Introduction
In 1993, Lee et al identified a non-coding
RNA (ncRNA), lin-42, in C. elegans (1). Subsequently, Fire and Mello found a
sequence-specific gene-silencing mechanism, referred to as RNA
interference (RNAi), which was induced by double-stranded RNA, and
proposed the concept of the regulation of gene expression by ncRNA
in 1998 (2). In 2000, let-73 was
identified as a short-stranded RNA, termed a microRNA (miRNA), that
regulated gene expression, development, cell division and
differentiation, and cell homeostasis. Subsequently, miRNAs have
been found in various organisms, including viruses and plants, and
the number of identified miRNAs continues to rise. A total of 530
human miRNAs were registered (miRecords: http://mirecords.biolead.org/) as of January 2008, and
more than 1,000 miRNAs are thought to exist.
miRNA-mediated RNAi is considered to be
post-transcriptional regulation, a third means of modulation of
gene expression, in addition to transcriptional regulation and
post-translational modification. miRNAs generally bind to the 3′
untranslated region (UTR) of a target messenger RNA (mRNA) via base
pairing and consequently control gene expression specifically by
inhibiting translation and decreasing mRNA stability. In
silico analysis has shown that one miRNA controls hundreds of
targets, and more than 60% of the approximately 20,000 human genes
that code proteins have at least one miRNA target site in the 3′
UTR. However, miRNA target sites are also included in coding
regions and in the 5′ UTR, and almost all mRNAs are regulated by
miRNAs. This suggests that miRNAs play a key role in the generation
of the transcriptome and proteome.
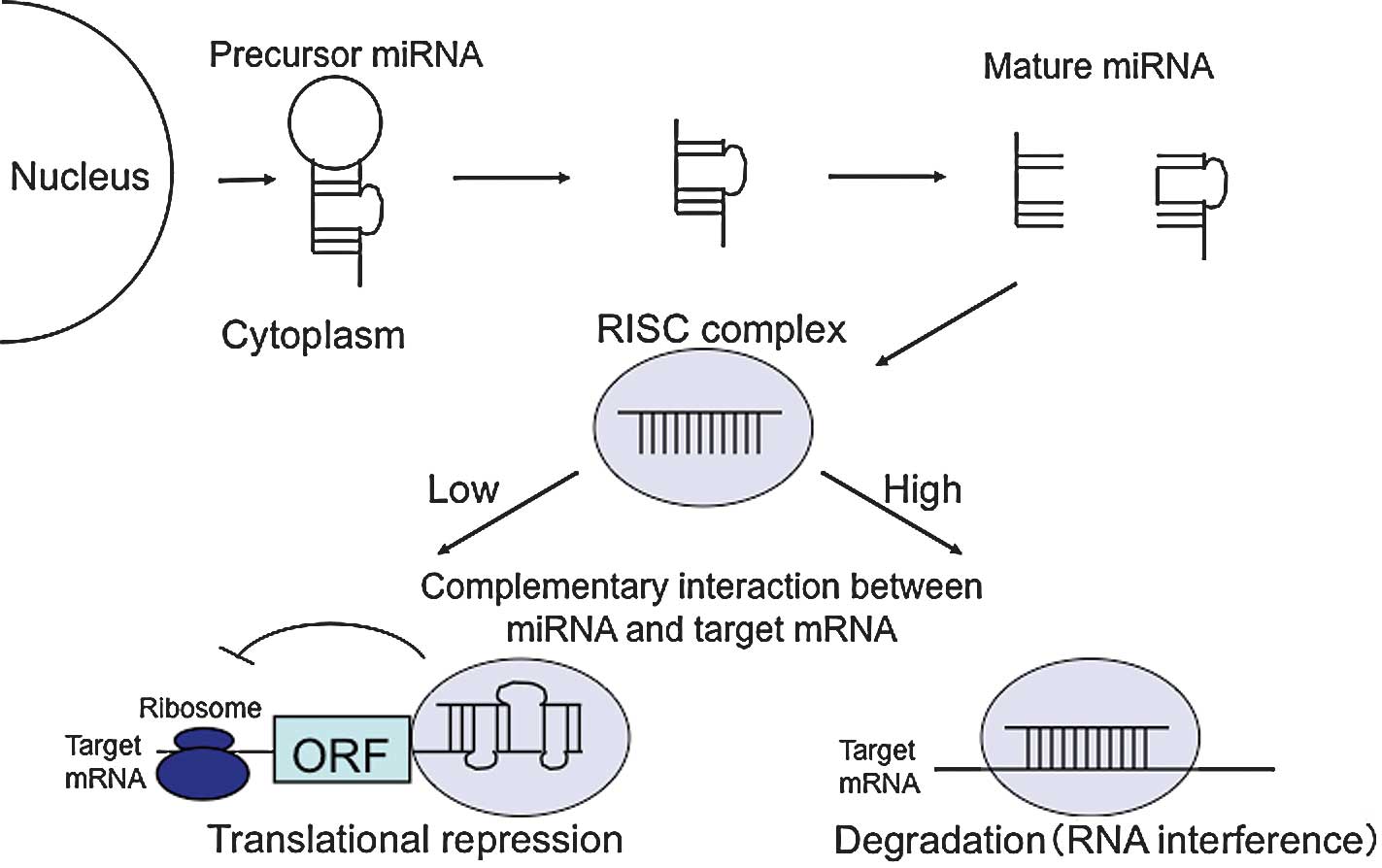
miRNA synthesis starts with the transcription of
miRNA genes to create a pri-miRNA, a long (hundreds or thousands of
nucleotides) precursor with poly(A) sequences. pri-miRNAs are
usually transcribed by RNA polymerase II and then cleaved into one
or a number of pre-miRNAs by the microprocessor complex, which
contains the nuclear RNase III Drosha-DGCR8 complex with a
double-stranded RNA-binding domain. Pre-miRNAs generally have a 70-
to 90-bp stem-loop structure. These molecules are transported from
the nucleus to the cytoplasm by exportin-5. The pre-miRNA hairpin
is cleaved by the Dicer complex, which includes helicase and RNase
III domains, and is processed to an approximately 21-bp
double-stranded miRNA, which is finally converted to a
single-stranded mature miRNA.
The mature miRNA is incorporated into the
RNA-induced silencing complex (RISC), a ribonucleoprotein complex,
to yield the miRISC, which recognizes target mRNA using the miRNA
