Introduction
Recombinant adeno-associated virus (rAAV),
ehxibiting minimal toxicity and long-term gene expression, is a
promising cancer gene therapy vector (1,2). In
particular, various studies have validated the potential of
rAAV-secreting therapeutic gene products as a gene transfer tool to
treat metastatic cancers (3,4). Among
these rAAV serotypes undergoing development as gene transfer tools,
the rAAV2 vector has been analyzed most extensively, and is
currently employed in clinical gene therapy (3,5).
Recently, we and other groups demonstrated the potential of the
rAAV5 serotype as an effective vector for antitumoral gene delivery
(6,7). Other studies reported the lowest
prevalence of AAV5-neutralizing antibodies and the highest
prevalence of AAV2-neutralizing antibodies in the normal human
population (8,9). These findings thus support the
hypothesis that rAAV5 vectors are beneficial in targeting
therapeutic genes to tumors without the barrier of humoral immunity
in humans.
However, rAAV5-mediated cancer gene therapy has
certain limitations in relation to systemic administration in
vivo. One of the major drawbacks of rAAV5-mediated gene
delivery is the broad tissue tropism of rAAV5, which is not
exclusively limited to tumor tissue. Transgene expression via rAAV5
is observed in tissues including the liver and spleen, unless
vectors are administered directly to the target tissue or organ
(10,11). These data suggest that it is crucial
to further improve the transduction properties of rAAV5 in order to
facilitate the preferential delivery of transgenes to target
tissues.
Consequently, the native tropism of rAAV5 was
modified by genetically inserting various tumor-targeting peptide
sequences into the capsid. The primary determinant of cell/tissue
tropism of rAAV is the binding specificity of the viral capsid to
cellular receptors (12).
Tumor-targeting motifs significantly enhance viral binding affinity
to cell surface proteins, including integrins, sialyl Lewis X
(sLeX), and tenascin C (TnC), which are preferentially expressed in
tumors (13–16). Since the initial attempt by Girod
et al to deliver genes via mutated rAAV2 to cells resistant
to rAAV2 infection (17), a number
of investigators have successfully achieved re-targeted gene
delivery, both in vivo and in vitro, by modifying the
rAAV2 capsid through insertional mutagenesis (18–20).
Currently, a limited number of studies focusing on re-targeted
rAAV5 vectors are available. Thus, the aim of the present study was
to identify the protruding loop regions capable of bearing
tumor-targeting motifs by modeling AAV5 capsid protein structures
using a computational approach with the X-ray crystal structure of
AAV2 (21). Hypothetical atomic
structures of insertion mutants at the expected loop region were
constructed. Using this approach, various rAAV5 mutants with
altered tropism were generated, which preferentially transduced
tumors and subsequently induced antitumoral effects.
Materials and methods
Cell culture, virus preparation and
transduction by rAAV vectors
293T and human cancer cells (hepatocellular SK-Hep1;
cervical HeLa; breast MDA-MB-231 and MDA-MD-435S; glioblastoma
U-251, U87-MG, U118-MG and U-251; and colon HCT-116) were purchased
from the American Type Culture Collection (ATCC; Manassas, VA,
USA). Cancer cells were maintained in Dulbecco's modified Eagle's
medium supplemented with 10% fetal bovine serum, L-glutamine (2
mM), penicillin (100 IU/ml) and streptomycin (50 μg/ml). The
CHO-Lec2 cell line purchased from ATCC was maintained in α-minimum
essential medium supplemented with 10% fetal bovine serum,
penicillin (100 IU/ml) and streptomycin (50 μg/ml).
Table I shows the
primers used to construct plasmids containing genes of mutant rAAV5
capsids. Using pSp72 plasmid containing the Rep gene of AAV2 and
the Cap gene of AAV5 as an original backbone, final constructs were
generated by a series of cloning steps and sequencing conformation.
Self-complementary rAAV vectors were generated as described
elsewhere (6). Titration of rAAVs
prepared using CsCl2 gradient ultracentrifugation was
performed using TaqMan-based Q-PCRs (iQTM Supermix, Bio-Rad,
Hercules, CA, USA).
 | Table IPrimer information used to construct
plasmids containing genes of mutant rAAV5 capsids. |
Table I
Primer information used to construct
plasmids containing genes of mutant rAAV5 capsids.
| Homing peptides | | Sequences of
oligonucleotides (5′ to 3′) |
|---|
| sLeX1 | Forward |
TACAGCTCCCCCGCCACCACTGGCGGAGTCCAGTTCAAC |
| Reverse |
TCTGGGGATCCAGTGGGCGTTATTTGTGCTCACGAAGCG |
| sLeX2 | Forward |
AGCTCCCCCGCCACCGGAGGCACTGCCCCCGCGACCGGCACGTACAACC |
| Reverse |
GTATCTGGGGATCCAGTGGGCTCCGGTGGAGCTCTGGTTGTTGGTGGC |
| RGD1 | Forward |
CGACAGTCACAGCCGGTGGAGCTCTGGTTGTT |
| Reverse |
AGGCGACTGTTTCTGTACTGCCCCCGCGACC |
| RGD2 | Forward |
TCCCTCGACAGTCACAGCCGGTGGAGCTCTGGTTGTT |
| Reverse |
CTGTTTCTGTGGCCTGTCAACTGCCCCCGCGACC |
| TnC1 | Forward |
ATGAAAGCCGCAATCGCAGCCGGTGGAGCTCTGGTT |
| Reverse |
AAACATAAAAGCTGCTTTTGCACTGCCCCCGCGACC |
| TnC2 | Forward |
TATGAAAGCCGGTGGAGCTCTGGTT |
| Reverse |
AACATAAAAGCACTGCCCCCGCGACC |
| TnC3 | Forward |
CAGCGCCGGGCTTTTATGTTTATG |
| Reverse |
AGCCCGGTGTGCTTTTGCACTGCC |
| TnC4 | Forward |
CTTTTATGTTTATGAAAGCCGGTGGAGCTCTGGTT |
| Reverse |
CCCGGCGCTGAGCCCGGTGACTGCCCCCGCGACC |
Cells were infected with various rAAVs, as
previously described (6). To
maximize the transduction of rAAV vectors, adenovirus 5 was
occasionally added. Transduction efficiencies (TEs) and means of
fluorescence intensities (MFIs) were determined using a flow
cytometer (FACSCalibur, Becton-Dickinson, San Jose, CA, USA) and
the CellQuest Plus program.
