Pruritus is a warning abnormal sensation elicited by
stimuli that frequently results in intense scratching impulses that
markedly impair the sleep, mood and quality of life of patients
(1), as well as potentially
causing skin damage, secondary infections or even life-threatening
complications (2–4), making it a key public health issue.
Pruritus has a complex etiology, with clinical classification
typically divided into four categories (5–7): i)
Cutaneous (derived from skin diseases such as allergic,
inflammatory or infectious conditions, as well as insect bites);
ii) systemic (extracutaneous diseases such as hepatic, renal,
hematological or drug-related disorders); iii) neuropathic
(secondary to neurological diseases); and iv) psychogenic
(conditions directly triggered by psychosocial factors are closely
associated with neuropsychiatric disorders, such as anxiety
disorders, depressive disorders and somatic symptom disorders).
These categories frequently coexist, resulting in mixed pruritus,
which is more common [in the etiological classification of chronic
pruritus, mixed etiology (e.g., coexistence of inflammatory and
neuropathic factors) is a critical consideration] in clinical
practice (8). For example, elderly
individuals are more likely to develop mixed pruritus due to
factors such as decreased sebum secretion and natural degradation
of their skin barrier (9), as well
as multiple chronic comorbidities (systemic) and neurodegenerative
changes (neuropathic), making them a particularly high-risk group.
Furthermore, environmental factors such as pollution weaken the
skin barrier, raising the global risk of pruritus-related disorders
in ~3 billion individuals (10),
highlighting the complexity and urgency of prevention and
therapy.
A deeper understanding of pruritus processes is
therefore necessary for developing novel therapies. According to a
previous study (11), pruritogens
stimulate sensory nerve terminals through damaged epidermal
barriers or immune cell activation, stimulating a number of
receptors [examples include G protein-coupled receptors (GPCRs)
such as protease-activated receptors 1/2 (PAR-1/2), Mas-related G
protein-coupled receptor (MRGPR) subtypes (e.g., MRGPRD, MRGPRX1,
MRGPRX2), serotonin (5-HT) receptors including 5-HTR2B, and
cytokine receptors such as interleukin-31 receptor A (IL-31RA)
mediating IL-31 signaling, and IL-17 receptor A (IL-17RA) binding
IL-17A] and changing transient receptor potential (TRP) as well as
voltage-gated sodium (NaV) channels to generate action potentials.
These signals are subsequently transmitted to regions in the brain
such as the parabrachial nucleus (PBN) and amygdala through spinal
gastrin-releasing peptide receptor (GRPR) neurons, whereby
affective and motivational states are modulated, ultimately
resulting in the ‘pruritus-scratch’ cycle.
Currently, Western medicine treats symptoms
primarily with antihistamines, glucocorticoids and
immunomodulators, which exhibit short-term efficacy but are
associated with long-term risks, such as skin atrophy (12), pigmentation, increased infection
susceptibility (13) and relapse
after discontinuation, limiting long-term relief (14). Traditional Chinese medicine (TCM)
has been used in the treatment of pruritus for a long time, with
emphasis on syndrome differentiation and general regulation. TCM
therapy has shown a unique ability to alleviate symptoms, reduce
recurrence and improve quality of life (15) by integrating synergistic
multi-component, multi-target and multi-pathway effects of single
herbal active components and compound formulations (16). However, despite its promising
efficacy and safety, the precise mechanisms of TCM treatment and
the complex interactions between its components are unclear
(15,16), hindering the development and
practical application of novel TCM therapies. To address this, in
the present review, a novel mechanistic framework for pruritus that
includes the sequential phases of
‘initiation-transduction-amplification-central integration’ was
developed, methodically clarifying the pathogenic underpinnings.
Furthermore, the present review describes TCM intervention
strategies at the molecular level, demonstrating that active
compounds from single herbs and conventional compound formulations
work together to modulate pruritus signaling networks through
multi-target interactions. In addition, the present review explores
multidimensional and multilevel action pathways, as well as
advancements from single herbs to compound formulations,
effectively bridging the gap between the theoretical framework of
TCM and the modern pathophysiological mechanisms of pruritus,
unlike existing studies that focus on monolithic Western medical
perspectives (17) and related
guidelines emphasizing clinical workflows over mechanistic
explanations (18). The aim was
not only to deepen the systemic understanding of pruritus
pathogenesis but also to provide a robust theoretical foundation
for novel drug development, clinical precision translation and
innovative integrative TCM-Western medicine therapeutic
strategies.
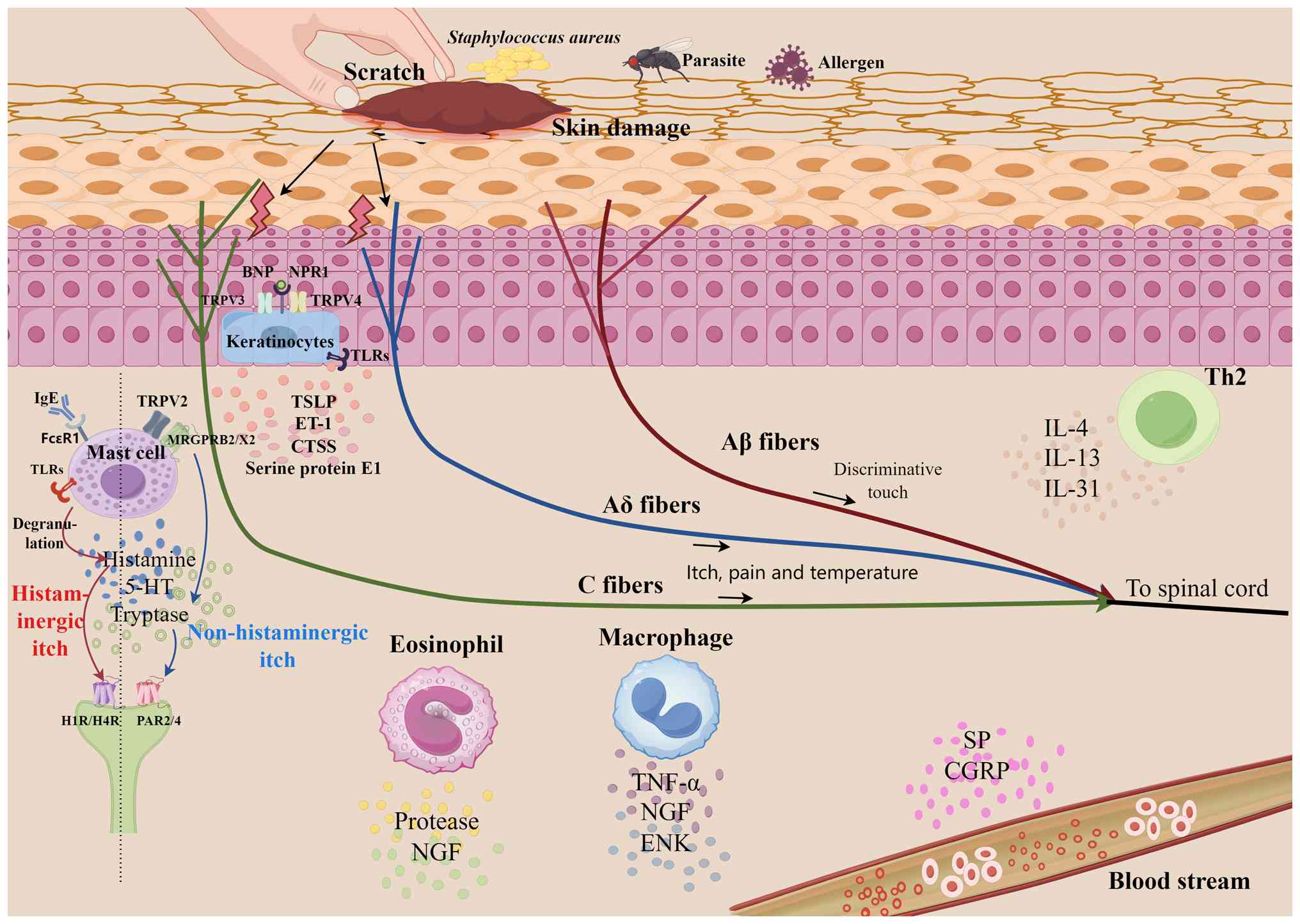
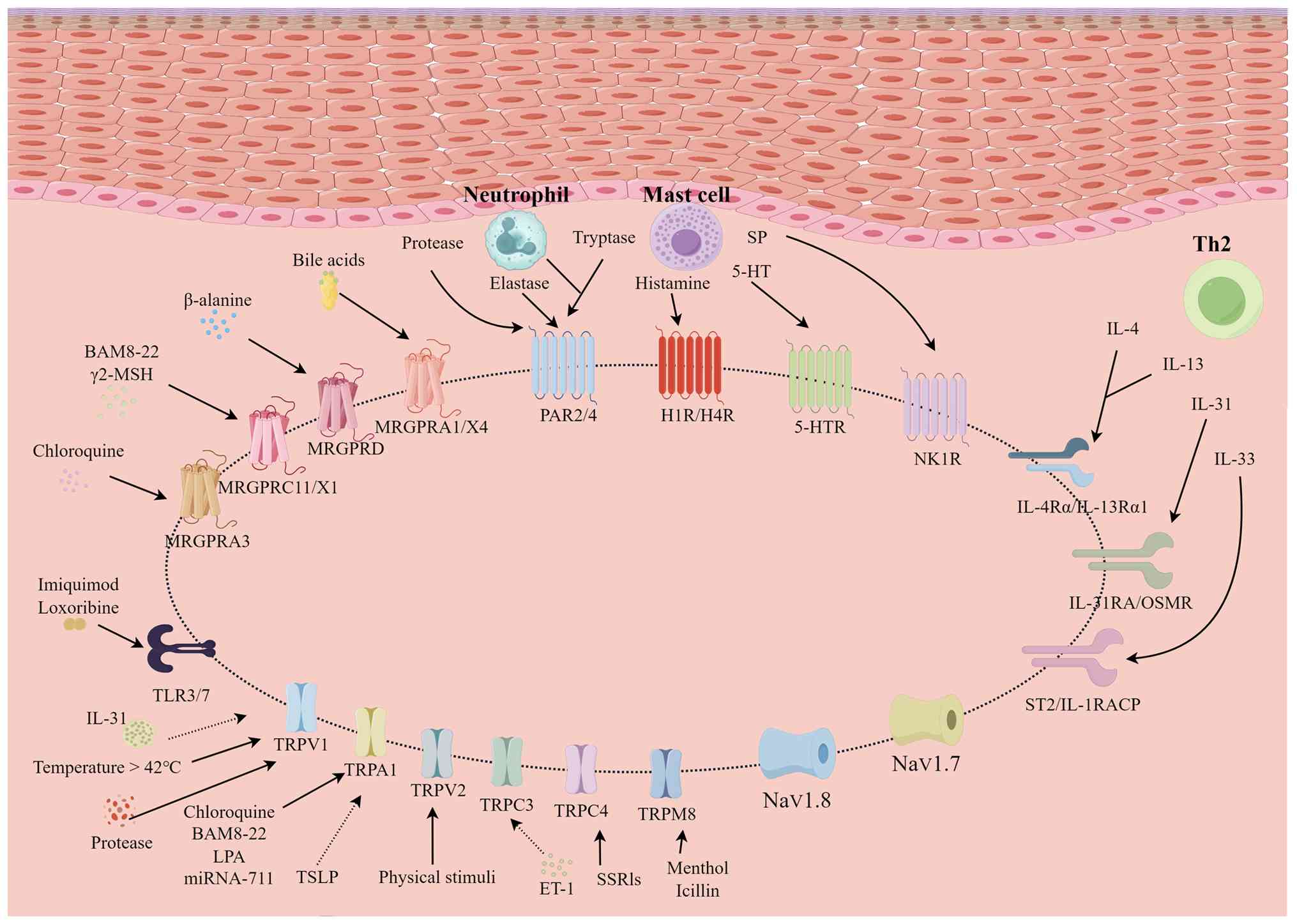
MRGPRs are primary regulators of
non-histamine-dependent pruritus, with pruritogens activating a
number of pathways [Gq protein-coupled signaling cascades (24), Gi protein signaling pathways
(25), calcium mobilization and
calcium-dependent pathways (26,27)
and TRP ion channel signaling routes (28)] (Fig.
3). The rodent MRGPRA/B/C and primate MRGPRX subfamilies are
particularly important in this area of pruritus research (29).
Chloroquine (CQ) treatment in patients with malaria
in Africa frequently causes intense pruritus (30). A study showed that mice lacking a
cluster of 12 MRGPR genes exhibit reduced scratching after CQ
administration, indicating that MRGPR-expressing neurons contribute
to itch perception (31). CQ
directly activates MRGPRA3, and ablation of MRGPRA3-expressing
neurons [which constitute 5–8% of dorsal root ganglion (DRG)
neurons] alleviates both acute and chronic itch symptoms (31). These findings have established
MRGPRA3 as a specific marker for neuronal pentraxin 2-positive
pruriceptive neurons.
Bovine adrenal medulla (BAM)-8-22 and γ2-melanocyte
stimulation hormone can activate human MRGPRX1 and mouse MRGPRC11,
which are orthologous receptors (32,33).
BAM8-22 is a strong agonist for both. Furthermore, activation of
MRGPRC11/X1 elicits varying effects depending on the site of
action. Namely, subcutaneous activation of cutaneous sensory fibers
causes itching (31), whereas
intrathecal activation causes analgesia (34,35).
As a result, an improved understanding of the functional roles of
MRGPRC11/X1 is required to create targeted therapeutic strategies
for pain- and itch-related diseases.
MRGPRB2 and its human ortholog MRGPRX2 are
specifically expressed in connective tissue mast cells and may be
activated by exogenous drugs or endogenous neuropeptides such as
substance P (SP) (42,43). Activation of MRGPRB2 primarily
triggers mast cell release of tryptase, which in turn drives
non-histaminergic itch pathways (44–46).
MRGPRX2 expression is upregulated in patients with AD, psoriasis,
allergic contact dermatitis and chronic urticaria (47). In disease models, MRGPRB2-deficient
mice exhibited markedly reduced itch and inflammation symptoms
(48,49), highlighting MRGPRX2 as a promising
therapeutic target for allergic pruritus.
Elevated bile acid levels in the serum of patients
with cholestatic disorders are associated with non-histaminergic
pruritus (50). Bile acids can
directly activate MRGPRX4 at physiological concentrations (51). Genetic deletion of its murine
ortholog, MRGPRA1, markedly reduces scratching behavior in mice
(52), whereas transgenic mice
expressing human MRGPRX4 exhibit exacerbated scratching in both
acute and chronic cholestasis models (53). These findings indicate that MRGPRX4
is a potential therapeutic target for alleviating
cholestasis-associated itch.
PARs act as a regulatory center, combining protease
signaling and epidermal barrier failure. Kallikreins and mast
cell-derived tryptase activate PAR2 and 4, resulting in pruritus.
Mast cell tryptase stimulates PAR2 through the phospholipase
C-β/inositol trisphosphate pathway, leading to IL-31 release and
keratinocyte production of thymic stromal lymphopoietin (TSLP).
This signaling cascade decreases filaggrin production through the
ERK/JNK/MAPK pathway. PAR2 activation further causes mast cell
degranulation, which releases cytokines that directly activate
nerve terminals, resulting in a self-amplifying pruritic cycle
(44,54,55).
In an animal study, administration of the PAR2 inhibitor PA-235 has
been shown to markedly reduce pruritic behaviors. Single-cell
sequencing results continue to show that PAR2 expression levels in
patients with AD lesional skin are positively associated with
‘SCORing Atopic Dermatitis’ scores (56,57).
Furthermore, Staphylococcus aureus serine protease V8
directly activates PAR1 on sensory neurons, causing bacterial
infection-induced itching (58).