as a guide and inhibits mRNA translation or cleaves the mRNA.
Cleavage occurs when the miRISC has high complementarity with the
mRNA, whereas inhibition of mRNA translation occurs when
miRISC/mRNA interaction has relatively low complementarity. In
animals, almost all miRNAs inhibit translation, whereas cleavage is
common in plants. Target mRNA in which translation is inhibited is
transported to and degraded in a processing body (P-body), a
granule localized in the cytoplasm (Fig. 1).
miRNA and small interfering RNA (siRNA) guide the
RISC to the target mRNA. However, siRNA requires complete
complementarity with the target, while miRNA functions with a
partial mismatch in base pairing with the target mRNA.
Consequently, one miRNA has numerous targets. As described above,
different modes of inhibition depend on these differences in
complementarity. Translation is inhibited when the base sequence in
the 3′ UTR of a target mRNA has incomplete complementarity with the
miRNA, but the detailed molecular mechanism has yet to be
elucidated.
2. miRNA in carcinogenesis
Relatively early studies on miRNA in cancer revealed
the following important findings. First, numerous miRNA genes are
located in regions with frequent copy number changes due to
amplification and deficit (fragile site) or with chromosomal
instability near an integration site of human papillomavirus type
16, suggesting an association of carcinogenesis with genomic
abnormalities in the primary structure of miRNA genes. Second,
miRNA expression profiles significantly differ among organs and
tissues, and miRNA levels in cancer tissues are generally lower
than those in normal tissues, suggesting that miRNAs act as tumor
suppressors.
It is now known that various miRNAs act in a manner
equivalent to oncogenes or tumor suppressor genes in cancer
development and progression. An association of miRNAs with cancer
stem cells has also gradually become apparent and miRNAs that
directly target oncogenes have been identified. miRNAs generally
decrease mRNA expression levels, and therefore, miRNAs targeting an
oncogene have a tumor suppressor action via control of the cell
cycle through reduction of the gene expression. By contrast,
certain miRNAs have an oncogene-like function, with a high miRNA
expression-inducing oncogenic transformation; i.e., the miRNA acts
similar to an oncogene. Recent studies have shown that the
expression of tumor suppressor miRNAs is regulated by aberrant DNA
methylation, revealing a mechanism in which the miRNA is
inactivated by epigenetic abnormalities (methylation) in a similar
manner to tumor suppressor (22).
