Introduction
Atherosclerosis (AS) is a chronic, progressive
disease primarily driven by lipid-induced inflammation,
predominantly affecting large- and medium-sized arteries. The
fundamental mechanism of AS involves a self-perpetuating
pathological cycle of dysregulated lipid metabolism and
inflammatory responses within the vascular wall that ultimately
results in plaque formation, luminal stenosis and thrombotic
events. AS, which is a leading global cause of cardiovascular
diseases, is characterized by multiple risk factors (such as
hyperlipidemia, hypertension, diabetes mellitus and smoking) and a
complex pathogenesis that involves various cell types and
biological processes (1).
Research has increasingly emphasized the role of
lipophagy, a form of cellular autophagy, in AS. Lipophagy is not
only involved in the degradation and recycling of intracellular
lipids, but it also modulates key pathological processes, including
inflammatory cell infiltration (2), foam cell formation (3), endothelial cell (EC) damage (4) and the proliferation and migration of
vascular smooth muscle cells (VSMCs) (5). Autophagy is a highly conserved
self-degradative cellular mechanism in which the dynamic formation
and maturation of double-membrane structures are central processes.
The key structures implicated in autophagy encompass phagophores,
autophagosomes (APs) and autolysosomes (ALs) (2). Lipophagy, as a pivotal subtype of
selective autophagy, is distinguished by lysosome-mediated lipid
droplet (LD) degradation and is integral to cellular lipid
homeostasis regulation. LDs are directed to lysosomes through
ubiquitination, which involves the recruitment of
lipophagy-specific factors and autophagy receptors (such as ORP8,
spartin and p62) (6–8).
Lipophagy markedly affects AS initiation and
progression. During AS pathogenesis, abnormal accumulation of LDs
(the primary organelles for intracellular neutral lipid storage) is
a hallmark of foam cell formation in macrophages and SMCs. LD
accumulation is markedly associated with disrupted lipid metabolism
and impaired lipophagy function. Lipophagy deficiency can result in
excessive intracellular lipid deposition, which promotes AS
progression (9). While
macroautophagy serves as a bulk degradation system for cytoplasmic
components, lipophagy represents a specialized, high-precision
defense mechanism against lipotoxicity. Unlike the passive storage
of neutral lipids, lipophagy actively mobilizes triglycerides (TGs)
and cholesteryl esters (CEs) from LDs for lysosomal hydrolysis.
This process is particularly critical in the lipid-rich milieu of
AS, in which lipophagic capacity determines whether a macrophage
resolves lipid burden or degenerates into a pro-inflammatory foam
cell (6,9).
Unlike previous reviews that primarily catalogue
individual lipophagic pathways, the present article used
established knowledge to propose two novel conceptual frameworks.
First, the spatiotemporal ‘double-edged sword’ model (Fig. 1), highlighting how the net effect
of lipophagy depends on the disease stage and the intensity of
autotoxic stress in different vascular cell types. Second, the
‘convergent therapeutic axis’ model (Fig. 2), which systematically categorizes
conventional agents, natural compounds and Traditional Chinese
Medicine (TCM) formulas based on their capacity to precisely target
terminal epigenetic/transcriptional nodes [such as the
SIRT-transcription factor EB (TFEB) axis] rather than merely
inhibit broad upstream kinases. By bridging microscopic molecular
signaling principles with macroscopic clinical and TCM theories,
the present review provided original insights to inform the
development of next-generation, targeted anti-atherosclerotic
therapeutics.
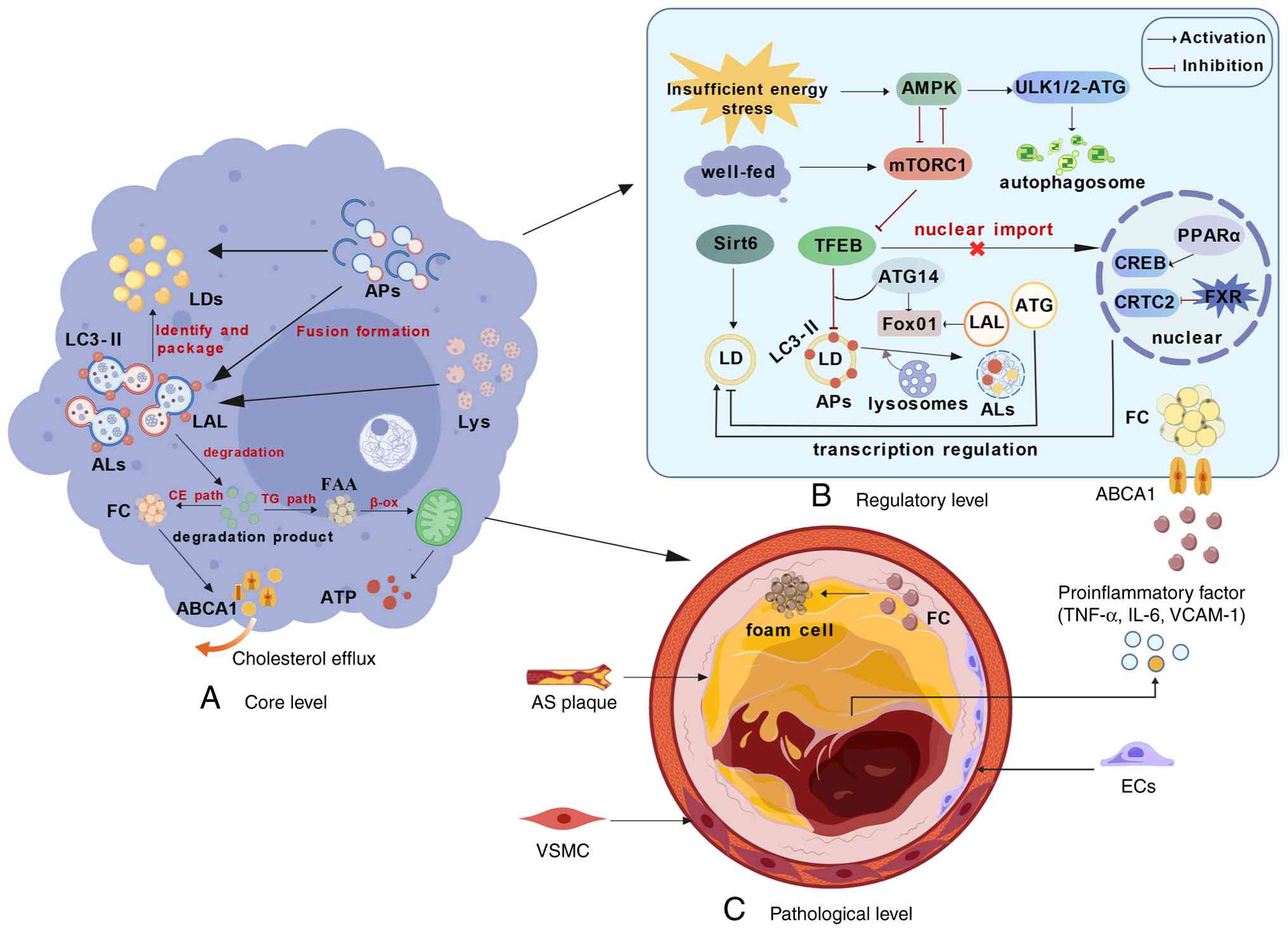 | Figure 1.Core mechanisms of lipophagy in AS
and therapeutically targetable pathways. Created with BioGDP.com
(142). (A) Core execution level:
Under lipotoxic stress, intracellular LDs are recognized and
sequestered by phagophores decorated with LC3-II, forming APs.
These APs subsequently fuse with lysosomes to generate
autolysosomes, in which lysosomal acid lipase hydrolyzes LDs. The
degradation products (free fatty acids) undergo mitochondrial
β-oxidation to produce ATP, while free cholesterol is expelled via
the ABCA1 transporter (cholesterol efflux). (B) Upstream regulatory
network: The initiation of lipophagy is strictly governed by
nutrient-sensing signaling cascades. AMPK activation triggers
autophagy by phosphorylating the ULK1/2 complex, whereas the
activated mTORC1 pathway suppresses this process. At the
transcriptional level, transcription factor EB serves as a master
regulator; its dephosphorylation facilitates nuclear translocation
to upregulate autophagy-related genes. Additionally, Sirt6
epigenetically promotes lipophagy, and the FXR-CREB axis modulates
this process: FXR inhibits the CREB-CRTC2 complex, suppressing
lipophagy, while PPARα counteracts this effect. (C) Pathological
manifestation level: In the arterial wall, impaired lipophagic flux
leads to massive lipid accumulation in macrophages, driving their
transformation into pro-inflammatory foam cells. Concurrently,
dysfunctional lipophagy in endothelial cells and vascular smooth
muscle cells exacerbates vascular inflammation and pathological
remodeling, ultimately accelerating atherosclerotic plaque
progression. LDs, lipid droplets; Aps, autophagosomes; ALs,
autolysosomes; LALs, lysosomal acid lipase; FFAs, free fatty acids;
Lys, lysosome; EC, endothelial cell; VSMC, vascular smooth muscle
cell; AS, atherosclerosis; TG, triglyceride; CE, cholesterol ester;
β-ox, β-oxidation; FC, free cholesterol. |
Literature search strategy
To comprehensively review the role of lipophagy in
AS, a systematic literature search was performed using PubMed
(https://pubmed.ncbi.nlm.nih.gov/), Web
of Science (https://www.webofscience.com/) and Embase (https://www.embase.com/). The search covered
literature published from database inception to October 2025. The
following keywords and combinations were used: i) Lipophagy
(Title/Abstract); ii) atherosclerosis (Title/Abstract); iii) and
lipophagy and atherosclerosis. Articles written in English,
including basic research, clinical studies and high-quality
reviews, were independently screened for relevance. Studies
primarily focusing on non-cardiovascular diseases or lacking direct
mechanistic links to lipid metabolism in AS were excluded.
Physiological functions of lipophagy
The core mechanisms of lipophagy encompass lipid
uptake, storage and degradation (10). As these processes facilitate the
maintenance of cellular lipid homeostasis, lipophagy is integral to
the pathogenesis of cardiovascular diseases (such as
cardiometabolic syndrome and diabetic cardiomyopathy) (10). Research has concentrated on
lipophagy as a notable physiological and pathological process that
is particularly relevant within the context of AS (10). Lipophagic degradation releases free
fatty acids and glycerol; this mechanism both supports energy
acquisition during periods of nutrient deprivation and regulates
intracellular lipid homeostasis, thereby mitigating the
cytotoxicity associated with excessive lipid accumulation (11). Within ALs, TG-rich LDs are degraded
into free fatty acids, which subsequently undergo β-oxidation,
producing adenosine triphosphate (ATP) (12,13).
Concurrently, LDs enriched with CE are hydrolyzed into free
cholesterol via ABCA1-mediated lipophagy, and the free cholesterol
is then exported to the extracellular space (14).
Farnesoid X receptor (FXR) and cAMP response
element-binding protein (CREB) have been identified as pivotal
transcription factors involved in lipophagy regulation, and the
FXR/CREB signaling pathway plays a notable role in modulating the
initiation of lipophagy (15). At
the molecular level, CREB is activated by phosphorylation at the
Ser133 residue, which triggers the recruitment of CREB-regulated
transcription coactivator 2 (CRTC2), resulting in the formation of
a transcriptionally active complex. This complex robustly
upregulates autophagy-related genes, including ATG7 and
ULK1 (16–18). By contrast, FXR exerts an
inhibitory effect by competitively binding to CRTC2. This
stoichiometric competition physically sequesters CRTC2 from CREB,
preventing the assembly of the CREB-CRTC2 transcriptional machinery
on the promoters of lysosomal genes. Thus, the dynamic equilibrium
between FXR and CREB in competition for CRTC2 binding serves as a
molecular switch for lipophagy initiation (19). Notably, PPARα activation
counteracts the suppressive influence of FXR on lipophagy (20,21)
(Fig. 1). The disruption of
lipophagy may result from excessive lipid accumulation in tissues,
as observed in conditions such as AS and hepatic steatosis
(11,22,23).
