Introduction
The 14-3-3 protein family, which comprises at least
seven isoforms, is a class of highly conserved proteins involved in
regulating signal transduction pathways, cellular proliferation,
differentiation and survival (1–5). Among
the 14-3-3 proteins, 14-3-3σ is the isoform most directly linked to
cancer and was initially identified as a human mammary
epithelium-specific marker (6). The
expression levels of 14-3-3σ are significantly lower in breast
cancer tissues than those in normal breast tissues, which may be
due to the hypermethylation of CpG islands in the 14-3-3σ gene
promoter (7–13).
14-3-3σ appears to be regulated by different
mechanisms following DNA damage, which contributes directly to
cancer development. The major regulator of 14-3-3σ is the tumor
suppressor p53 (14). Upon DNA
damage, p53 becomes dephosphorylated and is able to bind the
promoter region 1.8 kb upstream of the 14-3-3σ transcription start
site. The subsequent activation and increased expression of 14-3-3σ
leads to the arrest of G2/M, allowing sufficient time for DNA
repair (15–17). Yang et al demonstrated that
14-3-3σ stabilizes p53 and enhances its transcriptional activity
through the interaction with p53, suggesting that 14-3-3σ has a
positive feedback effect on p53 (18). However, p53 is mutated in over 50%
of all human cancers (19–21). In our previous study, 14-3-3σ was
found to be a direct transcriptional target of p73 and enhanced the
p73-mediated transcriptional activation as well as pro-apoptotic
activity (22). Thus, p73 may
functionally replace p53 to induce 14-3-3σ in p53-deficient breast
cancer cells (23). It is likely
that the p73/14-3-3σ pathway plays an important role in the
regulation of DNA damage-induced apoptosis in certain breast cancer
cells bearing p53.
In the present study, we aimed to determine the
possible link between the expression of p73 and 14-3-3σ in the
clinical breast specimens and the regulating mechanism of
p73/14-3-3σ in vivo.
Materials and methods
Clinical specimens
A total of 132 samples, 66 primary breast cancer
specimens and paired tumor-free breast specimens, analyzed in our
study were obtained from the same patients who had invasive breast
cancer and underwent surgical treatment at the Breast Center, the
Fourth Clinical Hospital of Hebei Medical University, China, in
2007. The patients, aged between 30 and 76 years, did not undergo
preoperative adjuvant chemotherapy and radiotherapy. Following
surgery, all 66 specimens were sent to the pathology department of
the hospital to be fixed and paraffin-embedded for routine
immunohistochemistry (IHC) analysis. Clinical data were collected
retrospectively from patient files and pathology reports, including
clinical stages, tumor size, pathological types, histological
grading, status of axillary nodes, ER status and c-erbB2 status.
Patients provided written informed consent prior to enrollment.
Immunohistochemistry
Sections (5 μm) were deparaffinized from the
formalin-fixed and paraffin-embedded tissue blocks. The sections
were then heated in a microwave oven for 5 min in 10 mmol/l
Na-citric buffer (pH 6.0) for antigen retrieval and washed with
phosphate-buffered saline (PBS, pH 7.2). The sections were immersed
in 0.3% hydrogen peroxide in methanol for 20 min to suppress
endogenous peroxidase activity. After washing with PBS, the
sections were incubated in 1:10 diluted normal goat serum at room
temperature in a humidified chamber for 30 min to prevent
non-specific immuno globulin binding. The sections were then
treated with the monoclonal anti-p73 (Ab-4, NeoMarkers, Fremont,
CA, USA) or with polyclonal anti-14-3-3σ (C-18, Santa Cruz
Biotechnology, Santa Cruz, CA, USA) antibody at 4°C overnight.
Normal IgG, which replaced the primary antibody, served as the
control. A streptoavidin-biotinylated horseradish peroxidase-based
detection system was used to reveal specific binding. Sections were
counterstained with hematoxylin for light microscopic review and
evaluation.
p73 and 14-3-3σ were always positively detected in
both the cytoplasm and the nucleus of primary breast cancer
specimens and tumor-free breast specimens. Immunoreactivity was
measured for p73 and 14-3-3σ and scored in the following way: -, no
positive cells (negative); +, <20% positive cells (‘mild
reaction’); ++, 21–50% positive cells (‘moderate reaction’); and
+++, >50% positive cells (‘strong reaction’). The
immunoreactivity scores for p73 were denoted as either ‘negative’
or ‘positive’, with positive including mild, moderate and strong
reactions. The immunoreactivity scores for 14-3-3σ were defined as
either ‘negative’ or ‘positive’, with positive including moderate
and strong reactions. The percentage of positive cells and staining
intensity were scored by 2 independent observers.