Computer modeling of capsid protein and
rAAV
Homology models of the capsid structures of
rAAV5-sLeX1, rAAV5- sLeX2 and rAAV5-RGD1 with a RGD-4C insert were
generated on the basis of the crystal structure of AAV2 capsid (PDB
ID code: 1LP3) using the homology modeling program Modeller 9v2
(22). The whole virion from a
single capsid structure model was generated using the CNSsolve
program v1.1 (23). Structural
figures were prepared using PyMol v0.98 (www.pymol.org).
Cell surface marker analysis
Cells were detected with primary antibodies for
αVβ3b and αVβ5 conjugated with FITC (LM609 and P1F6 clones:
Chemicon, Temecula, CA, USA). To measure α2,3-sialic acids on the
cell surface, cells were stained with primary MAL II lectin
conjugated with biotin (Vector Laboratories, Burlingame, CA, USA),
followed by the addition of secondary streptavidin-FITC. The
proportion of immune-stained cells and the MFI of cells was
measured using a flow cytometer.
RGD peptide and anti-tenascin C antibody
competition assays
For the RGD peptide competition assay, cells in
24-well plates were pre-incubated in fresh medium supplemented with
200, 100, 50, 20 or 10 mM active RGDS or RGES peptide (Sigma, St.
Louis, MO, USA) in the presence of 2% FBS for 30 min at room
temperature. Then, rAAV5 (HeLa, MOI 10; U87-MG, 20; and SK-Hep1,
100) or rAAV5-RGD1 (HeLa, MOI 5×103; U87-MG,
5×103; and SK-Hep1, 1×104) mutants expressing
GFP were added in the presence of peptides and adenovirus type 5
for 4 h. For the anti-tenascin C antibody competition assay, cells
in 96-well plates were pre-incubated in fresh medium supplemented
with 1 mg/ml anti-tenascin C IgG (R&D Systems, MAB2138,
Minneapolis, AT, USA) (1:50, 1:20, and 1:10 dilution) with 2% FBS
for 30 min at room temperature. rAAV5 or rAAV5-TnC4 vectors
containing a GFP-expressing cassette (virus titers at 20–50%
transduction) were added in the presence of antibodies. After 2
days, TEs were measured. At least three independent competition
assays were performed. Relative TEs were determined using the
formula: (TE of rAAV with peptides/TE of rAAV only) × 100%.
Cell cytotoxicity assay
U87-MG cells were seeded in a 6-well plate. For 100%
transgene transfer, cells were infected with rAAV5, rAAV5-RGD1 and
rAAV5-TnC4 at MOIs of 200, 20,000 and 50,000, respectively. For 20%
transduction, 5-fold lower MOIs were employed. The following day,
cells were infected with rAAV5 vectors expressing sc39TK for 24 h.
Cells (2×103/well) seeded in a 96-well plate were
treated with various concentrations of the prodrug ganciclovir
(Cymevene®, Roche, Basel, Switzerland) for 5 days.
Relative cell viabilities were determined as described previously
(24).
Statistics
Data were compared between groups using the
Student's unpaired t-test, and values were considered significant
at p<0.05.
Results
Successful generation of rAAV5 vectors
bearing genetically modified capsids
Alignment results with VP1 sequences of AAV2 and 5
previously obtained AAV2s (17,25)
indicate that peptides may be incorporated at positions 444 and 578
of rAAV5 VP1 (data not shown). Using eight different rep/cap
plasmids encoding mutant rAAV5, capsid proteins bearing the eight
peptide sequences (Table II) were
generated. Analysis of the packaging ability of the rAAV5 mutants
revealed that mutant virus production was generally as efficient as
that of wild-type rAAV5. The buoyant densities of mutants within a
narrow range of 1.370 to 1.385 g/ml of the CsCl2
gradient were similar to that of the wild-type virus (1.394±0.007
g/ml). Moreover, transmission electron microscopy data confirmed
that the mutant vectors maintained structural integrity (data not
shown). These findings suggest the possibility of insertional
modification of rAAV5 at positions 444 and 578 in VP1 without
disrupting the general conformation.
| Table IIA summary of the general properties
of the various rAAV5 mutants. |
Table II
A summary of the general properties
of the various rAAV5 mutants.
| Tumor markers | Viruses | Sites | Tumor-targeting
peptides | Titer (VG/ml) |
|---|
| WT | - | - |
1.3×1011 |
| sLeX | sLeX1 | 444 | AHWIPRYSSPAT |
0.7×1011 |
| sLeX2 | 578 |
G-AHWIPRYSSPAT-GG |
1.0×1011 |
| Integrins | RGD1 | 578 | G-CDCRGDCFC |
1.2×1011 |
| RGD2 | 578 |
G-CDCRGDCFC-GLS |
1.0×1011 |
| TnC | TnC1 | 578 |
GCDCG-FHKHK-SCFC |
1.1×1011 |
| TnC2 | 578 | G-FHKHKS |
1.9×1011 |
| TnC3 | 578 |
GCDCG-FHKHKSPALSPV-CFC |
2.4×1011 |
| TnC4 | 578 | G-FHKHKSPALSPV |
2.0×1011 |
rAAV5 containing the RGD motif transduces
a variety of human cancer cells in an integrin-dependent
manner
Various cancer cells expressing αVβ3 or αVβ5
integrins were transduced with rAAV5-RGD1 or rAAV5 expressing GFP.