Upon TLR detection of molecular patterns associated
with infection or damage, inflammatory signaling through the
myeloid differentiation primary response 88 (MyD88)/IL-1
receptor-associated kinase (IRAK)/TNF receptor-associated factor 6
(TRAF6) axis is activated. This activates the IKK complex,
transports NF-κB to the nucleus and promotes pro-inflammatory gene
transcription (59,60). Activation of the MAPK (ERK/p38/JNK)
pathway further initiates the production of cytokines (TNF-α,
IL-1/6/12, IL-8 and macrophage inflammatory protein-2) as well as
reactive oxygen/nitrogen species (61–63).
These mediators are produced by immune cells and astrocytes,
inducing local neuroinflammation and the transmission of pruritic
signals (64,65) (Fig.
3).
DRG neurons that express TLR3/TRPV1 drive both
histaminergic and non-histaminergic acute itching (66), as evidenced by agonist-induced
scratching behavior in mice. Notably, isothiocyanates (including
phenethyl isothiocyanate and sulforaphane) markedly reduce
poly(I:C)/CQ-induced acute itch and oxazolone-induced chronic itch
via inhibition of the TLR3 pathway (67).
TLR5, unlike other TLRs expressed in small-diameter
neurons, is predominantly expressed in medium-to-large DRG neurons
(74) corresponding to Aβ-LTMRs,
which mediate mechanical pruritus through activation of spinal
urocortin 3-positive interneurons (75).
TLR7, which is expressed in DRG neurons with TRP
ankyrin (TRPA)-1, is a key regulator of histamine-independent itch
(76–78). Ligands such as imiquimod directly
activate DRG neurons, causing scratching behavior and mediating
chronic itch through TRPA1-dependent processes (79). Let-7b, a secreted extracellular
microRNA, acts as an endogenous TLR7 ligand, contributing to
pruriceptive signaling (80). TLR2
and TLR7 on epidermal keratinocytes promote the release of
chemokine C-X-C motif ligand-1/2, IL-31, IL-33, IL-17A and TNF-α,
which contribute to chronic itching in dry skin and psoriasis
(81). Let-7b inhibits psoriatic
epidermal differentiation by downregulating IL-6 and inhibiting
ERK1/2 (82).
TRP superfamily ion channels serve as key components
in the regulatory network that coordinates the dynamic balance
between neuronal activation and epidermal barrier homeostasis in
the pathogenesis of cutaneous pruritus (83,84)
(Fig. 3). The TRP superfamily ion
channels are key downstream effectors of GPCRs and PARs (11,83).
The capsaicin receptor TRPV1 can detect high
temperatures (>42°C), low pH (<5.9) and capsaicin. It is
widely distributed throughout the skin and aids in the maintenance
of the epidermal barrier, activating sensory neurons by binding to
histamine receptors. PAR2 and PAR4 mediate non-histaminergic
pruritus and contribute to chronic neuroinflammation by sensitizing
TRPV1, thereby promoting pruritic responses (85).
Conversely, TRPA1 is a receptor for pain and itch,
triggered by cold temperatures (<17°C), menthol or
cinnamaldehyde. TRPA1 activity is regulated positively by numerous
pruritus-associated GPCRs (86).
TRPA1-knockout mice display markedly reduced acute scratching
reactions to CQ, adrenomedullin and sphingosine-1-phosphate
(87,88), as well as reduced scratching
activity in chronic dry skin-induced itch paradigms (89).
Loss-of-function mutations in TRPV3 impair PAR2
activity in keratinocytes and reduce neuronal activation (90,91).
By contrast, gain-of-function variants of TRPV3 are associated with
AD and Olmsted syndrome, with pharmacological inhibition of TRPV3
alleviating atopic itch (85,92).
Further studies have shown that IL-31 enhances TRPV3 expression in
keratinocytes through the natriuretic peptide receptor (NPR)-1
receptor in a brain natriuretic peptide (BNP)-dependent manner
(IL-31 stimulates sensory neurons to release BNP, which then
upregulates NPR1-mediated TRPV3 expression). This cascade promotes
the release of serine protease E1 and facilitates itch signaling
(93,94).
TRP melastatin (TRPM)-8 is a cold-sensitive ion
channel that operates within the temperature range of 8–28°C
(96,97). TRPM8 can be triggered by
methoxypropanediol to reduce irritation and improve skin barrier
restoration. TRPM8 optogenetic activation inhibits SP release from
neurons in the spinal dorsal horn that express GRPR, disrupting the
itch-scratch cycle (98).
TRP canonical (TRPC)-3 and TRPC4 are extensively
expressed in primary sensory neurons and have been associated with
pro-inflammatory sensitization (99). Mice that lack TRPC3/C4 exhibit
markedly reduced scratching reactions to non-histaminergic
pruritogens (99). In a contact
hypersensitivity model, TRPC3 expression and function in the
trigeminal ganglia were both increased (99). TRPC3 inhibition, either
pharmacological or genetic, reduces spontaneous scratching
(99). Additional research has
revealed that genetic deletion of TRPC4 notably reduced itching
caused by the selective serotonin reuptake inhibitor medicine
sertraline (100,101).
NaV channels control the intensity and duration of
itch signals by altering the action potential threshold (102,103): Higher NaV channel activity
increases neuronal excitability by lowering the action potential
threshold, resulting in more frequent action potentials and
enhanced pruritic signal intensity. The inactivation kinetics of
NaV channels determine the duration of action potentials: slower
inactivation prolongs action potential duration, sustaining
pruritic signals, whereas faster inactivation shortens action
potentials, leading to transient pruritic signaling. A study
employing sodium channel-specific knockout mouse models (104) have shown that NaV1.7 and NaV1.9
are primarily involved in acute pruritic signaling, while NaV1.8 is
associated with persistent itch pathogenesis. Additional research
(105) has revealed that DA-0218,
a computationally developed high-selectivity NaV1.7 inhibitor,
reduced both nociception and pruritus in animal models. This drug
exhibited broad-spectrum efficacy in inflammatory pain and
lymphoma-induced persistent pruritus, indicating that NaV1.7 may be
a suitable target for cross-disease pruritus treatment.
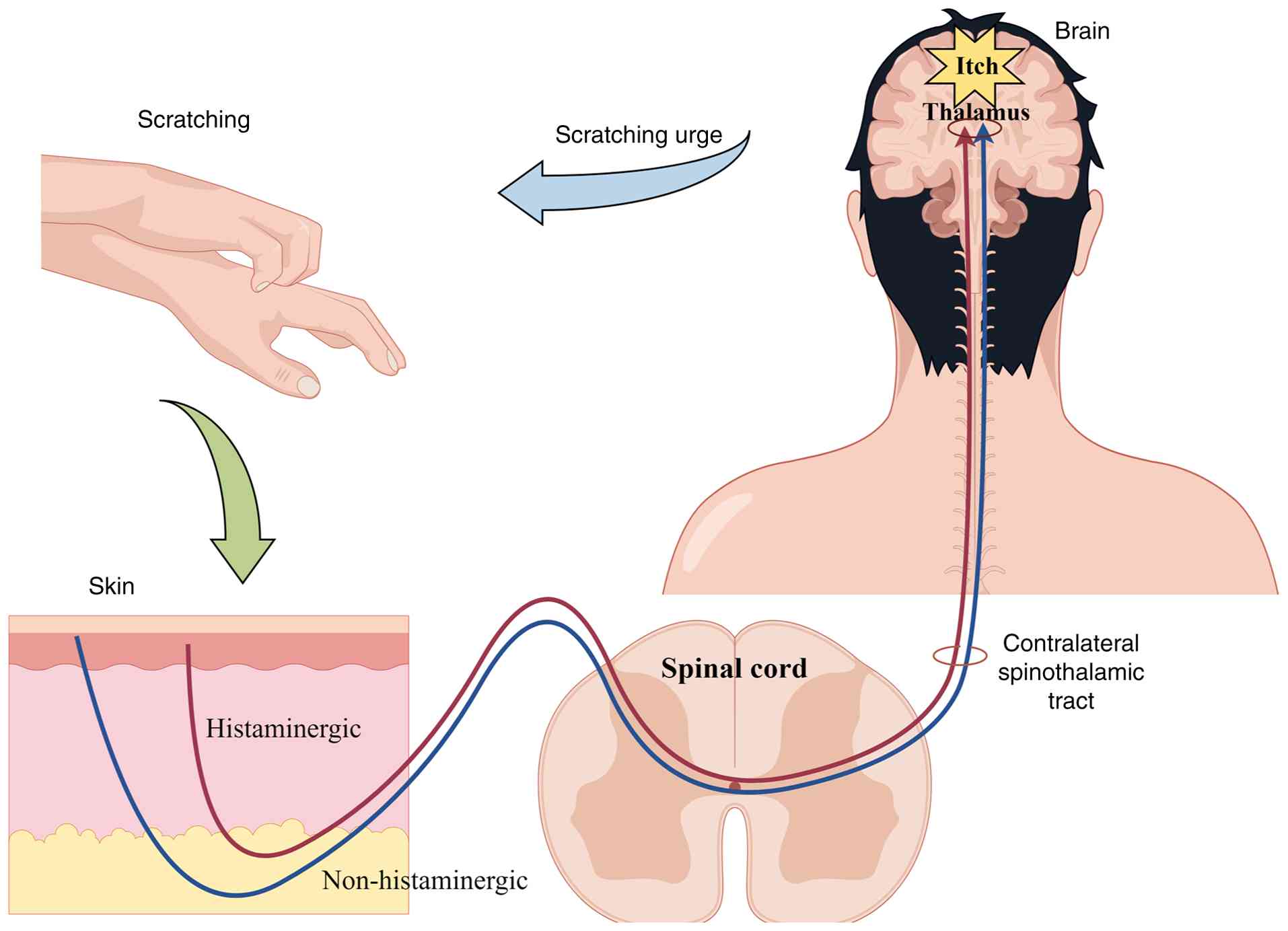
Spatiotemporal specificity of pruritic signaling is
controlled by the biogenic amine system through a network of
numerous receptor subtypes, including histamine and 5-HT (106). Histamine, a classical
pruritogenic mediator, is primarily stored in mast cells and
basophils, but keratinocytes produce trace amounts when disturbed
(107). Histamine produced
through IgE-Fc ε receptor I cross-linking or non-IgE mechanisms
(such as neuropeptides or thrombin) activates TRPV1/TRPA1 channels
through histamine receptor 1 (H1R)/H4R receptors (11), causing membrane depolarization
through the phospholipase C/12-lipoxygenase pathway. This
depolarization causes NaV1.7/NaV1.8-mediated action potentials in
the DRG, as well as neuropeptide release, resulting in neurogenic
inflammation and itching (11). In
animal models (108), combined
H1R/H4R blockade has been shown to reduce scratching more
effectively compared with single-target inhibition, indicating that
multi-receptor methods hold therapeutic promise. Furthermore, H4R
promotes Th2 cell IL-5/IL-13 secretion, which accelerates the
progression of AD (109),
suggesting that blocking H4R could be a novel therapeutic option
for AD.
5-HT regulates acute and chronic pruritus through a
number of mechanisms using unique peripheral receptor subtypes as
detailed in Table I (23,83,110–114), representing novel developments in
individualized chronic pruritus treatment: Higher NaV channel
activity increases neuronal excitability by lowering the action
potential threshold, resulting in more frequent action potentials
and enhanced pruritic signal intensity. The inactivation kinetics
of NaV channels determine the duration of action potentials: Slower
inactivation prolongs action potential duration, sustaining
pruritic signals, whereas faster inactivation shortens action
potentials, leading to transient pruritic signaling (83).
Neuropeptides serve an important role in regulating
the homeostasis of the ‘neural-immune-cutaneous’ interaction
network (Table II). SP activates
the mast cell surface receptors neurokinin 1 and MRGPRX2 (115,116), causing the production of
inflammatory mediators such as histamine and tryptase (117,118), resulting in a cycle of neurogenic
inflammation and pruritus (16).
Calcitonin gene-related peptide (CGRP), which acts as an
immunological modulator in AD, stimulates IL-13 production
(119). Conversely, fluctuations
in IL-13 levels reciprocally influence CGRP release and neuronal
sensitization (16). IL-31
activates its receptor IL-31RA to trigger CGRP secretion, while
CGRP subsequently suppresses CD4+ T cell proliferation and reduces
IL-13 generation, establishing an ‘IL-31→CGRP→IL-13’ negative
feedback axis. This axis dynamically modulates type 2 inflammatory
intensity and pruritic manifestations (120), with CGRP levels adjusting
downstream inflammatory outputs in response to upstream signals,
implying a dynamic relationship with pruritus intensity. BNP, a
pruritogenic neuropeptide, exhibits increased synthesis and release
mediated by IL-31 in the skin tissues and in vitro models of
nodular prurigo as well as the inflammatory microenvironment of AD.
BNP levels are markedly raised in the lesional skin of patients
with AD, where it activates the NPR1 receptor on keratinocytes
(121), promoting periostin
release and creating a positive feedback loop through the
‘BNP-periostin’ axis. Endothelin (ET)-1 promotes pruritus signaling
through the ET-A and ET-B receptor-mediated pathways (122).
Th2 inflammatory axes establish a pruritic signaling
amplification system by modulating the ‘neural-immune-epidermal’
interactive network (Table III;
Fig. 3). The IL-4/IL-13 axis
serves a central role in cutaneous inflammation and pruritus
(123–126). IL-31, a pro-inflammatory cytokine
secreted by Th2 cells, is a key driver of chronic pruritus when
upregulated (127–131). IL-33, a dual-function protein
from the IL-1 family, acts as a cytokine binding to the IL-1
receptor-like 1 (ST2) receptor on Th2 cells to regulate IL-17A and
IL-31 production as well as induce mast cell degranulation
(132). As a nuclear
transcriptional regulator, IL-33 interacts with the NF-κB p65
subunit in endothelial cells to facilitate inflammatory response
progression (133–136). The IL-23/IL-17 axis is primarily
implicated in the pathogenesis of psoriasis, AD and lupus
erythematosus (137–139). TSLP serves a pivotal role in
amplifying pruritic signaling in inflammatory skin diseases such as
AD by activating dendritic cells to drive Th2-type immune responses
(140–145).
The expression levels of periostin, an extracellular
matrix protein expressed by keratinocytes and fibroblasts, are
associated with the pathological course of pruritus (146–148). Concurrently, neurotrophic
factors, including nerve growth factor and brain-derived
neurotrophic factor, serve a role in pruritic and inflammatory
pathologies through distinct neuroimmunomodulatory mechanisms, with
a focus on the coordinated regulation of neuronal activation and
immune cell chemotaxis, collectively forming a molecular foundation
for neurogenic inflammation (149–153) (Table IV).
Endogenous opioids, including β-endorphin and
dynorphin A, affect pruritus through GPCRs, specifically the
µ-opioid receptor (MOR) and κ-opioid receptor (KOR). MOR is highly
expressed in inhibitory interneurons of the spinal dorsal horn,
such as NPY+ or Vgat+ neurons. β-endorphin, an endogenous MOR
agonist, inhibits the activity of these inhibitory neurons upon MOR
activation, thereby lifting their suppression on gastrin-releasing
peptide (GRP)-positive pruriceptive neurons (i.e.,
‘disinhibition’). This disinhibition hyperactivates the GRP-GRPR
microcircuit, amplifying pruritic signal transmission to the
brainstem (154,155). Specific knockout of the
Oprm1 gene in NPY or Vgat+ neurons completely abolishes
morphine-induced pruritus, confirming the central role of this
pathway (154,155). Under pathological conditions such
as atopic dermatitis, MOR activation upregulates pruritic mediators
(e.g., IL-31) and enhances TRP channel activity, intensifying
peripheral neuronal sensitization and exacerbating itch perception
(156). KOR agonists (e.g.,
nalfurafine) selectively suppress spinal efferent neurons
expressing GRPR projecting to the lateral parabrachial
nucleus/lateral spinal nucleus, blocking the transmission of
pruritic signals to higher brain centers (157). Activation of the β-endorphin-MOR
axis causes pruritus, while dynorphin A-KOR signaling reduces
symptoms, creating functionally opposing roles. An imbalance in
ligand binding to MOR and KOR in both plasma and the epidermis may
exacerbate pruritus in individuals with psoriasis, Alzheimer's
disease or liver disease (158–163).