Cell culture and cells transfection in
plasmid
Human breast cancer-derived MDA-MB-231 cells were
maintained in Dulbecco’s modified Eagle’s medium supplemented with
10% heat-inactivated fetal bovine serum (Invitrogen, Carlsbad, CA,
USA), 50 units of penicillin and 50 g/ml streptomycin. Cells were
grown at 37°C in a water-saturated atmosphere of 5% CO2
in air. Cells were transfected with empty, p53 expression, p73
expression, 14-3-3σ expression, p53 expression plasmid plus 14-3-3σ
expression plasmid or with p73 expression plasmid plus 14-3-3σ
expression plasmid using a Lipofectamine 2000 transfection reagent
according to the manufacturer’s recommendations (Invitrogen). The
expression plasmids were provided by Professor Nakagawara of the
Chiba Cancer Center.
RNA extraction and RT-PCR
Total RNA of cells was extracted using TRIzol
reagent (Invitrogen) according to the manufacturer’s instructions.
The RNA concentration was routinely measured by a spectrophotometer
and its quality was evaluated by visualization following agarose
gel electrophoresis and ethidium bromide staining. Total RNA (1μg)
was used to generate the first strand cDNA using random primers and
SuperScript II reverse transcriptase (Invitrogen). Reverse
transcription was carried out at 42°C for 1 h. The resultant cDNAs
were amplified by PCR-based strategy using rTaq DNA polymerase
(Takara, Ohtsu, Japan). cDNAs of p53, p73,
14-3-3σ and GAPDH were amplified by using primers as
shown in Table I. The expression of
GAPDH was measured as an internal control. PCR products were
separated on 2% agarose gel electrophoresis and visualized by
ethidium bromide staining.
 | Table IPrimer pairs used for amplification
reactions. |
Table I
Primer pairs used for amplification
reactions.
| Genes | Primers | Annealing (°C) | Cycles |
|---|
| p53 | F:
5′-CTGCCCTCAACAAGATGTTTTG-3′
R: 5′-CTATCTGAGCAGCGCTCATGG-3′ | 58 | 28 |
| p73 | F:
5′-TCTGGAACCAGACAGCACCT-3′
R: 5′-GTGCTGGACTGCTGGAAAGT-3′ | 58 | 28 |