rAAV5-RGD1 efficiently transduced integrin-positive cells (HeLa,
U87-MG; Fig. 1A, MDA-MB-435S, and
SK-Hep1; data not shown), but not integrin-negative MDA-MB-231
cells. Similar patterns of gene transfer were observed for
rAAV5-RGD2 (data not shown). In contrast, rAAV5 transduced all
cells expressing sialic acid, a major receptor, regardless of the
degree of integrin expression. The αVβ5 integrin-positive and
sialic acid-negative CHO-Lec2 cells were transduced by rAAV5-RGD1,
but not rAAV5 (Fig. 1B).
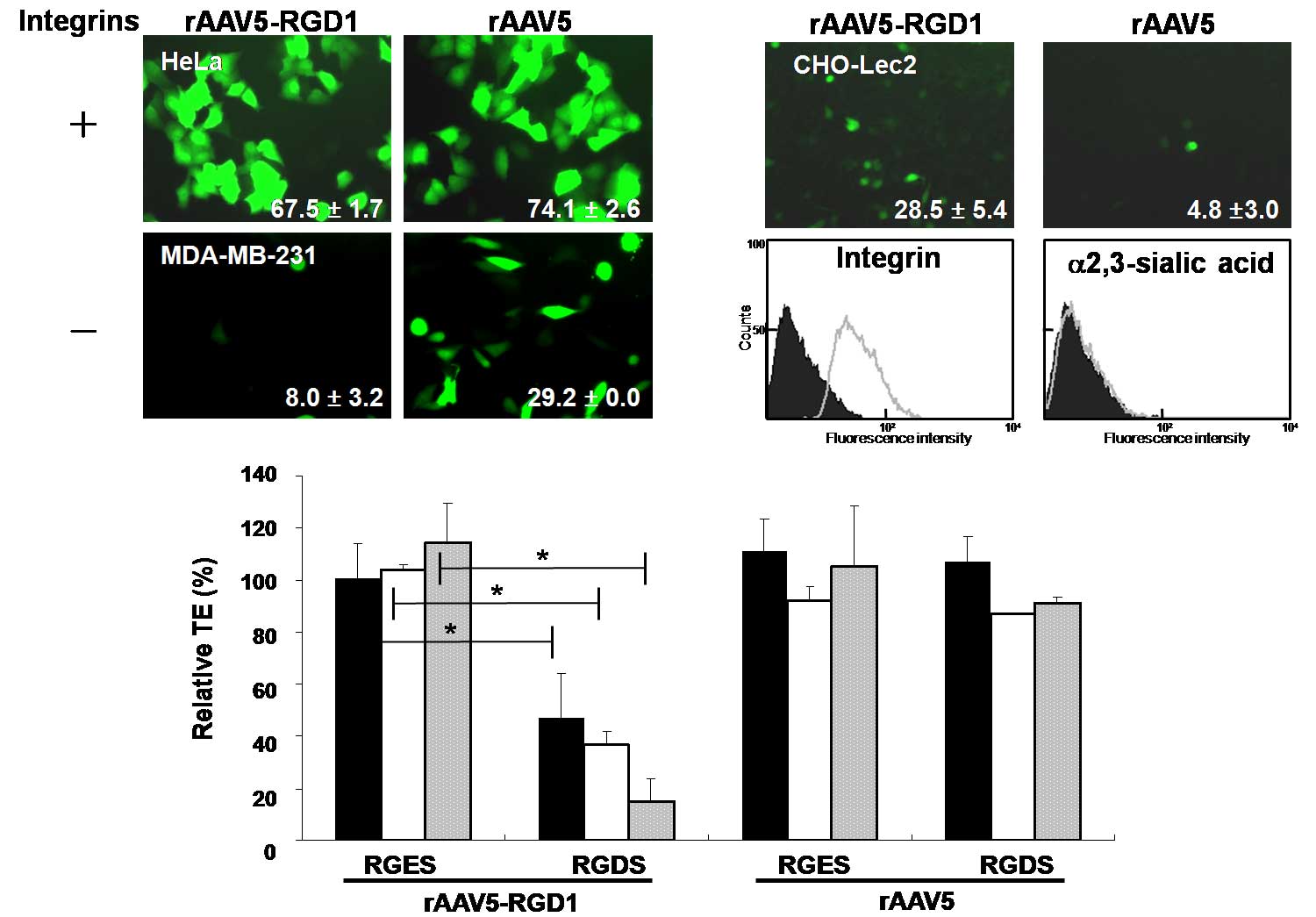 | Figure 1Integrin-dependent transduction of
human cancer cells by rAAV5-RGD1. (A) Integrin-positive cells were
infected with rAAV5-RGD1 (MOI 5,000) or rAAV5 (MOI 20) expressing
GFP. After 48 h, transduction efficiencies (TEs) were analyzed
using a flow cytometer, as indicated in each image (mean±SD, n≥3).