miR-124 is a particular miRNA for which an
association with methylation and cancer stem cells has been shown
in detail. miR-124 is located in or near a CpG island in the
chromosomal regions of 8p23.1, 8q12.3 and 20q13.33. miR-124
expression is inhibited by tumor-specific aberrant DNA methylation
in colon cancer HCT116 cells and is reactivated by treatment with
5-aza-2′-deoxycytidine (5-aza-dC). In HCT116 cells, the methyl-CpG
binding protein, MeCP2, and methyl-CpG binding domain protein,
MBD2, are bound to the promoter of the miR-124 gene, whereas no
binding of these proteins occurs in demethylated HCT116 cells
following knockout of DNA methyltransferase DNMT1 and DNMT3B genes
or treatment with 5-aza-dC. The results suggest that promoter
selectivity is due to methylation. The transcriptional product of
CDK6 is an oncogene and CDK6 mRNA is a direct target of miR-124.
Overexpression of miR-124 or miR-137 has also been shown to induce
morphological changes and differentiation into neurons, with an
enhanced expression of neuronal differentiation markers in mouse
neural stem cells and human glioblastoma (3). The studies suggest that miR-124 is
involved in the maintenance of cancer stem cell genotypes, raising
the possibility of treatment using the miRNA targeting of cancer
stem cells.
The expression of miR-127, located in the CpG island
of 14q32.31, is also reactivated by treatment with 5-aza-dC and
inhibited by elevated methylation in prostate and colon cancers
(4). This miRNA is of interest
since it acts as an oncogene in the targeting of BCL6 mRNA
(4). However, the most detailed
study of an oncogene-type miRNA was performed for miR-17-92
(2). Many miRNAs form clusters in
genomes, and miR-17-92 contains the sequence miR-17, miR-18a,
miR-19a, miR-20a, miR-19b-1 and miR-92-1, encoded in this order. In
B-cell lymphomas, the expression of the cluster is enhanced and
induced by the oncogene c-Myc, with studies in mouse models and
cell cultures showing subsequent inhibition of apoptosis and
enhanced cell division, suggesting that miR-17-92 acts as an
oncogene. Of the genes in this cluster, miR-17 and miR-20a target
the E2F1 gene, which is related to the apoptosis response, and the
down-regulation of E2F1 by miR-17-92 inhibits apoptosis. c-Myc
regulates E2F1 transcription and vice versa. Consequently, miR-17-2
forms a feedback loop with c-Myc and E2F1 (5). Expression of miR-17-92 is also
regulated by E2F3. This regulation indicates the presence of a
comprehensive system for the modulation of gene expression, in
which E2F1 activation by c-Myc is inhibited by miR-17-92, resulting
in the inhibition of apoptosis. Therefore, this system is crucial
for the survival of cancer cells. miR-17-92 also targets various
tumor suppressor genes and therefore plays a key role as an
oncogene-type miRNA (6).
The miR-15a/16-1 cluster consists of the miRNAs,
miR-15a and miR-16-1, and is found in the chromosomal region 13q14.
Frequent absence or silencing of this cluster occurs in chronic
lymphocytic leukemia. miR-15a and miR-16-1 inhibit BCL2, an
oncogene that inhibits apoptosis. Therefore, expression of these
miRNAs in leukemia cells induces apoptosis. A recent study showed
that miR-15a/16-1 targets the c-Myb and IGSF4 oncogenes, and a
microarray analysis showed that the miRNA cluster inhibits a number
of cancer-associated genes correlated with cell growth and
anti-apoptosis. Another miRNA, let-7, acts as a tumor suppressor by
inhibiting the oncogenes K-ras and HMGA2, suggesting that miRNAs
also play a key role in the inhibition of oncogenic transformation.
As described below, a reduced let-7 expression has been found in
patients with lung cancer and the let-7 expression level was
correlated with the outcomes in these patients (7).
3. miRNA as a diagnostic biomarker in human
cancer
miRNA expression in tissues is regulated by the
number of miRNAs present in a cell at a specific period of time.