Autophagic impairment has been prominently observed in
atherosclerotic plaques in both human and animal studies (24,25).
Impaired lipophagy, characterized by the suppression of lipid
droplet-lysosome fusion and inhibited lysosomal biogenesis, has
also been observed in foam cells, suggesting an association between
lipophagy and AS (26). During the
advanced stages of AS, dysregulation of lipophagy leads to
disrupted lipid metabolism. This disturbance subsequently affects
macrophage survival and apoptosis, potentially promoting plaque
instability. Ultimately, plaque rupture can occur, further
exacerbating pathological progression (27).
Molecular mechanisms regulating lipophagy in
AS
Molecularly, lipophagy involves a sophisticated
sequence of recognition and engulfment. Beyond general
macroautophagy, the specific recruitment of LC3 to the LD surface
is facilitated by adapter proteins such as p62 and NBR1, which bind
to ubiquitinated perilipins (PLINs) (28). Furthermore, the Rab family of small
GTPases (specifically Rab7 and Rab18) is a notable mediator of the
physical contact between LDs and the isolation membrane, ensuring
the precise sequestration of lipid cargo into nascent
autophagosomes (29,30).
Research has uncovered a complex regulatory network
governing these processes. Central to this network are several
master transcription factors, including TFEB (31), TFE3 (32), PPARα (33) and forkhead box protein O1 (FoxO1)
(34), alongside key epigenetic
modulators such as sirtuin 6 (Sirt6) (35). TFEB and Sirt6 are notable
protectors against AS progression that enhance lysosomal biogenesis
and macrophage lipid clearance, whereas FoxO1 regulates lipophagic
flux by activating ATG14 and lysosomal acid lipase (LAL) expression
(36,37). These diverse regulatory elements
are organized into several core signaling axes.
AMPK-mechanistic target of rapamycin
(mTORC1) energy sensing axis
The AMPK and mTOR signaling pathways are key
regulators of lipophagy efficiency owing to their influence on
lipid synthesis and oxidation processes (38). Analogous to that of autophagy, the
initiation of lipophagy is governed by the inhibition of specific
signaling pathways (particularly the PI3K/AKT/mTOR pathway). mTOR
predominantly regulates the lipophagic process through its complex,
mTORC1 (39). Lipophagy initiation
is dictated by the dynamic interplay between AMPK and mTORC1, which
are the primary cellular energy sensors (40). mTORC1 activation facilitates ULK1/2
and ATG13 phosphorylation, which in turn inhibits the formation of
the ULK1/2-ATG13 complex (41,42).
mTORC1 modulates lipophagy via two distinct mechanisms. Initially,
lipophagy is inhibited through the modulation of TFEB nuclear
translocation, and subsequently, suppression is accomplished by
regulation of ATG14 activity (43,44).
AMPK activation directly inhibits mTORC1 activity by
phosphorylating and enhancing RHEB GTPase-activating protein
activity, as well as by disrupting the stability of the mTORC1
complex through RAPTOR phosphorylation. Simultaneously, mTORC1
exerts negative feedback regulation by phosphorylating the AMPKα
subunits at the Ser347 and Ser345 residues. This reciprocal
modulation establishes a dynamically balanced metabolic regulatory
network (45). In this process,
lipophagy regulation is intricately linked to macrophage function
as well as the overall status of systemic lipid metabolism.
Moreover, disruptions in hepatic lipid metabolism are markedly
associated with increased AS risk, underscoring the notable role of
lipophagy in maintaining systemic lipid homeostasis (46). Heterogenous nuclear
ribonucleoprotein A2/B1 (hnRNPA2B1)-knockout hepatocytes exhibit
increased co-localization of APs and LDs and enhanced LD lysosomal
degradation, suggesting that lipophagy is crucial for reducing
lipid deposition. The absence of hnRNPA2B1 decreases lipid droplet
accumulation by preventing Atg5 degradation and boosting
autophagy-driven lipophagy (47).
TFEB, the master transcriptional
regulator
TFEB, recognized as a key regulator of autophagy and
lysosome biogenesis, is considered to play a marked role in various
pathological processes associated with the development of AS
(48). TFEB is a marked regulator
of lipid metabolism, overseeing transcriptional responses that
control lysosomal biogenesis and autophagy. With roles in autophagy
regulation, lysosomal formation, fusion and substrate degradation,
TFEB promotes lipid breakdown and transport, making it a potential
target for regulating lipophagy (49). TFEB has been shown to facilitate LD
degradation and enhance lipid metabolic homeostasis by activating
the lipophagy pathway. This mechanism may reduce lipid accumulation
in macrophages and attenuate inflammatory responses, potentially
impeding AS progression (50).
TFEB transcriptional activity is tightly regulated by
post-translational modifications, primarily phosphorylation and
acetylation, which dictate its subcellular localization. Under
nutrient-rich conditions, mTORC1 phosphorylates TFEB at specific
residues (such as Ser211), promoting its interaction with 14-3-3
proteins and resulting in its cytosolic sequestration. Conversely,
during lipid stress or atherogenic conditions, mTORC1 inhibition or
AMPK activation leads to TFEB dephosphorylation (51). Concurrently, NAD+-dependent
deacetylases, particularly SIRT1, deacetylate TFEB at key lysine
residues (such as Lys116) (52).
This synergistic dephosphorylation and deacetylation trigger the
rapid nuclear translocation of TFEB, where it binds to coordinated
lysosomal expression and regulation motifs. This binding markedly
upregulates the expression of essential lipophagy and cholesterol
efflux genes, including MAP1LC3B, SQSTM1 (p62) and
ABCA1, thereby accelerating lipid droplet degradation and
reducing lipotoxicity (53). TFEB
enhances lipid breakdown and cholesterol removal via the
autophagy-lysosome pathway and PPARα-PGC1α signaling while also
stabilizing AS plaques by inhibiting NLRP3 inflammasome activation
(38). Although the precise
molecular mechanisms by which TFEB regulates AS are not yet fully
understood, evidence suggests that TFEB expression levels vary
among different cell types (48).
These variations may affect AS pathological progression through
cell-specific regulatory networks.
SIRT6 and epigenetic regulation
Sirt6, a deacetylase enzyme, plays a notable role in
lipophagy and lipid metabolism. It has been implicated in the
pathogenesis of age-related diseases, cancer and diabetes (54–56),
and Sirt6 regulation offers a new strategy for AS treatment. Sirt6
knockout increases plaque area in ApoE-/-mice, while its
overexpression reduces plaque size and slows AS progression by
downregulating MMP-2 and suppressing hypoxia inducible factor-1α,
toll-like receptor-4 and von Willebrand factor. This enhances
plaque stability by reducing intraplaque neovascularization
(57). In macrophages, increasing
Sirt6 expression boosts autophagy, reducing macrophagic
interactions with ECs and decreasing adhesion and infiltration.
This suppresses macrophage apoptosis and enhances atherosclerotic
plaque stability (57). Studies
have further highlighted the multifaceted protective roles of SIRT6
in autophagy regulation and inflammation resolution across
different vascular cell types. For example, Li et al
(58) demonstrated that SIRT6
epigenetically enhances macrophage efferocytosis, a specialized
phagocytic and autophagic clearance mechanism, by hindering
miR-216/217 cluster maturation, which promotes the resolution of
chronic inflammation. Furthermore, Zi et al (59) showed that the protective role of
SIRT6 extends beyond macrophages by revealing that SIRT6-induced
autophagy effectively restricts TREM-1-mediated pyroptosis in
oxidized low-density lipoprotein (ox-LDL)-treated ECs. This effect
notably reduces endothelial damage and inflammatory cell death.
These findings collectively underscore that SIRT6 not only prevents
macrophage foam cell formation but also globally attenuates
vascular inflammation by modulating autophagic flux, reinforcing
its potential as a multidimensional epigenetic target for plaque
stabilization. Sirt6 also promotes lipophagy by inhibiting the Wnt1
pathway, which reduces the accumulation of LDs and associated
proteins, facilitating their degradation. These actions
collectively reduce foam cell formation and inhibit AS development
(35).
PLIN family proteins and selective
recognition via chaperone-mediated autophagy (CMA)
PLINs, the predominant proteins on LD surfaces,
localize to specific binding sites to modulate lipid storage and
hydrolysis (60). As notable
surface proteins associated with LDs, PLINs initiate lipophagy by
undergoing degradation (61). The
PLIN protein family, consisting of PLIN1-5, regulates lipase
binding to LDs during lipophagy, influencing both macroautophagy
and CAM. PLIN1, required for LD function, plays a notable role in
metabolic regulation. In adipocytes, its absence enhances
liposome-lysosome fusion and LD-lysosome interactions, accelerating
macroautophagy-mediated lipolysis (62). Structurally, the CMA pathway
selectively recognizes KFERQ-like motifs within the amino acid
sequences of PLIN2 and PLIN3, and the cytosolic chaperone heat
shock cognate 70 kDa protein (Hsc70) specifically identifies and
binds to these pentapeptide motifs. As a result of this structural
interaction, the PLIN-Hsc70 complex is delivered to the lysosomal
membrane, where it engages with the lysosome-associated membrane
protein 2A (LAMP-2A) receptor for translocation and subsequent
degradation. Moreover, PLIN2/3 removal via this process reveals the
lipid droplet surface, which is a prerequisite for the
macroautophagic machinery to recognize and engulf the lipid cargo.
PLIN2 also facilitates AMPK signaling activation by interacting
with the HSP70 chaperone protein, effectively coupling lipid
droplet sensing to lipophagy upstream modulation (63). Beyond structural interactions,
different PLIN family members exert distinct macroscopic effects on
AS. Research on human carotid plaques and macrophage models shows
that PLIN2 encourages early lipid droplet formation and
proinflammatory macrophage behavior, with higher mRNA expression
observed in symptomatic plaques (64). By contrast, PLIN1 stabilizes large
LDs and supports the anti-inflammatory balance by reducing
cholesterol efflux and markedly lowering TNFA, MMP2, ABCA1
and ABCG1 mRNA levels (64). Collectively, these functions
demonstrate the pivotal roles of PLIN family proteins in the
regulation of lipophagic processes.
Emerging external regulators: Gut
microbiota and metabolites
Research into metabolites as regulators has
confirmed that gut microbiota metabolites directly modulate AS
development through lipophagy. Pro-atherogenic substances such as
trimethylamine N-oxide promote lipid accumulation by specifically
inhibiting macrophage lipophagy. By contrast, beneficial
metabolites such as short-chain fatty acids and secondary bile
acids improve lipid metabolism by activating the GPCR/FXR signaling
pathway (65,66). Additionally, ferulic acid
restructures the gut microbiota by specifically reducing the
abundance of Firmicutes, Erysipelotrichaceae and
Ileibacterium, which subsequently modulates fecal
metabolites, activating the AMPK/SREBP1/ACC1 signaling pathway
(67). Activation of this pathway
mitigates lipid accumulation by promoting lipophagy, highlighting a
therapeutic mechanism involving the gut-vascular axis (67).
Role of lipophagy in AS
AS is a complex condition characterized by lipid
buildup in arterial walls, which leads to arterial narrowing and
plaque formation. Key features of AS include VSMC proliferation and
migration, endothelial dysfunction and vascular remodeling.
Lipophagy markedly influences these pathological changes throughout
AS development (38).
Macrophages
Through lipophagy, LDs in macrophages are
transported to lysosomes, where they are hydrolyzed into free
cholesterol by LAL. Subsequently, the free cholesterol is exported
to the extracellular environment via the cholesterol reverse
transport pathway, which is mediated by the ATP-binding cassette
transporters ABCA1 and ABCG1 (68). The clearance of excess lipids via
macrophagic lipophagy prevents the formation of foam cells,
delaying the progression of AS (69). Macrophage ACE expression regulates
lipid metabolism by activating PPARα, which boosts β-oxidation and
cholesterol efflux. This increases mitochondrial respiration and
ATP production, reducing foam cell formation and promoting M2
polarization and efferocytosis, notably slowing AS progression
(70). While lipophagy initially
plays an atheroprotective role by promoting cholesterol efflux, its
role in advanced AS represents its capacity to act as a
‘double-edged sword’. Under conditions of severe lipotoxic stress,
excessive or dysregulated lipophagy can overwhelm lysosomal
capacity, leading to lysosomal membrane permeabilization,
macrophage apoptosis and impaired efferocytosis (71). This transition ultimately
contributes to necrotic core expansion and plaque instability.