| 14-3-3σ | F:
5′-GAGCGAAACCTGCTCTCAGT-3′
R: 5′-CTCCTTGATGAGGTGGCTGT-3′ | 58 | 28 |
| GAPDH | F:
5′-ACCTGACCTGCCGTCTAGAA-3′
R: 5′-TCCACCACCCTGTTGCTGTA-3′ | 58 | 22 |
Experimental animals
All experiments with animals were conducted
according to the guidelines of our Institutional Animal Ethics
Committee. Thirty-six 3-week old female nude mice were purchased
from the Institute of Laboratory Animal Sciences, Chinese Academy
of Medical Sciences (CAMS) and maintained in the Animal Center, the
Fourth Hospital of Hebei Medical University, China. The mice were
randomly divided into 6 groups (n=6 per group). MDA-MB-231 cells
were transfected with the indicated combination of different
expression plasmids. The stable clones were subsequently
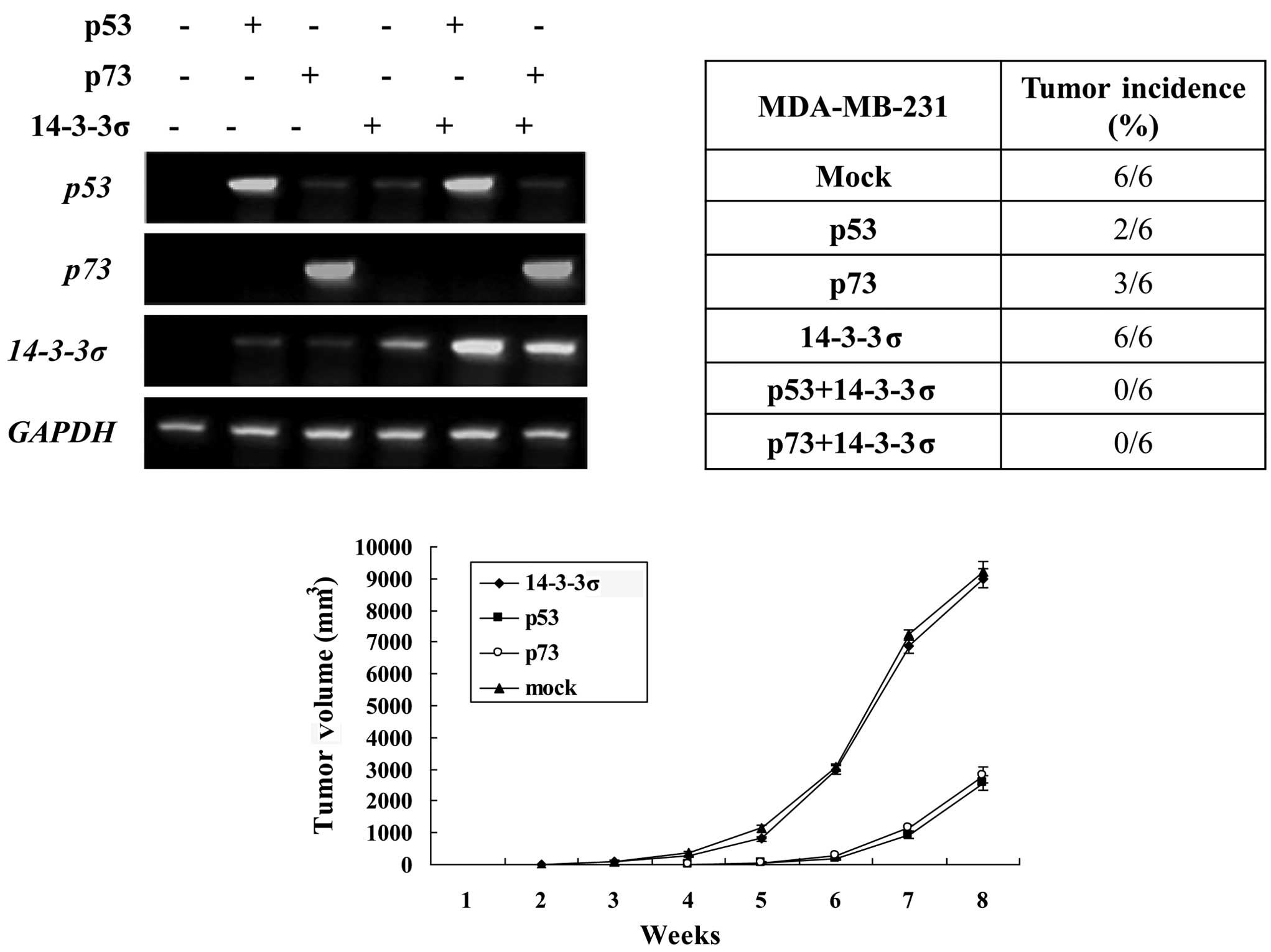
established, and the effects were examined with RT-PCR (Fig. 2A). MDA-MB-231 cells
(5×106) in 0.2 ml of PBS were harvested and injected
subcutaneously into the flank region of the female nude mice. Mice
were observed daily and palpated for tumor formation per one week.
When the tumor masses reached 50–80 mm3, the mice were
administered intraperitoneal injections of adriamycin (ADR) (15
mg/kg per day). Two-dimensional tumor measurements were made daily
after tumor formation. The tumor volume was calculated according to
the formula: volume = π (short diameter2) × (long
diameter)/6. After 8 weeks, the mice were sacrificed by cervical
dislocation. Tumors were excised and used for IHC as described. The
growth curve of the tumors was drawn.
Statistical analysis
Data were analyzed with SPSS 13.0 software.
Pearson’s χ2 test or continuity corrected Pearson’s
χ2 test was applied for the data analysis, expressed as
qualitative values. The κ consistency test was used to analyze the
correlation between the expression of p73 and 14-3-3σ in tumor-free
breast and breast cancer specimens. One way analysis of variance
(ANOVA) was adopted for determine the data of tumor growth.