Tissue-specific patterns of miRNA expression have also been shown
and analysis of these patterns may produce potential markers for
identifying the original source of a cancer, which is crucial in
establishing a treatment plan. miRNA analysis may also be useful
for distinguishing the primary lesion from metastases.
miRNA expression may serve as a prognostic
predictor, since the prognosis of cancer is correlated with the
expression levels of numerous miRNAs: has-let-7, has-let-7a-2 and
has-miR-155 in lung cancer; has-miR-125b in hepatocellular
carcinoma; has-miR-21 in breast, gastric, colon and pancreatic
cancers; and has-let-7b and has-miR-205 in head and neck squamous
cell carcinoma. These correlations are purely statistical and the
fundamental underlying mechanisms have yet to be elucidated. One
study showed that methylation of let-7a-3 was related to prognosis
in ovarian cancer. Poor outcomes were found in ovarian cancer
patients with low levels of mRNA for Dicer and Drosha, which are
involved in the maturation of miRNAs. If the miRNA precursor is not
processed efficiently, the mature miRNA-induced gene silencing
mechanism may not function adequately, resulting in abnormal
oncogenic effects. Lung cancer patients with low Dicer expression
levels also have a poor prognosis (8).
Numerous studies have been performed with regards to
the relationship between miRNA and drug resistance in cancer cells.
Meng et al showed that sensitivity to gemcitabine was
increased by inhibiting miR-21 and miR-200b, which have a higher
expression in bile duct cancer than in benign tumors (9). Yang et al showed that miR-21
introduced into ovarian cancer cells activated AKT through PTEN
inhibition and worsened cisplatin resistance (10). The relationships of miRNAs with
sensitivity to doxorubicin in breast cancer and sensitivity to
endocrine therapy have also been examined. Further studies on
multiple drug resistance may facilitate the choice of treatment
strategies based on the expression levels of miRNAs.
Since RNA is degraded by RNase in the serum, it was
initially thought that miRNAs would be absent. However, a number of
miRNAs, including miR-638, were subsequently detected in the serum.
The mechanisms by which miRNAs are released into and stabilized in
the serum have yet to be determined, but miRNAs appear to be
transported selectively based on specific requirements for the
molecules in the serum. Lawrie et al found a correlation
between serum miR-21 levels and the prognosis of patients with
diffuse large B-cell lymphoma (11), and Mitchell et al found high
serum miR-141 levels in patients with prostate cancer (12).
Tanaka et al found that the serum of patients
with acute leukemia included more miRNAs with low expression levels
than patients with high expression levels, in comparison to the
serum of healthy volunteers. In particular, the serum miR-92 level
was significantly reduced, whereas the miR-92 level in leukemia
cells was high (23). To explain
these results, Tanaka et al proposed the hypothesis that
leukemia cells degrade specific miRNAs or promote the uptake of
miR-92 from the blood (23). The
correlation between a specific cancer and a serum miRNA level
suggests the feasibility for diagnostic and prognostic predictions
based on miRNA levels in the blood. This is likely to become a key
area of research, since biomarker miRNAs may permit early
diagnosis, personalized treatment and prognostic prediction.
4. Applications of miRNA in cancer
therapy
Since miRNA expression levels are known to change in
cancer cells, inhibition of the overexpression of an oncogene-type
miRNA or elevation of the underexpression of a tumor
suppressor-type miRNA may have a therapeutic effect. miRNAs are
small RNAs of approximately 22 base pairs, thus, antisense
oligonucleotides are likely to be effective as base
sequence-specific inhibitors. Delivery methods for antisense agents
have improved, and these approaches should facilitate the targeting
of miRNA, including the use of RNA oligonucleotides with chemical
modifications, in order to overcome their lack of in vivo
stability.
Stoffel et al (13) synthesized an anti-miRNA
oligonucleotide (AMO) (an ‘antagomir’) with 2′-O-methyl,
phosphorothioate and cholesterol groups, and showed the efficacy of
this molecule in mice. At 24 h after antagomir-16 was injected in
the caudal vein, the miR-16 level was reduced in all organs except
the brain, indicating that the antagomir did not cross the
blood-brain barrier. However, the percutaneous injection of
antagomir-16 reduced miR-16 in the brain (13). A study on antagomir-122 targeting
liver-specific expression of miR-122 showed that this miRNA is
involved in the regulation of cholesterol biosynthesis (14). These results suggest that antagomirs
are effective for the treatment of diseases based on the targeting
of specific miRNAs.