Therefore, understanding the temporal dynamics of lipophagy is
required.
Lipophagy and VSMC proliferation,
migration and apoptosis
VSMCs are key in maintaining intravascular
homeostasis, which includes processes such as contraction, dilation
and vascular remodeling. As a result, VSMCs are recognized as
cellular determinants in the pathology of arterial walls and are
notably associated with cardiovascular diseases, including AS
(72). Recent transcriptomic and
multi-omics studies (15,57,67)
have shown that VSMC proliferation and migration, which occur as a
result of a switch from a contractile state to a synthetic state,
cause arterial wall thickening. Subsequently, atherosclerotic
plaques undergo growth and structural changes driven by the PDGF/BB
signaling pathway and lncRNA-ITGA2-related 3D genome interactions,
which are key in AS pathology (73). VSMCs are involved in vascular
remodeling and markedly contribute to neointimal formation through
their proliferative and migratory activities in AS (74). Enhanced lipophagy is intricately
associated with VSMC proliferation; this relationship is
particularly pronounced under hyperlipidemic conditions, wherein
the proliferation of VSMCs and their transformation into foam cells
are facilitated by the uptake of lipids, such as ox-LDL (75). Furthermore, peroxidase derived from
millet bran markedly inhibits lipid phagocytosis and cell
proliferation in human aortic smooth muscle cells (HASMCs). This
enzyme facilitates the transition of HASMCs toward a contractile
phenotype and concurrently suppresses their migratory capacity.
This transition inhibits the phosphorylation of STAT3, which
mediates inflammatory responses, reducing aortic plaque formation
(76). Concurrently, lipid
accumulation and enhanced lipophagy induce VSMC migration.
Migration is particularly augmented within inflammatory
microenvironments, where VSMCs express specific chemokines and cell
adhesion molecules, enabling motility (77).
Lipophagy also plays a notable role in apoptosis
regulation in VSMCs. AS progression is often associated with VSMC
apoptosis, and the degree of lipophagy markedly affects this
process (78). Excessive lipid
uptake by VSMCs can initiate cellular clearance by inducing
apoptosis. Nonetheless, excessive lipid accumulation and the
lipophagy process may further exacerbate cellular death (79). PPARγ coactivator-1 alpha (PGC-1α)
suppresses glucose-driven VSMC proliferation, migration and
inflammation, which are crucial aspects of AS pathology. PGC-1α
encourages VSMCs to differentiate into a macrophage-like form,
potentially increasing the quantity of VSMCs-derived foam cells in
plaques and aiding plaque stabilization (80). The gelsolin-mediated
lipophagy-related signaling pathway is considered a necessary
mechanism in safeguarding VSMCs against apoptosis, and this
discovery offers new therapeutic insights supporting AS treatment
(81).
As a pivotal regulator of VSMC function, lipophagy
is a prospective therapeutic target for AS (26,82).
The dual role of lipophagy in VSMCs must be considered in the
context of overall plaque stability. Specifically, the lipophagic
flux in VSMCs serves as a determinant of plaque structural
integrity. While physiological lipophagy prevents the
transdifferentiation of VSMCs into foam cells by mobilizing
intracellular lipids, excessive or dysfunctional autophagic flux
can trigger VSMC apoptosis (82).
This depletion of VSMCs within the fibrous cap directly compromises
its mechanical strength, as these cells are the primary source of
the extracellular matrix, particularly interstitial collagen
(83). A reduced VSMC population
leads to a net loss of collagen fibers and progressive thinning of
the fibrous cap, thereby transforming a stable lesion into a
vulnerable plaque prone to rupture. Thus, the ‘double-edged sword’
of lipophagy in VSMCs represents a pivotal link between cellular
lipid homeostasis and macroscopic plaque stability (84,85).
Thus, the net effect of VSMC lipophagy depends on the disease stage
and the intensity of autotoxic stress.
Association between lipophagy and EC
dysfunction
ECs are notable contributors to AS and serve as
regulatory centers throughout the progression of the disease. These
cells participate in various physiological processes, such as
intracellular signal transduction, cell adhesion, immune response
and inflammation (38). EC
dysfunction (ECD) involves a compromised endothelial barrier,
increased inflammation, oxidative stress imbalance and decreased
nitric oxide availability, leading to lipoprotein leakage, monocyte
recruitment and plaque formation. As plaques progress, ECD
exacerbates inflammation by releasing proinflammatory and
prothrombotic factors, contributing to plaque instability (86). Research has elucidated the notable
role of lipophagy in preserving EC function, and ECD is
acknowledged as an early indicator of AS. Lipophagic deficiency can
produce lipid accumulation within ECs, triggering oxidative stress
and inflammatory responses, which further aggravates ECD (87). ox-LDL binds to specific receptors
on ECs, facilitating EC lipid uptake; this mechanism induces lipid
overload and subsequently leads to EC apoptosis (88). Enhanced lipophagy in ECs helps
clear excess lipids, improving EC function and vascular health. It
also reduces inflammation, a key factor in AS development, by
suppressing EC levels of pro-inflammatory cytokines (89). For example, lipophagy mitigates
ox-LDL-induced inflammatory responses in ECs by facilitating
intracellular ox-LDL clearance, thus providing protection against
EC damage (90,91). Lipophagy mitigates inflammatory
responses by facilitating EC repair and regeneration through the
modulation of specific autophagy-related signaling pathways, such
as the CaMKKβ/AMPK, SOCE and PI3K/AKT pathways (92,93).
The role of lipophagy in EC inflammation extends beyond lipid
clearance; it also encompasses regulation of cytokine expression
and signaling pathway activity (93,94).
Enhanced lipophagy markedly improves EC function and augments
resistance to oxidative stress and inflammation. Certain natural
compounds, such as naringenin, have been shown to facilitate EC
lipophagy, thereby mitigating ox-LDL-induced cellular damage
(95). Lipophagy is integral to EC
functional recovery; consequently, lipophagy modulation has
considerable therapeutic potential in the context of AS.
Effects of lipophagy on vascular
remodeling
Vascular remodeling is a pathological mechanism
critical to AS progression, characterized by structural
reorganization of the vascular wall, diminished elasticity, tissue
calcification, plaque rupture and luminal narrowing. These
phenomena are manifested through a range of pathological processes
(70). For example, as a key
regulator of lipid metabolism and inflammatory responses, lipophagy
is crucial for VSMC remodeling in AS (96). VSMCs not only function as
structural support cells but also play a role in lipid metabolism
during atherogenesis. By promoting lipid degradation, lipophagy
mitigates lipid accumulation within VSMCs, thereby limiting AS
progression (97). Macrophages are
the primary cells responsible for lipid accumulation within
arterial walls. In macrophages, lipophagy mitigates the
intracellular accumulation of CEs and TG through LD degradation via
lysosomal pathways, thereby preventing foam cell formation and
promoting cholesterol efflux (6,68,98).
LAL is crucial in the lipophagy pathway, and reduced LAL expression
decreases its activity in macrophages, resulting in more LDs,
faster foam cell formation and vascular thickening. Conversely, LAL
overexpression reduces lipid buildup by boosting the expression of
genes such as Hmgb1, Hmgb2, Hspa5 and Scarb2. This
also decreases cholesterol efflux and enhances lipophagy
coordination (6,99). Lipophagic deficiency has been shown
to expedite the progression of unstable atherosclerotic plaques.
Impaired lipophagy within macrophages causes increased lipid core
size, diminished collagen content and thinning of the fibrous cap
in plaques, collectively enhancing the risk of plaque rupture
(66). Vascular endothelial injury
triggers vascular remodeling; however, lipophagy shields ECs from
ox-LDL and advanced glycation end product damage by maintaining
redox balance, clearing damaged mitochondria and lipotoxic
products, preventing lipid droplet buildup and reducing lipid
deposition. In addition, lipophagy also decreases inflammatory cell
secretion, further protecting vascular ECs (100). Preclinical studies indicate that
lipophagy activators, including berberine (BBR), decrease the
levels of inflammatory factors such as TNF-α and IL-6 through a
SIRT3-mediated lipophagy pathway. This mechanism enhances
endothelium-dependent vasodilation and attenuates the progression
of vascular stiffening (101).
Enhancing lipophagic activity may therefore constitute a pivotal
strategy for improving the structural integrity of the vascular
wall and preventing AS.
Other cell types
Beyond macrophages and VSMCs, emerging evidence
suggests that lipophagy serves as a notable metabolic rheostat in
other non-canonical cell types, notably T lymphocytes and
fibroblasts (102). In T
lymphocytes, lipophagy--mediated mobilization of triacylglycerols
provides free fatty acids for mitochondrial fatty acid oxidation,
an energetic process essential for T-cell activation and the
maintenance of regulatory T cell populations. A deficiency in
autophagic lipid processing can impair Treg suppressive function
and promote a pro-inflammatory Th17 phenotype, thereby intensifying
the inflammatory milieu within the plaque. Similarly, in vascular
fibroblasts, lipophagy-mediated lipid turnover maintains
intracellular lipid proteostasis, thereby suppressing cellular
senescence and the pathological fibroblast-to-myofibroblast
transition by mitigating oxidative stress (102–104). By governing the availability of
lipid-derived signaling molecules, lipophagy influences the
production of the extracellular matrix, which is required for the
structural integrity of the adventitia and media (105). These emerging insights suggest
that lipophagy represents a universal metabolic hub across diverse
cell populations in the progression of AS.
Therapeutics and active compounds for AS
prevention and treatment
In terms of clinical applications, targeting
lipophagy presents a viable yet complex therapeutic frontier.
Current pharmacological strategies can be broadly categorized into
two paradigms: Conventional lipid-lowering agents and holistic
multi-target modulators (such as natural compounds and TCM)
(106,107). While conventional drugs such as
statins primarily promote lipophagy indirectly as a secondary
consequence of systemic lipid reduction and upstream AMPK
activation, TCM formulas and their active botanical monomers (such
as BBR) often act directly on intracellular epigenetic and
transcriptional networks (such as the SIRT1-TFEB axis) to restore
specific autophagic flux. The contrast between these approaches
highlights a shift from broad systemic metabolic control toward
more nuanced, multi-targeted intracellular regulation. In the
following sections, these therapeutic agents are systematically
evaluated based on their mechanisms of action.
Autophagy activators
Autophagy activators are compounds employed to
stimulate or augment autophagy. These agents operate by activating
autophagy-related signaling pathways or modulating the expression
of notable autophagy-associated proteins. The underlying mechanisms
encompass negative regulation of the mTOR signaling pathway and
activation of AMPK, among others (108). mTOR is a highly conserved
serine/threonine protein kinase with a pivotal role in the
regulation of fundamental biological processes such as cell growth,
proliferation, protein synthesis, energy metabolism and autophagy
(109). Everolimus, the most
extensively researched mTOR inhibitor, has been thoroughly
investigated to determine its therapeutic potential against AS,
owing to its autophagy-inducing properties (110). It notably enhances plaque
stability, and the application of everolimus-eluting
cobalt-chromium platforms to cardiac stents facilitates the
development of a stable plaque phenotype (110,111). Everolimus has been shown to
inhibit AS progression, and its therapeutic efficacy appears to be
more pronounced in the early stages of AS than in the advanced
stages (112). However, the
clinical translation of broad mTOR inhibitors such as everolimus,
whose therapeutic efficacy varies between early and advanced
lesions in LDL-R-deficient mice and clinical studies, is markedly
limited by systemic immunosuppressive effects and metabolic side
effects, including treatment-induced hypercholesterolemia, systemic
immunosuppressive effects and metabolic side effects, emphasizing
the need for cell type-specific delivery systems (109–111).