Spinal pruriceptive signaling requires a dynamic
balance between excitatory and inhibitory circuits. GRP-expressing
neurons in the spinal cord use GRP receptors to relay peripheral
itch signals (164,165). Peripheral nociceptive inputs
inhibit the GRP-GRPR pathway by activating basic helix-loop-helix
family member E22 interneurons, which reduces chemical itch.
Somatostatin increases pruritus through binding to somatostatin 2
receptors, inhibiting dynorphinergic neurons and disinhibiting
GRPR+ neurons (166,167).
In pruritus research, the labelled line theory
posits that peripheral pruriceptors, specific receptors/ion
channels, spinal interneurons, ascending projection pathways and
cortical neurons form a dedicated ‘labelled’ pathway responsible
for itch signal recognition, transmission and perception. This
pathway diverges from other sensory modalities like pain at
critical nodes, explaining phenomena such as scratching alleviating
itch while exacerbating pain. For instance, itch and pain share
certain mediators (e.g., IL-33, TRP channels) but exhibit
modality-specific differences: IL-31/periostin predominantly drive
pruritus, whereas CCL2/CXCL dominate pain; µ-opioid receptor
activation alleviates pain but worsens itch. These observations
support the concept of partially overlapping yet functionally
segregated pathways (156). The
theory also guides targeted therapies: Dupilumab targets
Th2-associated pruritus (171),
Nalfurafine acts on the GRPR pathway (157) and gabapentin addresses
neuropathic itch (8), all
reflecting the ‘pathway-specific treatment’ paradigm.
Perceptions of pruritus involve interactions among
numerous neural circuits within the brain (172), including the primary
somatosensory cortex, thalamus, PBN, central amygdala (CeA),
periaqueductal gray and ventral tegmental area (VTA), among other
regions (Fig. 4). The PBN
constitutes a core hub for itch processing (173), where neurons are activated under
histamine and CQ stimulation. Deletion of the glutamate transporter
in this region reduces itch-related behaviors (174). The CeA integrates inputs from
numerous brain regions to modulate itch-associated affective
states. For example, histamine enhances neuronal activity in its
projection zones and optogenetic activation of CeA neurons evokes
scratching and anxiety-like behaviors (175). The VTA mediates itch-related
aversion through γ-aminobutyric acid-ergic neurons, while its
dopaminergic neurons are implicated in the rewarding effects of
scratching (176). Furthermore,
TAC1+ neurons in the lateral and ventrolateral
periaqueductal gray regulate spinal pruriceptive transmission
through the rostral ventromedial medulla pathway (176). Itch perception arises from the
coordinated activity of distributed brain networks processing
sensory (177), emotional
(178) and cognitive components
(179); however, the specific
neural circuitry mechanisms require further elucidation to guide
targeted therapies.
TCM has evolved a complete pathological
understanding of pruritus throughout millennia of clinical practice
(180,181), establishing a systematic
etiological and mechanistic framework that is consistent with
modern medical perspectives. In TCM theory, pruritus is classified
into the syndromes of itch-wind (yang-feng) or wind-induced
pruritus (180), with its core
pathogenesis scientifically interpreted as follows: External
factors such as allergen exposure, parasitic infections or
humid-hot environments disrupt the skin barrier integrity (stratum
corneum dysfunction) and trigger localized inflammation (local
inflammatory response) (182,183), leading to microcirculatory
dysregulation and heightened neuronal excitability (184). Alternatively, prolonged skin
dryness (xerosis), nutritional inadequacies or reduced epidermal
barrier function (epidermal barrier impairment) cause keratinocyte
dehydration (185), immune cell
infiltration and secondary neurogenic inflammation (186), which results in persistent
pruritus. This theoretical approach emphasizes symptom-based
phenotypic categorization for individualized diagnosis, creating
the pathophysiological groundwork for precision medicine in
contemporary dermatology. These ideas are associated with the
etiology-driven classification systems provided by the
International Forum for the Study of Itch (11,187–189), which merge classical conceptions
with modern scientific paradigms.
Clinical research on TCM for pruritic disorders
(including psoriasis, eczema, urticaria and AD) has primarily
focused on integrating disease-based diagnosis with TCM pattern
differentiation (190–193). A combined approach of internal
and external therapies has demonstrated enhanced efficacy, whereby
topical herbal formulations for eczema can precisely modulate
immune responses (190),
acupuncture or multi-herb formulas for urticaria show high response
rates (191), and integrative
TCM-Western medicine strategies for psoriasis improve therapeutic
outcomes and support early management of comorbidities (192). In the treatment of AD, TCM
emphasizes the use of topical herbal washes or ointments (193). These formulations not only
alleviate severe itching and skin dryness but also improve skin
barrier function, thereby delaying disease recurrence and reducing
dependence on corticosteroids (190–193). These interventions not only
alleviate symptoms and reduce recurrence but also minimize adverse
effects (such as skin atrophy, pigmentation, increased infection
susceptibility and relapse after discontinuation, limiting
long-term relief) associated with conventional pharmacotherapies
(such as glucocorticoids, immunosuppressants and biological agents,
among others), highlighting the holistic advantages of TCM.
Active pharmaceutical ingredients derived from TCM
represent a key component of modern TCM research, serving not only
as the pharmacodynamic material basis of compound formulations but
also as candidates for drug development. According to data from
China's National Medical Products Administration (194), a number of TCM formulations (such
as Yinxieling granules, Xiaofeng Zhiyang granules and Fushu Zhiyang
ointment), have entered the market, demonstrating efficacy in
treating a number of pruritic disorders. Concurrently, the
International Traditional Medicine Clinical Trial Registry
(195), highlights three primary
trends in TCM anti-pruritic clinical research: i) An increased
proportion of mechanism verification studies; ii) accelerated
development of novel TCM-based drugs; and iii) a marked rise in
multicenter trials. Numerous clinical trials are currently being
conducted, including a randomized, open-label, dose-finding,
positive drug-controlled clinical trial evaluating the efficacy and
safety of Jingfang mixture (ITMCTR2025000520) for chronic
urticaria, a randomized, double-blind, placebo-controlled trial
assessing Hefu Zhiyang lotion (ITMCTR2024000637) for chronic
eczema, and a study investigating the regulation of antimicrobial
peptides and Staphylococcus aureus colonization in AD by
Guben Huashi formula (ITMCTR2025001890). These efforts reflect the
growing integration of TCM with evidence-based methodologies to
advance pruritus management.
Overall, the aforementioned five classical active
components of TCM have similar modes of action in the treatment of
pruritus. These drugs target specific molecular nodes in pruritus
pathways, such as TLR4, TSLP, TRPV1 and TRPV3, reducing downstream
inflammatory signaling cascades such as NF-κB/MAPK, JAK/STAT and
JAK1/STAT3 (215). This
multi-target regulation decreases pro-inflammatory cytokines
(including TNF-α, IL-6 and IL-1β) and modulates anti-inflammatory
mediators (including IL-10) (216), thereby inhibiting inflammatory
amplification and pruritic signal transmission. These findings
establish the molecular basis for TCM's synergistic multi-target
regulation of pruritus in inflammatory skin disorders such as
atopic dermatitis (AD) and psoriasis, demonstrating its therapeutic
advantage through a multi-component, multi-target, and
multi-pathway approach.
Studies have indicated that active constituents
from additional medicinal herbs, including Saposhnikoviae Radix
(217–219), Dictamni Cortex (28,220), Moutan Cortex (221–223), Scutellariae Radix (224–227), Sophorae flavescentis Radix
(228–230), Cicadae Periostracum (231), Cnidii Fructus (232–235), Thymi Herba (236,237), Sophorae tonkinensis Radix
(238–240), Kochiae Fructus (241–243), Gentianae Radix (244–246), Tripterygii Radix (247,248), Arnebiae Radix (249,250), Tetradium ruticarpum
(251,252) and Artemisia annua
(253,254), also exhibit notable therapeutic
value in ameliorating cutaneous pruritus (Table V).
Chinese herbal formulae, as the primary
interventional technique within the diagnostic and therapeutic
systems of TCM (255), achieve
total organism management by combining the actions of diverse
elements (256). Combinatorial
approaches used in multi-herb formulations are key in the dynamic
principles of pattern discrimination and treatment (257). Si and Zhao (258) used bibliometric methods to
systematically analyze the composition principles of
pruritus-related formulae, identify core compatibility frameworks
using frequency statistics and herb cluster analysis, and conduct a
focused quantitative investigation of the distribution patterns and
cluster associations of high-frequency drugs. The findings showed
that traditional formulae such as Longdan Xiegan Tang (gentian
liver-draining decoction), Danggui Yinzi (angelica decoction),
Xiaofeng San (wind-dispersing powder), Dihuang Yinzi
(Rehmannia decoction) and Liuwei Dihuang Tang
(six-ingredient Rehmannia decoction) exhibited marked
therapeutic benefits in the treatment of cutaneous pruritus. Their
mechanisms of action are based on a mix of holistic pattern
discrimination and modern medical technology, as well as the
multitarget regulatory qualities of chemical formulations (259). Furthermore, modern modified
formulae and refined classical prescriptions have shown clear
therapeutic value in both mechanistic research and clinical
practice (260–262) by precisely optimizing herbal
compatibility ratios, demonstrating the systemic benefits of
compound formulations in regulating complex pathological
networks.
Danggui Yinzi, originating from the Song dynasty
medical text Chong Ding Yan Shi Ji Sheng Fang (265), consists of 10 herbal ingredients:
Angelica sinensis (Danggui), Paeonia lactiflora (Bai
Shao), Ligusticum chuanxiong (Chuan Xiong), Rehmannia
glutinosa (Sheng Di Huang), Schizonepeta tenuifolia
(Jing Jie Sui), Saposhnikovia divaricata (Fang Feng),
Tribulus terrestris (Bai Ji Li), Polygonum
multiflorum (He Shou Wu), Astragalus membranaceus (Huang
Qi) and Glycyrrhiza uralensis (Gan Cao). This formulation
effectively targets the IL-33/ST2-mediated pruritic signaling
pathway (266,267), inhibiting mast cell activation
and degranulation. It also reduces the expression of mast cell
tryptase and the release of pro-inflammatory cytokines such as
TNF-α and IL-1β (266).
Furthermore, it corrects the Th1/Th2 immunological imbalance by
upregulating IFN-γ, downregulating IL-4 and IgE, as well as
decreasing the production of leukotrienes (LTs; including LTB4 and
LTC4) and their receptors [cysteinyl (Cys)-lekotriene receptor 1
(LTR1) and CysLTR2] (267). These
effects notably disrupt the IL-33-induced neuroimmune-epidermal
crosstalk network, reducing inflammatory amplification loops and
easing pathological alterations such as dermal edema, capillary
dilatation and epidermal barrier failure. The molecular
pharmacological profile of Danggui Yinzi provides a mechanistic
basis for its therapeutic efficacy in chronic pruritic skin
diseases such as urticaria, which is associated with the
IL-33-driven Th2-type pruritic pathway.
These three TCM formulations contribute to curing
Th2-type pruritus by targeting key cytokine pathways, including the
TSLP/TSLPR/IL-7Rα, IL-31/OSMRβ and IL-33/ST2 signaling axes
(133,271). This multi-target approach
disrupts the inflammatory amplification cycle in the
‘neuro-immuno-epidermal’ crosstalk network and inhibits key
signaling pathways, including the JAK/STAT, MAPK and NF-κB
signaling pathways (272,273). As a result, these formulations
demonstrate a coordinated reduction in mast cell activation,
keratinocyte dysfunction and skin barrier deterioration (16). These findings provide the
groundwork for multi-target synergistic intervention in Th2-driven
chronic pruritic dermatoses such as AD and urticaria, determining
the TCM therapeutic premise of systemic control through
multi-pathway synergy.
In addition to the aforementioned Chinese herbal
formulae, other compound prescriptions such as Guizhi Mahuang Ge
Ban Tang (274) (cinnamon and
Ephedra half-and-half decoction), Taohong Siwu Tang
(275) (peach kernel and
Carthamus four-substance decoction), Huanglian Jiedu Tang
(276,277) (Coptis toxin-resolving
decoction), Jiawei Guomin Jian (278–281) (modified allergy-relieving
decoction), Danggui Kushen Wan (282,283) (Angelica and Sophora
pill), Jiuwei Yong'an Granule (284) (nine-ingredient Yong'an granule)
and Yupingfeng San (285,286) (jade wind-barrier powder) have
also been demonstrated to alleviate cutaneous pruritus (Table VI).
The present review provides a systematic review of
the diversity and complexity of pruritus mechanisms, with a focus
on the multi-pathway inhibition of pruritic signaling by active
components of single Chinese herbs and compound formulations
through synergistic modulation of the ‘neuro-immune-cutaneous’
interaction network, specifically the following interventions: i)
Suppression of receptor activation (including PARs and MRGPRs) and
subsequent sensitization of downstream TRP channels; ii) inhibition
of cytokine-mediated Th2 immune bias (for example, IL-4, IL-13 and
IL-31) and mast cell degranulation; and iii) promotion of epidermal
barrier repair (particularly in compound formulations). These
findings provide notable mechanistic evidence to support TCM
multi-target and holistic therapeutic strategies for pruritus.
Herbal components work synergistically to regulate peripheral
signaling and central integration, making them safer compared with
single-target therapy for interrupting the
‘itch-scratch-inflammation’ cycle (287).
However, certain limitations remain. Firstly,
interactions between components within compound formulations are
not fully understood and the clinical applicability of existing
experimental models needs to be validated by larger-sample clinical
trials. Future studies should aim to explore novel
pruritus-associated signaling pathways, multi-target synergistic
therapies and combined Chinese-Western therapeutic techniques to
improve overall efficacy.
The following suggestions represent treatment
approaches combining TCM and Western medicine: Firstly, following
the TCM principles of therapeutic customization and symptom
distinction may improve clinical outcomes. Future research should
aim to build upon conventional approaches while actively
investigating novel interventions and technologies. For example,
modern biotechnology could be utilized to enhance herbal extraction
and formulation techniques, thereby creating next-generation herbal
preparations. Similarly, the integration of external TCM therapies
such as acupuncture, cupping and herbal bathing with contemporary
physical therapies should be considered. Secondly, further
collaboration between TCM and Western medical professionals is
necessary to capitalize on the advantages of the other. To maximize
holistic regulation, lower recurrence rates, increase treatment
efficacy and improve the quality of life of patients with chronic
diseases, standardized, evidence-based diagnostic and therapeutic
protocols must be established. These integrated methods seek to
ensure clinical rigor and translational relevance in current
healthcare systems by bridging the gap between established clinical
practices and evidence-based standards.
To ensure scientific rigor and the preservation of
TCM characteristics, randomized controlled trials (RCTs) combining
TCM and Western medicine should strictly adhere to randomization,
control and blinding principles (288,289) while fully integrating TCM's
syndrome differentiation framework into treatment protocols.
Specific strategies include core grouping criteria based on TCM
syndrome differentiation and using clear (290), standardized syndrome diagnostic
criteria as the primary stratification component, rather than
relying primarily on Western disease categories. Blinding
implementation requires developing placebos that are comparable in
appearance, flavor and taste to active herbal medicines, as well as
keeping outcome assessors blind to group allocation to increase
objectivity (291). To guarantee
acceptable statistical power, the sample size should be determined
using mixed-effects models for continuous variables (for example,
pruritus ratings) and χ2 tests for categorical outcomes
(292). Multicenter coordination
requires unifying inclusion/exclusion criteria, diagnostic
methodologies and treatment processes across centers, as well as
centralizing training and developing a robust quality control
system to improve sample representativeness and result
dependability (293). Outcome
metrics include clinical indicators, quality-of-life assessments
and TCM-specific syndrome scores, which may help capture treatment
outcomes properly (294). To
address common issues, standardized syndrome differentiation
training must be implemented to ensure diagnostic consistency
(295), thereby establishing
clear protocols for treatment frequency, procedural standardization
and adherence optimization (for example, rationalizing treatment
schedules and improving follow-up). These efforts seek to
standardize and modernize TCM RCTs, increasing their scientific
validity and reputation.
