Introduction
The modification of N6-methyladenosine (m6A) is the most prevalent internal chemical modification in eukaryotic messenger RNA and it is a core component of epitranscriptomic regulation. By dynamically modulating RNA splicing, stability, translation and degradation, m6A modification participates in the regulation of gene expression and the maintenance of cellular function (1). This reversible modification network, orchestrated by methyltransferases (for example, the METTL3 complex), demethylases (for example FTO and ALKBH5) and reader proteins [for example the YTH domain-containing family (YTHDF) of proteins], plays a critical role in development, metabolism and disease pathogenesis (2). The reversible nature of m6A modification was first established by the discovery of FTO as an m6A demethylase in 2011, followed by the identification of ALKBH5. Concurrently, transcriptome-wide m6A mapping revealed its widespread distribution and conserved sequence motifs. These seminal studies transformed m6A from a static mark into a dynamic and reversible regulatory mechanism, sparking the field of epitranscriptomics. Recent studies have revealed that aberrant m6A modification is closely associated with various diseases, including cancer, neurodegenerative disorders and metabolic dysregulation, underscoring its potential as a biomarker and therapeutic target (3–5).
Retinal diseases, such as diabetic retinopathy (DR), age-related macular degeneration (AMD) and inherited retinal dystrophies, such as retinitis pigmentosa (RP), are leading causes of irreversible vision loss, with their pathogenesis closely linked to genetic and environmental factors (6). Epigenetic mechanisms, including DNA methylation, histone modification and RNA modification, play significant roles in retinal physiology and pathology (7). Among these, m6A modification has emerged as a novel regulatory mechanism that is highly enriched in retinal cells, which participates in processes such as angiogenesis, inflammation and neuronal survival (8). For instance, in DR, hyperglycemia induces alterations in the m6A modification profile, thereby affecting the expression of key molecules such as vascular endothelial growth factor and exacerbating retinal damage (9). Additionally, m6A modification is implicated in degenerative conditions such as AMD, suggesting its broad relevance in ophthalmic diseases (10,11).
The present review comprehensively summarizes the mechanisms and clinical implications of m6A modification in retinal diseases. It begins by outlining the fundamental regulatory network of m6A modification and its expression characteristics in retinal cells (12) It then analyzes its specific regulatory roles in diseases such as DR and AMD (13), discusses therapeutic strategies targeting the m6A pathway and their prospects (14), and finally addresses current research limitations and future directions, including the integration of multi-omics technologies and the potential of artificial intelligence in personalized medicine (15). By synthesizing advances in m6A research in retinal diseases, the present review aims to provide a theoretical foundation for understanding disease mechanisms and for developing therapeutic strategies (16,17).
Search strategy
A systematic literature search was performed using the PubMed(https://pubmed.ncbi.nlm.nih.gov/) and Web of Science databases(https://www.webofscience.com), covering the period from database inception to October 2025. The search strategy combined terms related to m6A modification and retinal diseases. For m6A modification, the following keywords were used: ‘N6-methyladenosine’, ‘m6A’, ‘METTL3’, ‘METTL14’, ‘WTAP’, ‘FTO’, ‘ALKBH5’, ‘YTHDF1’, ‘YTHDF2’ and ‘YTHDC1’. For search of retinal diseases, full list search terms used included ‘diabetic retinopathy’, ‘age-related macular degeneration’, ‘retinoblastoma’, ‘uveitis’ and ‘retinitis pigmentosa’. Boolean operators (AND/OR) were applied to combine these term groups. Additional relevant studies were identified by manually screening the reference lists of retrieved articles and previous reviews. Inclusion and exclusion criteria: Only peer-reviewed, original research articles and review papers published in English were included. The final reference list was selected based on relevance to the regulatory roles, pathological implications and therapeutic targeting of m6A modification in the retina.
Physiology and clinical management of the normal retina
The vertebrate retina is a highly organized layered structure comprising multiple cell types that work in concert to process visual information. Neuronal components include photoreceptors (rods and cones), bipolar cells, ganglion cells, horizontal cells and amacrine cells, with over 60 distinct neuronal subtypes identified in mammals (18). In addition to these neurons, Müller glial cells provide structural and metabolic support, and the retinal pigment epithelium (RPE) forms the outer blood-retinal barrier and maintains photoreceptor homeostasis (19). Rod photoreceptors dominate in number and mediate dim-light vision, whereas cones enable color and high-acuity vision. The cone output is decomposed into ~12 parallel channels via distinct bipolar cell types, which then relay specific visual features to different subsets of retinal ganglion cells (18). Amacrine cells, of which nearly 30 types exist, refine these signals through lateral and crossover inhibition, generating diverse encodings such as direction selectivity and motion detection (18).
From a clinical perspective, standard therapeutic approaches for common retinal diseases have been well established. For DR and neovascular AMD, intravitreal anti-vascular endothelial growth factor (anti-VEGF) agents represent first-line therapy; laser photocoagulation and corticosteroids are also used as adjunctive or alternative treatments (19). In early and intermediate AMD, lifestyle modifications (smoking cessation) and nutritional supplementation with the AREDS2 formulation (lutein, zeaxanthin, vitamins C and E, and zinc) are recommended to reduce the risk of progression (19). For RP, no curative therapy exists for most genetic subtypes, but gene replacement therapy has been approved for RPE65-associated RP, and optogenetic strategies are under investigation (20). Autoimmune retinopathy, a rare inflammatory condition, is diagnosed based on clinical, imaging and electrophysiological criteria, and treatment primarily aims to prevent progression (21). Understanding normal retinal physiology and the existing treatment landscape provides a foundation for exploring emerging epitranscriptomic regulators, such as m6A modification, in retinal diseases.
