Lu AQ: The biological function
of m6A reader YTHDF2 and its role in human disease. Cancer Cell
Int. 21:1092021. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Nguyen TT, Ma LN, Slovak ML, Bangs CD,
Cherry AM and Arber DA: Identification of novel Runx1 (AML1)
translocation partner genes SH3D19, YTHDf2, and ZNF687 in acute
myeloid leukemia. Genes Chromosomes Cancer. 45:918–932. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhang J, Pi J, Liu Y, Yu J and Feng T:
Knockdown of YTH N6-methyladenosine RNA binding protein
2 (YTHDF2) inhibits proliferation and promotes apoptosis in MGC-803
gastric cancer cells. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi.
33:1628–1634. 2017.(In Chinese). PubMed/NCBI
|
|
113
|
Chen M, Wei L, Law CT, Tsang FH, Shen J,
Cheng CL, Tsang LH, Ho DW, Chiu DK, Lee JM, et al: RNA
N6-methyladenosine methyltransferase-like 3 promotes liver cancer
progression through YTHDF2-dependent posttranscriptional silencing
of SOCS2. Hepatology. 67:2254–2270. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Xie H, Li J, Ying Y, Yan H, Jin K, Ma X,
He L, Xu X, Liu B, Wang X, et al: METTL3/YTHDF2 m6 A
axis promotes tumorigenesis by degrading SETD7 and KLF4 mRNAs in
bladder cancer. J Cell Mol Med. 24:4092–4104. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Yang J, Han Q, Li C, Yang H, Chen X and
Wang X: Circular RNA circ_0001105 Inhibits Progression and
Metastasis of Osteosarcoma by Sponging miR-766 and Activating
YTHDF2 Expression. Onco Targets Ther. 13:1723–1736. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Carissimi C, Laudadio I, Lorefice E,
Azzalin G, De Paolis V and Fulci V: Bisulphite miRNA-seq reveals
widespread CpG and non-CpG 5-(hydroxy)methyl-Cytosine in human
microRNAs. RNA Biol. 18:2226–2235. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Xiong Q and Zhang Y: Small RNA
modifications: Regulatory molecules and potential applications. J
Hematol Oncol. 16:642023. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Cheray M, Etcheverry A, Jacques C, Pacaud
R, Bougras-Cartron G, Aubry M, Denoual F, Peterlongo P, Nadaradjane
A, Briand J, et al: Cytosine methylation of mature microRNAs
inhibits their functions and is associated with poor prognosis in
glioblastoma multiforme. Mol Cancer. 19:362020. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Tang F, Liu Y, Sun Y, Xiong Y, Gu Y, Zhou
J, Ouyang Y and Zhang S: Construction of a serum diagnostic
signature based on m5C-related miRNAs for cancer detection. Front
Endocrinol (Lausanne). 14:10997032023. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Chen Q, Zhang X, Shi J, Yan M and Zhou T:
Origins and evolving functionalities of tRNA-derived small RNAs.
Trends Biochem Sci. 46:790–804. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Lyons SM, Achorn C, Kedersha NL, Anderson
PJ and Ivanov P: YB-1 regulates tiRNA-induced Stress Granule
formation but not translational repression. Nucleic Acids Res.
44:6949–6960. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Goodarzi H, Liu X, Nguyen HC, Zhang S,
Fish L and Tavazoie SF: Endogenous tRna-derived fragments suppress
breast cancer progression via YBX1 displacement. Cell. 161:790–802.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Di Fazio A and Gullerova M: An old friend
with a new face: TRNA-derived small RNAs with big regulatory
potential in cancer biology. Br J Cancer. 128:1625–1635.
2023.PubMed/NCBI
|
|
124
|
Kapranov P, Cheng J, Dike S, Nix DA,
Duttagupta R, Willingham AT, Stadler PF, Hertel J, Hackermüller J,
Hofacker IL, et al: RNA maps reveal new RNA classes and a possible
function for pervasive transcription. Science. 316:1484–1488. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Ma L, Cao J, Liu L, Du Q, Li Z, Zou D,
Bajic VB and Zhang Z: LncBook: A curated knowledgebase of human
long non-coding RNAs. Nucleic Acids Res. 47:D128–D134. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Statello L, Guo CJ, Chen LL and Huarte M:
Gene regulation by long non-coding RNAs and its biological
functions. Nat Rev Mol Cell Biol. 22:96–118. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Li Y, Xia Y, Jiang T, Chen Z, Shen Y, Lin
J, Xie L, Gu C, Lv J, Lu C, et al: Long noncoding RNA DIAPH2-AS1
promotes neural invasion of gastric cancer via stabilizing NSUN2 to
enhance the m5C modification of NTN1. Cell Death Dis. 14:2602023.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Zheng H, Zhu M, Li W, Zhou Z and Wan X:
m5 C and m6 A modification of long noncoding
NKILA accelerates cholangiocarcinoma progression via the
miR-582-3p-YAP1 axis. Liver Int. 42:1144–1157. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Valinluck V, Tsai HH, Rogstad DK, Burdzy
A, Bird A and Sowers LC: Oxidative damage to methyl-CpG sequences
inhibits the binding of the methyl-CpG binding domain (MBD) of
methyl-CpG binding protein 2 (MeCP2). Nucleic Acids Res.