Metformin (Met) is commonly prescribed as a
first-line pharmacological treatment for type 2 diabetes mellitus,
and it is acknowledged as an AMPK activator (112). Patients with AS exhibit AMPK and
ERK pathway suppression, which contributes to abnormal lipid
metabolism and inflammation (113). Met reduces plaque buildup caused
by a high-fat diet, exerting a protective effect against AS. Mice
fed a high-fat diet exhibit an increased proportion of
Ki67-positive proliferating cells and α-SMA-positive VSMCs within
the arterial media. Met decreases abundance of these dual-positive
cells and ERK levels while increasing AMPK and Pdlim5 levels. This
finding indicates that Met selectively counteracts the pathological
VSMC proliferation promoted by a high-fat diet, thereby stabilizing
the vascular wall and mitigating AS (112).
Atorvastatin is a primary therapeutic agent for
lipid reduction and plaque stabilization that exhibits
anti-oxidative, anti-inflammatory, anti-thrombotic and
anti-atherosclerotic properties (114). Additionally, it enhances
endothelial function and contributes to plaque stabilization
(115). It inhibits foam cell
formation by promoting lipophagy, marked by increased Atg protein
expression, a higher LC3-II/LC3-I ratio and reduced p62 levels. It
also enhances LC3 and LD co-localization in treated foam cells,
suggesting an ability to induce autophagy in macrophages and reduce
intracellular lipid buildup (116). Clinically, while atorvastatin is
widely prescribed for systemic lipid-lowering at standard doses of
20–80 mg/d, ongoing clinical evaluations are required to determine
whether these conventional regimens are sufficient to optimally
induce macrophage lipophagy within the complex plaque
microenvironment (117,118). Furthermore, the dose-dependent
risk of statin-associated muscle symptoms necessitates the
exploration of localized targeted delivery strategies (119,120).
Herbal monomers
Increased research into the anti-AS effects of TCM
extracts has confirmed the efficacy of numerous herbal monomers
against AS. Additionally, synergistic combinations of multiple
TCM/natural products (such as multicomponent botanical
compositions) demonstrate greater anti-AS activity and fewer side
effects than isolated compounds used alone (121). Geniposide (GE), a water-soluble
iridoid glycoside also referred to as genipin-1-β-gentiobioside,
has been identified in >40 plant species, and it exhibits a
range of pharmacological effects and biological activities
(122). GE boosts lipophagy by
inhibiting the PARP1/PI3K/AKT pathway, accelerating LD breakdown
and reducing lipid buildup. This slows AS development by preventing
apoptosis and foam cell marker expression in VSMC-derived foam
cells (123).
Epigallocatechin gallate (EGCG) is the most abundant
and biologically active compound in tea polyphenols; it exhibits
multiple biological effects, including anti-inflammatory and
anti-oxidant (124), anti-obesity
(121), as well as anti-infection
and anti-tumor activities (125).
EGCG activates autophagic flux via the Ca2+/CaMKKβ/AMPK
pathway, reducing lipid buildup in vascular ECs by promoting
lipophagy. EGCG releases Ca2+ from the endoplasmic
reticulum, triggering CaMKKβ and AMPK/ULK1 phosphorylation
(bypassing mTOR) and subsequently increasing LC3-II formation and
p62 degradation (126). EGCG
enhances lysosomal degradation by co-localizing LDs with LC3 and
lysosomes, ultimately reducing palmitate-induced lipid
accumulation, improving endothelial lipotoxicity and mitigating AS
(127,128).
BBR, an isoquinoline alkaloid derived from Coptis
chinensis and various Berberis species, demonstrates
anti-atherosclerotic properties through a range of mechanisms such
as lipid regulation, anti-inflammatory effects, oxidative stress
reduction, vascular endothelial dysfunction mitigation, modulation
of VSMC proliferation and migration and anti-thrombotic activity
(129). BBR boosts the
intracellular NAD+/NADH ratio by activating NAD+ biosynthesis
pathways, enhancing SIRT1 deacetylase activity (130,131). Activated SIRT1 then deacetylates
TFEB, increasing the LC3-II/LC3-I ratio and Beclin 1 expression
(132). This process promotes
autophagosome formation, inducing lipophagy and promoting LD
degradation while delaying cell apoptosis. Together, these effects
intervene in AS lipid metabolism. Despite these promising in
vitro mechanisms, the clinical translation of BBR is
significantly hindered by its poor oral bioavailability. Achieving
sustained therapeutic concentrations specifically within
atherosclerotic plaques remains a major challenge, emphasizing the
necessity for advanced cell-type-specific delivery systems
(130,131). Future clinical applications will
likely depend on the development of novel nanoparticle-based
delivery systems that enhance lipophagic efficacy in
vivo.
The therapeutic landscape of natural compounds
targeting AS is rapidly expanding beyond traditional signaling
pathways to encompass epigenetic regulation and intercellular
communication. Recent comprehensive studies have emphasized that
natural medicinal active ingredients and TCMs can effectively
prevent and treat AS by directly targeting epigenetic modifications
(133). For example,
intercellular crosstalk via extracellular vesicles (EVs), such as
EV-derived miR-146a targeting SMAD4, has recently been identified
as a novel pathogenic mechanism in high-fat diet-induced AS
(134). Botanical extracts
possess a profound capacity to modulate such epigenetic networks,
evidenced by their ability to mitigate chronic inflammatory
diseases by markedly altering micro RNA expression profiles
(135). Consequently, future
investigations into natural lipophagy modulators should explore
their potential to concurrently regulate EV miRNAs and the
epigenetic landscape, which would offer a multi-dimensional,
systemic approach to plaque stabilization.
Chinese herbal formulas
Research examining the therapeutic effects and
underlying mechanisms of TCM interventions in the context of AS has
markedly increased in recent years (136,137). TCM formulations demonstrate
efficacy in alleviating AS symptoms and decelerating disease
progression, attributed to multi-component actions, multi-target
mechanisms and minimal side effects (138). Tianxiangdan enhances lipophagy by
inhibiting the PI3K/Akt/mTOR pathway, reducing p85/Akt/mTOR
phosphorylation. This activates TFEB and the autophagy proteins
ULK1/LC3-II, boosting LD degradation. Tianxiangdan also increases
ABCA1 expression and suppresses expression of p62, raising
lipophagy levels, reducing foam cell formation, decreasing arterial
plaque lipid deposition and slowing AS progression (139). Recent molecular docking and
pharmacological analyses indicate that rather than acting as a
simple generalized inhibitor, the Huoxue Qutan Formula contains
primary active constituents such as specific flavonoids and
saponins that directly interact with upstream nutrient-sensing
kinases. This targeted interaction persistently suppresses mTORC1
phosphorylation, thereby mechanically uncoupling mTORC1 from TFEB
and enhancing TFEB nuclear translocation, increasing LC3-II/I and
Beclin1 levels, and reducing p62 levels. This process leads to
improved co-localization of LDs with autophagosomes and upregulates
the expression of the cholesterol efflux proteins ABCA1 and SCARB1
through TFEB pathways, reducing foam cell formation and lipid
deposition in atherosclerotic plaques (140). The Gualou-Xiebai herbal
combination inhibits P2RY12 activation, downregulating p62 and
Plin2 expression and upregulating LC3II expression. The quantity of
autophagosomes is also increased, markedly enhancing lipophagy
activity. Consequently, foam cell formation is reduced, and
anti-atherosclerotic effects are achieved (141).
In conclusion, accumulating evidence suggests that
pharmacological agents and active compounds that modulate lipophagy
exhibit notable therapeutic potential for AS prevention and
treatment. Research priorities in this domain include autophagy
inducers, monomeric TCM components and TCM compounds.
Mechanistically, these agents operate by activating
autophagy-related signaling pathways, regulating the activity of
key autophagy proteins, improving dysregulated lipid metabolism and
inflammatory responses, inhibiting foam cell formation, exerting
anti-inflammatory and antioxidant effects, ameliorating vascular
endothelial dysfunction and modulating VSMC proliferation and
migration (Table I). Further
investigation is necessary to elucidate the precise mechanisms and
assess the clinical feasibility of these pharmacological
agents.
 | Table I.Pharmacological agents and active
compounds targeting lipophagy for the prevention and treatment of
AS. |
Table I.
Pharmacological agents and active
compounds targeting lipophagy for the prevention and treatment of
AS.
| A, Autophagy
activator |
|---|
|
|---|
| Drug
ingredients | Mechanism of
action | Method | Model | Targets | Actions | (Refs.) |
|---|
| Everolimus | 1. Inhibits mTORC1
signaling. | In vivo
and | In
vitro: | mTORC1,
Survivin, | 1. Selectively
reduced foam cell viability. | (108–110) |
|
| 2. Induces foam
cell apoptosis | In
vitro | 1. Human THP-1 | Clusterin,
MAP1LC3, | 2. Induced
autophagy |
|
|
| and autophagy. |
|
macrophage-derived | MMP1 and MCP-1 | 3. Inhibited matrix
degradation. |
|
|
| 3. Suppresses
matrix degradation |
| foam cells |
| 4. Reduced
pro-inflammatory secretion. |
|
|
| and
pro-inflammatory cytokine |
| 2. HCASMCs |
| 5. Decreased
macrophage infiltration |
|
|
| secretion. |
| 3. HUVECs |
| in
vivo. |
|
|
|
|
| In
vivo: |
|
|
|
|
|
|
| 1.