Role of m6A modification in DR
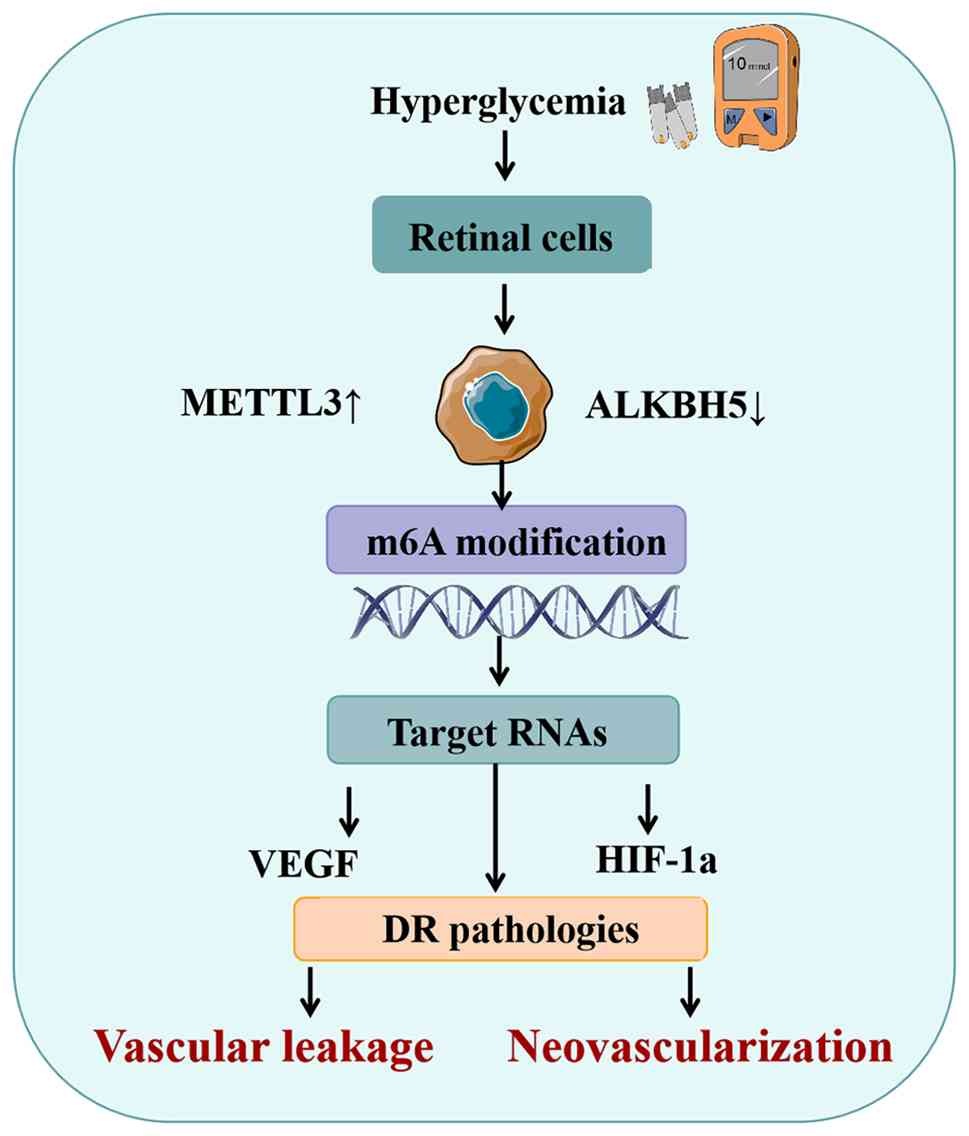
DR is a common microvascular complication of diabetes that represents a leading cause of blindness worldwide (22). Its pathological features include increased vascular permeability, neovascularization, inflammatory responses and neurodegeneration, ultimately leading to retinal dysfunction (23). Hyperglycemia drives DR progression through mechanisms such as oxidative stress, inflammatory activation and dysregulation of the VEGF and hypoxia induced factor 1α signaling pathway. A recent study indicated that m6A modification participates in the pathogenesis of DR, providing new insights into its molecular mechanisms (24).
METTL3, a key methyltransferase, shows upregulated expression in DR and regulates downstream genes via m6A modification. For instance, METTL3-mediated m6A modification enhances the stability of piezo-type mechanosensitive ion channel component 1 (PIEZO1) mRNA, yet promotes its degradation via YTHDF2, thereby exacerbating the inflammatory response in Müller cells (25). METTL3 also cooperates with YTH domain-containing protein 1 (YTHDC1) to upregulate the long non-coding RNA OGRU, activating oxidative stress pathways and contributing to Müller cell injury (26). The reader protein YTHDF2 plays a significant role in DR as well. Downregulation of circular RNA circFAT1 influences autophagy and pyroptosis in RPE cells by interacting with YTHDF2, aggravating cellular damage (27). The demethylase ALKBH5, which is downregulated in DR, may suppress ferroptosis through YTHDF1-mediated degradation of acyl-CoA synthetase long chain family member 4 (ACSL4) mRNA, potentially delaying disease progression (28). These findings suggest that m6A modification participates in multi-level regulation of gene networks associated with DR, such as modulating the VEGF signaling pathway and inflammatory factor expression, serving as a central regulatory mechanism in the hyperglycemic microenvironment (29,30).
Experimental studies further support the functional role of m6A modification in DR. Elevated global m6A levels, along with increased expression of METTL3 and YTHDF2, have been observed in human RPE cells (ARPE-19) and Müller cells under high-glucose conditions. Knockdown of METTL3 alleviates high glucose-induced cellular injury (31). In diabetic mouse models, abnormal retinal m6A modification levels have been reported, and interference with METTL3 expression ameliorates vascular leakage and neurodegeneration (29). Analysis of clinical samples reveals reduced ALKBH5 expression in the retinas of patients with DR, which correlates with disease severity (32). Collectively, these data underscore the critical involvement of m6A modification in the pathogenesis of DR and provide a rationale for its targeting as a therapeutic strategy (Fig. 1).
Role of m6A modification in AMD
AMD is a leading cause of vision loss in the elderly, characterized by RPE atrophy and choroidal neovascularization (CNV) (33). Its pathogenesis involves oxidative stress, inflammatory responses, VEGF signaling dysregulation and epigenetic disturbances (34). A previous study has identified m6A modification as a significant contributor to RPE degeneration and CNV formation (35).
m6A modification participates in various pathological processes of AMD by regulating RNA metabolism. METTL3, which is upregulated in the RPE of patients with AMD, enhances high mobility group AT-hook 2 mRNA stability through m6A modification, thereby activating the epithelial-mesenchymal transition (EMT) pathway and promoting fibrosis (33). The demethylase ALKBH5, which is elevated in wet AMD, stabilizes DNA binding 2 (ID2) mRNA to facilitate migration and angiogenesis of retinal microvascular endothelial cells (36). Downregulated circular RNA circSPECC1 affects the miR-145-5p/CDKN1A axis via the m6A-YTHDC1 pathway, inhibiting oxidative stress-induced ferroptosis (37). Furthermore, FTO activates the NLR family pyrin domain-containing 3 (NLRP3) inflammasome by modulating m6A modification of the long non-coding RNA Neat1, contributing to RPE pyroptosis (38,39). These mechanisms illustrate the multi-level regulatory role of m6A in AMD pathogenesis.