P<0.05 was considered to be statistically significant.
Results
General
Our results, explained in detail below, showed that
the expression of both p73 and 14-3-3σ was significantly lower than
that in tumor-free breast specimens. Moreover, 14-3-3σ expression
was positively correlated with the expression of p73. Furthermore,
overexpression of 14-3-3σ counteracts tumorigenicity by positively
regulating p73 in p53-mutated or -deficient cancers in vivo.
These results may lead to the use of 14-3-3σ in the therapeutic
application for the p53-mutated and p73-expressed breast cancer
patients.
Clinical data
A total of 132 samples were obtained as paired
samples of tumor and tumor-free breast tissues from 66 individuals.
The patients were females, aged between 30 and 76 years. The
following clinical data were collected retrospectively from patient
files and pathology reports: clinical stages, tumor size,
pathological types, histological grading, status of axillary nodes,
ER status and c-erbB2 status. The tumor samples included 47
infiltrating ductal carcinoma, 17 infiltrating lobular carcinoma, 1
medullary carcinoma and 1 mucinous adenocarcinoma. There were 16
cases in Stage I, 27 cases in Stage II and 23 cases in Stage III.
Patient characteristics are shown in Table III.
| Table IIIRelationship between p73 or 14-3-3σ
expression and clinical parameters of breast cancer patients. |
Table III
Relationship between p73 or 14-3-3σ
expression and clinical parameters of breast cancer patients.
| p73/IHC | | 14-3-3σ/IHC | |
|---|
|
| |
| |
|---|
| Clinical
parameters | − | +, ++ | P-valuea | −, + | ++, +++ | P-valuea |
|---|
| Tumor size (cm) | | | 0.848 | | | 0.936 |
| ≤2 | 5 | 3 | | 6 | 2 | |
| >2 and ≤5 | 26 | 10 | | 25 | 11 | |
| >5 | 15 | 7 | | 15 | 7 | |
| Clinical stage | | | 0.994 | | | 0.744 |
| I | 11 | 5 | | 10 | 6 | |
| II | 19 | 8 | | 19 | 8 | |
| III | 16 | 7 | | 17 | 6 | |
| Pathological type of
carcinoma | | | 0.671 | | | 0.817 |
| Infitrating ductal
carcinoma | 31 | 16 | | 32 | 15 | |
| Infiltrating
lobular carcinoma | 13 | 4 | | 12 | 5 | |
| Medullary
carcinoma | 1 | 0 | | 1 | 0 | |
| Mucinous
adenocarcinoma | 1 | 0 | | 1 | 0 | |
| Histological
grading | | | 0.392 | | | 0.151 |
| I | 5 | 3 | | 5 | 3 | |
| II | 29 | 9 | | 30 | 8 | |
| III | 12 | 8 | | 11 | 9 | |
| Axillary lymph
nodes | | | 0.671 | | | 0.929 |
| − | 25 | 12 | | 26 | 11 | |
| + | 21 | 8 | | 20 | 9 | |
| ER | | | 0.793 | | | 0.348 |
| − | 26 | 12 | | 25 | 13 | |
| + | 20 | 8 | | 21 | 17 | |
| HER-2 | | | 0.485 | | | 0.04 |
| −, ++ | 21 | 11 | | 17 | 15 | |