Hypercholesterolemic |
|
|
|
|
|
|
| rabbit iliac
artery |
|
|
|
|
|
|
| model |
|
|
|
| Metformin | 1. Activates the
AMPK pathway. | In vivo | C57/B6 mice | AMPK, ERK and | 1. Attenuated
atherosclerotic deposition. | (111,112) |
|
| 2. Inhibits the ERK
pathway. |
|
| Pdlim5 | 2. Inhibited VSMC
proliferation and |
|
|
|
|
|
|
| migration. |
|
| Atorvastatin | 1. Upregulates
phosphorylation | In
vitro | Macrophages | Atg, LC3-II/LC3-I
and | 1. Inhibited foam
cell formation. | (114) |
|
| of AMPK. |
|
| p62 | 2. Reduced lipid
droplet accumulation. |
|
|
| 2. Downregulates
phospho- |
|
|
|
|
|
|
| rylation of the
mTOR. |
|
|
|
|
|
|
| 3. Promotes
lipophagy. |
|
|
|
|
|
|
| B, Herbal
monomers |
|
| Drug
ingredients | Mechanism of
action | Method | Model | Targets | Actions | (Refs.) |
|
| Geniposide | Inhibits the
PARP1/PI3K/AKT | In vivo
and | In vitro:
High-fat | PARP1, PI3K,
AKT | 1. Inhibited lipid
accumulation, apoptosis, | (123) |
|
| signaling
pathway | In
vitro | diet-fed
ApoE-/-mice. |
| and foam cell
marker expression. |
|
|
|
|
| In vivo:
VSMCs |
| 2. Accelerated
lipid droplet degradation. |
|
|
|
|
|
|
| 3. Suppressed
VSMC-derived foam cell |
|
|
|
|
|
|
| formation. |
|
|
Epigallocatechin | Activates the
CaMKKβ/AMPK | In
vitro | Primary bovine
aortic | CaMKKβ, AMPK, | 1. Promoted
autophagic flux and | (126–128) |
| gallate | pathway |
| endothelial
cells. | ULK1 and
LC3-II. | lipophagy. |
|
|
|
|
|
|
| 2. Inhibited lipid
droplet accumulation |
|
|
|
|
|
|
| and clearance of
ectopic lipid deposits. |
|
|
|
|
|
|
| 3. Ameliorated
lipotoxicity in vascular |
|
|
|
|
|
|
| endothelial
cells. |
|
| Berberine | 1. Inhibits the
PI3K/AKT/mTOR | In
vitro | Peritoneal
macrophages | SIRT1, TFEB,
LC3-II/ | 1. Induced
lipophagy. | (130,131) |
|
| signaling
pathway. |
|
| LC3-I and
Beclin1. | 2. Promoted lipid
droplet degradation. |
|
|
| 2. Activates the
SIRT1/TFEB |
|
|
| 3. Delayed cellular
apoptosis. |
|
|
| pathway. |
|
|
|
|
|
|
| 3. Upregulates the
NAD+/NADH |
|
|
|
|
|
|
| ratio. |
|
|
|
|
|
|
| C, Chinese
herbal formula |
|
| Drug
ingredients | Mechanism of
action | Method | Model | Targets | Actions | (Refs.) |
|
| Tianxiangdan | Suppresses the
PI3K/Akt/mTOR | In vivo | High-fat
diet-fed | PI3K, Akt,
mTOR, | 1. Activated
lipophagy. | (139) |
|
| pathway and
activates TFEB |
| ApoE-/-mice | TFEB, LC3II/I,
ULK1, | 2. Promoted lipid
droplet degradation. |
|
|
|
|
|
| ABCA1 and p62 | 3. Reduced foam
cell formation and |
|
|
|
|
|
|
| intra-plaque lipid
deposition. |
|
| Huayu Qutan | 1. Regulates the
mTORC1/TFEB | In vivo
and | In vivo:
ApoE-/-mice | mTORC1, TFEB, | 1. Activated
lipophagy. | (140) |
| Recipe | signaling
pathway. | in
vitro | In vitro:
RAW 264.7 | LC3-II/I, Beclin1
and | 2. Upregulated
ABCA1 and SCARB1. |
|
|
| 2. Inhibits the
PI3K/Akt/mTOR |
| cells | p62 | 3. Modulated serum
lipid profiles. |
|
|
| pathway and
activates TFEB. |
|
|
| 4. Reduced lipid
accumulation in |
|
|
|
|
|
|
| macrophages. |
|
|
|
|
|
|
| 5. Inhibited foam
cell formation. |
|
|
|
|
|
|
| 6. Promoted
autophagosome formation. |
|
|
|
|
|
|
| 7. Facilitated
lipid degradation in foam |
|
|
|
|
|
|
| cells via the
autophagic-lysosomal |
|
|
|
|
|
|
| pathway. |
|
| Gualou-Xiebai | 1. Downregulates
P2RY12, p62 | In vivo | High-fat
diet-fed | P2RY12, LC3II,
p62 | 1. Enhanced
lipophagy. | (141) |
| herb pair | and Plin2. |
| ApoE-/-mice | and PLIN2 | 2. Increased the
number of |
|
|
| 2. Upregulates
LC3-II protein |
|
|
|
autophagosomes. |
|
|
| expression |
|
|
| 3. Reduced foam
cell formation. |
|
Convergence of therapeutic
mechanisms
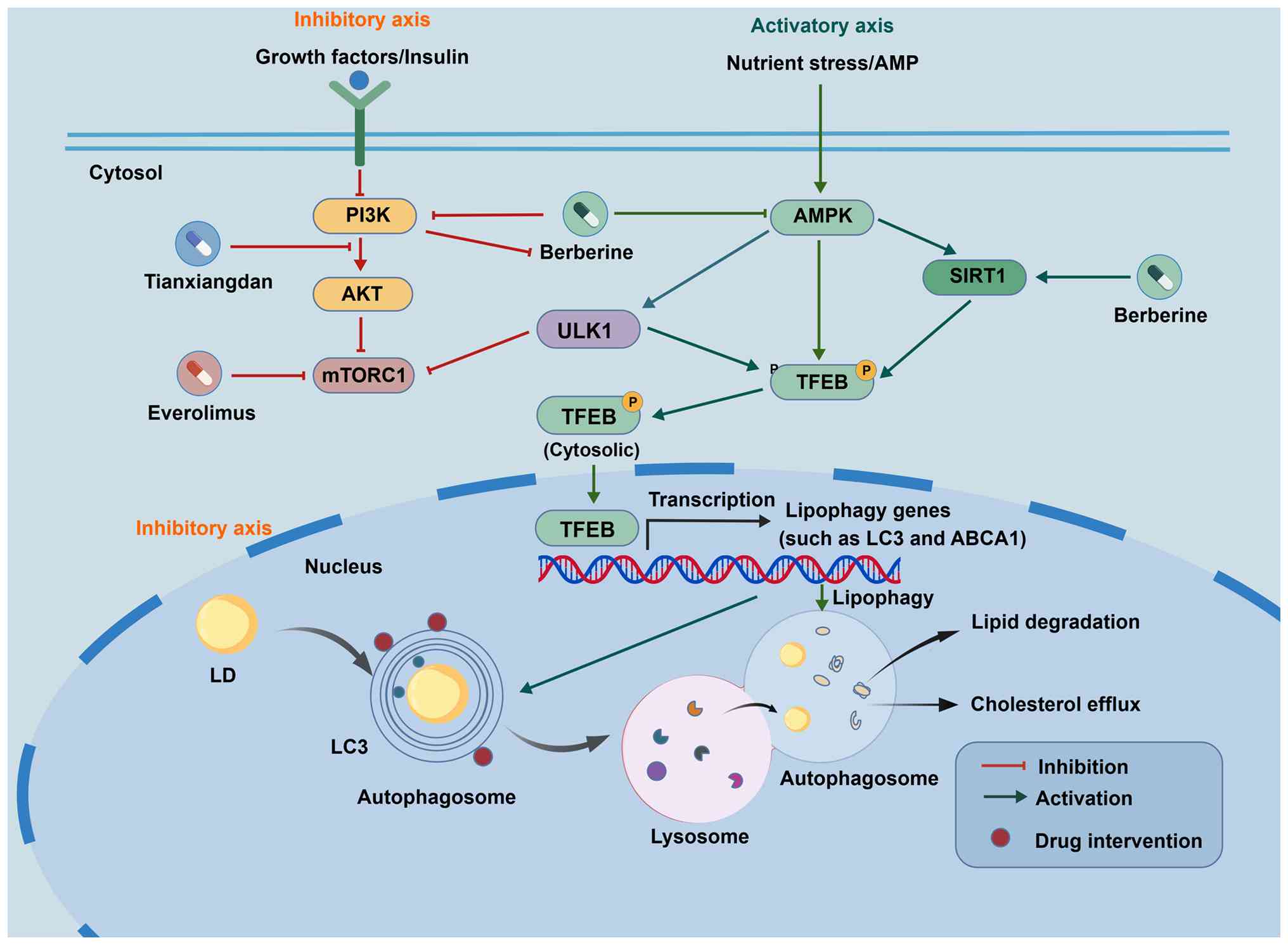
As illustrated in the regulatory network (Fig. 2), pharmacological agents modulate
lipophagy through two distinct but converging signaling axes.
First, in the inhibitory axis, agents such as everolimus and the
TCM formula Tianxiangdan function by inhibiting the PI3K/Akt/mTOR
pathway. This inhibition suppresses mTORC1 phosphorylation, thereby
relieving the suppression of ULK1/2 and TFEB, which subsequently
activates autophagy proteins and increases LD degradation. Second,
in the activating axis, natural compounds such as BBR and EGCG
predominantly target the AMPK-SIRT1 axis. BBR activates NAD+
biosynthesis, which enhances SIRT1 deacetylase activity and leads
to TFEB deacetylation, ultimately increasing the LC3-II/LC3-I
ratio. Similarly, EGCG activates the CaMKKβ/AMPK pathway, bypassing
mTOR and directly stimulating autophagic flux. These interventions
ultimately converge to upregulate the expression of cholesterol
efflux proteins (ABCA1) and reduce foam cell formation.
Discussion and future perspectives
AS pathological mechanisms are intricate, involving
a multitude of cell types and biomolecules. Lipophagy, a variant of
autophagy, has garnered notable attention as a focal point of
research. Lipophagy not only plays a pivotal role in modulating
inflammatory responses but also directly influences lipid
metabolism and cellular functions, contributing to AS initiation
and progression (6,14). Recent studies indicate that
lipophagy modulates macrophage functionality, either facilitating
or inhibiting lipid deposition within arterial walls and thus
impacting plaque stability and formation (68,98).
Synthesizing these multidimensional findings, our proposed
‘stage-dependent double-edged sword’ model challenges the
traditional paradigm that simply views autophagy activation as
universally beneficial. Instead, the present model underscores a
critical pathogenic threshold where adaptive lipid clearance
transitions into maladaptive lysosomal membrane permeabilization
and cell death (110).
Recognizing this threshold is an original insight that is necessary
in order to transition from broad, systemic autophagy inducers to
the precise, context-dependent interventions outlined in our
therapeutic framework.
To address the existing challenges in the field,
future research should prioritize several critical aspects. First,
it is imperative to develop more specific lipophagy modulators to
enhance therapeutic efficacy and minimize potential side effects.
Current agents often possess broad mechanisms of action, which can
result in inconsistent effectiveness and safety concerns (110,111). Therefore, precisely targeted
lipophagy drugs are required for ensuring therapeutic success. An
evaluation of regulatory networks revealed that not all signaling
axes hold equal therapeutic promise. While the upstream AMPK-mTORC1
axis acts as the primary cellular energy sensor, its broad
influence on global protein synthesis and cell growth makes it
susceptible to systemic side effects (114). Consequently, downstream
transcriptional and epigenetic regulators, specifically the TFEB
and SIRT1/6 axes, are more viable therapeutic targets (82,130). These downstream nodes are well
suited for a ‘precision strike’ strategy, which would directly
mobilize lysosomal biogenesis and autophagic flux without
disrupting fundamental cellular metabolism. Therefore, future drug
development should prioritize molecules that specifically calibrate
these terminal transcriptional effectors rather than broad upstream
kinases.
Second, there should be an emphasis on optimizing
combination therapies. Single-drug treatments are often inadequate
for addressing complex pathological conditions, whereas rationally
designed multi-drug regimens have the potential to improve
therapeutic outcomes and reduce the risk of drug resistance.
Through systematic clinical trials and personalized strategies,
drugs with complementary mechanisms can be strategically combined
to achieve enhanced efficacy. Furthermore, advancing translational
research is a key priority. While basic studies provide a
theoretical foundation for lipophagy-based therapies, further work
is needed to translate these discoveries into clinical practice.
The integration of multidisciplinary approaches, such as molecular
biology, is essential to facilitate this transition. From a
translational perspective, the molecular mechanism of lipophagy
aligns well with the TCM principle of resolving phlegm (Qutan).
Formulas such as Huoxue Qutan and Gualou-Xiebai promote lipophagy
by upregulating LC3-II and reducing p62/Plin2 levels. By
facilitating the lysosomal degradation of ‘turbid’ lipids (LDs)
(140,141), these herbal pairs effectively
reduce the necrotic core and stabilize plaques. This intricate
molecular crosstalk provides a compelling modern biological basis
for classical TCM therapies, highlighting their potential as
multi-targeted modulators for AS.
Lipophagy has been established as a critical process
in AS development and progression that is characterized by complex
and dual regulatory mechanisms. Future research should aim to
elucidate the specific functions of lipophagy and its interactions
with other biological processes. The therapeutic potential of
targeting lipophagy for clinical management warrants further
scientifically rigorous research to facilitate the development of
novel therapeutic strategies and provide new approaches for AS
prevention and treatment.
Acknowledgements
Not applicable.
Funding
The present review was supported by the National Natural Science
Foundation of China (grant no. 81573919) and the Henan Provincial
Key Research and Development Program (grant no. 221111310500).
Availability of data and materials
Not applicable.
Authors' contributions
SN drafted the manuscript and prepared the figures.
CP, XM, JJ and FZ conducted the literature search and contributed
to the writing and editing of the manuscript. YC proposed the
research design, supervised the study and critically revised the
manuscript for important intellectual content. Data authentication
is not applicable. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, artificial
intelligence tools were used to improve the readability and
language of the manuscript. Subsequently, the authors thoroughly
reviewed and edited the content produced by the artificial
intelligence tools as necessary, and the manuscript was further
refined by professional native-speaking editors from LetPub. The
authors take full responsibility for the integrity and ultimate
content of the present manuscript.
References
|
1
|
Libby P, Buring JE, Badimon L, Hansson JK,
Deanfield J, Bittencourt SM, Tokgözoğlu L and Lewis EF:
Atherosclerosis. Nat Rev Dis Primers. 5:562019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Filali-Mouncef Y, Hunter C, Roccio F,
Zagkou S, Dupont N, Primard C, Proikas-Cezanne T and Reggiori F:
The ménage à trois of autophagy, lipid droplets and liver disease.