(B) Similarly, CHO-Lec2 cells were treated with rAAV5-RGD1 (MOI
10,000) or rAAV5 (MOI 1,000). Flow cytometry was performed after
incubating cells with either integrin antibody or lectin (n=3). (C)
U87-MG cells were infected with rAAV5 or rAAV5-RGD1 in the
absence/presence of an integrin-binding motif, RGDS peptide (black
bars, 50 μM; white bars, 100 μM; gray bars, 200 μM), and the
relative TE was determined. The non-specific RGES peptide was
employed as a control. rAAV5-RGD1 efficiently transduced cells
expressing integrins (n=3, *p<0.005). |
Gene transfer by rAAV5-RGD1 in integrin-positive
cells was significantly inhibited by the RGDS peptide in a
concentration-dependent manner, but not by the non-specific RGES
peptide (Fig. 1C). As expected, the
TE of the rAAV5 virus was not disrupted by either peptide. These
results collectively indicate that rAAV5 bearing the RGD motif
preferentially transduces human cancer cells expressing integrins,
regardless of the presence of sialic acid, the native receptor of
the wild-type virus.
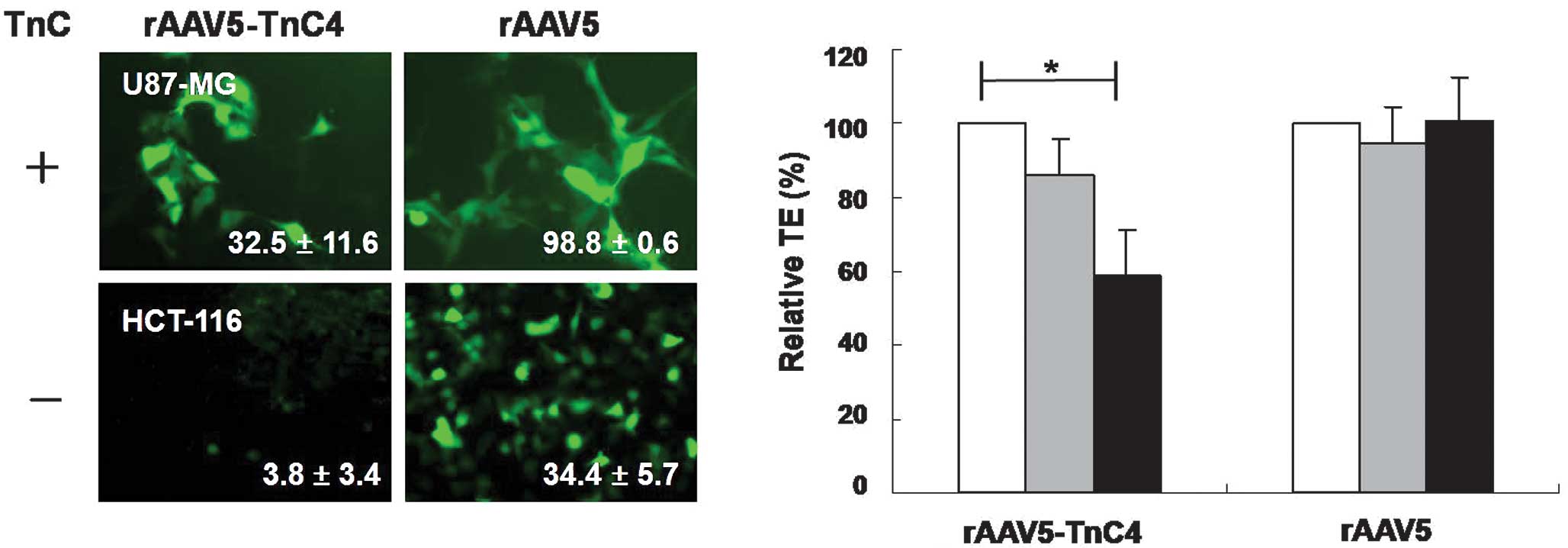
rAAV5-TnC variants preferentially
transduce TnC-expressing cells
The TEs of rAAV5-TnC mutants measured in TnC-
positive or -negative human cancer cells were dependent on TnC
expression. rAAV5-TnC4 efficiently transduced TnC-positive U87-MG,
but not TnC-negative HCT-116 (Fig.
2A). The anti-TnC antibody specifically decreased the
transduction level of rAAV5-TnC4 (Fig.
2B). The TEs of rAAV5-TnC4 in U87-MG cells decreased to
13.9±10.1 and 40.9±12.0% at 1:50 and 1:20 antibody dilutions,
respectively, compared to those of the mock-treated group. In
contrast, TEs of rAAV5 were not altered in the presence of anti-TnC
antibody. The data indicate that rAAV5-TnC specifically infects
human cancer cells in a TnC-dependent manner. In conjunction with
data from Fig. 2, the results
suggest that the newly inserted peptides into the 578 amino acid
position of rAAV5 VP1 facilitate recognition of their corresponding
surface receptors and enhance virus targeting.