Emerging technologies present promising prospects
for mechanistic study and medication development. Structural
biology and multi-omics methods, such as cryo-electron microscopy
and single-cell sequencing, can identify molecular connections
between herbal components and pruritic targets. Artificial
intelligence and metabolomics can clarify the holistic effects,
compatibility principles and pharmacodynamic material basis of
compound formulations. International multicenter RCTs will
establish more evidence-based efficacy evaluation systems.
Exploring developing domains (such as skin microbiota and
epigenetics) may improve the understanding of TCM mechanisms in
controlling ‘neuro-immune-microecological’ cross-system
interactions.
These advancements present novel possibilities for
combining Chinese and Western medicine in pruritus therapy,
stressing the necessity of multi-target and systemic approaches.
TCM is well-positioned to provide more systematic and accurate
solutions for global pruritus regulation through interdisciplinary
collaboration and technological integration, as well as strong
scientific support for developing integrated diagnostic and
treatment paradigms.
The present study was supported by The Li Yongji National Master
Traditional Pharmacist Legacy Studio Initiative (grant no.
14061240013) and The Open Project of the State Key Laboratory for
Innovation and Integration of Classical Formulations and Modern
Chinese Medicine (grant no. LSLSKL20240403).
Not applicable.
HZ wrote, reviewed and edited the original draft.
HZ was also responsible for conceptualization, conducting the
investigation and contributing towards the resources used. YoL
conducted project administration, provided supervision and
contributed towards the resources used, and writing, reviewing and
editing of the manuscript. YuL conceptualized the present study,
curated the data, conducted the investigation, and wrote, reviewed
and edited the manuscript. DF, LZ and RC conceptualized the study,
and contributed towards conducting the investigation, the resources
used, and writing, reviewing and editing of the manuscript. XZ and
JD acquired the funding and contributed towards conceptualizing the
present study, conducting the investigation, the resources used,
and writing, reviewing and editing of the manuscript. Data
authentication is not applicable. All authors have read and
approved the final version of the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
During the preparation of this work, artificial
intelligence tools (Grammarly) were used to improve the readability
and language of the manuscript or to generate images, and
subsequently, the authors revised and edited the content produced
by the artificial intelligence tools as necessary, taking full
responsibility for the ultimate content of the present
manuscript.
|
1
|
Whang KA, Khanna R, Williams KA, Mahadevan
V, Semenov Y and Kwatra SG: Health-related QOL and economic burden
of chronic pruritus. J Invest Dermatol. 141:754–760.e1. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Feng J, Zhao Y, Xie Z, Zang K, Sviben S,
Hu X, Fitzpatrick JAJ, Wen L, Liu Y, Wang T, et al: Miswiring of
merkel cell and pruriceptive C fiber drives the itch-scratch cycle.
Sci Transl Med. 14:eabn48192022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wimalasena NK, Milner G, Silva R, Vuong C,
Zhang Z, Bautista DM and Woolf CJ: Dissecting the precise nature of
itch-evoked scratching. Neuron. 109:3075–3087.e2. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu J, Zhao Q, Che D, Peng B, Wang X, Wang
K, Li L and Geng S: Epidermal mechanical scratching-induced ROS
exacerbates the itch-scratch cycle via TRPA1 activation on mast
cells in atopic dermatitis. J Invest Dermatol. 145:2034–2048.e7.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kahremany S, Hofmann L, Gruzman A and
Cohen G: Advances in understanding the initial steps of
pruritoceptive itch: How the itch hits the switch. Int J Mol Sci.
21:48832020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huet F, Taieb C, Corgibet F, Brenaut E,
Richard MA and Misery L: Pruritus, pain, and depression associated
with the most common skin diseases: Data from the french study
‘objectifs peau.’. Dermatol (Basel Switz). 238:448–453. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kwatra SG, Kambala A and Dong X:
Neuropathic pruritus. J Allergy Clin Immunol. 152:36–38. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Butler DC, Berger T, Elmariah S, Kim B,
Chisolm S, Kwatra SG, Mollanazar N and Yosipovitch G: Chronic
pruritus: A review. JAMA. 331:2114–2124. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Steinhoff M, Al-Khawaga S and Buddenkotte
J: Itch in elderly patients: Origin, diagnostics, management. J
Allergy Clin Immunol. 152:42–49. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Local Burden of Disease Household Air
Pollution Collaborators, . Mapping development and health effects
of cooking with solid fuels in low-income and middle-income
countries, 2000–18: A geospatial modelling study. Lancet Glob
Health. 10:e1395–e1411. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Misery L, Pierre O, Le Gall-Ianotto C,
Lebonvallet N, Chernyshov PV, Le Garrec R and Talagas M: Basic
mechanisms of itch. J Allergy Clin Immunol. 152:11–23. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Butala S and Paller AS: Optimizing topical
management of atopic dermatitis. Ann Allergy Asthma Immunol.
128:488–504. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin J, Yu H, Huang Y, Yuan J, Wang C and
Bai B: Bacterial cellulose membrane integrating phase-transited
lysozyme nanofilm loaded with biguanides for dressing treatment of
atopic dermatitis. Int J Biol Macromol. 323:1470472025. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim Y, Park K and Kim MS: Recent advances
in polymer-based drug delivery systems for atopic dermatitis:
Enhancing therapeutic efficacy and outcomes. Mater Today Bio.
35:1025172025. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sum CH, Ching J, Zhang H, Loo S, Lo CW,
Lai MK, Cheong PK, Yu CL and Lin ZX: Integrated Chinese and western
medicine interventions for atopic dermatitis: A systematic review
and meta-analysis. Chin Med. 16:101–116. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li D, Han Y, Zhou J, Yang H, Chen J, Tey
HL and Tan TTY: Mast cell-neuron axis as a core mechanism in
chronic pruritus of atopic dermatitis: From mechanistic insights to
therapeutic targets. Front Immunol. 16:16450952025. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song J, Xian D, Yang L, Xiong X, Lai R and
Zhong J: Pruritus: Progress toward pathogenesis and treatment.
Biomed Res Int. 2018:96259362018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chinese guidelines for the management of
chronic pruritus (2018). Int J Dermatol Venereol. 3:1–7. 2020.
|
|
19
|
Sutaria N, Adawi W, Goldberg R, Roh YS,
Choi J and Kwatra SG: Itch: Pathogenesis and treatment. J Am Acad
Dermatol. 86:17–34. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Agelopoulos K, Pereira MP, Wiegmann H and
Ständer S: Cutaneous neuroimmune crosstalk in pruritus. Trends Mol
Med. 28:452–462. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shin J, Kim B and Jang S: Diospyros
lotus leaf extract and its main component, myricitrin, inhibit
both histamine-dependent and histamine-independent itching. Exp
Ther Med. 29:1212025. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guo L, Zhang Y, Fang G, Tie L, Zhuang Y,
Xue C, Liu Q, Zhang M, Zhu K, You C, et al: Ligand recognition and
G protein coupling of the human Itch receptor MRGPRX1. Nat Commun.
14:50042023. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
De Logu F, Maglie R, Titiz M, Poli G,
Landini L, Marini M, Souza Monteiro de Araujo D, De Siena G,
Montini M, Cabrini DA, et al: miRNA-203b-3p induces acute and
chronic pruritus through 5-HTR2B and TRPV4. J Invest Dermatol.
143:142–153.e10. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gan B, Yu L, Yang H, Jiao H, Pang B, Chen
Y, Wang C, Lv R, Hu H, Cao Z and Ren R: Mechanism of
agonist-induced activation of the human itch receptor MRGPRX1. PLoS
Biol. 21:e30019752023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suzuki S, Iida M, Hiroaki Y, Tanaka K,
Kawamoto A, Kato T and Oshima A: Structural insight into the
activation mechanism of MrgD with heterotrimeric gi-protein
revealed by cryo-EM. Commun Biol. 5:7072022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wolf K, Kühn H, Boehm F, Gebhardt L,
Glaudo M, Agelopoulos K, Ständer S, Ectors P, Zahn D, Riedel YK, et
al: A group of cationic amphiphilic drugs activates MRGPRX2 and
induces scratching behavior in mice. J Allergy Clin Immunol.
148:506–522.e8. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
In Kim H, Lee GB, Song DE, Sanjel B, Lee
WJ and Shim WS: FSLLRY-NH2, a protease-activated receptor 2 (PAR2)
antagonist, activates mas-related G protein-coupled receptor C11
(MrgprC11) to induce scratching behaviors in mice. Life Sci.
325:1217862023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang N, Shao H, Deng J, Yang Y, Tang Z, Wu
G and Liu Y: Dictamnine ameliorates chronic itch in DNFB-induced
atopic dermatitis mice via inhibiting MrgprA3. Biochem Pharmacol.
208:1153682023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Al Hamwi G, Riedel YK, Clemens S,
Namasivayam V, Thimm D and Müller CE: MAS-related G protein-coupled
receptors X (MRGPRX): Orphan GPCRs with potential as targets for
future drugs. Pharmacol Ther. 238:1082592022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fricke TC, Pantke S, Lüttmann B,
Echtermeyer FG, Herzog C, Eberhardt MJ and Leffler A: Molecular
mechanisms of MrgprA3-independent activation of the transient
receptor potential ion channels TRPA1 and TRPV1 by chloroquine. Br
J Pharmacol. 180:2214–2229. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu Q, Tang Z, Surdenikova L, Kim S, Patel
KN, Kim A, Ru F, Guan Y, Weng HJ, Geng Y, et al: Sensory
Neuron-Specific GPCR mrgprs are itch receptors mediating
Chloroquine-induced pruritus. Cell. 139:1353–1365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lembo PMC, Grazzini E, Groblewski T,
O'Donnell D, Roy MO, Zhang J, Hoffert C, Cao J, Schmidt R,
Pelletier M, et al: Proenkephalin a gene products activate a new
family of sensory neuron-specific GPCRs. Nat Neurosci. 5:201–209.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Heller D, Doyle JR, Raman VS, Beinborn M,
Kumar K and Kopin AS: Novel probes establish mas-related G
protein-coupled receptor X1 variants as receptors with loss or gain
of function. J Pharmacol Exp Ther. 356:276–283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guan Y, Liu Q, Tang Z, Raja SN, Anderson
DJ and Dong X: Mas-related G-protein-coupled receptors inhibit
pathological pain in mice. Proc Natl Acad Sci. 107:15933–15938.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Z, He SQ, Xu Q, Yang F, Tiwari V, Liu
Q, Tang Z, Han L, Chu YX, Wang Y, et al: Activation of MrgC
receptor inhibits N-type calcium channels in small-diameter primary
sensory neurons in mice. Pain. 155:1613–1621. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Klein A, Solinski HJ, Malewicz NM, Ieong
HF, Sypek EI, Shimada SG, Hartke TV, Wooten M, Wu G, Dong X, et al:
Pruriception and neuronal coding in nociceptor subtypes in human
and nonhuman primates. Elife. 10:e645062021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang S, Edwards TN, Chaudhri VK, Wu J,
Cohen JA, Hirai T, Rittenhouse N, Schmitz EG, Zhou PY, McNeil BD,
et al: Nonpeptidergic neurons suppress mast cells via glutamate to
maintain skin homeostasis. Cell. 184:2151–2166.e16. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang C, Liu Y, Lanier M, Yeager A, Singh
I, Gumpper RH, Krumm BE, DeLeon C, Zhang S, Boehm M, et al:
High-affinity agonists reveal recognition motifs for the MRGPRD
GPCR. Cell Rep. 43:1149422024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu Q, Sikand P, Ma C, Tang Z, Han L, Li
Z, Sun S, LaMotte RH and Dong X: Mechanisms of itch evoked by
β-alanine. J Neurosci. 32:14532–14537. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Christensen J, Vecchio S, Elberling J,
Arendt-Nielsen L and Andersen H: Assessing punctate administration
of beta-alanine as a potential human model of non-histaminergic
itch. Acta Derm Venereol. 99:222–223. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Qu L, Fan N, Ma C, Wang T, Han L, Fu K,
Wang Y, Shimada SG, Dong X and LaMotte RH: Enhanced excitability of
MRGPRA3- and MRGPRD-positive nociceptors in a model of inflammatory
itch and pain. Brain. 137:1039–1050. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Weisshaar E, Szepietowski JC, Dalgard FJ,
Garcovich S, Gieler U, Giménez-Arnau AM, Lambert J, Leslie T,
Mettang T, Misery L, et al: European S2k guideline on chronic
pruritus. Acta Derm Venereol. 99:469–506. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chompunud Na Ayudhya C, Roy S, Thapaliya M
and Ali H: Roles of a mast Cell-specific receptor MRGPRX2 in host
defense and inflammation. J Dent Res. 99:882–890. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Meixiong J, Anderson M, Limjunyawong N,
Sabbagh MF, Hu E, Mack MR, Oetjen LK, Wang F, Kim BS and Dong X:
Activation of mast-cell-expressed mas-related G-protein-coupled
receptors drives non-histaminergic itch. Immunity. 50:1163–1171.e5.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang Z, Franke K, Bal G, Li Z, Zuberbier T
and Babina M: MRGPRX2-Mediated degranulation of human skin mast
cells requires the operation of Gαi, Gαq, Ca++ Channels, ERK1/2 and
PI3K-interconnection between early and late signaling. Cells.
11:9532022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Treudler R and Simon J: Developments and
perspectives in allergology. J Dtsch Dermatol Ges. 21:399–403.
2023. View Article : Google Scholar
|
|
47
|
Chaki S, Alkanfari I, Roy S, Amponnawarat
A, Hui Y, Oskeritzian CA and Ali H: Inhibition of orai channel
function regulates mas-related G protein-coupled receptor-mediated
responses in mast cells. Front Immunol. 12:8033352022. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thapaliya M, Chompunud Na Ayudhya C,
Amponnawarat A, Roy S and Ali H: Mast cell-specific MRGPRX2: A key
modulator of neuro-immune interaction in allergic diseases. Curr
Allergy Asthma Rep. 21:32021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jia T, Che D, Zheng Y, Zhang H, Li Y, Zhou
T, Peng B, Du X, Zhu L, An J and Geng S: Mast cells initiate type 2
inflammation through tryptase released by MRGPRX2/MRGPRB2
activation in atopic dermatitis. J Invest Dermatol. 144:53–62.e2.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kremer AE, Mayo MJ, Hirschfield GM, Levy
C, Bowlus CL, Jones DE, Johnson JD, McWherter CA and Choi YJ:
Seladelpar treatment reduces IL-31 and pruritus in patients with
primary biliary cholangitis. Hepatology. 80:27–37. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yu H, Zhao T, Liu S, Wu Q, Johnson O, Wu
Z, Zhuang Z, Shi Y, Peng L, He R, et al: MRGPRX4 is a bile acid
receptor for human cholestatic itch. Elife. 8:e484312019.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Meixiong J, Vasavda C, Green D, Zheng Q,
Qi L, Kwatra SG, Hamilton JP, Snyder SH and Dong X: Identification
of a bilirubin receptor that may mediate a component of cholestatic
itch. Elife. 8:e441162019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Meixiong J, Vasavda C, Snyder SH and Dong
X: MRGPRX4 is a G protein-coupled receptor activated by bile acids
that may contribute to cholestatic pruritus. Proc Natl Acad Sci.