| +++ | 25 | 9 | | 29 | 5 | |
Expression of p73 and 14-3-3σ in
tumor-free breast and breast cancer specimens
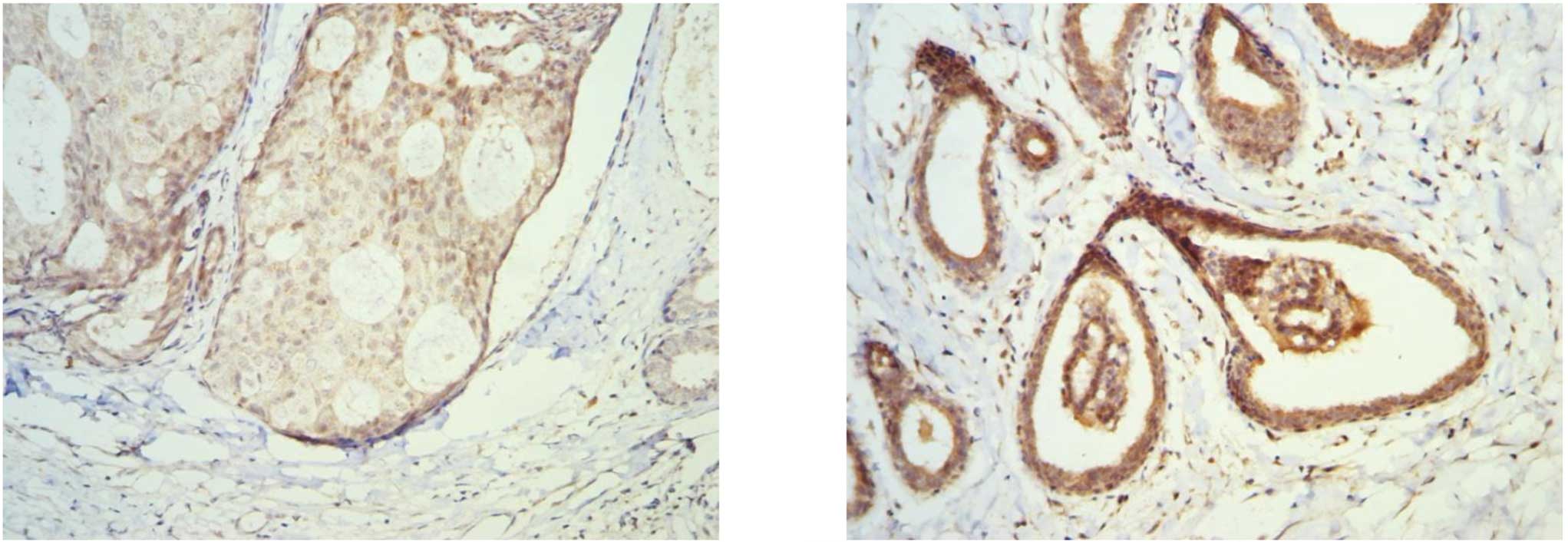
To examine the expression patterns of p73 and
14-3-3σ in breast tissues, the immunohistochemical staining with
p73 and 14-3-3σ antibodies was carried out in 66 paired tumor-free
breast and primary breast cancer specimens. As shown in Fig. 1, p73 and 14-3-3σ were always
positively detected in the cytoplasm and the nucleus. Overall, 50
out of 66 tumor-free breast specimens (75.8%) were found to be
positive (+, ++) with p73 antibody, whereas only 20 out of 66
breast cancer specimens (30.3%) were found to be positive (+, ++)
with p73 antibody (Table II). The
expression of p73 in tumor-free breast specimens was significantly
higher than that in breast cancer specimens (P=0.000). Forty-two
out of 66 tumor-free breast specimens (66.7%) were found to be
positive (++, +++) with the 14-3-3σ antibody, whereas only 20 out
of 66 breast cancer specimens (30.3%) were found to be positive
(++, +++) with the 14-3-3σ antibody (Table II). The expression of 14-3-3σ in
tumor-free breast specimens was significantly higher than that in
breast cancer specimens (P=0.000).
| Table IIExpression of p73 and 14-3-3σ in 66
breast cancer specimens and tumor-free breast specimens detected by
immuno-histochemistry. |
Table II
Expression of p73 and 14-3-3σ in 66
breast cancer specimens and tumor-free breast specimens detected by
immuno-histochemistry.
| p73/IHC | | 14-3-3σ/IHC | |
|---|
|
| |
| |
|---|
| Tissues | − | +, ++ | P-valuea | −, + | ++, +++ | P-valuea |
|---|
| Tumor-free
breast | 16 | 50 | | 24 | 42 | |
| Breast cancer | 46 | 20 | 0.000 | 46 | 20 | 0.000 |
Relationship between p73 or 14-3-3σ
expression and clinical parameters of breast cancer patients
The relationship between p73 or 14-3-3σ expression
with clinical parameters of breast cancer patients was
statistically evaluated with Pearson’s χ2 test or the
continuity corrected Pearson χ2 test. As shown in
Table III, no correlation was
observed between p73 expression and clinical parameters of breast
cancer patients such as tumor size, clinical stage, pathological
types, histological grading, status of axillary lymph nodes, ER
status and c-erbB2 status (P>0.05). 14-3-3σ expression was not
associated with tumor size, clinical stage, pathological types,
histological grading, status of axillary lymph nodes and ER status
(P>0.05), but was negatively associated with the c-erbB2 status
of breast cancer patients (P=0.04).