Locked nucleic acid (LNA) is a
2′-O,4′-C-methylene-linked RNA derivative that is
nuclease-resistant and forms highly stable and specific
double-stranded helices. Therefore, LNA is used as a probe for
hybridization and as a double-dye probe in real-time PCR. LNA is
water-soluble and less cytotoxic than other antisense agents, and
DNA/LNA mixed oligomers retain the advantage of allowing RNase H
activation after hybridization with the target RNA. Use of a
DNA/LNA AMO to target miR-21 overexpression in glioblastoma was
shown to induce apoptosis, and similar targeting of the
overexpression of miR-221 and miR-222 in prostate cancer cells
decreased the clonogenicity of the cells (15).
Tumor suppressor miRNAs are used to supplement miRNA
underexpression as agonists. In cellular studies, the effects of
miRNA have been investigated by transfection with an expression
vector, but small RNAs, including AIV10 and siRNA, are preferable
for use as miRNA-based therapeutic agents. Inhibition of cell
growth has been shown with transfection of sense miR-15/miR-16
oligoRNA in chronic lymphocytic leukemia and transfection of let-7
as dsRNA in lung cancer cells. The design of miR precursors that
allows the sense strand of the miRNA to be efficiently incorporated
into RISC has been reported using chemically modified dsRNA, and
transfection of miR-34a in glioblastoma cells using a pre-miR-34a
precursor was shown to increase apoptosis (16).
It remains to be elucidated whether transfected
miRNAs have equivalent actions to endogenous miRNA. It is unclear
whether exogenous miRNAs have sufficient activity in vivo
and there are numerous concerns about adverse reactions and the
local distribution of miRNA. Furthermore, although the delivery
methods for miRNA (including across the blood-brain barrier) have
been greatly improved, only a few options are applicable in
practice.
5. miRNA in the endometrium
The endometrium is a tissue that undergoes
substantial changes in parallel with the menstrual cycle in humans.
This tissue is changed for ovum implantation and embryo development
by ovary-releasing sex hormones that regulate gene expression.
These changes are histopathologically classified into apoptosis,
cell growth, neovascularization, cell differentiation and tissue
remodeling. The changes resemble an inflammatory response and
changes in the activities of endocrine and paracrine humoral
factors, including growth factors, chemokines, cytokines and
proteases, as well as alterations in the extracellular matrix are
involved. The rapid turnover of the cell cycle is undesirable in
the prevention of carcinogenesis. Intrinsic control of gene
expression by miRNAs is crucial in this environment and various
changes in the expression of miRNAs have been suggested in normal
endometrium, endometriotic tissue, metrorrhagia and endometrial
cancer.
Pan et al investigated miRNA profiles in the
uterine interstitium and granular epithelium and found that 32
miRNAs showed characteristic expression levels (17). In particular, estradiol-17β and
progesterone regulated the expression of miR-20a, miR-21, miR-23,
miR-26a, miR-18a, miR-181a, miR-206 and miR-142-5p. These miRNAs
are involved in the regulation of expression in numerous genes with
key actions in the uterine tissue, including transforming growth
factor β (TGF-β), TGF-β receptor, estrogen receptor, progesterone
receptor and CYP-19A1 (aromatase). A number of inflammation and
cell growth-related factors, including nuclear factor
κ-light-chain-enhancer of activated B cells (NF-κB), are also
regulated by miRNAs in local inflammatory microenvironments in the
uterine tissue. In this context, inhibition of the expression of
tumor necrosis factor α by miR-125b and miR-155 has been analyzed
(17).
6. miRNA and endometrial cancer
miRNA-related findings in endometrial cancer have
not emerged to the same extent as those in gastrointestinal cancer.
However, studies are ongoing in various institutions, including in
Japan (Table I). Boren et al
found that 13 miRNAs (hsa-let-7i, hsa-miR-221, hsa-miR-30c,
hsa-miR-152, hsa-miR-193, hsa-miR-185, hsa-miR-106a, hsa-miR-181a,
hsa-miR-210, hsa-miR-423, hsa-miR-103, hsa-miR-107 and hsa-let-7c)
and 90 mRNAs derived from human endometrial cancer and normal
uterine tissues were involved in endometrial cancer growth, and
proposed that the 13 miRNAs recognized 26 (29%) of the 90 mRNAs as
targets. A total of 11 of these 26 mRNA genes (KCNMB1, IGFBP-6,
ENPP2, TBL1X, CNN1, MYH11, KLF2, TGFB1/1, MYL9, SNCAIP and RAMP1)
were shown to form a network and to be involved in cell growth
(18).
 | Table IEndometrial cancer-associated
miRNAs. |
Table I
Endometrial cancer-associated
miRNAs.