116:10525–10530. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Meixiong J, Basso L, Dong X and Gaudenzio
N: Nociceptor-mast cell sensory clusters as regulators of skin
homeostasis. Trends Neurosci. 43:130–132. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Thibeault PE and Ramachandran R: Role of
the helix-8 and C-terminal tail in regulating proteinase activated
receptor 2 signaling. ACS Pharmacol Transl Sci. 3:868–882. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fan M, Fan X, Lai Y, Chen J, Peng Y, Peng
Y, Xiang L and Ma Y: Protease-activated receptor 2 in inflammatory
skin disease: Current evidence and future perspectives. Front
Immunol. 15:14489522024. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Stefansson K, Brattsand M, Roosterman D,
Kempkes C, Bocheva G, Steinhoff M and Egelrud T: Activation of
proteinase-activated receptor-2 by human kallikrein-related
peptidases. J Invest Dermatol. 128:18–25. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Deng L, Costa F, Blake KJ, Choi S,
Chandrabalan A, Yousuf MS, Shiers S, Dubreuil D, Vega-Mendoza D,
Rolland C, et al: S. aureus drives itch and scratch-induced skin
damage through a V8 protease-PAR1 axis. Cell. 186:5375–5393.e25.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chai Z, Zhou Y, Yang L, Zhang Y, Hossain
S, Hajimirzaei S, Qi J, Zhang G, Wei Y and Li Z: MyD88 contributes
to TLR3-mediated NF-κB activation and cytokine production in
macrophages. Cells. 14:15072025. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Araldi D, Khomula EV, Bonet IJM, Bogen O,
Green PG and Levine JD: Role of pattern recognition receptors in
chemotherapy-induced neuropathic pain. Brain. 147:1025–1042. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen X, Zhang J, Wang Y, Hu Q, Zhao R,
Zhong L, Zhan Q and Zhao L: Structure and immunostimulatory
activity studies on two novel Flammulina velutipes polysaccharides:
Revealing potential impacts of →6)-α-D-Glc p(1→ on the
TLR-4/MyD88/NF-κB pathway. Food Funct. 15:3507–3521. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wei X, Sun W, Zhu P, Ou G, Zhang S, Li Y,
Hu J, Qu X, Zhong Y, Yu W, et al: Refined polysaccharide from
dendrobium devonianum resists H1N1 influenza viral infection in
mice by activating immunity through the TLR4/MyD88/NF-κB pathway.
Front Immunol. 13:9999452022. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Feng Z, Meng R, Li Q, Li D and Xu Q:
5-aza-2′-deoxycytidine may regulate the inflammatory response of
human odontoblast-like cells through the NF-κB pathway. Int Endod
J. 54:1105–1117. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yang H, Wang YY, Chang W, Zhai M, Du WJ,
Jiang W, Xiang YW, Qin G, Yu J, Gong Y and Han Q: Primary sensory
neuron-derived miR-let-7b underlies stress-elicited psoriasis.
Brain Behav Immun. 123:997–1010. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Liu X, Jiang J, Lin S, Ge W, Tao Q, Liu S,
Zhanmu O, Yang Y, Chai B, Zhang J, et al: From skin to spinal cord:
How IL-17a drives psoriatic chronic itch. Brain Behav Immun.
132:1062182026. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu T, Berta T, Xu ZZ, Park CK, Zhang L,
Lü N, Liu Q, Liu Y, Gao YJ, Liu YC, et al: TLR3 deficiency impairs
spinal cord synaptic transmission, central sensitization, and
pruritus in mice. J Clinl Invest. 122:2195–2207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Moriyama M, Konno M, Serizawa K, Yuzawa N,
Majima Y, Hayashi I, Suzuki T and Kainoh M: Anti-pruritic effect of
isothiocyanates: Potential involvement of toll-like receptor 3
signaling. Pharmacol Res Perspect. 10:e010382022. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Min H, Lee H, Lim H, Jang YH, Chung SJ,
Lee CJ and Lee SJ: TLR4 enhances histamine-mediated pruritus by
potentiating TRPV1 activity. Mol Brain. 7:592014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu B, Escalera J, Balakrishna S, Fan L,
Caceres AI, Robinson E, Sui A, McKay MC, McAlexander MA, Herrick CA
and Jordt SE: TRPA1 controls inflammation and pruritogen responses
in allergic contact dermatitis. FASEB J. 27:3549–3563. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Shiratori-Hayashi M, Koga K, Tozaki-Saitoh
H, Kohro Y, Toyonaga H, Yamaguchi C, Hasegawa A, Nakahara T,
Hachisuka J, Akira S, et al: STAT3-dependent reactive astrogliosis
in the spinal dorsal horn underlies chronic itch. Nat Med.
21:927–931. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Koga K, Yamagata R, Kohno K, Yamane T,
Shiratori-Hayashi M, Kohro Y, Tozaki-Saitoh H and Tsuda M:
Sensitization of spinal itch transmission neurons in a mouse model
of chronic itch requires an astrocytic factor. J Allergy Clin
Immunol. 145:183–191.e10. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Feng J, Luo J, Mack MR, Yang P, Zhang F,
Wang G, Gong X, Cai T, Mei Z, Kim BS, et al: The antimicrobial
peptide human beta-defensin 2 promotes itch through toll-like
receptor 4 signaling in mice. J Allergy Clin Immunol.
140:885–888.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kim HJ, Lee EH, Lim YH, Jeong D, Na HS and
Jung Y: Pathophysiological role of TLR4 in chronic relapsing itch
induced by subcutaneous capsaicin injection in neonatal rats.
Immune Netw. 22:e202022. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Xu ZZ, Kim YH, Bang S, Zhang Y, Berta T,
Wang F, Oh SB and Ji RR: Inhibition of mechanical allodynia in
neuropathic pain by TLR5-mediated a-fiber blockade. Nat Med.
21:1326–1331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Pan H, Fatima M, Li A, Lee H, Cai W,
Horwitz L, Hor CC, Zaher N, Cin M, Slade H, et al: Identification
of a spinal circuit for mechanical and persistent spontaneous itch.
Neuron. 103:1135–1149.e6. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chen O, Jiang C, Berta T, Powell Gray B,
Furutani K, Sullenger BA and Ji RR: MicroRNA let-7b enhances spinal
cord nociceptive synaptic transmission and induces acute and
persistent pain through neuronal and microglial signaling. Pain.
165:1824–1839. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Proskocil BJ, Wai K, Lebold KM, Norgard
MA, Michaelis KA, De La Torre U, Cook M, Marks DL, Fryer AD, Jacoby
DB and Drake MG: TLR7 is expressed by support cells, but not
sensory neurons, in ganglia. J Neuroinflammation. 18:209–222. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Liu X, Wang Y, Zeng Y, Wang D, Wen Y, Fan
L, He Y, Zhang J, Sun W, Liu Y and Tao A: Microglia-neuron
interactions promote chronic itch via the NLRP3-IL-1β-GRPR axis.
Allergy. 78:1570–1584. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Liu T, Xu ZZ, Park CK, Berta T and Ji RR:
Toll-like receptor 7 mediates pruritus. Nat Neurosci. 13:1460–1462.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Park CK, Xu ZZ, Berta T, Han Q, Chen G,
Liu XJ and Ji RR: Extracellular MicroRNAs activate nociceptor
neurons to elicit pain via TLR7 and TRPA1. Neuron. 82:47–54. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wang Z, Feng Y and Hu Q: Keratinocyte TLR2
and TLR7 contribute to chronic itch through pruritic cytokines and
chemokines in mice. J Cell Physiol. 238:257–273. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wu Y, Liu L, Bian C, Diao Q, Nisar MF,
Jiang X, Bartsch JW, Zhong M, Hu X and Zhong JL: MicroRNA let-7b
inhibits keratinocyte differentiation by targeting IL-6 mediated
ERK signaling in psoriasis. Cell Commun Signal. 16:582018.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Sanjel B, Kim BH, Song MH, Carstens E and
Shim WS: Glucosylsphingosine evokes pruritus via activation of
5-HT2A receptor and TRPV4 in sensory neurons. Br J Pharmacol.
179:2193–2207. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Xie MX, Rao JH, Tian XY, Liu JK, Li X,
Chen ZY, Cao Y, Chen AN, Shu HH and Zhang XL: ATF4 inhibits TRPV4
function and controls itch perception in rodents and nonhuman
primates. Pain. 165:1840–1859. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Shirolkar P and Mishra SK: Role of TRP ion
channels in pruritus. Neurosci Lett. 768:1363792022. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hu Z, Zhang Y, Yu W, Li J, Yao J, Zhang J,
Wang J and Wang C: Transient receptor potential ankyrin 1 (TRPA1)
modulators: Recent update and future perspective. Eur J Med Chem.
257:1153922023. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wilson SR, Gerhold KA, Bifolck-Fisher A,
Liu Q, Patel KN, Dong X and Bautista DM: TRPA1 is required for
histamine-independent, mas-related G protein-coupled
receptor-mediated itch. Nat Neurosci. 14:595–602. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Hill XRZ, Morita XT, Brem XRB and Bautista
XDM: S1PR3 mediates itch and pain via distinct TRP
channel-dependent pathways. J Neurosci. 38:7833–7843. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wilson SR, Nelson AM, Batia L, Morita T,
Estandian D, Owens DM, Lumpkin EA and Bautista DM: The ion channel
TRPA1 is required for chronic itch. J Neurosci. 33:9283–9294. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Mahmoud O, Soares GB and Yosipovitch G:
Transient receptor potential channels and itch. Int J Mol Sci.
24:4202022. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhao J, Munanairi A, Liu XY, Zhang J, Hu
L, Hu M, Bu D, Liu L, Xie Z, Kim BS, et al: PAR2 mediates itch via
TRPV3 signaling in keratinocytes. J Invest Dermatol. 140:1524–1532.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Han Y, Luo A, Kamau PM, Takomthong P, Hu
J, Boonyarat C, Luo L and Lai R: A plant-derived TRPV3 inhibitor
suppresses pain and itch. Br J Pharmacol. 178:1669–1683. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Larkin C, Chen W, Szabó IL, Shan C,
Dajnoki Z, Szegedi A, Buhl T, Fan Y, O'Neill S, Walls D, et al:
Novel insights into the TRPV3-mediated itch in atopic dermatitis. J
Allergy Clin Immunol. 147:1110–1114.e5. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Tsagareli MG, Follansbee T, Iodi Carstens
M and Carstens E: Targeting transient receptor potential (TRP)
channels, mas-related G-protein-coupled receptors (mrgprs), and
protease-activated receptors (PARs) to relieve itch.
Pharmaceuticals (Basel). 16:17072023. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Chen Y, Fang Q, Wang Z, Zhang JY, MacLeod
AS, Hall RP and Liedtke WB: Transient receptor potential vanilloid
4 ion channel functions as a pruriceptor in epidermal keratinocytes
to evoke histaminergic itch. J Biol Chem. 291:10252–10262. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Courtin AS and Mouraux A: Combining
topical agonists with the recording of event-related brain
potentials to probe the functional involvement of TRPM8, TRPA1 and
TRPV1 in heat and cold transduction in the human skin. J Pain.
23:754–771. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Paricio-Montesinos R, Schwaller F,
Udhayachandran A, Rau F, Walcher J, Evangelista R, Vriens J, Voets
T, Poulet JFA and Lewin GR: The sensory coding of warm perception.
Neuron. 106:830–841.e3. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Becker J, Ellerkmann CS, Schmelzer H,
Hermann C, Lützel K, Gudermann T, Konrad DB, Trauner D, Storch U,
Mederos Y and Schnitzler M: Optical control of TRPM8 channels with
photoswitchable menthol. Angew Chem Int Ed Engl. 64:e2024165492025.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Liu Y, Liu Y, Limjunyawong N, Narang C,
Jamaldeen H, Yu S, Patiram S, Nie H, Caterina MJ, Dong X and Qu L:
Sensory neuron-expressed TRPC3 mediates acute and chronic itch.
Pain. 164:98–110. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Morita T, McClain SP, Batia LM, Pellegrino
M, Wilson SR, Kienzler MA, Lyman K, Olsen AS, Wong JF, Stucky CL,
et al: HTR7 mediates serotonergic acute and chronic itch. Neuron.
87:124–138. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xie Z and Hu H: TRP channels as drug
targets to relieve itch. Pharmaceuticals (Basel). 11:10002018.
View Article : Google Scholar
|
|
102
|
Middleton SJ, Perini I, Themistocleous AC,
Weir GA, McCann K, Barry AM, Marshall A, Lee M, Mayo LM, Bohic M,
et al: Nav1.7 is required for normal C-low threshold
mechanoreceptor function in humans and mice. Brain. 145:3637–3653.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Fauqueux J, Chaton L, Cleuziou P,
Diependaële AS, Bach N, Gruchy N, Gerard M, Meneboo JP, Villenet C,
Figeac M, et al: Long-read sequencing of recurrent FGF12
duplications in epilepsy: Insights into structural mechanisms and
aberrant isoforms. Epilepsia. 66:5014–5032. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Kühn H, Kappes L, Wolf K, Gebhardt L,
Neurath MF, Reeh P, Fischer MJM and Kremer AE: Complementary roles
of murine NaV1.7, NaV1.8 and NaV1.9 in acute itch signalling. Sci
Rep. 10:23262020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Dong F, Shi H, Yang L, Xue H, Wei M, Zhong
YQ, Bao L and Zhang X: FGF13 is required for Histamine-induced Itch
sensation by interaction with NaV1.7. J Neurosci. 40:9589–9601.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wiebe D, Limberg MM, Gray N and Raap U:
Basophils in pruritic skin diseases. Front Immunol. 14:12131382023.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Auyeung KL and Kim BS: Emerging concepts
in neuropathic and neurogenic itch. Ann Allergy Asthma Immunol.
131:561–566. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Tang H, Hou T, Wang J and Zhou W: Research
progress of histamine H4 receptor and its antagonists. Progress
Pharmaceutical Sci. 48:929–941. 2024.(In Chinese).
|
|
109
|
Nikolouli E, Mommert S, Dawodu DM,
Schaper-Gerhardt K, Stark H, Dittrich-Breiholz O, Gutzmer R and
Werfel T: The stimulation of TH2 cells results in increased IL-5
and IL-13 production via the H4 receptor. Allergy. 79:2186–2196.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Zhao ZQ, Liu XY, Jeffry J, Karunarathne
WK, Li JL, Munanairi A, Zhou XY, Li H, Sun YG, Wan L, et al:
Descending control of itch transmission by the serotonergic system
via 5-HT1A-facilitated GRP-GRPR signaling. Neuron. 84:821–834.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Stantcheva KK, Iovino L, Dhandapani R,
Martinez C, Castaldi L, Nocchi L, Perlas E, Portulano C, Pesaresi
M, Shirlekar KS, et al: A subpopulation of itch-sensing neurons
marked by ret and somatostatin expression. EMBO Rep. 17:585–600.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Domocos D, Selescu T, Ceafalan LC, Iodi
Carstens M, Carstens E and Babes A: Role of 5-HT1A and 5-HT3
receptors in serotonergic activation of sensory neurons in relation
to itch and pain behavior in the rat. J Neurosci Res. 98:1999–2017.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Miyahara Y, Funahashi H, Haruta-Tsukamoto
A, Kogoh Y, Kanemaru-Kawazoe A, Hirano Y, Nishimori T and Ishida Y:
Differential contribution of 5-HT4, 5-HT5, and 5-HT6 receptors to
acute pruriceptive processing induced by chloroquine and histamine
in mice. Biol Pharm Bull. 46:1601–1608. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Liu BW, Li ZX, He ZG, Wang Q, Liu C, Zhang
XW, Yang H and Xiang HB: Altered expression of itch-related
mediators in the lower cervical spinal cord in mouse models of two
types of chronic itch. Int J Mol Med. 44:835–846. 2019.PubMed/NCBI
|
|
115
|
Bawazir M, Sutradhar S, Roy S and Ali H:
MRGPRX2 facilitates IgE-mediated systemic anaphylaxis in a newly
established knock-in mouse model. J Allergy Clin Immunol.
155:974–987.e1. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Bandyopadhyay M, Morelli AE, Balmert SC,
Ward NL, Erdos G, Sumpter TL, Korkmaz E, Kaplan DH, Oberbarnscheidt
MH, Tkacheva O, et al: Skin codelivery of contact sensitizers and
neurokinin-1 receptor antagonists integrated in microneedle arrays
suppresses allergic contact dermatitis. J Allergy Clin Immunol.
150:114–130. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wang Z and Babina M: MRGPRX2 signals its
importance in cutaneous mast cell biology: Does MRGPRX2 Connect
mast cells and atopic dermatitis? Exp Dermatol. 29:1104–1111. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Liu AW, Zhang YR, Chen CS, Edwards TN,
Ozyaman S, Ramcke T, McKendrick LM, Weiss ES, Gillis JE, Laughlin
CR, et al: Scratching promotes allergic inflammation and host
defense via neurogenic mast cell activation. Science.