Correlations between the expression of
p73 and 14-3-3σ in tumor-free breast and breast cancer
specimens
In the present study, we aimed to investigate the
link between p73 and 14-3-3σ in breast tissues. As shown in
Table IV, 37 out of 50 (74%)
p73-positive (+, ++) tumor-free breast specimens showed a positive
14-3-3σ expression (++, +++). A total of 11 out of 16 (68.8%)
p73-negative (−) tumor-free breast specimens showed a negative
14-3-3σ expression (−, +). As shown in Table V, 14 out of 20 (70%) p73-positive
(+, ++) breast cancer specimens showed a positive 14-3-3σ
expression (++, +++). A total of 40 out of 46 (87.0%) p73-negative
(−) breast cancer specimens showed a negative 14-3-3σ expression
(−, +). These results suggest that the 14-3-3σ expression was
positively correlated with the expression of p73 in both tumor-free
breast and breast cancer specimens.
| Table IVCorrelation between the expression of
p73 and 14-3-3σ in tumor-free breast specimens detected by
immunohistochemistry. |
Table IV
Correlation between the expression of
p73 and 14-3-3σ in tumor-free breast specimens detected by
immunohistochemistry.
| 14-3-3σ/IHC | |
|---|
|
| |
|---|
| p73 | −, + | ++, +++ | P-valuea |
|---|
| − | 11 | 5 | |
| +, ++ | 13 | 37 | 0.000 |
| Table VCorrelation between the expression of
p73 and 14-3-3σ in breast cancer specimens detected by
immunohistochemistry. |
Table V
Correlation between the expression of
p73 and 14-3-3σ in breast cancer specimens detected by
immunohistochemistry.
| 14-3-3σ/IHC | |
|---|
|
| |
|---|
| p73 | −, + | ++, +++ | P-valuea |
|---|
| − | 40 | 6 | |
| +, ++ | 6 | 14 | 0.000 |
Overexpression of 14-3-3σ counteracts
tumorigenicity by positively regulating p73 and p53 in vivo
The most important tumor suppressor p53 has been
successfully employed as a molecular target for cancer gene
therapy. Yang et al demonstrated that an enforced expression
of 14-3-3σ suppressed breast tumor growth by positively regulating
p53 (5). However, p53 is highly
mutated in over 50% of human cancers. The p53 family member p73 has
been found to promote cell cycle arrest and/or apoptosis in
response to DNA damage similar to p53, whereas it is rarely mutated
in human cancers. Therefore, p73 may be an effective molecular
target for cancer gene therapy in p53-mutated or -deficient
cancers. Findings of our previous study indicated that 14-3-3σ
increased the stability and activity of p73. Therefore, we
hypothesized that the overexpression of 14-3-3σ counteracted
tumorigenicity by positively regulating p73 in p53-mutated or
-deficient cancers. To explore the tumor-suppressive effect of
14-3-3σ, we established a breast cancer xenograft nude mouse model
with an inducible expression of 14-3-3σ or with an inducible
expression of p53/p73 plus 14-3-3σ with ADR treatment. Tumor
formation was then assayed (Fig. 2B and
C). Tumorigenicity was found in all six mice of the mock
control group and 14-3-3σ expressing group in the 2nd week.
Markedly less tumorigenicity was found in the p53- (2/6) and p73-
(3/6) expressing group than in the control group, and
tumorigenicity was started from the 4th week, suggesting that an
enforced expression of p53 or p73 was capable of eradicating
tumorigenicity in breast cancer cells. However, tumorigenicity was
observed not altered in the 14-3-3σ-expressing group (6/6) as
compared with the control group, suggesting that an enforced
expression of 14-3-3σ alone was not sufficient to eradicate
tumorigenicity in breast cancer cells. No tumorigenicity was
observed in the cells with an expression of 14-3-3σ plus expression
of p53/p73, suggesting that the overexpression of 14-3-3σ
counteracts tumorigenicity by positively regulating p73 and p53
in vivo. Taken together, our present results strongly
suggested that the p73/14-3-3σ pathway plays an important role in
p53-mutated or deficient breast cancer tumorigenicity.