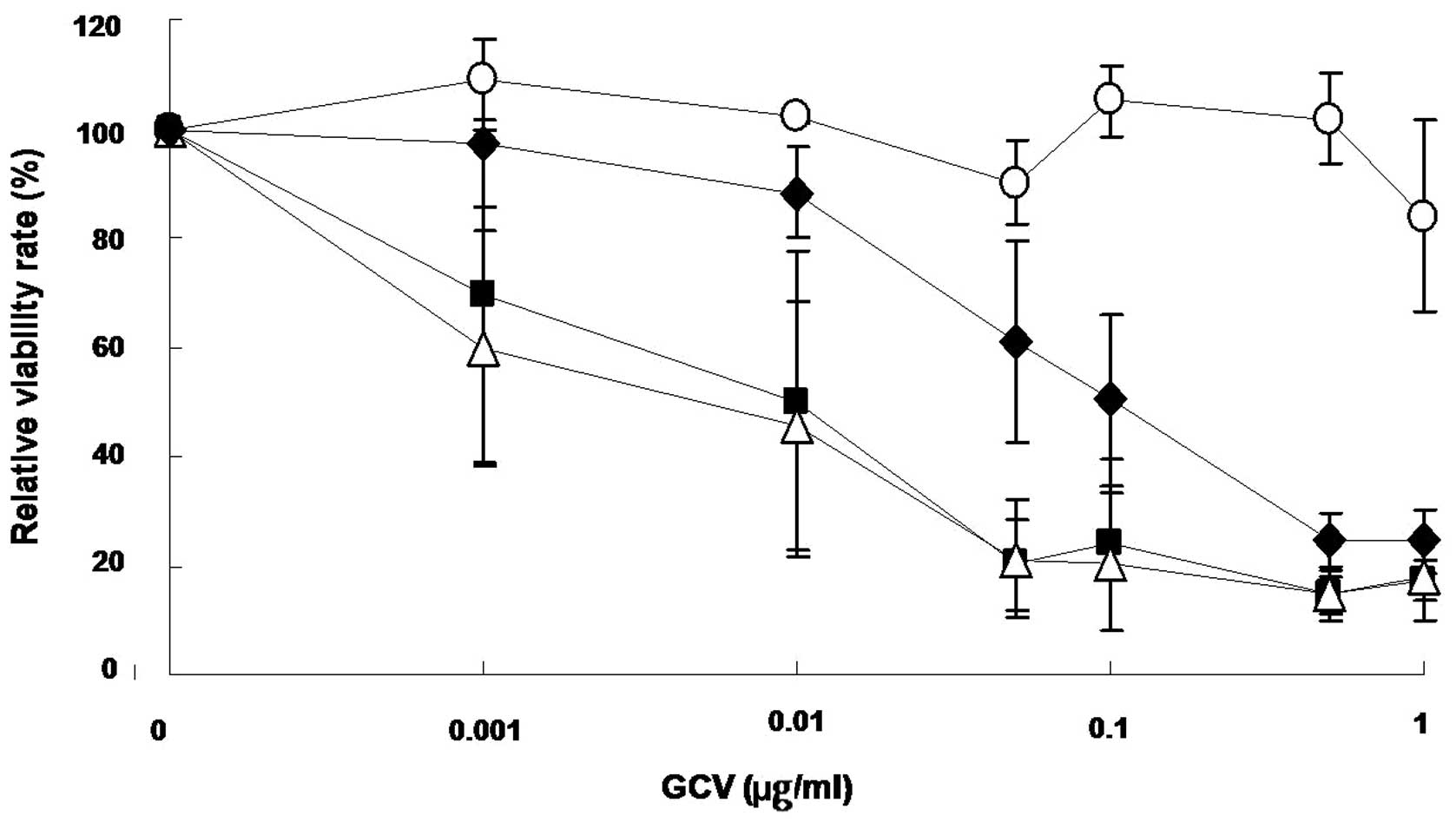
Genetically re-targeted rAAV5s expressing
sc39TK induces antitumoral effects
To assess the therapeutic effects of re-targeted
rAAV5, we utilized the herpes simplex type thymidine kinase
(HSV-sc39TK) gene/prodrug ganciclovir (GCV) system. sc39TK is a
splicing-corrected version of sr39TK, a mutant HSV-TK with
increased affinity for GCV or acyclovir (26,27).
The sc39TK gene was delivered to U87-MG cells via rAAV5-RGD1,
rAAV5-TnC4 and rAAV5 (Fig. 3). The
sc39TK gene transfer induced selective cytotoxicity in the presence
of GCV, while GFP did not. Thus, it appears that re-targeted
antitumoral gene transfer by mutant rAAV5s induces cytotoxicity in
transduced tumor cells. Moreover, the degree of antitumoral effects
differs with various rAAV5 mutants, possibly due to differences in
gene transfer efficacy.
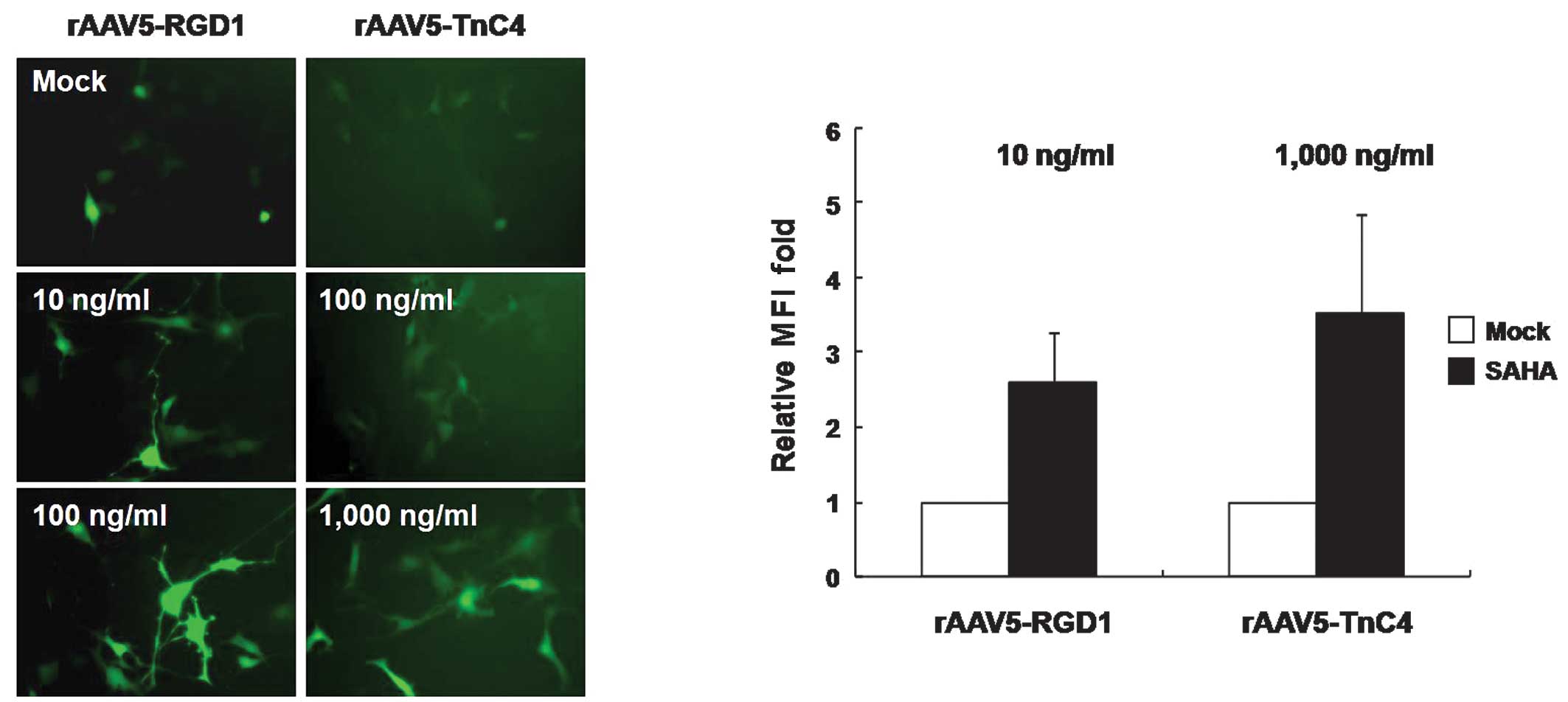
The HDAC inhibitor enhances gene delivery
efficacy of re-targeted rAAV5 vectors
To further enhance rAAV5-RGD1 or -TnC4-mediated gene
expression in target cancer cells, various concentrations of the
antitumoral HDAC inhibitor, suberoylanilide hydroxamic acid (SAHA),
were added prior to transduction with rAAV5-RGD1 and -TnC4.
Enhanced GFP gene expression was observed with the two rAAV5
mutants in a dose-dependent manner following SAHA treatment
(Fig. 4A). Additionally, SAHA
enhanced the intensity of GFP expression per cell, as noted in the
MFI values (Fig. 4B). In other
words, a similar degree of gene expression may be achieved at 10
times lower the total particle concentrations if SAHA is
introduced. Therefore, a combination of SAHA treatment and
tumor-targeted rAAV5 vectors encoding antitumoral genes may
significantly enhance therapeutic effects.
Discussion
For genetic re-targeting of rAAV against its natural
tropism, rAAV5 has considerable potential as an attractive backbone
due to favorable characteristics, such as cancer tropism, a low
level of neutralizing antibodies in human populations, and superior
virus production yield over other serotypes (6,9,28).
Limited information is available on rAA5 modification at present
(29), although several studies
have focused on genetic modification of rAAV1 and 2 capsids
(20,30,31).
In the present study, it was demonstrated that i) rAAV5 vectors may
be genetically modified within the capsid region, particularly at
VP1 positions 444 and 578, ii) mutants bearing insertion sequences
within a certain size limit are efficiently produced with no loss
of viral properties, iii) the insertion of tumor-binding motifs in
the rAAV5 capsid protein suppresses the native tropism of the rAAV5
vector, and iv) tumor-targeted rAAV5 vectors are capable of
selectively transducing human cancer cells, depending on their own
target receptor properties.
αV integrin, sLeX and TnC are often over-expressed
on the surfaces of a wide variety of tumor cells (14–16).