Autophagy. 18:50–72. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chistiakov DA, Orekhov AN and Bobryshev
YV: LOX-1-Mediated effects on vascular cells in atherosclerosis.
Cell Physiol Biochem. 38:1851–1859. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baumer Y, McCurdy S, Weatherby TM, Mehta
NN, Halbherr S, Halbherr P, Yamazaki N and Boisvert WA:
Hyperlipidemia-induced cholesterol crystal production by
endothelial cells promotes atherogenesis. Nat Commun. 8:11292017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Giannotti KC, Weinert S, Viana MN, Leiguez
E, Araujo TLS, Laurindo FRM, Lomonte B, Braun-Dullaeus R and
Teixeira C: A secreted phospholipase A2 induces formation of smooth
muscle foam cells which transdifferentiate to Macrophage-like
state. Molecules. 24:32442019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Robichaud S, Fairman G, Vijithakumar V,
Mak E, Cook DP, Pelletier AR, Huard S, Vanderhyden BC, Figeys D,
Lavallée-Adam M, et al: Identification of novel lipid droplet
factors that regulate lipophagy and cholesterol efflux in
macrophage foam cells. Autophagy. 17:3671–3689. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pu M, Zheng W, Zhang H, Wan W, Peng C,
Chen X, Liu X, Xu Z, Zhou T, Sun Q, et al: ORP8 acts as a lipophagy
receptor to mediate lipid droplet turnover. Protein Cell.
14:653–667. 2023.PubMed/NCBI
|
|
8
|
Chung J, Park J, Lai ZW, Lambert TJ,
Richards RC, Zhang J, Walther TC and Farese RV Jr: The Troyer
syndrome protein spartin mediates selective autophagy of lipid
droplets. Nat Cell Biol. 25:1101–1110. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jin Y, Zhang L, Huang X, Ma Y, Liu J,
Zhang H and Li X: Role of inhibition of cellular foaming by
lipophagy in atherosclerosis. Chin J Pathophysiol. 40:564–571.
2024.
|
|
10
|
Yang M, Zhang Y and Ren J: Autophagic
regulation of lipid homeostasis in cardiometabolic syndrome. Front
Cardiovasc Med. 5:382018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang S, Peng X, Yang S, Li X, Huang M,
Wei S, Liu J, He G, Zheng H, Yang L, et al: The regulation,
function, and role of lipophagy, a form of selective autophagy, in
metabolic disorders. Cell Death Dis. 13:1322022. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu K and Czaja MJ: Regulation of lipid
stores and metabolism by lipophagy. Cell Death Differ. 20:3–11.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ward C, Martinez-Lopez N, Otten EG,
Carroll B, Maetzel D, Singh R, Sarkar S and Korolchuk VI:
Autophagy, lipophagy and lysosomal lipid storage disorders. Biochim
Biophys Acta. 1861:269–284. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ouimet M, Franklin V, Mak E, Liao X, Tabas
I and Marcel YL: Autophagy regulates cholesterol efflux from
macrophage foam cells via lysosomal acid lipase. Cell Metab.
13:655–667. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Seok S, Fu T, Choi SE, Li Y, Zhu R, Kumar
S, Sun X, Yoon G, Kang Y, Zhong W, et al: Transcriptional
regulation of autophagy by an FXR-CREB axis. Nature. 108–111. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Z, Meng X, Lu R, Meng X, Li S, Wang Y,
Liu X, Liu X and Liu J: PBX1 improves cognition and reduces
Amyloid-β pathology in APP/PS1 mice by transcriptionally activating
the CRTC2-CREB pathway. Aging Cell. 25:e703112026. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paul S, Chatterjee A, Das K, Ray A, Basu
A, Mukhopadhyay S and Sen P: Thrombin confers chemotherapeutic
resistance by promoting transcriptional induction and
post-translational stabilization of pro-survival MCL1 in TNBC. J
Biol Chem. 301:1080252025. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zou L, Hong D, Li K and Jiang B:
Salt-inducible kinase 2 (SIK2) inhibitor ARN-3236 attenuates
bleomycin-induced pulmonary fibrosis in mice. BMC Pulm Med.
22:1402022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wei N, Feng W, Pan M, Xu X, Zheng S and
Jin H: Curcumol ameliorates high-fat diet-induced hepatic fibrosis
via dual regulation of FXR-CREB and Rab18-mediated hepatic stellate
cell lipophagy. Biochim Biophys Acta Mol Cell Biol Lipids.
1871:1597102026. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee JM, Wagner M, Xiao R, Kim KH, Feng D,
Lazar MA and Moore DD: Nutrient-sensing nuclear receptors
coordinate autophagy. Nature. 516:112–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao T, Wu K, Hogstrand C, Xu YH, Chen GH,
Wei CC and Luo Z: Lipophagy mediated carbohydrate-induced changes
of lipid metabolism via oxidative stress, endoplasmic reticulum
(ER) stress and ChREBP/PPARγ pathways. Cell Mol Life Sci.
77:1987–2003. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen K, Yuan R, Zhang Y, Geng S and Li L:
Tollip deficiency alters atherosclerosis and steatosis by
disrupting lipophagy. J Am Heart Assoc. 6:e0040782017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schwerbel K, Kamitz A, Krahmer N, Hallahan
N, Jähnert M, Gottmann P, Lebek S, Schallschmidt T, Arends D,
Schumacher F, et al: Immunity-related GTPase induces lipophagy to
prevent excess hepatic lipid accumulation. J Hepatol. 73:771–782.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li W, Sultana N, Siraj N, Ward LJ, Pawlik
M, Levy E, Jovinge S, Bengtsson E and Yuan XM: Autophagy
dysfunction and regulatory cystatin C in macrophage death of
atherosclerosis. J Cell Mol Med. 20:1664–1672. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sergin I, Evans TD, Zhang X, Bhattacharya
S, Stokes CJ, Song E, Ali S, Dehestani B, Holloway KB, Micevych PS,
et al: Exploiting macrophage autophagy-lysosomal biogenesis as a
therapy for atherosclerosis. Nat Commun. 8:157502017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pi H, Wang Z, Liu M, Deng P, Yu Z, Zhou Z
and Gao F: SCD1 activation impedes foam cell formation by inducing
lipophagy in oxLDL-treated human vascular smooth muscle cells. J
Cell Mol Med. 23:5259–5269. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fu Y, Deng Y, Zhang J, Chua SL and Khoo
BL: Biofilms exacerbate atherogenesis through macrophage-induced
inflammatory responses in a fibrous plaque microsystem model. Acta
Biomater. 168:333–345. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shroff A and Nazarko TY: SQSTM1, lipid
droplets and current state of their lipophagy affairs. Autophagy.
19:720–723. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Runsala M, Kuokkanen E, Uski E, Šuštar V,
Balci MÖ, Rajala J, Paavola V and Mattila PK: The Small GTPase Rab7
regulates antigen processing in B cells in a possible interplay
with autophagy machinery. Cells. 12:25662023. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kamerkar S, Singh J, Tripathy S, Bhonsle
H, Kumar M and Mallik R: Metabolic and immune-sensitive contacts
between lipid droplets and endoplasmic reticulum reconstituted in
vitro. Proc Natl Acad Sci USA. 119:e22005131192022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Zhao H, Li X, Wang Q, Yan M, Zhang
H, Zhao T, Zhang N, Zhang P, Peng L and Li P: Formononetin
alleviates hepatic steatosis by facilitating TFEB-mediated lysosome
biogenesis and lipophagy. J Nutr Biochem. 73:1082142019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoo J, Jeong IK, Ahn KJ, Chung HY and
Hwang YC: Fenofibrate, a PPARα agonist, reduces hepatic fat
accumulation through the upregulation of TFEB-mediated lipophagy.
Metabolism. 120:1547982021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sinha RA, Rajak S, Singh BK and Yen PM:
Hepatic lipid catabolism via PPARα-lysosomal crosstalk. Int J Mol
Sci. 21:23912020. View Article : Google Scholar
|
|
34
|
Kim KH, Oprescu SN, Snyder MM, Kim A, Jia
Z, Yue F and Kuang S: PRMT5 mediates FoxO1 methylation and
subcellular localization to regulate lipophagy in myogenic
progenitors. Cell Rep. 42:1133292023. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang T, Cheng Z, Zhao R, Cheng J, Ren H,
Zhang P, Liu P, Hao Q, Zhang Q, Yu X, et al: Sirt6 enhances
macrophage lipophagy and improves lipid metabolism disorder by
regulating the Wnt1/β-catenin pathway in atherosclerosis. Lipids
Health Dis. 22:1562023. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li M, Wang Z, Wang P, Li H and Yang L:
TFEB: A emerging regulator in lipid homeostasis for
atherosclerosis. Front Physiol. 12:6399202021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bao Y, Cao Y and Wu H: Research progress
on the role of lipophagy in atherosclerosis and traditional Chinese
medicine intervention. Chin J Arteriosclerosis. 30:753–763.
2022.
|
|
38
|
Blagov AV, Churov AV, Golovyuk AL, Lee AA,
Kashtalap VV, Sukhorukov VN and Orekhov AN: The role of metabolic
disorders in the development of atherosclerosis. Cell Mol Biol
(Noisy-le-Grand). 70:148–155. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Garcia-Macia M, Santos-Ledo A, Leslie J,
Paish HL, Collins AL, Scott RS, Watson A, Burgoyne RA, White S,
French J, et al: A mammalian target of Rapamycin-Perilipin 3
(mTORC1-Plin3) pathway is essential to activate lipophagy and
protects against hepatosteatosis. Hepatology. 74:3441–3459. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yano K, Yamaguchi K, Seko Y, Okishio S,
Ishiba H, Tochiki N, Takahashi A, Kataoka S, Okuda K, Liu Y, et al:
Hepatocyte-specific fibroblast growth factor 21 overexpression
ameliorates high-fat diet-induced obesity and liver steatosis in
mice. Lab Invest. 102:281–289. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zachari M and Ganley IG: The mammalian
ULK1 complex and autophagy initiation. Essays Biochem. 61:585–596.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim YC and Guan KL: mTOR: A pharmacologic
target for autophagy regulation. J Clin Invest. 125:25–32. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Park JM, Jung CH, Seo M, Otto NM, Grunwald
D, Kim KH, Moriarity B, Kim YM, Starker C, Nho RS, et al: The ULK1
complex mediates MTORC1 signaling to the autophagy initiation
machinery via binding and phosphorylating ATG14. Autophagy.
12:547–564. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Roczniak-Ferguson A, Petit CS, Froehlich
F, Qian S, Ky J, Angarola B, Walther TC and Ferguson SM: The
transcription factor TFEB links mTORC1 signaling to transcriptional
control of lysosome homeostasis. Sci Signal. 5:ra422012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gonzalez A, Hall MN, Lin SC and Hardie DG:
AMPK and TOR: The yin and yang of cellular nutrient sensing and
growth control. Cell Metab. 31:472–492. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang S, Hong F, Ma C and Yang S: Hepatic
lipid metabolism disorder and atherosclerosis. Endocr Metab Immune
Disord Drug Targets. 22:590–600. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang H, Xia P, Yang Z, Liu J, Zhu Y,
Huang Z, Zhang Z and Yuan Y: Cullin-associated and
neddylation-dissociated 1 regulate reprogramming of lipid
metabolism through SKP1-Cullin-1-F-boxFBXO11-mediated heterogeneous
nuclear ribonucleoprotein A2/B1 ubiquitination and promote
hepatocellular carcinoma. Clin Transl Med. 13:e14432023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang Y, Yang J, Zhang J and Xiang Z: The
role and function of transcription factor EB in atherosclerosis: A
comprehensive review. Chin J Hypertension. 30:1142–1146. 2022.
|
|
49
|
Wang MY, Li EW, Gao G, Fu ZX, Zhang XW,
Wang H, Wang P, Zhang ZQ, Xu JY and Xie ZS: Zexie decoction
regulates Akt/TFEB signaling pathway to promote lipophagy in
hepatocytes. Zhongguo Zhong Yao Za Zhi. 47:6183–6190. 2022.(In
Chinese). PubMed/NCBI
|
|
50
|
Zhao X: Effects of hepatic lysosome
function on atherosclerotic mice with different lipemia. Wuhan
University; 2021
|
|
51
|
Acharya A and Demetriades C: mTORC1
activity licenses its own release from the lysosomal surface. Mol
Cell. 84:4385–4400.e7. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu Q, Lin R, Jiang T, Deng L, Wu Y, Yuan
Q, Qi X, Mu P, Jiang J, Deng Y and Wen J: Enhanced SIRT1-TFEB
interaction promotes lysosome biogenesis and autophagy by reducing
TFEB acetylation: Revealing the enterotoxicity disparity of
deoxynivalenol and T-2 toxin. J Agric Food Chem. 73:12993–13005.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Negoita F, Fraguas Bringas C, Hellberg K,
Luda KM, Liu H, Li Z, Cuenco J, Zhao JF, Sathe G, Ganley IG, et al:
AMPK promotes TFEB transcriptional activity through
dephosphorylation at both MTORC1-dependent and -independent sites.