387:eadn93902025. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Antúnez C, Torres MJ, López S,
Rodriguez-Pena R, Blanca M, Mayorga C and Santamaría-Babi LF:
Calcitonin gene-related peptide modulates interleukin-13 in
circulating cutaneous lymphocyte-associated antigen-positive T
cells in patients with atopic dermatitis. Br J Dermatol.
161:547–553. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Fassett MS, Braz JM, Castellanos CA,
Salvatierra JJ, Sadeghi M, Yu X, Schroeder AW, Caston J,
Munoz-Sandoval P, Roy S, et al: IL-31-dependent neurogenic
inflammation restrains cutaneous type 2 immune response in allergic
dermatitis. Sci Immunol. 8:eabi68872023. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Wheeler JJ, Williams N, Yu J and Mishra
SK: Brain natriuretic peptide exerts inflammation and peripheral
itch in a mouse model of atopic dermatitis. J Invest Dermatol.
144:705–707. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Wong LS, Yen YT, Lin SH and Lee CH: IL-17A
induces endothelin-1 expression through p38 pathway in prurigo
nodularis. J Invest Dermatol. 140:702–706.e2. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Ulzii D, Kido-Nakahara M, Nakahara T,
Tsuji G, Furue K, Hashimoto-Hachiya A and Furue M: Scratching
counteracts IL-13 signaling by upregulating the decoy receptor
IL-13Rα2 in keratinocytes. Int J Mol Sci. 20:33242019. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Wiegmann H, Renkhold L, Zeidler C,
Agelopoulos K and Ständer S: Interleukin profiling in atopic
dermatitis and chronic nodular prurigo. Int J Mol Sci. 25:84452024.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Guttman-Yassky E, Blauvelt A, Eichenfield
LF, Paller AS, Armstrong AW, Drew J, Gopalan R and Simpson EL:
Efficacy and safety of lebrikizumab, a high-affinity interleukin 13
inhibitor, in adults with moderate to severe atopic dermatitis: A
phase 2b randomized clinical trial. JAMA Dermatol. 156:411–420.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Hashimoto T, Yokozeki H, Karasuyama H and
Satoh T: IL-31-generating network in atopic dermatitis comprising
macrophages, basophils, thymic stromal lymphopoietin, and
periostin. J Allergy Clin Immunol. 151:737–746.e6. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Kawai R, Ichimasu N and Katagiri K: IL-4
and IL-13 are not involved in IL-31-induced itch-associated
scratching behaviour in mice. Exp Dermatol. 33:e151152024.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Girolomoni G, Maurelli M and Gisondi P:
The emerging role of the neuroimmune cytokine interleukin-31 in
chronic inflammatory skin diseases. Ital J Dermatol Venereol.
157:306–312. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Suehiro M, Numata T, Saito R, Yanagida N,
Ishikawa C, Uchida K, Kawaguchi T, Yanase Y, Ishiuji Y, McGrath J
and Tanaka A: Oncostatin M suppresses IL31RA expression in dorsal
root ganglia and interleukin-31-induced itching. Front Immunol.
14:12510312023. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Miron Y, Miller PE, Hughes C, Indersmitten
T, Lerner EA and Cevikbas F: Mechanistic insights into the
antipruritic effects of lebrikizumab, an anti-IL-13 mAb. J Allergy
Clin Immunol. 150:690–700. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Xu J, Zanvit P, Hu L, Tseng PY, Liu N,
Wang F, Liu O, Zhang D, Jin W, Guo N, et al: The cytokine TGF-β
induces interleukin-31 expression from dermal dendritic cells to
activate sensory neurons and stimulate wound itching. Immunity.
53:371–383.e5. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Imai Y: Interleukin-33 in atopic
dermatitis. J Dermatol Sci. 96:2–7. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Trier AM, Mack MR, Fredman A, Tamari M,
Ver Heul AM, Zhao Y, Guo CJ, Avraham O, Ford ZK, Oetjen LK, et al:
IL-33 signaling in sensory neurons promotes dry skin itch. J
Allergy Clin Immunol. 149:1473–1480.e6. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Bodoor K, Al-Qarqaz F, Heis LA, Alfaqih
MA, Oweis AO, Almomani R and Obeidat MA: IL-33/13 axis and IL-4/31
axis play distinct roles in inflammatory process and itch in
psoriasis and atopic dermatitis. Clin Cosmet Investig Dermatol.
13:419–424. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Kolkhir P, Pyatilova P, Ashry T, Jiao Q,
Abad-Perez AT, Altrichter S, Vera Ayala CE, Church MK, He J, Lohse
K, et al: Mast cells, cortistatin, and its receptor, MRGPRX2, are
linked to the pathogenesis of chronic prurigo. J Allergy Clin
Immunol. 149:1998–2009.e5. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Franke K, Wang Z, Zuberbier T and Babina
M: Cytokines stimulated by IL-33 in human skin mast cells:
Involvement of NF-κB and p38 at distinct levels and potent
Co-operation with FcεRI and MRGPRX2. Int J Mol Sci. 22:35802021.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Tollenaere MAX, Hebsgaard J, Ewald DA,
Lovato P, Garcet S, Li X, Pilger SD, Tiirikainen ML, Bertelsen M,
Krueger JG and Norsgaard H: Signalling of multiple interleukin
(IL)-17 family cytokines via IL-17 receptor a drives
psoriasis-related inflammatory pathways. Br J Dermatol.
185:585–594. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Liu T, Li S, Ying S, Tang S, Ding Y, Li Y,
Qiao J and Fang H: The IL-23/IL-17 pathway in inflammatory skin
diseases: From bench to bedside. Front Immunol. 11:5947352020.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Lin W, Zhou Q, Liu C, Ying M and Xu S:
Increased plasma IL-17, IL-31, and IL-33 levels in chronic
spontaneous urticaria. Sci Rep. 7:177972017. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Wang ZY, Zheng YX, Xu F, Cui YZ, Chen XY,
Chen SQ, Yan BX, Zhou Y, Zheng M and Man XY: Epidermal
keratinocyte-specific STAT3 deficiency aggravated atopic
dermatitis-like skin inflammation in mice through TSLP
upregulation. Front Immunol. 14:12731822023. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Wang SH and Zuo YG: Thymic stromal
lymphopoietin in cutaneous immune-mediated diseases. Front Immunol.
12:6985222021. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Yosipovitch G Berger T and Fassett MS:
Neuroimmune interactions in chronic itch of atopic dermatitis. J
Eur Acad Dermatol Venereol. 34:239–250. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Schaper-Gerhardt K, Rossbach K, Nikolouli
E, Werfel T, Gutzmer R and Mommert S: The role of the histamine
H4 receptor in atopic dermatitis and psoriasis. Br J
Pharmacol. 177:490–502. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Dai X, Muto J, Shiraishi K, Utsunomiya R,
Mori H, Murakami M and Sayama K: TSLP impairs epidermal barrier
integrity by stimulating the formation of nuclear
IL-33/phosphorylated STAT3 complex in human keratinocytes. J Invest
Dermatol. 142:2100–2108.e5. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Xia Q, Liu T, Wang J, Sun L, Chen X, Liu
Y, Lu X, Liu H, Sun Y and Zhang F: Mast cells and thymic stromal
lymphopoietin (TSLP) expression positively correlates with pruritus
intensity in dermatitis herpetiformis. Eur J Dermatol. 30:499–504.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Hashimoto T, Nattkemper L, Kim H,
Kursewicz CD, Fowler E, Shah SM, Nanda S, Fayne RA, Romanelli P and
Yosipovitch G: Dermal periostin: A new player in itch of prurigo
nodularis. Acta Derm Venereol. 101:adv003752021. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Hashimoto T, Kursewicz CD, Fayne RA, Nanda
S, Shah SM, Nattkemper L, Yokozeki H and Yosipovitch G:
Pathophysiologic mechanisms of itch in bullous pemphigoid. J Am
Acad Dermatol. 83:53–62. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Hashimoto T, Mishra SK, Olivry T and
Yosipovitch G: Periostin, an emerging player in itch sensation. J
Invest Dermatol. 141:2338–2343. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Sari DW, Minematsu T, Yoshida M,
Noguchi-Watanabe M, Tomida S, Kitamura A, Abe M and Sanada H:
Validity of skin Blot examination for albumin and nerve growth
factor β to detect itching of the skin in indonesian older adults.
J Tissue Viability. 30:42–50. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Wong LS, Lee CH and Yen YT: Increased
epidermal nerve growth factor without small-fiber neuropathy in
dermatomyositis. Int J Mol Sci. 23:90302022. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Deng J, Parthasarathy V, Marani M,
Bordeaux Z, Lee K, Trinh C, Cornman HL, Kambala A, Pritchard T,
Chen S, et al: Extracellular matrix and dermal nerve growth factor
dysregulation in prurigo nodularis compared to atopic dermatitis.
Front Med (Lausanne). 9:10228892022. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Solinski HJ, Rukwied R and Schmelz M:
Microinjection of pruritogens in NGF-sensitized human skin. Sci
Rep. 11:214902021. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Guseva D, Rüdrich U, Kotnik N, Gehring M,
Patsinakidis N, Agelopoulos K, Ständer S, Homey B, Kapp A, Gibbs
BF, et al: Neuronal branching of sensory neurons is associated with
BDNF-positive eosinophils in atopic dermatitis. Clin Exp Allergy.
50:577–584. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Zeng Q, Li Y, Wu Y, Wu J, Xu K, Chen Y,
Rao Y, Li N, Luo Y, Jiang C, et al: Neuropeptide Y neurons mediate
opioid-induced itch by disinhibiting GRP-GRPR microcircuits in the
spinal cord. Nat Commun. 16:70742025. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Wang Z, Jiang C, Yao H, Chen O, Rahman S,
Gu Y, Zhao J, Huh Y and Ji RR: Central opioid receptors mediate
morphine-induced itch and chronic itch via disinhibition. Brain.
144:665–681. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Yosipovitch G, Kim B, Luger T, Lerner E,
Metz M, Adiri R, Canosa JM, Cha A and Ständer S: Similarities and
differences in peripheral itch and pain pathways in atopic
dermatitis. J Allergy Clin Immunol. 153:904–912. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Sheahan TD, Warwick CA, Cui AY, Baranger
DAA, Perry VJ, Smith KM, Manalo AP, Nguyen EK, Koerber HR and Ross
SE: Kappa opioids inhibit spinal output neurons to suppress itch.
Sci Adv. 10:eadp60382024. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Kupczyk P, Hołysz M and Jagodziński PP:
Opioid receptors in psoriatic skin: Relationship with itch. Acta
Derm Venereol. 97:564–570. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Taneda K, Tominaga M, Negi O, Tengara S,
Kamo A, Ogawa H and Takamori K: Evaluation of epidermal nerve
density and opioid receptor levels in psoriatic itch. Br J
Dermatol. 165:277–284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Kim BS, Inan S, Ständer S, Sciascia T,
Szepietowski JC and Yosipovitch G: Role of kappa-opioid and
mu-opioid receptors in pruritus: Peripheral and central Itch
circuits. Exp Dermatol. 31:1900–1907. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Bigliardi-Qi M, Lipp B, Sumanovski LT,
Buechner SA and Bigliardi PL: Changes of epidermal mu-opiate
receptor expression and nerve endings in chronic atopic dermatitis.
Dermatology. 210:91–99. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Moniaga C, Iwamoto S, Kitamura T,
Fujishiro M, Takahashi N, Kina K, Ogawa H, Tominaga M and Takamori
K: Plasma dynorphin a concentration reflects the degree of pruritus
in chronic liver disease: A preliminary report. Acta Derm Venereol.
99:442–443. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Kremer AE: Endogenous opioid levels do not
correlate with itch intensity and therapeutic interventions in
hepatic pruritus. Front Med. 8:6411632021. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Sun S, Xu Q, Guo C, Guan Y, Liu Q and Dong
X: Leaky gate model: Intensity-dependent coding of pain and itch in
the spinal cord. Neuron. 93:840–853. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Lay M and Dong X: Neural mechanisms of
itch. Annu Rev Neurosci. 43:187–205. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Huang J, Polgár E, Solinski HJ, Mishra SK,
Tseng PY, Iwagaki N, Boyle KA, Dickie AC, Kriegbaum MC, Wildner H,
et al: Circuit dissection of the role of somatostatin in itch and
pain. Nat Neurosci. 21:707–716. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Ross SE, Mardinly AR, McCord AE, Zurawski
J, Cohen S, Jung C, Hu L, Mok SI, Shah A, Savner EM, et al: Loss of
inhibitory interneurons in the dorsal spinal cord and elevated itch
in Bhlhb5 mutant mice. Neuron. 65:886–898. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Acton D, Ren X, Di Costanzo S, Dalet A,
Bourane S, Bertocchi I, Eva C and Goulding M: Spinal neuropeptide
Y1 receptor-expressing neurons form an essential excitatory pathway
for mechanical itch. Cell Rep. 28:625–639.e6. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Ren X, Liu S, Virlogeux A, Kang SJ, Brusch
J, Liu Y, Dymecki SM, Han S, Goulding M and Acton D: Identification
of an essential spinoparabrachial pathway for mechanical itch.
Neuron. 111:1812–1829.e6. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Gao ZR, Chen WZ, Liu MZ, Chen XJ, Wan L,
Zhang XY, Yuan L, Lin JK, Wang M, Zhou L, et al: Tac1-expressing
neurons in the periaqueductal gray facilitate the itch-scratching
cycle via descending regulation. Neuron. 101:45–59.e9. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Wang L, Peng J and Chen J: Case report:
Dupilumab: A promising treatment option for adult linear IgA
bullous dermatosis with severe pruritus. Front Immunol.
15:14095562024. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Piyush Shah D and Barik A: The
spino-parabrachial pathway for itch. Front Neural Circuits.
16:8058312022. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Dong X and Dong X: Peripheral and central
mechanisms of itch. Neuron. 98:482–494. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Mu D, Deng J, Liu KF, Wu ZY, Shi YF, Guo
WM, Mao QQ, Liu XJ, Li H and Sun YG: A central neural circuit for
itch sensation. Science. 357:695–699. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Sanders XKM, Sakai K, Henry TD, Hashimoto
XT and Akiyama XT: A subpopulation of amygdala neurons mediates the
affective component of itch. J Neurosci. 39:3345–3356. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Su XY, Chen M, Yuan Y, Li Y, Guo SS, Luo
HQ, Huang C, Sun W, Li Y, Zhu MX, et al: Central processing of itch
in the midbrain reward center. Neuron. 102:858–872.e5. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Chen XJ, Liu YH, Xu NL and Sun YG: Itch
perception is reflected by neuronal ignition in the primary
somatosensory cortex. Natl Sci Rev. 9:nwab2182022. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Zheng J, Zhang XM, Tang W, Li Y, Wang P,
Jin J, Luo Z, Fang S, Yang S, Wei Z, et al: An insular cortical
circuit required for itch sensation and aversion. Curr Biol.
34:1453–1468.e6. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Wu GY, Zheng XX, Zhao SL, Wang Y, Jiang S,
Wang YS, Yi YL, Yao J, Wen HZ, Liu J, et al: The prelimbic cortex
regulates itch processing by controlling attentional bias.
iScience. 26:1058292023. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Zhang Z, Meng H and Li L: A preliminary
study on traditional Chinese medicine in the development of senil
antipruritic cosmetics. J Light Industry. 32:37–42. 2017.(In
Chinese).
|
|
181
|
Ning Z, Yuxin Z, Yanan Z, Xiaohui Y,
Guanru L and Fengjie Z: Systematic review and Meta-analysis of the
clinical efficacy of oral Chinese medicine on pruritus. World Chin
Med. 16:934–941. 2021.
|
|
182
|
Cristaudo A, Pigliacelli F, Sperati F,
Orsini D, Cameli N, Morrone A and Mariano M: Instrumental
evaluation of skin barrier function and clinical outcomes during
dupilumab treatment for atopic dermatitis: An observational study.