| Up-regulated | Down-regulated | Reference |
|---|
| miR-185 | miR-Let7e | Boren et al
(18) |
| miR-106a | miR-221 | Wu et al
(24) |
| miR-181a | miR-30c | Chung et al
(21) |
| miR-210 | miR-152 | |
| miR-423 | miR-193 | |
| miR-103 | miR-204 | |
| miR-107 | miR-99b | |
| miR-Let7c | miR-193b | |
| miR-205 | | |
| miR-449 | | |
| miR-429 | | |
Myatt et al found that the expression of the
tumor suppressor gene FOXO1 in endometrial cancer cells was
down-regulated in comparison to that in the normal endometrium
(19). Immunohistochemical studies
showed that the level of FOXO1 was highest in the normal
endometrium, lower in endometrial hyperplasia and further reduced
in an endometrial tumor. However, the FOXO1 level is relatively
high in HEC-1B endometrial cancer cells and lower in Ishikawa
endometrial cancer cells. In HEC-1B cells, the levels of miR-9,
miR-27, miR-96, miR-153, miR-182, miR-183 and miR-186 were high,
whereas little expression of miR-29a, miR-128, miR-152 and miR-486
was found. Therefore, this pattern of miRNA expression appears to
produce only a limited inhibition of FOXO1 expression. In Ishikawa
cells, the levels of miR-27, miR-96, miR-128, miR-153, miR-182,
miR-183 and miR-186 were higher than the respective levels in
HEC-1B cells, which may reflect the lower FOXO1 level in Ishikawa
cells compared to HEC-1B cells. Furthermore, in Ishikawa cells,
inhibition of the expression of miR-9, miR-27, miR-96, miR-153,
miR-183 and miR-186 induced FOXO1-dependent G1 cell cycle arrest
and cell death with a gradual decrease of RNAi with FOXO1
expression. The results suggest that miRNA-dependent FOXO1
down-regulation plays a key role in the survival of endometrial
tumor cells.
Myatt et al concluded that the abnormal
expression of miRNAs affect the regulation of apoptosis-associated
genes and plays a specific role in the development of endometrial
cancer (19). This study is notable
as it showed that miRNAs in a gene network developed in
silico inhibited tumor suppressor genes and induced
tumorigenesis. The results also showed that the tumor size is
reduced by inhibiting these miRNAs, which suggests the potential of
targeting of miRNAs as a therapeutic strategy in patients with
endometrial cancer.
Hiroki et al conducted a comparative study of
carcinoma and normal endometrial tissues derived from 21 patients
with serous endometrial adenocarcinoma between January 2001 and
December 2006 and analyzed the relationship of miRNA levels with
clinical outcomes, including clinicopathological features and
survival rate (20). Out of 120
miRNAs that were analyzed, the expression levels of 54, including
miR-101, miR-10b*, miR-152 and miR-29b, were
down-regulated, and those of the remaining 66, including miR-200a,
miR-200b and miR-205, were up-regulated. miR-205 was also
investigated as a predictor for head and neck squamous cell
carcinoma and significant correlations were found between
inhibition of the expression of miR-10b*, miR-29b and
miR-455-5p and vascular invasion (P=0.048, P=0.013 and P=0.032,
respectively). A low expression of miR-101, miR-10b*,
miR-139-5p, miR-152, miR-29b and miR-455-5p was a significant
prognostic factor for poor overall survival (P<0.05) and a low
expression of miR-152, miR-29b and miR-455-5p was a significant
prognostic factor for poor disease-free survival (P<0.05).
Results of the multivariate analysis revealed that a low expression
of miR-152 was an independent risk factor for poor overall survival
(P=0.021), and a low expression of miR-101 and miR-152 was an
independent risk factor for poor disease-free survival (P=0.016 and
P=0.010, respectively). In vitro studies showed that
transfection with miR-101 and miR-152 precursors inhibited cell
growth in serous endometrial adenocarcinoma (P<0.0001 and
P<0.01, respectively), and a strong positive correlation was
found between miR-101 down-regulation and the immune response to
cyclooxygenase-2 (COX-2) (P=0.035). Hiroki et al concluded
that these data showed that insufficient regulation of miRNA
expression was an effective indicator for predicting a poor
prognosis in patients with serous endometrial adenocarcinoma
(20). This study is crucial since
it indicates an actual relationship between prognosis and the miRNA
levels in endometrial cancer.