Homing peptides of various sizes ranging from 7 to 20 amino acids
were successfully inserted, which included sequences responding to
tumor surface markers. Introduction of these peptides did not alter
the physical or biological properties of viruses. The data suggest
that rAAV5 tolerates up to 20 amino acids in terms of insertion
size. Previously, Grifman et al performed an alignment of
the AAV1-5 capsid sequences, and demonstrated that loops III and IV
including Arg at positions 447 and 587 of rAAV2, respectively,
could be freely modified to alter rAAV2 tropism without losing
virus replication ability (18).
Consistent with earlier findings, the two loops containing amino
acids 444 and 578 of rAAV5 were genetically altered with no effects
on rAAV5 production ability.
rAAV5 mutants harboring peptide sequences with
affinities for specific tumor markers facilitate recognition of the
corresponding cell surface receptors, and not the native sialic
acid receptor. Along with systemic mutation analyses (25), the atomic structure of AAV2 strongly
suggests that the primary receptor binding motif is located in the
IV loop at position 587 near the 3-fold proximal peak. Another loop
III, responsible for receptor recognition, resides on the distal
floor of the peak. Walters et al suggested that three amino
acids (I528, N546 and M547) located in the depression at the 2-fold
axis are involved in the receptor binding of rAAV5 (32). However, we observed significant
decreases in TE through the introduction of homing peptides at
positions 444 and 578. Thus, two regions near positions 444 and 578
located on the 3-fold proximal peak are potentially responsible for
rAAV5 binding to α2,3-linked sialic acid on host cells. Further
studies are required to elucidate the receptor binding phenotype of
rAAV5.
Cytotoxicity of rAAV5-RGD1 and -TnC4-mediated sc39TK
expression in conjunction with GCV treatment suggests that genetic
modification of rAAV5 at the capsid provides a reasonable basis for
transductional selectivity and specificity of these mutant viruses
for cancer cells. However, our data show that the TEs of
re-targeted rAAV5 vectors were not as high as expected, which
limits their practical utility. To overcome this limitation, we
further investigated the effectiveness of combination treatment
with chemical compounds, including anti-cancer agents or protease
inhibitors (33,34). Several HDAC inhibitors significantly
enhance adenovirus-mediated and rAAV2-mediated transduction
efficiencies (34). SAHA, a HDAC
inhibitor, was approved by the US Food and Drug Administration for
treatment of cutaneous T-cell lymphoma in 2006 (35). As expected, SAHA clearly improved
re-targeted rAAV5-mediated gene transfer in a dose-dependent
manner. HDAC inhibitors may enhance the utility of re-targeted
rAAV5-mediated therapeutic antitumoral gene transduction
strategies, promoting synergistic therapeutic effects.