Experimental evidence further supports its functional significance. In a laser-induced CNV model, knockdown of WT1-associated protein (WTAP) suppressed pathological angiogenesis (40). Intervention in METTL3 or ALKBH5 expression in RPE cells influenced EMT and angiogenic processes, respectively (33,36). Clinical sample analyses suggest that m6A-related factors such as METTL3 are associated with AMD risk (41), while FTO inhibitors exhibit anti-inflammatory effects in experimental models (38). Collectively, these studies demonstrate the critical regulatory function of m6A modification in AMD development.
Role of m6A modification in RB and uveitis: Current evidence and limitations
Although RB (a malignant tumor) and uveitis (an autoimmune inflammatory disease) differ fundamentally in etiology and pathology, they share a common challenge in m6A research: Direct evidence remains extremely scarce. The majority of our current understanding of m6A modification in these two conditions is extrapolated from studies on other cancers (for RB) or other inflammatory/autoimmune diseases (for uveitis). While such extrapolations offer valuable mechanistic insights and potential therapeutic directions, they also carry a high risk of overgeneralization. Therefore, the present review reports the current knowledge on m6A in RB and uveitis in the following two subsections, followed by a consolidated discussion of limitations and future priorities.
m6A in RB and uveitis
RB is the most common intraocular malignancy in children, primarily caused by biallelic mutations in the RB1 gene. Clinical manifestations include leukocoria, strabismus or visual impairment, with potential metastasis and poor treatment outcomes in advanced cases (42,43). Recent research has highlighted the role of m6A modification in tumor progression, as it regulates gene expression at the post-transcriptional level and may contribute to the malignant progression of RB (44).
Although studies on m6A in RB remain limited, mechanisms observed in other cancers may provide insights. METTL3-mediated m6A modification enhances the stability and translation efficiency of oncogenic transcripts, promoting tumor progression (45). YTHDF2 recognizes m6A marks to facilitate the degradation of target mRNAs, affecting genes related to cell cycle and apoptosis (46). FTO regulates signaling pathways such as MYC through demethylation (47). These mechanisms suggest potential analogous functions of m6A modification in RB.
Indirect experimental evidence supports its potential role. In lung cancer, METTL3 knockdown suppresses the expression of genes such as cyclin-dependent kinase 6 (CDK6) and matrix metalloproteinase-2, delaying tumor progression (48). In hepatocellular carcinoma, ALKBH5 influences malignant phenotypes by regulating signal transducer and activator of transcription 1 mRNA stability (49). In bladder cancer, YTHDF2 promotes tumor growth by degrading period circadian regulator 1 mRNA (50). These findings offer a reference for understanding the role of m6A in RB.
Therapeutic strategies targeting the m6A regulatory network show promising potential. The METTL3 inhibitor STM2457 suppresses acute myeloid leukemia cell proliferation (51). The FTO inhibitor FB23-2 induces cell differentiation and apoptosis (52). The YTHDF1 inhibitor ebselen exhibits antitumor effects in breast cancer (53). These discoveries provide new directions for RB treatment.
Uveitis is an inflammatory disease involving the iris, ciliary body and choroid that is frequently associated with autoimmune dysregulation. Characterized by immune cell infiltration and breakdown of the blood-retinal barrier, it represents a significant cause of vision impairment in young and middle-aged adults (54). m6A modification, which is critically involved in immunoregulation, may contribute to the development of uveitis. METTL3-mediated m6A modification promotes the stability of T cell immunoreceptor with Ig and ITIM domains (TIGIT) mRNA, thereby regulating T cell function. During T-cell exhaustion, METTL3 upregulates TIGIT expression by enhancing its mRNA stability, leading to T-cell dysfunction. T cell-mediated autoimmune responses play a central role in its pathogenesis (55). This suggests that m6A modification may influence immune homeostasis in uveitis by modulating immune checkpoint molecules.
Furthermore, m6A modification participates in innate immune regulation. In conditions such as juvenile idiopathic arthritis, decreased expression of FTO and ALKBH5 in monocytes at inflammatory sites leads to elevated m6A levels, promoting aberrant monocyte activation (56). Given that monocyte/macrophage infiltration is a key feature of uveitis, m6A may influence the inflammatory process by regulating their activation state.
An experimental study indicated that METTL3 knockdown ameliorates T cell exhaustion (57), and peripheral blood m6A levels correlate with disease activity in autoimmune disorders. Although direct evidence in uveitis is limited, these findings imply that m6A may participate in disease pathogenesis through its regulation of T cells and monocytes/macrophages.
Therapeutic targeting of m6A regulators, such as the METTL3 inhibitor STM2457, have demonstrated anti-inflammatory effects in various models, offering new avenues for immunotherapy in uveitis
Limitations and future directions
Due to the scarcity of direct studies on m6A modification in RB and uveitis, the above discussion partly relies on findings from other cancers or inflammatory diseases. These extrapolations should be interpreted with caution, and future work using RB-specific and uveitis-specific models (for example patient-derived xenografts for RB, experimental autoimmune uveitis models) is urgently needed to validate the proposed mechanisms and to explore the therapeutic potential of targeting m6A regulators in these conditions.
Role of m6A modification in RP
RP is an inherited retinal degenerative disorder characterized primarily by photoreceptor cell apoptosis. It is frequently caused by mutations in genes such as RP GTPase regulator (RPGR), and effective treatments are currently lacking (58,59).
A study has shown that m6A modification contributes to the progression of RP by dynamically regulating gene expression. METTL14 expression is reduced in patients, and it regulates microtubule-associated protein 2 (MAP2) expression via m6A modification, influencing phagocytosis, proliferation and apoptosis in retinal pigment epithelial cells. YTHDF2 recognizes m6A marks on MAP2 mRNA, affecting its stability and subsequently modulating factors such as neuronal differentiation 1, thereby playing a role in maintaining cellular function (60).
Additionally, circular RNAs derived from the RPGR gene locus contain m6A modification motifs, which may participate in the regulation of RPGR pre-mRNA and influence photoreceptor cell survival (61). A study has demonstrated that METTL14 knockdown impairs phagocytosis and increases apoptosis in ARPE-19 cells, with MAP2 identified as a key target (60). In animal models, METTL14 deficiency exacerbates retinal degeneration, and m6A modification of circular RNAs may be involved in translational regulation (61), supporting the functional relevance of m6A in this disease.
m6A modification may also influence disease progression by regulating oxidative stress and inflammatory responses. Targeted strategies such as METTL3 inhibitors have shown protective effects in glaucoma models (62), and FTO/ALKBH5 inhibitors have exhibited efficacy in other retinal disorders, suggesting their therapeutic potential for RP.