Autophagy. 1–15. 2026.doi: 10.1080/15548627.2026.2629720 (Epub
ahead of print).
|
|
54
|
You Y and Liang W: SIRT1 and SIRT6: The
role in aging-related diseases. Biochim Biophys Acta Mol Basis Dis.
1869:1668152023. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Herranz D, Muñoz-Martin M, Cañamero M,
Mulero F, Martinez-Pastor B, Fernandez-Capetillo O and Serrano M:
Sirt1 improves healthy ageing and protects from metabolic
syndrome-associated cancer. Nat Commun. 1:32010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang Y, Liu T, Cai Y, Liu W and Guo J:
SIRT6′s function in controlling the metabolism of lipids and
glucose in diabetic nephropathy. Front Endocrinol (Lausanne).
14:12447052023. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang T: Sirt6 enhances macrophage
lipophagy and improves lipid metabolism disorder to increase
atherosclerotic plaque stability. Air Force Medical University;
2020
|
|
58
|
Li B, Xin Z, Gao S, Li Y, Guo S, Fu Y, Xu
R, Wang D, Cheng J, Liu L, et al: SIRT6-regulated macrophage
efferocytosis epigenetically controls inflammation resolution of
diabetic periodontitis. Theranostics. 13:231–249. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zi Y, Yi-An Y, Bing J, Yan L, Jing T,
Chun-Yu G, Fan P, Hao L, Jia-Ni T, Han-Jin H, et al: Sirt6-induced
autophagy restricted TREM-1-mediated pyroptosis in ox-LDL-treated
endothelial cells: Relevance to prognostication of patients with
acute myocardial infarction. Cell Death Discov. 5:882019.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang Y, Zou Y, Wang X, Liu J and Pang Y:
PLINs: Lipid droplet-associated proteins involved in lipolysis. J
Northeast Agricultural Univ. 52:82–89. 962021.
|
|
61
|
Hsieh K, Lee YK, Londos C, Raaka BM, Dalen
KT and Kimmel AR: Perilipin family members preferentially sequester
to either triacylglycerol-specific or cholesteryl-ester-specific
intracellular lipid storage droplets. J Cell Sci. 125:4067–4076.
2012.PubMed/NCBI
|
|
62
|
Lizaso A, Tan KT and Lee YH: β-adrenergic
receptor-stimulated lipolysis requires the RAB7-mediated
autolysosomal lipid degradation. Autophagy. 9:1228–1243. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kaushik S and Cuervo AM: Degradation of
lipid droplet-associated proteins by chaperone-mediated autophagy
facilitates lipolysis. Nat Cell Biol. 17:759–770. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cho KY, Miyoshi H, Nakamura A, Greenberg
AS and Atsumi T: Lipid droplet protein PLIN1 regulates inflammatory
polarity in human macrophages and is involved - in atherosclerotic
plaque development by promoting stable lipid storage. J Atheroscler
Thromb. 30:170–181. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ouyang Y, Hu M, Zhang Y, Lin H, Tang W,
Peng M, He P and Ouyang X: The role of gut microbiota and its
metabolites in the development of atherosclerosis. Chemistry of
Life. 45:43–54. 2025.(In Chinese).
|
|
66
|
Tang WH, Kitai T and Hazen SL: Gut
microbiota in cardiovascular health and disease. Circ Res.
120:1183–1196. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Gu Y, Zhang Y, Li M, Huang Z, Jiang J,
Chen Y, Chen J, Jia Y, Zhang L and Zhou F: Ferulic acid ameliorates
atherosclerotic injury by modulating gut microbiota and lipid
metabolism. Front Pharmacol. 12:6213392021. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Jeong SJ, Lee MN and Oh GT: The role of
macrophage lipophagy in reverse cholesterol transport. Endocrinol
Metab (Seoul). 32:41–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu Q, Wang YM and Gu HF: Lipophagy in
atherosclerosis. Clin Chim Acta. 511:208–214. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Björkegren JLM and Lusis AJ:
Atherosclerosis: Recent developments. Cell. 185:1630–1645. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Guo X, Hou P, Zhang S, Xu Q, Zhou M, Tang
W, Jin F, Zhang B, Guo Z, Zhao X, et al: Enhancer-associated
LncRNA-ITGA2 promotes vascular remodeling through ITGA2. Circ Res.
136:1610–1628. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Bennett MR, Sinha S and Owens GK: Vascular
smooth muscle cells in atherosclerosis. Circ Res. 118:692–702.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zeng C, Peng Z, Huang S, Xu Z, Peng Z, Wu
Z, Lei J, Zhang X, Qin J, Ye K, et al: Metal-organic
framework-based nanoplatforms for synergistic anti-atherosclerosis
therapy by regulating the PI3K/AKT/MSR1 pathway in macrophages.
Nanoscale. 17:3071–3085. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Liu F, Shan S, Li H and Li Z: Treatment of
peroxidase derived from foxtail millet bran attenuates
atherosclerosis by inhibition of CD36 and STAT3 in vitro and in
vivo. J Agric Food Chem. 68:1276–1285. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Liu F, Shan S, Li H, Shi J, Yang R and Li
Z: Millet shell polyphenols ameliorate atherosclerosis development
by suppressing foam cell formation. J Nutr Biochem. 115:1092712023.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Nahapetyan H, Moulis M, Grousset E,
Faccini J, Grazide MH, Mucher E, Elbaz M, Martinet W and Vindis C:
Altered mitochondrial quality control in Atg7-deficient VSMCs
promotes enhanced apoptosis and is linked to unstable
atherosclerotic plaque phenotype. Cell Death Dis. 10:1192019.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ravi S, Martin LC, Krishnan M, Kumaresan
M, Manikandan B and Ramar M: Interactions between macrophage
membrane and lipid mediators during cardiovascular diseases with
the implications of scavenger receptors. Chem Phys Lipids.
258:1053622024. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Qi X, Zhang Y, Li J, Hou D and Xiang Y:
Effect of PGC-1α on proliferation, migration, and
transdifferentiation of rat vascular smooth muscle cells induced by
high glucose. J Biomed Biotechnol. 2012:7564262012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhang Q, Wen XH, Tang SL, Zhao ZW and Tang
CK: Role and therapeutic potential of gelsolin in atherosclerosis.
J Mol Cell Cardiol. 178:59–67. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Gimbrone MA Jr and Garcia-Cardena G:
Endothelial cell dysfunction and the pathobiology of
atherosclerosis. Circ Res. 118:620–636. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Pu Y, Cheng CK, Zhang H, Luo JY, Wang L,
Tomlinson B and Huang Y: Molecular mechanisms and therapeutic
perspectives of peroxisome proliferator-activated receptor alpha
agonists in cardiovascular health and disease. Med Res Rev.
43:2086–2114. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Chen Z, Ouyang C, Zhang H, Gu Y, Deng Y,
Du C, Cui C, Li S, Wang W, Kong W, et al: Vascular smooth muscle
cell-derived hydrogen sulfide promotes atherosclerotic plaque
stability via TFEB (transcription factor EB)-mediated autophagy.
Autophagy. 18:2270–2287. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Goncalves I, Pan M, Singh P, Wang W, Zhao
J, Dib L, Sundius L, Persson A, Gialeli C, Fountas P, et al:
Spatial transcriptomics reveals a key role of fibroblast-like
vascular smooth muscle cells in human atherosclerotic cell
crosstalk and stability. Eur Heart J. ehaf10912026.doi:
10.1093/eurheartj/ehaf1091 (Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhao L, Zhao L, Liu D, Huang F, Peng Q, Lu
J, Zhou J, Zheng S and Liu X: Vascular smooth muscle cells: A
therapeutic target in atherosclerosis. Rev Cardiovasc Med.
26:282402025. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Tang HY, Chen AQ, Zhang H, Gao XF, Kong XQ
and Zhang JJ: Vascular smooth muscle cells phenotypic switching in
cardiovascular diseases. Cells. 11:40602022. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Zhang J, Chang J, Chen V, Beg MA, Huang W,
Vick L, Wang Y, Zhang H, Yttre E, Gupta A, et al: Oxidized LDL
stimulates PKM2-mediated mtROS production and phagocytosis. J Lipid
Res. 66:1008092025. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Bardin M, Pawelzik SC, Lagrange J, Mahdi
A, Arnardottir H, Regnault V, Fève B, Lacolley P, Michel JB,
Mercier N and Bäck M: The resolvin D2-GPR18 axis is expressed in
human coronary atherosclerosis and transduces atheroprotection in
apolipoprotein E deficient mice. Biochem Pharmacol. 201:1150752022.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Pirillo A, Norata GD and Catapano AL:
LOX-1, OxLDL, and atherosclerosis. Mediators Inflamm.
2013:1527862013. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Daub K, Seizer P, Stellos K, Krämer BF,
Bigalke B, Schaller M, Fateh-Moghadam S, Gawaz M and Lindemann S:
Oxidized LDL-activated platelets induce vascular inflammation.
Semin Thromb Hemost. 36:146–156. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Hammoutene A, Biquard L, Lasselin J,
Kheloufi M, Tanguy M, Vion AC, Mérian J, Colnot N, Loyer X, Tedgui
A, et al: A defect in endothelial autophagy occurs in patients with
non-alcoholic steatohepatitis and promotes inflammation and
fibrosis. J Hepatol. 72:528–538. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Sotler T and Sebestjen M: PCSK9 as an
atherothrombotic risk factor. Int J Mol Sci. 24:19662023.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
El Bounkari O, Zan C, Yang B, Ebert S,
Wagner J, Bugar E, Kramer N, Bourilhon P, Kontos C, Zarwel M, et
al: An atypical atherogenic chemokine that promotes advanced
atherosclerosis and hepatic lipogenesis. Nat Commun. 16:22972025.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Jiang L, Bai D, Yang Y, Wu Y, Chen Y, Wang
M, Wang W, Wang H, Xiong Y, Zhu S, et al: Naringenin attenuates
Ox-LDL-induced Injury in HUVECs by regulating lipophagy partly
through the SOCE pathway. Curr Pharm Des. 31:2025–2037. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Wang J, Peng M, Gao J and Xu G:
Atherosclerotic vascular remodeling induced by phenotypic switching
of vascular smooth muscle cell. Chin J Arteriosclerosis.
33:269–276. 2025.
|
|
95
|
Zhuang W, Yang Y, Li H and Liang J:
Research advance of Nrf2 on atherosclerosis by regulating vascular
smooth muscle cell. Zhejiang Da Xue Xue Bao Yi Xue Ban. 50:390–395.
2021.PubMed/NCBI
|
|
96
|
Boström P, Magnusson B, Svensson PA,
Wiklund O, Borén J, Carlsson LM, Ståhlman M, Olofsson SO and Hultén
LM: Hypoxia converts human macrophages into triglyceride-loaded
foam cells. Arterioscler Thromb Vasc Biol. 26:1871–1876. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Yang R, Zhu Y, Wang Y, Ma W, Wang X, Han X
and Liu N: Recent progress in autophagy and vascular calcification.