Skin Res Technol. 27:810–813. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Steinhoff M, Ahmad F, Pandey A, Datsi A,
AlHammadi A, Al-Khawaga S, Al-Malki A, Meng J, Alam M and
Buddenkotte J: Neuroimmune communication regulating pruritus in
atopic dermatitis. J Allergy Clin Immunol. 149:1875–1898. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Tamari M and Ver Heul AM: Neuroimmune
mechanisms of type 2 inflammation in the skin and lung. Allergol
Int. 74:177–186. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Tominaga M and Takamori K: Peripheral itch
sensitization in atopic dermatitis. Allergol Int. 71:265–277. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Torres T, Mendes-Bastos P, Cruz MJ, Duarte
B, Filipe P, Lopes MJP and Gonçalo M: Interleukin-4 and atopic
dermatitis: Why does it matter? A narrative review. Dermatol Ther.
15:579–597. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Biazus Soares G, Hashimoto T and
Yosipovitch G: Atopic dermatitis itch: Scratching for an
explanation. J Invest Dermatol. 144:978–988. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Garcovich S, Maurelli M, Gisondi P, Peris
K, Yosipovitch G and Girolomoni G: Pruritus as a distinctive
feature of type 2 inflammation. Vaccines. 9:303–318. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Wollenberg A and Giménez-Arnau A:
Sensitive skin: A relevant syndrome, be aware. J Eur Acad Dermatol
Venereol. 36:3–5. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Dong H, Peng M, Wang L, An Y and Miao M:
Modern mechanisms and new advances in the treatment of external use
of Chinese medicine in the treatment of chronic eczema. China J
Traditional Chin Med Pharmacy. 40:4176–4180. 2025.(In Chinese).
|
|
191
|
Yue Y and Hou S: Research progress on
pharmacological effects and clinical application of traditional
Chinese medicine in treatment of chronic urticaria. Chin
Traditional Herbal Drugs. 53:7596–7605. 2022.(In Chinese).
|
|
192
|
Liu L, Sun X, Mo M, Zhou Y, Li B, Zhang X
and Li X: Clinical Diseases Responding Specially to TCM Treatment:
Psoriasis. Chin J Exp Traditional Med Formulae. 31:260–268.
2025.(In Chinese).
|
|
193
|
Liu M, He B, Hu J, Dai Y, Ren L, Ge S, Li
K, Jin Q, Song P and Chi H: Randomized Controlled Trials on Chinese
Herbal Medicine Therapy for Atopic Dermatitis: An Evidence Map.
Chin J Exp Traditional Med Formulae. 31:138–145. 2025.(In
Chinese).
|
|
194
|
Xu Y, Guo Q, Chen Z, Liu Y and Yang Y:
Overview of new indications for novel drugs approved in China
between 2018 and 2024. Drug Discovery Today. 30:1043422025.
View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Tian S, Zhang J, Yuan S, Wang Q, Lv C,
Wang J, Fang J, Fu L, Yang J, Zu X, et al: Exploring
pharmacological active ingredients of traditional Chinese medicine
by pharmacotranscriptomic map in ITCM. Brief Bioinform.
24:bbad0272023. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Li K, Liu J, Wang T and Wu G: Rule of
Prescriptions for Treatment of Pruritus by Modern Famous Chinese
Medicine Based on Association Rule. Liaoning J Traditional Chin
Med. 50:156–160. 2023.(In Chinese).
|
|
197
|
Zhang W, Cui N, Su F, Liu M, Li B, Sun Y,
Zeng Y, Yang B, Kuang H and Wang Q: Effects of rehmanniae radix
praeparata polysaccharides on LPS-induced immune activation in mice
based on gut microbiota, metabolomics and transcriptomics. Int J
Biol Macromol. 311:1439812025. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Zhang H, Wu ZM, Yang YP, Shaukat A, Yang
J, Guo YF, Zhang T, Zhu XY, Qiu JX, Deng GZ and Shi DM: Erratum to:
Catalpol ameliorates LPS-induced endometritis by inhibiting
inflammation and TLR4/NF-κB signaling. J Zhejiang Univ Sci B.
21:341. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Lu MK, Chang CC, Chao CH and Hsu YC:
Structural changes, and anti-inflammatory, anti-cancer potential of
polysaccharides from multiple processing of Rehmannia
glutinosa. Int J Biol Macromol. 206:621–632. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Xin Q, Yuan R, Shi W, Zhu Z, Wang Y and
Cong W: A review for the anti-inflammatory effects of paeoniflorin
in inflammatory disorders. Life Sci. 237:1169252019. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Zhang L and Wei W: Anti-inflammatory and
immunoregulatory effects of paeoniflorin and total glucosides of
paeony. Pharmacol Ther. 207:1074522020. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Kong X and Dongmei S: The applications and
mechanisms of paeoniflorin in dermatological disorders. Chin J
Dermatovenerol Integrated Traditional Western Med. 17:473–476.
2018.
|
|
203
|
Chen J, He Q and Jin J: Targeting
dendritic cell activation: The therapeutic impact of paeoniflorin
in cortosteroid-dependent dermatitis management. Arch Dermatol Res.
316:348–359. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Chen L, Wang Y and Guan Z: Auxiliary
effects of Heling emollient in children with mild atopic
dermatitis. Chin J Practical Pediatr. 38:869–872. 2023.(In
Chinese).
|
|
205
|
Ning J, Deng X, Chen L, Hang L, Zhu Y, Wu
L, Xue Y and Yuan H: Construction and in vitro anti-inflammatory
evaluation of a drug-loaded one-piece licorice exocyst-like
particle-loaded licorice chalcone A nano-delivery system. Acta
Pharmaceutica Sinica. 60:1147–1155. 2025.(In Chinese).
|
|
206
|
Shu J, Cui X, Liu X, Yu W, Zhang W, Huo X
and Lu C: Licochalcone a inhibits IgE-mediated allergic reaction
through PLC/ERK/STAT3 pathway. Int J Immunopathol Pharmacol.
36:39463202211354622022. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Wang Y, Yang H, Chen L, Jafari M and Tang
J: Network-based modeling of herb combinations in traditional
Chinese medicine. Briefings Bioinf. 22:bbab1062021. View Article : Google Scholar
|
|
208
|
Gao K, Liu L, Lei S, Li Z, Huo P, Wang Z,
Dong L, Deng W, Bu D, Zeng X, et al: HERB 2.0: An updated database
integrating clinical and experimental evidence for traditional
Chinese medicine. Nucleic Acids Res. 53:D1404–D1414. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Zhang Y, Li X, Shi Y, Chen T, Xu Z, Wang
P, Yu M, Chen W, Li B, Jing Z, et al: ETCM v2.0: An update with
comprehensive resource and rich annotations for traditional Chinese
medicine. Acta Pharm Sin B. 13:2559–2571. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Li M, Ren Y, Lin Z, Liu L, Li Y, Li S, Guo
R, Li P and Du B: Structural identification and anti-stomatitis
activity of one arabinose-rich polysaccharide from Rehmannia
glutinosa. Int J Biol Macromol. 284:1380062025. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Liu Y, Dong H, Sun-Waterhouse D, Li W,
Zhang B, Yu J, Qiu Z and Zheng Z: Three anti-inflammatory
polysaccharides from Lonicera japonica thunb: Insights into
the structure-function relationships. Food Sci Hum Wellness.
13:2197–2207. 2024. View Article : Google Scholar
|
|
212
|
Bai X, Rao X, Wang Y, Shen H and Jin X: A
homogeneous Lonicera japonica polysaccharide alleviates
atopic dermatitis by promoting Nrf2 activation and NLRP3
inflammasome degradation via p62. J Ethnopharmacol. 309:1163442023.
View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Zhang T, Rao X, Song S, Tian K, Wang Y,
Wang C, Bai X and Liu P: WLJP-025p, a homogeneous Lonicera
japonica polysaccharide, attenuates atopic dermatitis by
regulating the MAPK/NFκB/AP-1 axis via Act1. Int J Biol Macromol.
256:1284352024. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Zhang H, Sun X, Qi H, Ma Q, Zhou Q, Wang W
and Wang K: Pharmacological inhibition of the Temperature-sensitive
and Ca2+-Permeable transient receptor potential vanilloid TRPV3
channel by natural forsythoside B attenuates pruritus and
cytotoxicity of keratinocytes. J Pharmacol Exp Ther. 368:21–31.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Guttman-Yassky E, Irvine AD, Brunner PM,
Kim BS, Boguniewicz M, Parmentier J, Platt AM and Kabashima K: The
role of Janus kinase signaling in the pathology of atopic
dermatitis. J Allergy Clin Immunol. 152:1394–1404. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Shi HX, Gao YJ and Wang SR:
Electroacupuncture in non-surgical management of lumbar spinal
stenosis: Mechanistic potential in attenuating ligamentum flavum
thickening via inflammatory factor modulation. Front Immunol.
16:16443942025. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Yan J, Ye F, Ju Y, Wang D, Chen J, Zhang
X, Yin Z, Wang C, Yang Y, Zhu C, et al: Cimifugin relieves pruritus
in psoriasis by inhibiting TRPV4. Cell Calcium. 97:1024292021.
View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Zheng J, Gu A, Kong L, Lu W, Xia J, Hu H
and Hong M: Cimifugin relieves histamine-independent itch in atopic
dermatitis via targeting the CQ receptor MrgprA3. ACS Omega.
9:7239–7248. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Wang X, Wang Q, Zhang F and Wang J: The
therapeutic effect of cimifugin on atopic dermatitis in mice and
its mechanism. Chin Pharmacological Bulletin. 12:1648–1654.
2023.(In Chinese).
|
|
220
|
Yang N, Shao H, Deng J, Jin H, Xu L and
Liu Y: Comparison and mechanism study on the anti-dermatitis
effects of the main pharmacodynamic components of cortex dictamni
including obacunone, dictamnine, and fraxinellone. J Nanjing Med
Univ. 43:1636–1642. 16492023.(In Chinese).
|
|
221
|
Wu D, Cao X and An Y: Syndrome
Differentiation and Treatment of Psoriasis by Regulating Classical
JAK/STAT Signaling Pathway with Traditional Chinese Medicine: A
Review. Chin J Exp Traditional Med Formulae. 29:258–269. 2023.(In
Chinese).
|
|
222
|
Yan S, Xindi Z and Hongying Z: Clinical
study on Paeonol Ointment combined with tacrolimus in treatment of
skin pruritus. Drugs Clinic. 38:1438–1441. 2023.
|
|
223
|
Hu Y, Zhou H, Huang Q and Yao Y: Effect of
paeonol on the expression of cytokines production and maturation of
DCs co-stimulated by FSL-1 and IL-4. Chin J Clin Pharmacol
Therapeutics. 24:14–19. 2019.(In Chinese).
|
|
224
|
Wang D, Sun Z, Guo Q, Li X, Bai Y, Wei L
and He Y: Therapeutic effect and mechanism of the topical
preparation of baicalein onatopic dermatitis. J China Pharm Univ.
56:99–109. 2025.(In Chinese).
|
|
225
|
Du LX, Gao XY, Ren XQ, Yang YY, Ding YY,
Xu A, Wang XY, Zhang YX, Shu S, Yang YF, et al: Baicalein
ameliorates chronic itch in ACD mice by suppressing the spinal
astrocytic STAT3-LCN2 cascade. Acta Pharmacol Sin. 46:366–379.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
226
|
Wang Y, Tan L, Jiao K, Xue C, Tang Q,
Jiang S, Ren Y, Chen H, El-Aziz TMA, Abdelazeem KNM, et al:
Scutellarein attenuates atopic dermatitis by selectively inhibiting
transient receptor potential vanilloid 3 channels. Br J Pharmacol.
179:4792–4808. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Um JY, Kim HB, Kim JC, Park JS, Lee SY,
Chung BY, Park CW and Kim HO: TRPV3 and Itch: The role of TRPV3 in
chronic pruritus according to clinical and experimental evidence.
Int J Mol Sci. 23L:149622022. View Article : Google Scholar
|
|
228
|
Lin Y, Chen XJ, He L, Yan XL, Li QR, Zhang
X, He MH, Chang S, Tu B, Long QD and Zeng Z: Systematic elucidation
of the bioactive alkaloids and potential mechanism from sophora
flavescens for the treatment of eczema via network pharmacology. J
Ethnopharmacol. 301:1157992023. View Article : Google Scholar : PubMed/NCBI
|
|
229
|
Luo Z, Zhao T, Yi M, Wang T, Zhang Z, Li
W, Lin N, Liang S, Verkhratsky A and Nie H: The exploration of the
potential mechanism of oxymatrine-mediated antipruritic effect
based on network pharmacology and weighted gene co-expression
network analysis. Front Pharmacol. 13:9466022022. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Kim M, Yuk HJ, Min Y, Kim DS and Sung YY:
Securinega suffruticosa extract alleviates atopy-like lesions in
NC/Nga mice via inhibition of the JAK1-STAT1/3 pathway. Biomed
Pharmacother. 169:1159032023. View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Park G, Kwon N, Kim MH and Yang WM: The
slough of cicadidae periostracum ameliorated lichenification by
inhibiting interleukin (IL)-22/janus kinase (JAK) 1/signal
transducer and activator of transcription (STAT) 3 Pathway in
atopic dermatitis. Food Sci Anim Resour. 43:859–876. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Neuberger A, Nadezhdin KD, Zakharian E and
Sobolevsky AI: Structural mechanism of TRPV3 channel inhibition by
the plant-derived coumarin osthole. EMBO Rep. 22:e532332021.
View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Callahan BN, Kammala AK, Syed M, Yang C,
Occhiuto CJ, Nellutla R, Chumanevich AP, Oskeritzian CA, Das R and
Subramanian H: Osthole, a natural plant derivative inhibits MRGPRX2
induced mast cell responses. Front Immunol. 11:7032020. View Article : Google Scholar : PubMed/NCBI
|
|
234
|
Yang Q, Kong L, Huang W, Mohammadtursun N,
Li X, Wang G and Wang L: Osthole attenuates ovalbumin-induced lung
inflammation via the inhibition of IL-33/ST2 signaling in asthmatic
mice. Int J Mol Med. 46:1389–1398. 2020.PubMed/NCBI
|
|
235
|
Cheng Q, Zhuo Z, Duhua C, Ziyi L, Xuesong
Y and Jianzhou Y: The role of transient receptor potential
vanilloid 3 in the pathogenesis of atopic dermatitis. Chin J
Dermatovenereol. 1–11. 2025.
|
|
236
|
Wang W, Wang H, Zhao Z, Huang X, Xiong H
and Mei Z: Thymol activates TRPM8-mediated Ca2+ influx for its
antipruritic effects and alleviates inflammatory response in
imiquimod-induced mice. Toxicol Appl Pharmacol. 407:1152472020.
View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Wang M, Luo H, Wei T, Lan C, Deng J and
Jia J: Research progress on the physiological function and
molecular mechanism of thymol. China Feed. 13:7–11. 2023.(In
Chinese).
|
|
238
|
Jiang S, Zhang H, Li Y, Zhi W, Zong S and
Liu Y: Research Progress on Pharmacological Effects of Sophocarpine
Based on TLR4-NF-κ B/MAPK-Inflammatory Axis. Chin Med J Res Prac.
37:99–102. 2023.(In Chinese).
|
|
239
|
Yu D, Wang S, Fu Y, Yang L and Deng Z:
Toxicity and Mechanism of Alkaloids in Sophorae Tonkinensis Radix
et Rhizoma: A Review. Chin J Exp Tradit Med Formulae. 28:262–271.
2022.(In Chinese).
|
|
240
|
Zeng H, Zhang Z, Zhou D, Wang R,
Verkhratsky A and Nie H: Investigation of the anti-inflammatory,
anti-pruritic, and analgesic effects of sophocarpine inhibiting TRP
channels in a mouse model of inflammatory itch and pain. J
Ethnopharmacol. 337:1188822025. View Article : Google Scholar : PubMed/NCBI
|
|
241
|
Luo M, Zeng B, Wang H, Yang Z, Peng Y,
Zhang Y and Wang C: Kochia scoparia saponin momordin ic modulates
HaCaT cell proliferation and apoptosis via the wnt/β-catenin
pathway. Evid Based Complement Alternat Med. 2021:55221642021.