Discussion and future perspectives
Critical synthesis of contradictory findings
The m6A regulatory network exhibits marked context-dependent functions across different retinal diseases. An example is ALKBH5, a m6A demethylase. In DR, ALKBH5 exerts protective effects by reducing ferroptosis through YTHDF1-mediated degradation of ACSL4 mRNA (28). Conversely, in AMD, ALKBH5 promotes choroidal neovascularization and subretinal fibrosis by stabilizing ID2 mRNA (36). Such opposing roles highlight that the therapeutic outcome of targeting a single m6A regulator may vary dramatically depending on disease context, cell type and microenvironment. Similarly, METTL3 has been consistently reported as pathogenic in both DR and AMD, driving inflammation in the former and fibrosis in the latter (33,63). However, functional redundancy among methyltransferases (for example METTL3 and METTL14) may complicate the interpretation of single-enzyme inhibition studies. These discrepancies underscore the need for cell-type-specific and disease-stage-specific investigations before clinical translation (41).
Common m6A-regulated modules across retinal diseases
Despite disease-specific differences, several common m6A-regulated pathological modules recur across DR, AMD, RP and other retinal disorders. First, inflammation is a contributing factor. METTL3-mediated m6A modification enhances PIEZO1 mRNA stability in DR (25) and activates the NLRP3 inflammasome in AMD (38), both leading to inflammatory damage. Second, angiogenesis represents another convergent module. ALKBH5-driven ID2 upregulation promotes choroidal neovascularization in AMD (36), while METTL3 influences VEGF expression in DR (29). Third, cell death pathways including ferroptosis, pyroptosis and autophagy are commonly regulated by m6A. For instance, ALKBH5 protects against ferroptosis in DR via ACSL4 (28), whereas FTO promotes pyroptosis in AMD via Neat1 (38), and METTL3 modulates autophagy in retinal ganglion cells (64). These shared modules suggest that targeting a single m6A regulator may have pleiotropic effects across multiple diseases, but also raises concerns about off-target outcomes.
Positioning of m6A-targeted therapy relative to current treatments
Current standard therapies for major retinal diseases include anti-VEGF agents (for DR and neovascular AMD), laser photocoagulation, corticosteroids and, for selected RP cases, gene replacement therapy (65,66). While these treatments have proven efficacy, limitations such as frequent intravitreal injections, non-response in a subset of patients (for example anti-VEGF resistance in DR), and lack of curative options for degenerative conditions persist. In this context, m6A-targeted agents could be positioned in several ways: i) As adjunctive therapy combined with anti-VEGF to enhance efficacy or reduce injection frequency; ii) as an alternative or add-on for anti-VEGF-resistant patients, particularly those with persistent inflammation or ferroptosis-driven pathology; and iii) for early-stage intervention, targeting m6A modules before irreversible vascular or neuronal damage occurs. For RP and uveitis, where anti-VEGF is not the mainstay, m6A modulators may complement gene therapy (by stabilizing photoreceptor transcripts) or immunosuppressants (by fine-tuning T cell responses) (67).
Delivery and safety challenges
Systemic administration of m6A-targeting small molecules (for example STM2457 and FB23-2) poses a risk of off-target effects due to the ubiquitous expression of m6A machinery across tissues. However, the eye offers unique advantages for local delivery. Intravitreal injection can achieve high retinal drug concentrations while minimizing systemic exposure (68). Nevertheless, several challenges remain. First, current inhibitors lack long-term safety data in large animal models; non-human primate studies are urgently needed. Second, retinal cell-type specificity is difficult to achieve with small molecules alone. Emerging delivery strategies include adeno-associated virus (AAV) vectors for targeted expression of m6A enzyme modulators (41), nanoparticle-based carriers to enhance retinal penetration (69), and biodegradable hydrogels for sustained release. Third, potential immune responses against viral vectors or repeated injections must be addressed. These safety and delivery considerations are critical for clinical translation.
Future perspectives
Future research should prioritize the development of tissue-specific regulatory tools, such as AAV vectors for precise editing of m6A enzymes (41). Mapping m6A epitranscriptomes across disease stages via single-cell sequencing will help elucidate their dynamic changes (70). Concurrently, deeper investigation into the interplay between m6A and other epigenetic mechanisms such as histone modifications and DNA methylation is warranted (71). Optimizing nanoparticle-based delivery systems may enhance retinal drug accumulation and reduce systemic toxicity (72). Establishing non-human primate models will be crucial for evaluating long-term safety and efficacy. Furthermore, advancing the clinical translation of m6A-related biomarkers (for example blood METTL14 levels) could support personalized treatment strategies (73).
Notably, m6A-targeted agents may synergize with conventional therapies. In diabetic retinopathy, combining m6A regulators with anti-VEGF drugs could enhance efficacy, while in RP, integrating gene therapy with m6A modulation may yield synergistic protective effects. Artificial intelligence-driven multi-omics analyses hold promise for uncovering novel m6A regulatory networks, providing a theoretical foundation for combination therapies. Progress in these areas will require interdisciplinary collaboration across epigenetics, bioinformatics, materials science and clinical medicine.
In summary, m6A modification serves as a critical epitranscriptomic regulatory mechanism in retinal diseases (Fig. 2). Although m6A-targeted therapies face multiple challenges, advancing mechanistic studies, developing precise regulatory tools and promoting clinical validation may offer new hope for patients. Future efforts should focus on multi-omics integration, delivery system optimization, and rigorous clinical trial design to accelerate the translation of m6A-based treatments into clinical practice.