Chung et al (21) investigated miRNA expression and
clinicopathological features of endometrial cancer in 30 specimens
collected from patients with endometrioid endometrial
adenocarcinoma in Hong Kong. The results were compared to those for
22 normal tissue specimens. Carcinoma was detected in 25 specimens
of stages I and II, and 5 of stage III. High levels of miR-95,
miR-103, miR-106a, miR-151, miR-155, miR-182, miR-183, miR-194,
miR-200a, miR-200c, miR-203, miR-205, miR-210 and miR-223 were
found. A total of 30 miRNAs correlated with features associated
with prognosis, including disease period, invasion into muscle
layers, recurrence and invasion of lymph nodes. A total of 68 genes
were identified as potential targets of the 30 miRNAs. Chung et
al (21) noted that the
expression level of miR-205 in cancer tissue was extremely high, at
27.2-fold that in normal tissues, also noted by Hiroki et al
(20). The miR-205 level showed a
correlation with invasion into muscle layers and recurrence. Chung
et al showed that the transfection of sequences inhibiting
miR-205 into the endometrial cancer cell line RL95-2 inhibited the
expression of miR-205 by 64.9% and increased the expression of the
tumor suppressor gene JPH4 by 14.2% (21). JPH4 shows abnormalities in
endometrioid endometrial adenocarcinoma and these in vitro
and in vivo results suggest that it is a target of miR-205.
The results of Chung et al are significant since they show
the relationship between miRNA levels and clinicopathological
features in endometrioid adenocarcinoma, which accounts for 80–90%
of endometrial cancer.
7. Conclusion
This review evaluated recent findings on miRNA
expression and activities and on the relationship between miRNA and
endometrial cancer. Techniques for diagnosis and treatment based on
miRNA have yet to be established and the majority of results remain
at the in vitro level. However, the utility of miRNAs as
prognostic predictors has been shown. The use of miRNAs for the
treatment of malignant tumors involves delivery of a single miRNA
that may modify the expression of approximately 100 genes. This
type of treatment is significantly different from conventional gene
therapy, in which only one gene is targeted, despite a malignant
tumor being a phenotype of cumulative abnormalities. This
phenomenon is a limitation of conventional gene therapy that may be
overcome by treatment with miRNAs. However, inhibition of the
expression of genes unrelated to carcinogenesis, but with an mRNA
sequence similar to the target gene, may present a difficulty. As
for conventional antitumor agents, a delivery system with
selectivity for local lesions is required for the targeting of
miRNA drugs to focus on the tumor and avoid effects on other
tissues that may cause serious adverse reactions.
References
|
1
|
Lee RC, Feinbaum RL and Ambros V: The
C. elegans heterochronic gene lin-4 encodes small RNAs with
antisense complementarity to lin-14. Cell. 75:843–854. 1993.
|
|
2
|
Fire A, Xu S, Montgomery MK, Kostas SA,
Driver SE and Mello CC: Potent and specific genetic interference by
double-stranded RNA in Caenorhabditis elegans. Nature.
91:806–811. 1998. View
Article : Google Scholar
|
|
3
|
Silber J, Lim DA, Petritsch C, Persson AI,
Maunakea AK, Yu M, Vandenberg SR, Ginzinger DG, James CD, Costello
JF, Bergers G, Weiss WA, Alvarez-Buylla A and Hodgson JG: miR-124
and miR-137 inhibit proliferation of glioblastoma multiforme cells
and induce differentiation of brain tumor stem cells. BMC Med.
6:142008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Saito Y, Liang G, Egger G, Friedman JM,
Chuang JC, Coetzee GA and Jones PA: Specific activation of
microRNA-127 with downregulation of the proto-oncogene BCL6 by
chromatin-modifying drugs in human cancer cells. Cancer Cell.
9:435–443. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Coller HA, Forman JJ and Hegesse-Miller A:
Myc’ed messages: myc induces transcription of E2Fl while inhibiting
its translation via a microRNA polycistron. PLoS Genet.