View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Wang C, Jiao S, Zhou R, Huang P, Zeng B,
Yang Z and Wang J: Momordin Ic ameliorates psoriasis skin damage in
mice via the IL-23/IL-17 axis. Arch Dermatol Res. 316:4742024.
View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Lan J, Ma W and Chen L: Advances in
researches on chemical constituents and pharmacological activities
of Kochia Scoparia. J Chin Med Materials. 47:506–512. 2024.(In
Chinese).
|
|
244
|
Yu H, Gachmaa B, Yu J, Liang T, Uranghai
X, Guo G, Xu W, Wang P, Liu J, Jukov A, et al: A comprehensive and
systemic review of the Gentiana: Ethnobotany, traditional
applications, phytochemistry, pharmacology, and toxicology in the
mongolian plateau. J Ethnopharmacol. 345:1195732025. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Zhao K, Pu S, Sun L and Zhou D:
Gentiopicroside-loaded chitosan nanoparticles inhibit TNF-α-induced
proliferation and inflammatory response in HaCaT keratinocytes and
ameliorate imiquimod-induced dermatitis lesions in mice. Int J
Nanomed. 18:3781–3800. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
246
|
Qin C, Zhang S and Li Y: Study on
Processing History, Chemical Composition and Pharmacological
Effects of Longdan (Gentianae Radix et Rhizoma). Chin Arch
Traditional Chin Med. 44:134–140. 2026.(In Chinese).
|
|
247
|
Gao Y, Ma R, Weng W, Zhang H, Wang Y, Guo
R, Gu X, Yang Y, Yang F, Zhou A, et al: TRPV1 SUMOylation
suppresses itch by inhibiting TRPV1 interaction with H1 receptors.
Cell Rep. 39:1109722022. View Article : Google Scholar : PubMed/NCBI
|
|
248
|
Song Y, Zheng W and Hua Z: Effect
mechanism of Tripterygium wilfordii in treatment of psoriasis based
on network pharmacology and molecular docking. Chemistry Life.
42:1188–1200. 2022.(In Chinese).
|
|
249
|
Song Y, Ding Q, Hao Y, Cui B, Ding C and
Gao F: Pharmacological effects of shikonin and its potential in
skin repair: A review. Molecules. 28:79502023. View Article : Google Scholar : PubMed/NCBI
|
|
250
|
Mu Z, Guo J, Zhang D, Xu Y, Zhou M, Guo Y,
Hou Y, Gao X, Han X and Geng L: Therapeutic effects of shikonin on
skin diseases: A review. Am J Chin Med. 49:1871–1895. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
251
|
Liang J, Chen W, Zhou Y, Meng W, Xie M,
Weng Y, Qin L, Li J and Wu G: Inhibitory effect of evodiamine on
psoriasis lesions and itching in mice. Clin Cosmet Investig
Dermatol. 17:1527–1541. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
252
|
Hu M, Liu Q, Zang X, Hu J, Du J and Zhou
W: Effect of Rutaecarpine on Imiquimod (IMQ)-induced Psoriasis-like
Mouse Model. Chin J Dermatovenereol. 34:1366–1371. 2020.(In
Chinese).
|
|
253
|
Wei Q, Jin Q, Jin L, Yu R, Li F, Li H, Jin
D, Meng F and Jin G: Dihydroartemisinin alleviates psoriasis-like
skin inflammation in mice by inhibiting proliferation of
keratinocytes and expression of pro-inflammatory cytokines. Chin J
Immunol. 36:543–548. 2020.(In Chinese).
|
|
254
|
Yang J, Wang R, Wang R and Zhang S:
Preparation of dihydroartemisinin-loaded liposomes and their
topical efficacy in alleviating psoriasis-like lesions in mice.
Drug Evaluation Res. 48:1853–1868. 2025.(In Chinese).
|
|
255
|
Cui HR, Zhang JY, Cheng XH, Zheng JX,
Zhang Q, Zheng R, You LZ, Han DR and Shang HC: Immunometabolism at
the service of traditional chinese medicine. Pharmacol Res.
176:1060812022. View Article : Google Scholar : PubMed/NCBI
|
|
256
|
Wang X, Shi X, Xi Z, Zhang Z, Luo Z, Wang
J and Shan J: The scientific basis of synergy in traditional
chinese medicine: Physicochemical, pharmacokinetic, and
pharmacodynamic perspectives. Chin Med. 21:15–35. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
257
|
Yang X, Luo Y and Ding C: MRHAF:
Multi-relational hierarchical attention with hybrid knowledge
fusion for explainable herb recommendations. IEEE J Biomed Health
Inform. December 15–2025.(Epub ahead of print). View Article : Google Scholar
|
|
258
|
Si F and Zhao Z: Distribution of TCM
syndromes and prescription rules of skin pruritus. Traditi Chin Med
Res. 34:39–42. 2021.(In Chinese).
|
|
259
|
Liu XY, Zheng HW, Wang FZ, Atia TW, Fan B
and Wang Q: Developments in the study of chinese herbal medicine's
assessment index and action mechanism for diabetes mellitus. Anim
Models Exp Med. 7:433–443. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
260
|
Cui J, May BH, Lin W, Luo Q, Zhang AL, Guo
X, Lu C, Li Y and Xue CC: Chinese herbal medicines for
rhinosinusitis: A text-mining study with comparisons to
contemporary research and clinical guidance. BMC Complement Med
Ther. 25:1652025. View Article : Google Scholar : PubMed/NCBI
|
|
261
|
Li C, Jia W, Yang J, Cheng C and Olaleye
OE: Multi-compound and drug-combination pharmacokinetic research on
Chinese herbal medicines. Acta Pharmacol Sin. 43:3080–3095. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
262
|
Li HH, Livneh H, Huang HL, Wang YH, Lu MC,
Chen WJ and Tsai TY: Integrating Chinese herbal medicine into
conventional care was related to lower risk of sarcopenia among
rheumatid arthritis patients: A retrospective, population-based
study. J Multidiscip Healthc. 39:411–415. 2025.(In Chinese).
|
|
263
|
Wang S, Ma Y, Cui Z and Diao J: A case of
factitious disorders as Henoch-Schonlein purpura. Chin Mental
Health J. 39:411–415. 2025.(In Chinese).
|
|
264
|
Zhang M, Xue P, Zhao Y, Gao Y, Dong W, Ma
W, Wang Z and Zhang C: Protective Effect of Longdan Xiegan
Decoction on Eczema Rats Regulating H1R/TRPV1, PAR-2/TRPV1 and
P38MAPK Signal Pathway. World Sci Technol Modernization Traditi
Chin Med. 24:2331–2339. 2022.(In Chinese).
|
|
265
|
Ni Z, Cheng-Yan W, Qing-Wu S and Zi-He W:
Textual research on the classical prescription danggui decoction.
Modern Chin Med. 25:888–899. 2023.
|
|
266
|
Zhang X, Wei Q, Cai C, Pang Y, He Y, Wu S,
Guo J and Zeng J: Effect of Danggui Yinzi on Relieving Allergic
Reactions in UL Mice by Inhibiting IL-33-mediated Degranulation of
Mast Cells. Chin Arch Traditional Chin Med. 39:144–148. 268–270.
2021.(In Chinese).
|
|
267
|
Yao B, Wang J and Ran Z: Effect and
Mechanism of Danggui Yinzi on Eczema Model of Blood Deficiency and
Wind Dryness. World Sci Technol Modernization Traditi Chin Med.
24:4055–4062. 2022.
|
|
268
|
Li P, Yu Q, Nie H, Yin C and Liu B:
IL-33/ST2 signaling in pain and itch: Cellular and molecular
mechanisms and therapeutic potentials. Biomed Pharmacother.
165:1151432023. View Article : Google Scholar : PubMed/NCBI
|
|
269
|
Wu S and Lan C: Clinical study on the
efficacy and safety of liangxue xiaofeng san combined with
levocetirizine oral solution in treating chronic spontaneous
urticaria with blood-heat and wind-generation pattern in Children.
Lishizhen Med Materia Med Res. 34:2168–2171. 2023.(In Chinese).
|
|
270
|
Li'Na F, Yige W, Yi H, Ke X, Jin H,
Shijiao S and Guang Z: Study on regulation mechanism of xiaofeng
powder for damp-heat accumulation skin type epidermal barriers. J
Sichuan Traditi Chin Med. 39:43–48. 2021.
|
|
271
|
Roh YS, Choi J, Sutaria N, Belzberg M,
Kwatra MM and Kwatra SG: IL-31 inhibition as a therapeutic approach
for the management of chronic pruritic dermatoses. Drugs.
81:895–905. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
272
|
Huang IH, Chung WH, Wu PC and Chen CB:
JAK-STAT signaling pathway in the pathogenesis of atopic
dermatitis: An updated review. Front Immunol. 13:10682602022.
View Article : Google Scholar : PubMed/NCBI
|
|
273
|
Kim SY, Han SD, Kim M, Mony TJ, Lee ES,
Kim KM, Choi SH, Hong SH, Choi JW and Park SJ: Mentha arvensis
essential oil exerts anti-inflammatory in LPS-stimulated
inflammatory responses via inhibition of ERK/NF-κB signaling
pathway and anti-atopic dermatitis-like effects in
2,4-dinitrochlorobezene-induced BALB/c mice. Antioxidants (Basel).
10:19412021. View Article : Google Scholar : PubMed/NCBI
|
|
274
|
Fu B, Ji Y, Li J, Pei M and Yang H:
Network Pharmacology-based Mechanism of Guizhi Mahuang Geban
Decoction in the Treatment of Uraemic Pruitus. New Chinese Med and
Clin Pharmacology. 32:1675–1684. 2021.(In Chinese).
|
|
275
|
Niu F, Li B, Wang S, Li Y and Zhou W:
Taohong Siwutang Exerts Anti-inflammatory Effect and Regulates
Caspase-14 and EVPL Expression in Treatment of Psoriasis. Chin J
Exp Traditi Med Formulae. 24:12–21. 2024.(In Chinese).
|
|
276
|
Zhang Z, Zhang H, Guan J and Chen Y: Study
on the Effect and Mechanism of Huanglian Jiedu Decoction in
Improving Itching Symptoms in 1-chloro-2,4-dinitrobenzene-Induced
Atopic Dermatitis Mice. Tradit Chin Drug Res Clin Pharmacol.
6:1713–1720. 2023.(In Chinese).
|
|
277
|
Li D, Xing Y and Wang B: Pharmacological
Mechanism of Huanglian Jiedutang: A Review. Chin J Exp Tradit Med
Formulae. 24:275–283. 2025.(In Chinese).
|
|
278
|
Long S, Wen C, Zhu H, Wang Y, Li H and Wu
X: Effects of modified guominjian decoction on degranulation of
mast cells in larva of urticaria model mice. J Tradit Chin Med.
60:322–326. 2019.(In Chinese).
|
|
279
|
Gao J, Chen G, Tan W, Lan J, Zhang J, Rao
G, Wang R and Zhang Y: Interventional effects of Guo Min Jian
(Anti-Allergy Decoction) on atopic dermatitis in a murine model.
Lishizhen Med Materia Med Res. 33:1806–1810. 2022.(In Chinese).
|
|
280
|
Juan G, Guifang C, Wei T, Jie Z, Gaoxiong
R, Ruirui W and Yi Z: Mechanism of guomin decoction to improve
atopic dermatitis by inhibiting mast cell degranulosis through
IgE/FcεRI pathway. Chin Arch Traditi Chin Med. 42:135–140. 283–284.
2024.
|
|
281
|
Huang J, Xu Y, Yang X, Wang J, Zhu Y and
Wu X: Jiawei guomin decoction regulates the degranulation of mast
cells in atopic dermatitis mice via the HIS/PAR-2 pathway. J
Ethnopharmacol. 321:1174852024. View Article : Google Scholar : PubMed/NCBI
|
|
282
|
Tan W, Liu Y, Deng S, Wang R, Rao G, Zhang
L and Zhang Y: Preliminary mechanism study on the antipruritic
effect of Danggui Kushen Pill. Chin J Hosp Pharmacy. 43:1786–1794.
2023.(In Chinese).
|
|
283
|
Xing C, Mei W, Lixia H, Xinhong L, Qiuying
W and Jing T: Mechanism of Danggui Kushen pill in treating eczema
based on network pharmacology. J Shenyang Pharmaceutical Univ.
38:845–854. 2021.
|
|
284
|
Gu Q, Lin J, Lu X, Chen T, Wu Y and Yang
Y: Jiu-Wei-Yong-An formula suppresses JAK1/STAT3 and MAPK signaling
alleviates atopic dermatitis-like skin lesions. J Ethnopharmacol.
295:1154282022. View Article : Google Scholar : PubMed/NCBI
|
|
285
|
Yu H, Zhang W, Rao Q and Zhou M:
Yupingfeng Granules Relieving Pruritus via Inhibiting Mast Cells
and IL-31 in Atopic Dermatitis Rats. New Drugs Clin Pharmacology
Trad Chin Med. 33:1017–1024. 2022.(In Chinese).
|
|
286
|
Wen W, Zhang H, Lin M, Yu C, Xu E and Xu
J: Effects of Yupingfeng Powder Combined with Desloratadine on
Serum Inflammatory Factors and Quality of Life in Children with
Chronic Urticaria. Med Innovation China. 19:78–81. 2022.
|
|
287
|
Wang J, Chen Y, Yang X, Huang J, Xu Y, Wei
W and Wu X: Efficacy and safety of Chinese herbal medicine in the
treatment of chronic pruritus: A systematic review and
meta-analysis of randomized controlled trials. Front Pharmacol.
13:10299492023. View Article : Google Scholar : PubMed/NCBI
|
|
288
|
Ferrannini E and Rosenstock J: Clinical
translation of cardiovascular outcome trials in type 2 diabetes: Is
there more or is there less than meets the eye? Diabetes Care.
44:641–646. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
289
|
Zhang Y and Chow SC: Mapping of subjective
measurements in traditional Chinese medicine to objective clinical
endpoints in western medicine. Biologics. 18:433–452.
2024.PubMed/NCBI
|
|
290
|
Liu Y, Xu J, Yu Z, Chen T, Wang N, Du X,
Wang P, Zhou X, Xu H and Zhang Y: Ontology characterization,
enrichment analysis, and similarity calculation-based evaluation of
disease-syndrome-formula associations by applying SoFDA. Imeta.
2:e802023. View Article : Google Scholar : PubMed/NCBI
|
|
291
|
Lin YH, Sahker E, Shinohara K, Horinouchi
N, Ito M, Lelliott M, Cipriani A, Tomlinson A, Baethge C and
Furukawa TA: Assessment of blinding in randomized controlled trials
of antidepressants for depressive disorders 2000–2020: A systematic
review and meta-analysis. EClinicalMedicine. 50:1015052022.
View Article : Google Scholar : PubMed/NCBI
|
|
292
|
Ren Y, Jia Y, Yang M, Yao M, Wang Y, Mei
F, Li Q, Li L, Li G, Huang Y, et al: Sample size calculations for
randomized controlled trials with repeatedly measured continuous
variables as primary outcomes need improvements: A cross-sectional
study. J Clin Epidemiol. 166:1112352024. View Article : Google Scholar : PubMed/NCBI
|
|
293
|
Llovera G, Langhauser F, Isla Cainzos S,
Hoppen M, Abberger H, Mohamud Yusuf A, Mencl S, Heindl S, Ricci A,
Haupeltshofer S, et al: Stroke of consistency: Streamlining
multicenter protocols for enhanced reproducibility of infarct
volumes in preclinical stroke research. Stroke. 55:2522–2527. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
294
|
Day J, Antony A, Tillett W and Coates LC:
The state of the art-psoriatic arthritis outcome assessment in
clinical trials and daily practice. Lancet Rheumatol. 4:e220–e228.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
295
|
Leung AYL, Zhang J, Chan CY, Chen X, Mao
J, Jia Z, Li X and Shen J: Validation of evidence-based
questionnaire for TCM syndrome differentiation of heart failure and
evaluation of expert consensus. Chin Med. 18:702023. View Article : Google Scholar : PubMed/NCBI
|