The transduction potencies of genetically modified
rAAV5s varied significantly in different mutants. The TE of
rAAV5-RGD1 was the most superior, followed by rAAV5-TnC3 and -TnC4
(data not shown). In contrast, mutants harboring the sLeX-binding
motif did not show any specific transducing ability in
sLeX-positive cancer cells (data not shown). Discrepancies in the
TEs of rAAV5 mutants may primarily be attributed to differences in
binding affinity to their receptors. Trafficking steps, such as
inefficient endocytic processes or premature virus degradation by
proteasomes following virus entry, may be another reason for
variations in the TEs of mutant viruses. Recent studies have shown
that the mutation of a surface-exposed tyrosine of rAAV2, 8 and 9
enhances transduction, both in vivo and in vitro,
through the evasion of tyrosine phosphorylation and prevention of
proteasome-mediated degradation (36). Accordingly, similar types of
tyrosine mutations in rAAV5 may increase the TE by allowing mutant
rAAV5 to escape from the ubiquitin-proteasome system. We are
currently investigating this possibility by generating various
mutations at the possible ubiquitin recognition sites.
Acknowledgements
This study was partly supported by grants to H. Lee
from the Korea Science and Engineering Foundation (M1053404002) and
the National Research Foundation of Korea (NRF-2010-R13-0029521),
Republic of Korea.
References
|
1
|
Hacker UT, Wingenfeld L, Kofler DM,
Schuhmann NK, Lutz S, Herold T, King SB, Gerner FM, Perabo L,
Rabinowitz J, McCarty DM, Samulski RJ, Hallek M and Buning H:
Adeno-associated virus serotypes 1 to 5 mediated tumor cell
directed gene transfer and improvement of transduction efficiency.
J Gene Med. 7:1429–1438. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li C, Bowles DE, van Dyke T and Samulski
RJ: Adeno-associated virus vectors: potential applications for
cancer gene therapy. Cancer Gene Ther. 12:913–925. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Daya S and Berns KI: Gene therapy using
adeno-associated virus vectors. Clin Microbiol Rev. 21:583–593.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Higano CS, Corman JM, Smith DC, Centeno
AS, Steidle CP, Gittleman M, Simons JW, Sacks N, Aimi J and Small
EJ: Phase 1/2 dose-escalation study of a GM-CSF-secreting,
allogeneic, cellular immunotherapy for metastatic
hormone-refractory prostate cancer. Cancer. 113:975–984. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mueller C and Flotte TR: Clinical gene
therapy using recombinant adeno-associated virus vectors. Gene
Ther. 15:858–863. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee HS, Shin OK, Kim SJ, Lee WI, Jeong S,
Park K, Choe H and Lee H: Efficient gene expression by
self-complementary adeno-associated virus serotype 2 and 5 in
various human cancer cells. Oncol Rep. 18:611–616. 2007.PubMed/NCBI
|
|
7
|
Thorsen F, Afione S, Huszthy PC, Tysnes
BB, Svendsen A, Bjerkvig R, Kotin RM, Lonning PE and Hoover F:
Adeno-associated virus (AAV) serotypes 2, 4 and 5 display similar
transduction profiles and penetrate solid tumor tissue in models of
human glioma. J Gene Med. 8:1131–1140. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Blacklow NR, Hoggan MD, Sereno MS, Brandt
CD, Kim HW, Parrott RH and Chanock RM: A seroepidemiologic study of
adenovirus-associated virus infection in infants and children. Am J
Epidemiol. 94:359–366. 1971.PubMed/NCBI
|
|
9
|
Halbert CL, Miller AD, McNamara S, Emerson
J, Gibson RL, Ramsey B and Aitken ML: Prevalence of neutralizing
antibodies against adeno-associated virus (AAV) types 2, 5, and 6
in cystic fibrosis and normal populations: Implications for gene
therapy using AAV vectors. Hum Gene Ther. 17:440–447. 2006.
View Article : Google Scholar
|
|
10
|
Gao G, Vandenberghe LH, Alvira MR, Lu Y,
Calcedo R, Zhou X and Wilson JM: Clades of adeno-associated viruses
are widely disseminated in human tissues. J Virol. 78:6381–6388.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nathwani AC, Gray JT, McIntosh J, Ng CY,
Zhou J, Spence Y, Cochrane M, Gray E, Tuddenham EG and Davidoff AM:
Safe and efficient transduction of the liver after peripheral vein
infusion of self-complementary AAV vector results in stable
therapeutic expression of human FIX in nonhuman primates. Blood.
109:1414–1421. 2007. View Article : Google Scholar
|
|
12
|
Berns KI and CRP: Parvoviridae. Field's
Virology. Knipe DM and PMH: Lippincott Williams & Wilkins;
Philadelphia: 2007
|
|
13
|
Cruet-Hennequart S, Maubant S, Luis J,
Gauduchon P, Staedel C and Dedhar S: α(v) integrins regulate cell
proliferation through integrin-linked kinase (ILK) in ovarian
cancer cells. Oncogene. 22:1688–1702. 2003.
|
|
14
|
Moschos SJ, Drogowski LM, Reppert SL and
Kirkwood JM: Integrins and cancer. Oncology (Williston Park).
21:13–20. 2007.
|
|
15
|
Wiksten JP, Lundin J, Nordling S, Lundin
M, Kokkola A, von Boguslawski K and Haglund C: Tenascin-C
expression correlates with prognosis in gastric cancer. Oncology.