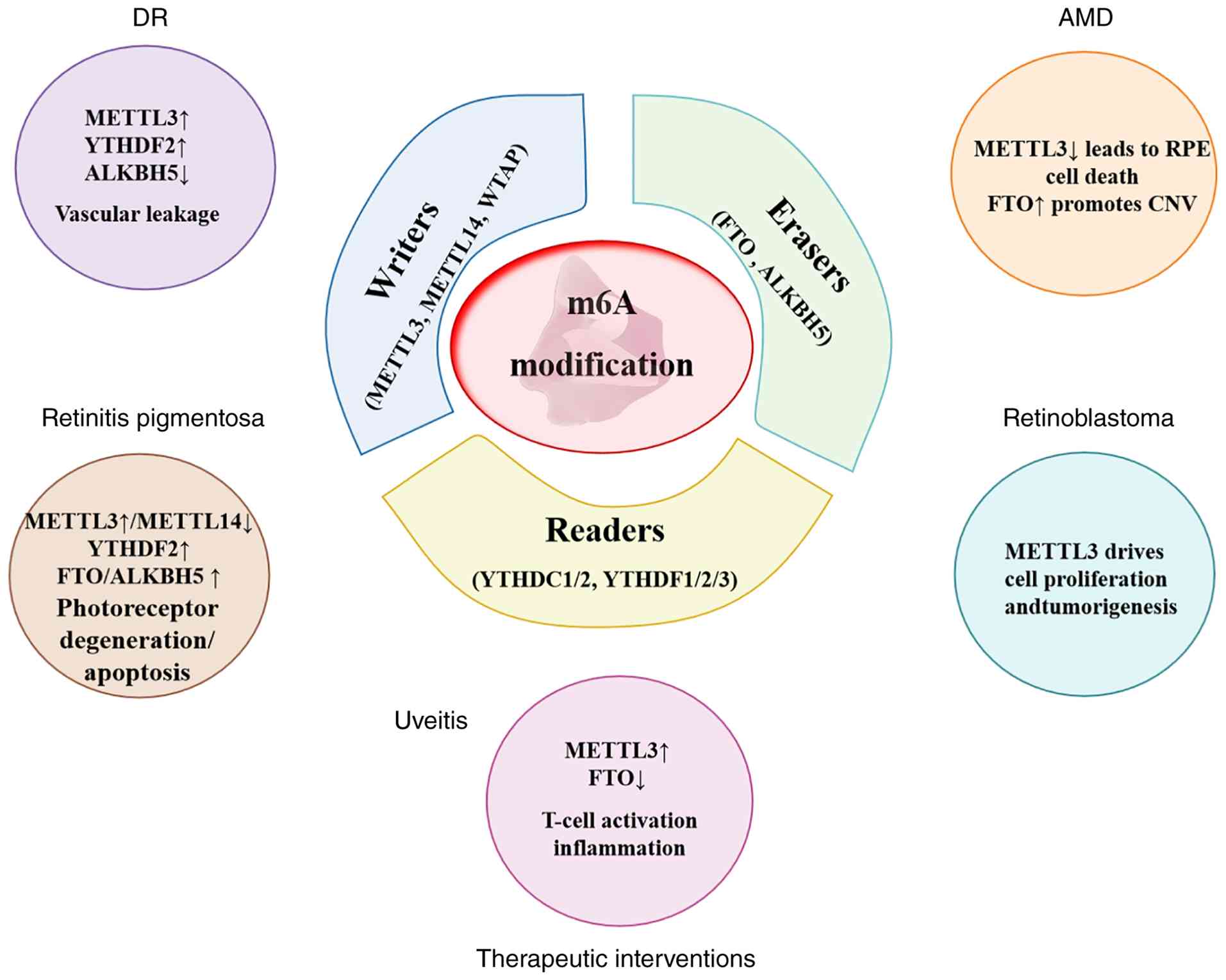 |
Figure 2.
Central role of m6A modification in retinal diseases. This graphical abstract illustrates m6A RNA modification as a dynamic regulatory hub, orchestrated by writers (methyltransferases), erasers (demethylases) and readers (binding proteins), in the pathogenesis of major retinal diseases, including DR, AMD, retinoblastoma, uveitis and retinitis pigmentosa. Although the figure presents disease-specific pathways, several m6A-regulated pathological modules-such as inflammation (DR, uveitis and AMD), angiogenesis (DR and AMD) and cell death, including ferroptosis and pyroptosis (DR, AMD and RP) - recur across multiple diseases, representing shared mechanisms. Arrows indicate the dysregulation of key m6A-modifying enzymes (for example METTL3 and ALKBH5) and their influence on disease-specific pathological processes. The lower section highlights the therapeutic potential of targeting the m6A pathway with small-molecule inhibitors. m6A, N6-methyladenosine; DR, diabetic retinopathy; AMD, age-related macular degeneration; RPE, retinal pigment epithelium; CNV, choroidal neovascularization.
|
Conclusion
Collectively, the present review delineates the critical involvement of m6A modification in the pathogenesis of various retinal diseases, including diabetic retinopathy, age-related macular degeneration, RB, uveitis and RP. The dynamic and reversible nature of m6A modification, governed by methyltransferases, demethylases and reader proteins, exerts multifaceted control over key pathological processes such as aberrant angiogenesis, inflammation, cellular death and photoreceptor degeneration. Emerging evidence highlights the therapeutic potential of targeting specific m6A regulators, exemplified by small-molecule inhibitors like STM2457 and FB23-2 in preclinical models (Fig. 3).
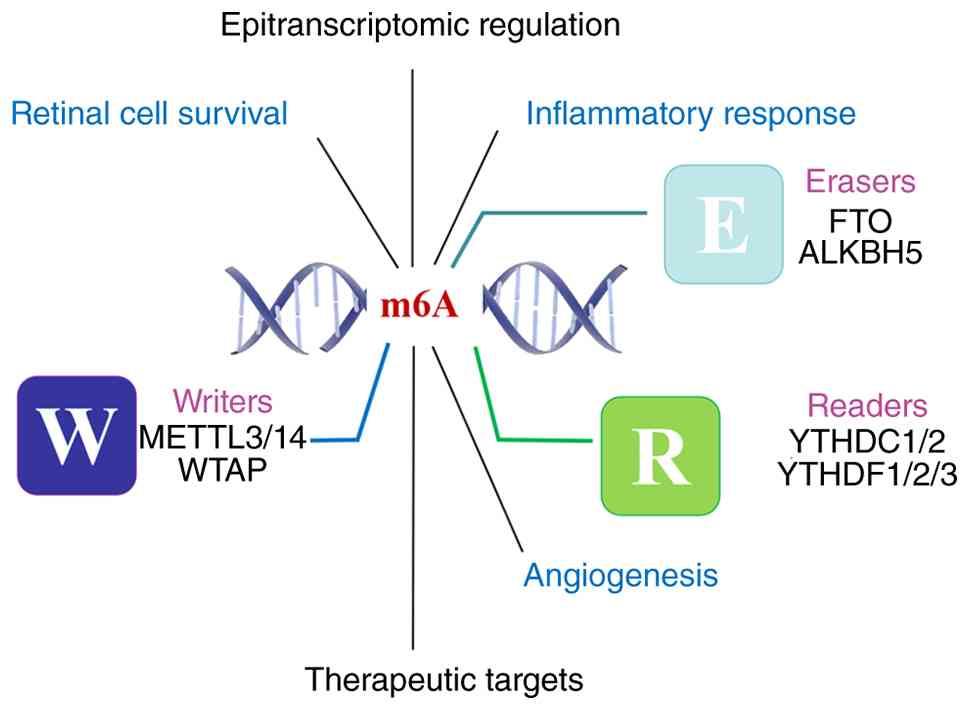 |
Figure 3.