3:e1462007.
|
|
6
|
He L, Thomson JM, Hemann MT,
Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe
SW, Hannon GJ and Hammond SM: A microRNA polycistron as a potential
human oncogene. Nature. 435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dews M, Homayouni A, Yu D, Murphy D,
Sevignani C, Wentzel E, Furth EE, Lee WM, Enders GH, Mendell JT and
Thomas-Tikhonenko A: Augmentation of tumor angiogenesis by a
Myc-activated microRNA cluster. Nat Genet. 38:1060–1065. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Karube Y, Tanaka H, Osada H, Tomida S,
Tatematsu Y, Yanagisawa K, Yatabe Y, Takamizawa J, Miyoshi S,
Mitsudomi T and Takahashi T: Reduced expression of Dicer associated
with poor prognosis in lung cancer patients. Cancer Sci.
96:111–115. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Meng F, Henson R, Lang M, Wehbe H,
Maheshwari S, Mendell JT, Jiang J, Schmittgen TD and Patel T:
Involvement of human micro-RNA in growth and response to
chemotherapy in human cholangiocarcinoma cell lines.
Gastroenterology. 130:2113–2129. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang N, Kaur S, Volinia S, et al: MicroRNA
microarray identifies Let-7i as a novel biomarker and therapeutic
target in human epithelial ovarian cancer. Cancer Res.
68:10307–10314. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lawrie CH, Gal S, Dunlop HM, Pushkaran B,
Liggins AP, Pulford K, Banham AH, Pezzella F, Boultwood J,
Wainscoat JS, Hatton CS and Harris AL: Detection of elevated levels
of tumour-associated microRNAs in serum of patients with diffuse
large B-cell lymphoma. Br J Haematol. 141:672–675. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mitchell PS, Parkin RK, Kroh EM, et al:
Circulating microRNAs as stable blood-based markers for cancer
detection. Proc Natl Acad Sci USA. 105:10513–10518. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Krützfeldt J, Kuwajima S, Braich R, Rajeev
KG, Pena J, Tuschl T, Manoharan M and Stoffel M: Specificity,
duplex degradation and subcellular localization of antagomirs.
Nucleic Acids Res. 35:2885–2892. 2007.PubMed/NCBI
|
|
14
|
Esau C, Davis S, Murray SF, et al: miR-122
regulation of lipid metabolism revealed by in vivo antisense
targeting. Cell Metab. 3:87–98. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Galardi S, Mercatelli N, Giorda E,
Massalini S, Frajese GV, Ciafrè SA and Farace MG: miR-221 and
miR-222 expression affects the proliferation potential of human
prostate carcinoma cell lines by targeting p27Kip1. J Biol Chem.
282:23716–23724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Welch C, Chen Y and Stallings RL:
MicroRNA-34a functions as a potential tumor suppressor by inducing
apoptosis in neuroblastoma cells. Oncogene. 26:5017–5022. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pan Q and Chegini N: MicroRNA signature
and regulatory functions in the endometrium during normal and
disease states. Semin Reprod Med. 26:479–493. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boren T, Xiong Y, Hakam A, Wenham R, Apte
S, Wei Z, Kamath S, Chen DT, Dressman H and Lancaster JM: MicroRNAs
and their target messenger RNAs associated with endometrial
carcinogenesis. Gynecol Oncol. 110:206–215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Myatt SS, Wang J, Monteiro LJ, Christian
M, Ho KK, Fusi L, Dina RE, Brosens JJ, Ghaem-Maghami S and Lam EW:
Definition of microRNAs that repress expression of the tumor
suppressor gene FOXO1 in endometrial cancer. Cancer Res.
70:367–377. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hiroki E, Akahira J, Suzuki F, Nagase S,
Ito K, Suzuki T, Sasano H and Yaegashi N: Changes in microRNA
expression levels correlate with clinicopathological features and
prognoses in endometrial serous adenocarcinomas. Cancer Sci.
101:241–249. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chung TK, Cheung TH, Huen NY, et al:
Dysregulated microRNAs and their predicted targets associated with
endometrioid endometrial adenocarcinoma in Hong Kong women. Int J
Cancer. 124:1358–1365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Osaki M, Takeshita F and Ochiya T:
MicroRNAs as biomarker and therapeutic drugs in human cancer.
Biomarkers. 13:658–670. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tanaka M, Oikawa K, Takanashi M, Kudo M,
Ohyashiki J, Ohyashiki K and Kuroda M: Down-regulation of miR-92 in
human plasma is a novel marker for acute leukemia patients. PLoS
One. 4:e55322009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu W, Lin Z, Zhang Z and Liang X:
Expression profile of mammalian microRNAs in endometrial carcinoma.
Eur J Cancer Prev. 18:50–55. 2009. View Article : Google Scholar
|