32:4100–4108. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zhang C, Gao J, Xiong D and Zhao Y:
Transcriptome-Wide 5-methylcytosine profiling of lncRNAs in the
mouse cerebral ischemia model. Pharmaceuticals (Basel). 17:3842024.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Cusenza VY, Tameni A, Neri A and Frazzi R:
The lncRNA epigenetics: The significance of m6A and m5C lncRNA
modifications in cancer. Front Oncol. 13:10636362023. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Xu F, Xiao Q, Du WW, Wang S and Yang BB:
CircRNA: Functions, applications and prospects. Biomolecules.
14:15032024. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Chen S, Cai D, Zhao Q, Wu J, Zhou X, Xu H,
Li X, Zhang R, Peng W, Li G and Nan A: NSUN2-mediated m5C
modification of circFAM190B promotes lung cancer progression by
inhibiting cellular autophagy. Int J Biol Macromol. 306:1415282025.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Wu J, Zhao Q, Chen S, Xu H, Zhang R, Cai
D, Gao Y, Peng W, Chen X, Yuan S, et al: NSUN4-mediated m5C
modification of circERI3 promotes lung cancer development by
altering mitochondrial energy metabolism. Cancer Lett.
605:2172662024. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Cai D, Chen X, Xu H, Zhao Q, Zhou X, Wu J,
Yuan S, Gao Y, Li D, Zhang R, et al: m5C-modified circRREB1
promotes lung cancer progression by inducing mitophagy. J Exp Clin
Cancer Res. 44:2032025. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Li M, Li J, Wen H, Li J, Wang S, Guo J,
Zhang D, Zhang A, Cui C, Fu R, et al: NSUN2-mediated m5C
hypermethylation of hsa_circ_0004516 promotes breast cancer brain
metastasis by activating AKT signaling. Noncoding RNA Res.
14:177–190. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Wu L, Gu L, Zheng Y, Liu J, Wei Z, Liu F,
Li J, Meng L, Sang Y, Sang M, et al: CircPRKCA facilitates
esophageal squamous cell carcinoma metastasis via m5C-dependent
CSF2 mRNA stabilization. J Transl Med. 23:3852025. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Diamantopoulos PT, Pappa V, Symeonidis A,
Kotsianidis I, Galanopoulos A, Papadaki H, Anagnostopoulos A,
Vassilopoulos G, Zikos P, Hatzimichael E, et al: Characteristics of
Long-term survival in patients with myelodysplastic syndrome
treated with 5-Azacyditine: Results from the hellenic 5-Azacytidine
registry. Clin Lymphoma Myeloma Leuk. 20:114–121. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Song H, Zhang J, Liu B, Xu J, Cai B, Yang
H, Straube J, Yu X and Ma T: Biological roles of RNA m5C
modification and its implications in cancer immunotherapy. Biomark
Res. 10:152022. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Chen J, Ying C, Gu Z, Zhu B, Wang J, Qian
Y, Zheng H, Zhang J and Wang Y: Construction of a prognostic model
using RNA processing factor genes and the key role of NSUN6 in
glioma outcomes. J Cell Mol Med. 29:e706682025. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Wang W, Ding Y, Zhao H, Wang S, Huang J
and Sun L: NSUN2-tRNA(Val-CAC)-axis-regulated codon-biased
translation drives triple-negative breast cancer glycolysis and
progression. Cell Mol Biol Lett. 30:1002025. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Fang L, Huang H, Lv J, Chen Z, Lu C, Jiang
T, Xu P, Li Y, Wang S, Li B, et al: m5C-methylated lncRNA NR_033928
promotes gastric cancer proliferation by stabilizing GLS mRNA to
promote glutamine metabolism reprogramming. Cell Death Dis.
14:5202023. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Sun Z, Xue S, Zhang M, Xu H, Hu X, Chen S,
Liu Y, Guo M and Cui H: Aberrant NSUN2-mediated m5C
modification of H19 lncRNA is associated with poor differentiation
of hepatocellular carcinoma. Oncogene. 39:6906–6919. 2020.
View Article : Google Scholar : PubMed/NCBI
|