Schematic representation of the m6A regulatory network in retinal pathophysiology. This model illustrates the central role of m6A RNA modification, mediated by writers (W), erasers (E) and readers (R), in coordinating key biological processes within the retina. The dynamic interplay of these regulatory proteins influences a network of downstream pathways, including epitranscriptomic regulation, inflammatory response, angiogenesis and cell survival, which collectively contribute to retinal disease pathogenesis. The convergence of these pathways highlights the potential of targeting the m6A machinery for therapeutic intervention. m6A, N6-methyladenosine; W, writers; E, erasers; R, readers; WTAP, WT1-associated protein; YTHDC1, YTH domain-containing protein 1.
|
However, translating these findings into clinical applications faces challenges, primarily concerning cell-type specificity, efficient retinal drug delivery and functional redundancy within the m6A network. Future research should prioritize the integration of multi-omics approaches, the development of targeted delivery systems and rigorous validation in physiologically relevant models. Overcoming these hurdles will be instrumental in harnessing the epitranscriptome for novel diagnostic and therapeutic strategies against retinal diseases, ultimately aiming to preserve vision in affected patients.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (grant no. 82271094), the Science and Technology Projects in Guangzhou (grant no. 202201010618) and the Medical Scientific Research Foundation of Guangdong Province of China (grant no. A2022415).
Availability of data and materials
Not applicable.
Authors' contributions
WZ conceived the article and wrote the manuscript. WZ and JC performed the literature search and revised the manuscript. JC and JZ critically revised the manuscript. All authors read and approved the final manuscript. Data authentication is not applicable.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
|
1
|
Li S, Luo P, Fan J, Li Y, Tu J and Long X: RNA modifications in health and disease. MedComm (2020). 6:e703412025. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gao X, Wang H, Zhang H, Zhan J, Liu Z and Cheng Y: The METTL family as regulators of methylation and therapeutic targets in cardiovascular diseases. Eur J Pharmacol. 1008:1783442025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lagunas-Rangel FA: METTL3 in colorectal cancer: Molecular insights and clinical implications. Mol Biol Rep. 53:982025. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fu X, Ruan X and He J: METTL3-driven m6A epigenetics in gastric cancer: Unveiling oncogenic networks and clinical translation from tumorigenesis to therapy resistance. Cell Biol Toxicol. 41:1322025. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu S, Liu J, Chen S, Li B and Ye Z: M(6)A methylation in tumor immune microenvironment: Multidimensional mechanism and targeted therapy strategies. Biochim Biophys Acta Rev Cancer. 1880:1894892025. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gettinger K, Lee D, Tomita Y, Negishi K and Kurihara T: Diabetic retinopathy, a comprehensive overview on pathophysiology and relevant experimental models. Int J Mol Sci. 26:98822025. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu T, Yang XJ, Zhou L, Gan M, He TT, Hong S, Feng YY, Su G, Zhao YX, Cao Y and Zeng QF: Epigenetic regulation in cognitive impairment: Focus on N6-methyladenosine modification and its potential role in perioperative neurocognitive disorders. Ibrain. 11:332–346. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kumari N, Karmakar A, Ahamad Khan MM and Ganesan SK: The potential role of m6A RNA methylation in diabetic retinopathy. Exp Eye Res. 208:1086162021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lei C, Lv Z, Ran Q, Jiang F and Zhang M: Homocysteine and diabetic retinopathy. Exp Eye Res. 262:1107292026. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kadonosono K, Hayashi A and de Juan E Jr: Endovascular surgery in the field of ophthalmology. Jpn J Ophthalmol. 65:1–5. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nowak JZ: Age-related macular degeneration (AMD): Pathogenesis and therapy. Pharmacol Rep. 58:353–363. 2006.PubMed/NCBI
|
|
12
|
Li X, Xu YR and Zhang J: The role of E3 ubiquitin ligases in vertebrate eye development: Mechanisms and emerging perspectives. Exp Eye Res. 262:1107022026. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kalogeropoulos D, Moussa K, Copado IA, Habot-Wilner Z, Touhami S, Ploumi I and Androudi S: Non-neoplastic disorders mimicking posterior segment inflammation. Ocul Immunol Inflamm. 33:2486–2497. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Le Du J and Ronco C: Therapeutic strategies targeting ocular vasculopathies: Current advances and emerging challenges. Drug Discov Today. 30:1044962025. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kiraly P and Fischer MD: Artificial intelligence applications in inherited retinal dystrophies. Graefes Arch Clin Exp Ophthalmol. 264:299–307. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schleiss MR: The urgent search for predictive biomarkers in the emerging era of universal congenital cytomegalovirus screening. Philos Trans R Soc Lond B Biol Sci. 380:202404342025. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun K, Li J, Chen C, Zhou X, Ma G, Mao L, Tang Q, Ma B, Li D, Chen Z, et al: Advances in nanopore direct RNA sequencing and its impact on biological research. Biotechnol Adv. 85:1087102025. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Masland RH: The neuronal organization of the retina. Neuron. 76:266–280. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