J China Pharmaceutical Univ. 49:401–406. 2018.(In Chinese).
|
|
98
|
Wang Z, Nong T and Zheng J: Biological
connotation of Disease-stage-state of hypertension in concept of
State-target differentiation and treatment from perspective of
lipophagy. Chin J Exp Tradit Med Formulae. 30:193–201. 2024.
|
|
99
|
Chen W, Jin T, Xie Y, Zhong C, Gao H,
Zhang L, Ju J, Cheng T, Li M, Wang H, et al: Berberine partially
ameliorates cardiolipotoxicity in diabetic cardiomyopathy by
modulating SIRT3-mediated lipophagy to remodel lipid droplets
homeostasis. Br J Pharmacol. 182:5038–5056. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Hou Y and Zhao Y: The mechanisms of action
and clinical application progress of autophagy activators. J Sun
Yat-Sen Univ (Med Sci). 46:371–383. 2025.
|
|
101
|
Sabers CJ, Martin MM, Brunn GJ, Williams
JM, Dumont FJ, Wiederrecht G and Abraham RT: Isolation of a protein
target of the FKBP12-rapamycin complex in mammalian cells. J Biol
Chem. 270:815–822. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zhang Y, Li T, Pan M, Wang W, Huang W,
Yuan Y, Xie Z, Chen Y, Peng J, Li X and Meng Y: SIRT1 prevents
cigarette smoking-induced lung fibroblasts activation by regulating
mitochondrial oxidative stress and lipid metabolism. J Transl Med.
20:2222022. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yang Y, Bai D, Jiang L, Chen Y, Wang M,
Wang W, Wang H, He Q, Bu G, Long J and Yuan D: Stilbene glycosides
alleviate atherosclerosis partly by promoting lipophagy of
dendritic cells. Int Immunopharmacol. 143:1132232024. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Cheng H, Xu B, Zhang L, Wang Y, Chen M and
Chen S: Bortezomib alleviates antibody-mediated rejection in kidney
transplantation by facilitating Atg5 expression. J Cell Mol Med.
25:10939–10949. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Widgerow AD, Casas LA, Claytor B, Zeidler
KR and Shafiq F: Topical peptide technology and the scarring
spectrum: Linking mechanistic pathways to internal scarring
(Fibrous Banding) and external scarring (Incisional) and outcomes
in postsurgical healing. Aesthet Surg J Open Forum. 8:ojaf1692026.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Ferri N, Ruscica M, Fazio S and Corsini A:
Correction: Ferri et al. Low-density lipoprotein
cholesterol-lowering drugs: A narrative review. J. Clin. Med.
2024.13:943J Clin Med. 13:45822024. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Beshir SA, Hussain N, Elnor AA and Said
ASA: Umbrella review on non-statin lipid-lowering therapy. J
Cardiovasc Pharmacol Ther. 26:437–452. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Shao B, Liu C, Su D and Miao C: Research
progress in the roles of macrophage autophagy in atherosclerosis
and related drugs. Prog Pharmaceutical Sci. 41:490–496. 2017.
|
|
109
|
Watanabe T, Kotani J, Murata Y, Seguchi O,
Yanase M and Nakatani T: Tissue characterization of progressive
cardiac allograft vasculopathy in patients with everolimus therapy
compared with donor-transmitted atherosclerosis assessed using
serial intravascular imaging: A case report. Transplant Proc.
46:2456–2461. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Beutner F, Brendel D, Teupser D, Sass K,
Baber R, Mueller M, Ceglarek U and Thiery J: Effect of everolimus
on pre-existing atherosclerosis in LDL-receptor deficient mice.
Atherosclerosis. 222:337–343. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zhang Y, Wang Y, Bao C, Xu Y, Shen H, Chen
J, Yan J and Chen Y: Metformin interacts with AMPK through binding
to γ subunit. Mol Cell Biochem. 368:69–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Song Q, Bi L, Zhang J, Ren R and Zhao X:
Research progress in the mechanism of atorvastatin on No-reflow of
coronary artery. Herald Med. 40:641–644. 2021.
|
|
113
|
Li Y and Liu X: Metformin improves the
abnormal proliferation of vascular smooth muscle cells and
atherosclerosis in atherosclerotic mice by regulating AMPK/ERK
pathway. J Univ South China (Med Ed). 51:32–36. 2023.
|
|
114
|
Zheng S, Du Y, Ye Q, Zha K and Feng J:
Atorvastatin enhances foam cell lipophagy and promotes cholesterol
efflux through the AMP-activated protein Kinase/Mammalian target of
rapamycin pathway. J Cardiovasc Pharmacol. 77:508–518. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zhao L, Yu H and Tian W: Research progress
of traditional Chinese medicine monomer regulating autopha of
vascular endothelial cells and interfering atherosclersi. Chin Arch
Tradit Chin Med. 39:117–120. 2021.
|
|
116
|
Liu L, Wu Q, Chen Y, Gu G, Gao R, Peng B,
Wang Y, Li A, Guo J, Xu X, et al: Updated pharmacological effects,
molecular mechanisms, and therapeutic potential of natural product
geniposide. Molecules. 27:33192022. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Zhang L, Li J, Kou Y, Shen L, Wang H, Wang
Y, Ma R, Wu T, Yang X, Gu Y and Yi L: Mechanisms and treatment of
atherosclerosis: Focus on macrophages. Front Immunol.
15:14903872024. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Sadeghi M, Khayati S, Dehnavi S, Almahmeed
W, Sukhorukovi VN and Sahebkar A: Regulatory impact of statins on
macrophage polarization: Mechanistic and therapeutic implications.
J Pharm Pharmacol. 76:763–775. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Wiggins BS, Backes JM and Hilleman D:
Statin-associated muscle symptoms-A review: Individualizing the
approach to optimize care. Pharmacotherapy. 42:428–438. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Al-Makhamreh HK, Toubasi AA, Obaid YY and
Albustanji FH: Significance of Statin-associated muscle symptoms
and its impact on patients adherence and outcomes. J Cardiovasc
Pharmacol. 81:185–191. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Gerber T, Nunes A, Moreira BR and
Maraschin M: Yerba mate (Ilex paraguariensis A. St.-Hil.) for new
therapeutic and nutraceutical interventions: A review of patents
issued in the last 20 years (2000–2020). Phytother Res. 37:527–548.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Ran D, Hong W, Yan W and Mengdie W:
Properties and molecular mechanisms underlying geniposide-mediated
therapeutic effects in chronic inflammatory diseases. J
Ethnopharmacol. 273:1139582021. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Lin J, Wang X, Gu M, Chen Y, Xu J, Chau
NV, Li J, Ji X, Chu Q, Qing L and Wu W: Geniposide ameliorates
atherosclerosis by restoring lipophagy via suppressing
PARP1/PI3K/AKT signaling pathway. Phytomedicine. 129:1556172024.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Kim HS, Montana V, Jang HJ, Parpura V and
Kim JA: Epigallocatechin gallate (EGCG) stimulates autophagy in
vascular endothelial cells: A potential role for reducing lipid
accumulation. J Biol Chem. 288:22693–22705. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Li S, Yu H and Tian W: Research progress
of effect of traditional Chinese medicine in prevention and
treatment of atherosclerosis based on signal pathway related to
inflammatory response. Chin J Exp Tradit Med Formulae. 26:180–186.
2020.
|
|
126
|
Li Y, Xue X, Yu L, Qian J, Li X, Tian M,
Yang J, Deng R, Lu C, Xiao C and Liu Y: Recombinant high-density
lipoprotein targeted delivery of celastrol to promote foam cells
lipophagy against early atherosclerosis. J Nanobiotechnology.
23:2372025. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Zhao X, Li K, Liu Q, Jiang M and Cao Y:
Oral exposure to SiO2 nanoparticles promotes foam cell areas in
aortic sinus of ApoE-/-Mice and epigallocatechin gallate attenuates
the effects via metabolic restoration. Cardiovasc Toxicol.
25:1837–1849. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Wang L, Pan Q and Tang C: Mechanisms
underlying the Anti-atherosclerotic effects of EGCG. Curr Mol Med.
25:1327–1335. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Xing L, Zhou X, Li AH, Li HJ, He CX, Qin
W, Zhao D, Li PQ, Zhu L and Cao HL: Atheroprotective effects and
molecular mechanism of berberine. Front Mol Biosci. 8:7626732021.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zheng Y, Kou J, Wang P, Ye T, Wang Z, Gao
Z, Cong L, Li M, Dong B, Yang W, et al: Berberine-induced TFEB
deacetylation by SIRT1 promotes autophagy in peritoneal
macrophages. Aging (Albany NY). 13:7096–7119. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Zhu L, Xu JJ, Li HD, Li JJ, Cheng M, Niu
XN, Jia PC, Liu JY, Huang C, Lv XW and Li J: Berberine ameliorates
abnormal lipid metabolism via the adenosine Monophosphate-activated
protein Kinase/Sirtuin 1 pathway in alcohol-related liver disease.
Lab Invest. 103:1000412023. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Xu Y and Wan W: Acetylation in the
regulation of autophagy. Autophagy. 19:379–387. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Wang W, Li H, Shi Y, Zhou J, Khan GJ, Zhu
J, Liu F, Duan H, Li L and Zhai K: Targeted intervention of natural
medicinal active ingredients and traditional Chinese medicine on
epigenetic modification: Possible strategies for prevention and
treatment of atherosclerosis. Phytomedicine. 122:1551392024.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Zhai K, Deng L, Wu Y, Li H, Zhou J, Shi Y,
Jia J, Wang W, Nian S, Jilany Khan G, et al: Extracellular
vesicle-derived miR-146a as a novel crosstalk mechanism for
high-fat induced atherosclerosis by targeting SMAD4. J Adv Res.
73:729–741. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Duan H, Li H, Feng J, Zhoua J, Shi Y, Wang
L, Dong Z, Zhai K, Hu K and Wei Z: Isodon Suzhouensis extract
interferes with miRNA expression profile for prevention and
treatment of chronic obstructive pneumonia disease. Food Sci Hum
Wellness. 14:92502502025. View Article : Google Scholar
|
|
136
|
Zhi W, Liu Y, Wang X and Zhang H: Recent
advances of traditional Chinese medicine for the prevention and
treatment of atherosclerosis. J Ethnopharmacol. 301:1157492023.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Tan M, Wang J, Chen Z and Xie X: Exploring
global research trends in Chinese medicine for atherosclerosis: A
bibliometric study 2012–2023. Front Cardiovasc Med. 11:14001302024.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Yang L, Wang Y, Ye X, Liu Q, Qu D and Chen
Y: Traditional Chinese medicine-based drug delivery systems for
anti-tumor therapies. Chin J Nat Med. 22:1177–1192. 2024.PubMed/NCBI
|
|
139
|
Zhang YJ, He H, Sawuer G, Ma XK, Ainiwaer
Z, Wu DD, Zhang XX and An DQ: Tianxiangdan suppresses foam cell
formation by enhancing lipophagy and reduces the progression of
atherosclerosis. In Vitro Cell Dev Biol Anim. 61:298–310. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Li Y, Pan J, Yu JJJ, Wu X, Yang G, Pan X,
Sui G, Wang M, Cheng M, Zhu S, et al: Huayu Qutan Recipe promotes
lipophagy and cholesterol efflux through the
mTORC1/TFEB/ABCA1-SCARB1 signal axis. J Cell Mol Med.
28:e182572024. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Bao Y, Zhu L, Wang Y, Liu J, Liu Z, Li Z,
Zhou A and Wu H: Gualou-Xiebai herb pair and its active ingredients
act against atherosclerosis by suppressing VSMC-derived foam cell
formation via regulating P2RY12-mediated lipophagy. Phytomedicine.
128:1553412024. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Jiang S, Li H, Zhang L, Mu W, Zhang Y,
Chen T, Wu J, Tang H, Zheng S, Liu Y, et al: Generic diagramming
platform (GDP): A comprehensive database of high-quality biomedical
graphics. Nucleic Acids Res. 53:D1670–D1676. 2025. View Article : Google Scholar : PubMed/NCBI
|