64:245–250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamaguchi A, Ding K, Maehara M, Goi T and
Nakagawara G: Expression of nm23-H1 gene and Sialyl Lewis X antigen
in breast cancer. Oncology. 55:357–362. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Girod A, Ried M, Wobus C, Lahm H, Leike K,
Kleinschmidt J, Deleage G and Hallek M: Genetic capsid
modifications allow efficient re-targeting of adeno-associated
virus type 2. Nat Med. 5:1052–1056. 1999. View Article : Google Scholar
|
|
18
|
Grifman M, Trepel M, Speece P, Gilbert LB,
Arap W, Pasqualini R and Weitzman MD: Incorporation of
tumor-targeting peptides into recombinant adeno-associated virus
capsids. Mol Ther. 3:964–975. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
White AF, Mazur M, Sorscher EJ, Zinn KR
and Ponnazhagan S: Genetic modification of adeno-associated viral
vector type 2 capsid enhances gene transfer efficiency in polarized
human airway epithelial cells. Hum Gene Ther. 19:1407–1414. 2008.
View Article : Google Scholar
|
|
20
|
Work LM, Buning H, Hunt E, Nicklin SA,
Denby L, Britton N, Leike K, Odenthal M, Drebber U, Hallek M and
Baker AH: Vascular bed-targeted in vivo gene delivery using
tropism-modified adeno-associated viruses. Mol Ther. 13:683–693.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie Q, Bu W, Bhatia S, Hare J,
Somasundaram T, Azzi A and Chapman MS: The atomic structure of
adeno-associated virus (AAV-2), a vector for human gene therapy.
Proc Natl Acad Sci USA. 99:10405–10410. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sali A and Blundell TL: Comparative
protein modelling by satisfaction of spatial restraints. J Mol
Biol. 234:779–815. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brunger AT, Adams PD, Clore GM, DeLano WL,
Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu
NS, Read RJ, Rice LM, Simonson T and Warren GL: Crystallography
& NMR system: A new software suite for macromolecular structure
determination. Acta Crystallogr D Biol Crystallogr. 54:905–921.
1998.
|
|
24
|
Chang JW, Lee H, Kim E, Lee Y, Chung SS
and Kim JH: Combined antitumor effects of an adenoviral cytosine
deaminase/thymidine kinase fusion gene in rat C6 glioma.
Neurosurgery. 47:931–938; discussion 938–939, 2000.
|
|
25
|
Wu P, Xiao W, Conlon T, Hughes J,
Agbandje-McKenna M, Ferkol T, Flotte T and Muzyczka N: Mutational
analysis of the adeno-associated virus type 2 (AAV2) capsid gene
and construction of AAV2 vectors with altered tropism. J Virol.
74:8635–8647. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Black ME, Newcomb TG, Wilson HM and Loeb
LA: Creation of drug-specific herpes simplex virus type 1 thymidine
kinase mutants for gene therapy. Proc Natl Acad Sci USA.
93:3525–3529. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Blumenthal M, Skelton D, Pepper KA, Jahn
T, Methangkool E and Kohn DB: Effective suicide gene therapy for
leukemia in a model of insertional oncogenesis in mice. Mol Ther.
15:183–192. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Grimm D, Kay MA and Kleinschmidt JA:
Helper virus-free, optically controllable, and two-plasmid-based
production of adeno-associated virus vectors of serotypes 1 to 6.
Mol Ther. 7:839–850. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Arnold GS, Sasser AK, Stachler MD and
Bartlett JS: Metabolic biotinylation provides a unique platform for
the purification and targeting of multiple AAV vector serotypes.
Mol Ther. 14:97–106. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stachler MD and Bartlett JS: Mosaic
vectors comprised of modified AAV1 capsid proteins for efficient
vector purification and targeting to vascular endothelial cells.
Gene Ther. 13:926–931. 2006.PubMed/NCBI
|
|
31
|
White K, Buning H, Kritz A, Janicki H,
McVey J, Perabo L, Murphy G, Odenthal M, Work LM, Hallek M, Nicklin
SA and Baker AH: Engineering adeno-associated virus 2 vectors for
targeted gene delivery to atherosclerotic lesions. Gene Ther.
15:443–451. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Walters RW, Agbandje-McKenna M, Bowman VD,
Moninger TO, Olson NH, Seiler M, Chiorini JA, Baker TS and Zabner
J: Structure of adeno-associated virus serotype 5. J Virol.
78:3361–3371. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim SJ, Nam YR, Shin O, Choi J, Lee B,
Chang JW, Kwon YK, Park K and Lee H: Treatment with hydroxyurea and
tyrphostin-1 significantly improves the transduction efficiency of
recombinant adeno-associated viruses in human cancer cells. Oncol
Rep. 14:1475–1479. 2005.PubMed/NCBI
|
|
34
|
Okada T, Uchibori R, Iwata-Okada M,
Takahashi M, Nomoto T, Nonaka-Sarukawa M, Ito T, Liu Y, Mizukami H,
Kume A, Kobayashi E and Ozawa K: A histone deacetylase inhibitor
enhances recombinant adeno-associated virus-mediated gene
expression in tumor cells. Mol Ther. 13:738–746. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Marks PA and Breslow R: Dimethyl sulfoxide
to vorinostat: development of this histone deacetylase inhibitor as
an anticancer drug. Nat Biotechnol. 25:84–90. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Petrs-Silva H, Dinculescu A, Li Q, Min SH,
Chiodo V, Pang JJ, Zhong L, Zolotukhin S, Srivastava A, Lewin AS
and Hauswirth WW: High-efficiency transduction of the mouse retina
by tyrosine-mutant AAV serotype vectors. Mol Ther. 17:463–471.
2009. View Article : Google Scholar : PubMed/NCBI
|