García-Layana A, Cabrera-López F, García-Arumí J, Arias-Barquet L and Ruiz-Moreno JM: Early and intermediate age-related macular degeneration: update and clinical review. Clin Interv Aging. 12:1579–1587. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hoon M, Okawa H, Della Santina L and Wong RO: Functional architecture of the retina: development and disease. Prog Retin Eye Res. 42:44–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Modjtahedi BS, Palestine AG, Jampol LM, Sarraf D, Sen HN, Sobrin L, Chen JJ, Yang P, Adamus G, Fong DS, et al: Guidelines for the diagnosis, management, and study of autoimmune retinopathy from the American academy of ophthalmology's task force. Ophthalmol Retina. 9:1005–1016. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He W, Tang P and Lv H: Targeting oxidative stress in diabetic retinopathy: Mechanisms, pathology, and novel treatment approaches. Front Immunol. 16:15715762025. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McLellan FC, Huang K, Wong E and Shu DY: The angiofibrotic switch in retinal and choroidal vascular diseases: Mechanistic drivers of angiogenesis and endothelial-mesenchymal transition. Am J Pathol. 195:1363–1375. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang Z, Guan J, Zhang X, Hu Q, Huang J, Sang W, Wang X, Jiang B and Sun D: METTL14-dependent regulation of HMGB1 attenuates inflammation in diabetic retinopathy. Acta Diabetol. 63:245–257. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han N, Yu N and Yu L: The mRNA stability of PIEZO1, regulated by methyltransferase-like 3 via N(6)-Methylation of adenosine modification in a YT521-B homology domain family 2-Dependent manner, facilitates the progression of diabetic retinopathy. Am J Pathol. 195:265–280. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fu S, Zhou Q, Peng X, Hu Y, Xiong J and Liu F: METTL3/YTHDC1 mediates up-regulation of lncRNA OGRU in an m6A-dependent manner involving in oxidative stress and inflammation of HG-induced Müller cells. Immunol Lett. 272:1069722025. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang C, Qi P, Cui H, Lu Q and Gao X: CircFAT1 regulates retinal pigment epithelial cell pyroptosis and autophagy via mediating m6A reader protein YTHDF2 expression in diabetic retinopathy. Exp Eye Res. 222:1091522022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liao Q, Li Y, Cui M and Liu M: m6A demethylase ALKBH5 reduces ferroptosis in diabetic retinopathy through the m6A-YTHDF1-ACSL4 axis. Diabet Med. 42:e700332025. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang H, Fu W, Cai Y and Xu H: METTL3-Mediated N6-Methyladenosine modification regulates the progression of diabetic retinopathy. Appl Biochem Biotechnol. 197:5992–6008. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hua Z, Zhong F, Xu C and Liang S: METTL14 aggravates pyroptosis in diabetic cardiomyopathy by promoting m6A modification of NLRP3. Immunol Res. 73:1452025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo D and Zhang B: METTL3-Mediated m(6)A Regulation of GDF11 promotes socket healing in diabetic rats. Diabetes Metab Syndr Obes. 18:3505–3514. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheng Q, Zhou L, Fan X, Ma M, Zhang C, Zha X and Zhang Y: FTO-mediated Nrf2 demethylation alleviates high glucose-induced oxidative stress and apoptosis in retinal pigment epithelial cells. Mol Biol Rep. 52:2892025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Chen Y, Liang J, Jiang M, Zhang T, Wan X, Wu J, Li X, Chen J, Sun J, et al: METTL3-mediated m6A modification of HMGA2 mRNA promotes subretinal fibrosis and epithelial-mesenchymal transition. J Mol Cell Biol. 15:mjad0052023. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li B, Wang Z, Zhou H, Zou J, Yoshida S and Zhou Y: N6-methyladenosine methylation in ophthalmic diseases: From mechanisms to potential applications. Heliyon. 10:e236682023. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li X, Ma B, Zhang W, Song Z, Zhang X, Liao M, Li X, Zhao X, Du M, Yu J, et al: The essential role of N6-methyladenosine RNA methylation in complex eye diseases. Genes Dis. 10:505–520. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wei Y, Hu X, Liu Y, Zhang J, Zhao L, Yang J, Xie Z, Shi D and Ma L: ALKBH5-mediated m6A modification of ID2 mRNA promotes choroidal neovascularization and subretinal fibrosis. Cell Signal. 138:1122372026. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen X, Wang Y, Wang JN, Cao QC, Sun RX, Zhu HJ, Zhang YR, Ji JD and Liu QH: m(6)A modification of circSPECC1 suppresses RPE oxidative damage and maintains retinal homeostasis. Cell Rep. 41:1116712022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xue Q, Huang J, Wang B, Ji J, Wang L, Kumari S, Lan C and Xiao M: FTO Fuels Aβ(1–40)-Induced retinal pigment epithelium cell injury associated with pyroptosis by erasing m6A methylation of the lncRNA Neat1. FASEB J. 39:e711662025. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hu Y, Chen J, Wang Y, Sun J, Huang P, Feng J, Liu T and Sun X: Fat mass and obesity-associated protein alleviates Aβ(1–40) induced retinal pigment epithelial cells degeneration via PKA/CREB signaling pathway. Cell Biol Int. 47:584–597. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gong Q, Hu L, Liu G, Yin X, Zhao X, Li Q, Li Y, Sun Y, Zhou Y, Guo C and Du Z: WTAP-mediated N6-methyladenosine mRNA methylation regulates laser-induced macular neovascularization. Mol Vis. 30:336–347. 2024.PubMed/NCBI
|
|
41
|
Gong YJ, Zou ZL, Qiu KR, Wang Q and Zhou XL: Integration of multi-omics data reveals dysregulated RNA methylation in retinal pigment epithelium drives age-related macular degeneration. Int J Ophthalmol. 18:1626–1639. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lohmann DR and Gallie BL: Retinoblastoma: Revisiting the model prototype of inherited cancer. Am J Med Genet C Semin Med Genet. 129C:23–28. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu H, Zhang Y, Zhang YY, Li YP, Hua ZQ, Zhang CJ, Wu KC, Yu F, Zhang Y, Su J and Jin ZB: Human embryonic stem cell-derived organoid retinoblastoma reveals a cancerous origin. Proc Natl Acad Sci USA. 117:33628–33638. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen J and Zeng B: METTL14-mediated m6A modification of LINC00340 exerts oncogenic role in retinoblastoma by regulating Notch signaling pathway. Int Ophthalmol. 45:732025. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ran Q, Gao J, Li G, Wang J, Li X, Xiong A, Zhang Y, Xiong Y and He X: METTL3-driven m(6)A modification orchestrates mitophagy-dependent ferroptosis in PM2.5-induced lung injury. Front Immunol. 16:16838192025. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tang D, Cao C, Huang S, He Q and Wang A: YTHDF2 drives oral squamous cell carcinoma progression via m(6)A-dependent degradation of MTUS1/ATIP1 mRNA and mitochondrial dysregulation. Cell Signal. 136:1121452025. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tian J, He Q, Li N, Sun Y, Zhang A and Wang H: FTO-Mediated m6A Demethylation of MZF1 Regulates DECR1 to promote fatty acid oxidation and exacerbate myocardial ischemia/reperfusion injury: FTO-Mediated m6A Demethylation of MZF1 enhances fatty acid oxidation and aggravates myocardial I/R injury. Biochem Genet. Sep 26–2025.(Epub ahead of print).
|
|
48
|
Suzuki R, Terashima M, Ishimura A, Meguro-Horike M, Horike SI, Wanna-Udom S, Takino T and Suzuki T: METTL3 contributes to osimertinib resistance in non-small cell lung cancer cell lines by regulating CDC25A and AURKB mRNA stability. Cell Signal. 136:1121562025. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang KT, Yang S, Zhao ZH, Rui CH, Shen SY, Cao D, Guo LN, Tang SH, Chen L, Wang HY and Qiu XY: ALKBH5 deficiency suppresses hepatocarcinogenesis in mice via m(6)A-dependent STAT1 restoration. Acta Pharmacol Sin. 47:209–221. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang L, Ren D, Cai Z, Hu W, Chen Y and Zhu X: O-GlcNAcylated YTHDF2 promotes bladder cancer progression by regulating the tumor suppressor gene PER1 via m(6)A modification. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 50:827–839. 2025.(In Chinese English). PubMed/NCBI
|
|
51
|
Shi Y, Liu J, Lin T, Lu W, Yang L and Huang S: Methyltransferase WTAP participates in shrimp defense against Vibrio parahaemolyticus infection. Dev Comp Immunol. 172:1055102025. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Song M, Lu S, Tang H, Lv K, Li J, Shi J and Xu Y: FTO-engineered extracellular vesicles from bone marrow mesenchymal stem cells ameliorate Staphylococcus aureus-induced osteomyelitis via m6A-dependent suppression of autophagy and pyroptosis. J Biol Eng. 19:1002025. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Vasukutty A, Bhattarai PY and Choi HS: Ebselen suppresses breast cancer tumorigenesis by inhibiting YTHDF1-Mediated c-Fos expression. Int J Mol Sci. 26:94162025. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Du L, Ho BM, Zhou L, Yip YWY, He JN, Wei Y, Tham CC, Chan SO, Schally AV, Pang CP, et al: Growth hormone releasing hormone signaling promotes Th17 cell differentiation and autoimmune inflammation. Nat Commun. 14:32982023. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Cai Z, Liu R, Zhao L, Zhou L, Li Q and He H: Immunological dynamics in PCOS T cell exhaustion TIGIT upregulation regulated by METTL3-Mediated N6 RNA methylation. Am J Reprod Immunol. 94:e701802025. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Abu-Tawil HI, Picavet LW, van Vroonhoven ECN, Bodelón A, Scholman RC, Ter Haar N, Boltjes A, Vastert SJ and van Loosdregt J: Reduced Expression of m(6)A Demethylases FTO and ALKBH5 in monocytes from the site of inflammation in patients with juvenile idiopathic arthritis. Int J Mol Sci. 26:92482025. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sui M, Wu S, Song Q, Chen S and Lu Y: FOXO6, stabilized by METTL3-Mediated m6A modification, accelerates polycystic ovary syndrome progression by transcriptional activation of TXNIP. FASEB J. 39:e711412025. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cehajic-Kapetanovic J, Xue K, Martinez-Fernandez de la Camara C, Nanda A, Davies A, Wood LJ, Salvetti AP, Fischer MD, Aylward JW, Barnard AR, et al: Initial results from a first-in-human gene therapy trial on X-linked retinitis pigmentosa caused by mutations in RPGR. Nat Med. 26:354–359. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Field MG, Kuznetsoff JN, Zhang MG, Dollar JJ, Durante MA, Sayegh Y, Decatur CL, Kurtenbach S, Pelaez D and Harbour JW: RB1 loss triggers dependence on ESRRG in retinoblastoma. Sci Adv. 8:eabm84662022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yin L, Ma C, Hou S and Ma X: Methyltransferase-like (METTL)14-mediated N6-methyladenosine modification modulates retinal pigment epithelial (RPE) activity by regulating the methylation of microtubule-associated protein (MAP)2. Bioengineered. 13:4773–4785. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Appelbaum T, Aguirre GD and Beltran WA: Identification of circular RNAs hosted by the RPGR ORF15 genomic locus. RNA Biol. 20:31–47. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang C, Feng L, Fang W, Zhang C, Zhang W, Zhu W, He Y, Xia Z, Song W and Xia X: Inhibition of Mettl3-mediated m6A RNA modification of HMGCS1 protects retinal ganglion cells from glutamate excitotoxicity-induced ferroptosis in a rat model of glaucoma. Int J Surg. 111:9147–9165. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Xiao B, Zhu Y, Liu M, Chen M, Huang C, Xu D, Wang F, Sun S, Huang J, Sun N and Yang F: Correction: Mir-340-3p-modified bone marrow mesenchymal stem cell-derived exosomes inhibit ferroptosis through METTL3-mediated m6A modification of HMOX1 to promote recovery of injured rat uterus. Stem Cell Res Ther. 15:3572024. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhu F, Feng J, Pan Y, Ouyang L, He T and Xing Y: Mettl3-Mediated N6-methyladenosine modification mitigates ganglion cell loss and retinal dysfunction in retinal ischemia-reperfusion injury by inhibiting FoxO1-mediated autophagy. Invest Ophthalmol Vis Sci. 66:582025. View Article : Google Scholar
|
|
65
|
Stitt AW, Curtis TM, Chen M, Medina RJ, McKay GJ, Jenkins A, Gardiner TA, Lyons TJ, Hammes HP, Simó R and Lois N: The progress in understanding and treatment of diabetic retinopathy. Prog Retin Eye Res. 51:156–186. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Flaxel CJ, Adelman RA, Bailey ST, Fawzi A, Lim JI, Vemulakonda GA and Ying GS: Age-related macular degeneration preferred practice pattern®. Ophthalmology. 127:P1–P65. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
He S, Li W, Wang G, Wang X, Fan W, Zhang Z, Li N and Hou S: FTO-mediated m6A modification alleviates autoimmune uveitis by regulating microglia phenotypes via the GPC4/TLR4/NF-κB signaling axis. Genes Dis. 10:2179–2193. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Mondal AK, Gaur M, Advani J and Swaroop A: Epigenome-metabolism nexus in the retina: Implications for aging and disease. Trends Genet. 40:718–729. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Luo C, Zheng ZG, Zeng MQ, Xu H, Yu XM, Sun D and He DJ: Curcumol targets the FTO/MAFG-AS1 axis to alleviate diabetic retinopathy via epigenetic remodeling and nanodelivery-based microenvironment modulation. World J Diabetes. 16:1070172025. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhu XJ, Jiang XY, Liu WJ, Fan YD, Liu G, Yao S, Sun KX, Chen JY, Lei B and Yang YM: Single-cell sequencing analysis reveals the essential role of the m(6)A reader YTHDF1 in retinal visual function by regulating TULP1 and DHX38 translation. Zool Res. 46:429–445. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Del Amo EM, Rimpelä AK, Heikkinen E, Kari OK, Ramsay E, Lajunen T, Schmitt M, Pelkonen L, Bhattacharya M, Richardson D, et al: Pharmacokinetic aspects of retinal drug delivery. Prog Retin Eye Res. 57:134–185. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wang X, Li X, Zong Y, Yu J, Chen Y, Zhao M, Wu D, Liao Y, Jiang C and Zhu H: Identification and validation of genes related to RNA methylation modification in diabetic retinopathy. Curr Eye Res. 48:1034–1049. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang SB, Nagasaka Y, Argyle D, Nagasaka A, Yerramothu P, Gelfand BD and Ambati J: Targeting the m6A mRNA demethylase FTO suppresses vascular endothelial growth factor release and choroidal neovascularization. Signal Transduct Target Ther. 8:722023. View Article : Google Scholar : PubMed/NCBI
|