Discussion
14-3-3σ, which belongs to the 14-3-3 family, was
initially identified as a human mammary epithelium-specific marker
1 (24). By using a proteomic
approach, Vercoutter-Edouart et al found that 14-3-3σ is
strongly down-regulated in breast cancer in comparison to normal
breast tissue (25). Mounting
evidence has shown that the down-regulation of 14-3-3σ in breast
cancer is due to the hypermethylation of CpG islands in the
14-3-3σ gene (7–13).
14-3-3σ has been found to be strongly induced in
response to DNA damage, and its expression is directly regulated by
p53. Furthermore, 14-3-3σ stabilizes p53 and directly increases its
transcriptional activity through the interaction with p53,
suggesting a positive feedback loop between 14-3-3σ and p53
(18). In our previous study,
14-3-3σ was found to be a direct transcriptional target of p73 that
enhances the p73-mediated transcriptional as well as pro-apoptotic
activity (22). The purpose of the
present study was to find a possible link between p73 and 14-3-3σ
expression in clinical breast specimens and to characterize the
regulating mechanism of p73/14-3-3σ in vivo. We performed
the immunohistochemical staining with p73 and 14-3-3σ antibodies in
66 paired tumor-free breast and primary breast cancer specimens, to
analyze the expression patterns of p73 and 14-3-3σ and their
correlation in breast cancer. The results show that the expression
of p73 and 14-3-3σ in breast cancer specimens was significantly
lower than the expression in tumor-free breast specimens. Moreover,
14-3-3σ expression is positively correlated with the expression of
p73 in tumor-free breast and breast cancer specimens. Furthermore,
the increased 14-3-3σ expression is associated with a low
expression of c-erbB2, suggesting that a high expression of 14-3-3σ
plays a protective role in the development of breast cancer.
Results of our previous study showed a positive
feedback regulation between p73 and 14-3-3σ in vitro.
Cosequently, we hypothesized that the overexpression of 14-3-3σ may
counteract tumorigenicity by positively regulating p73 in
p53-mutated or -deficient cancers. To prove our hypothesis, the
breast cancer xenograft nude mouse model was established with an
inducible expression of 14-3-3σ or an inducible expression of
p53/p73 plus 14-3-3σ with ADR. The tumor formation assay shows that
overexpression of 14-3-3σ alone is not sufficient to eradicate
tumorigenicity in breast cancer cells. However, a tumor-suppressive
effect was evident only in the presence of p53 or p73. Therefore,
our study demonstrates that an enforced expression of 14-3-3σ may
suppress breast tumor growth by positively regulating p73 in
p53-mutated breast cancers.
Acknowledgements
This study was supported by the National Nature
Science Foundation of China (No. 81001178). We thank Dr Nakagawara
of the Chiba Cancer Center for the p53, p73 and 14-3-3σ expression
plasmids. We would also like to extend our thanks to Dr Qianglin
Duan, a skilled English proofreader from Tongji University for
revision of this study.
References
|
1
|
Mhawech P: 14-3-3 proteins - an update.
Cell Res. 15:228–236. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rosenquist M: 14-3-3 proteins in
apoptosis. Braz J Med Biol Res. 36:403–408. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fu H, Subramanian RR and Masters SC:
14-3-3 proteins: structure, function, and regulation. Annu Rev
Pharmacol Toxicol. 40:617–647. 2000. View Article : Google Scholar
|
|
4
|
Muslin AJ and Xing H: 14-3-3 proteins:
regulation of subcellular localization by molecular interference.
Cell Signal. 12:703–709. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Van Hemert MJ, Steensma HY and van Heusden
GP: 14-3-3 proteins: key regulators of cell division, signalling
and apoptosis. Bioessays. 23:936–946. 2001.PubMed/NCBI
|
|
6
|
Hermeking H: The 14-3-3 cancer connection.
Nature Rev. 3:931–943. 2003.PubMed/NCBI
|
|
7
|
Ferguson AT, Evron E, Umbricht CB, et al:
High frequency of hypermethylation at the 14-3-3σ locus leads to
gene silencing in breast cancer. Proc Natl Acad Sci USA.
97:6049–6054. 2000.
|
|
8
|
Lodygin D and Hermeking H: The role of
epigenetic inactivation of 14-3-3sigma in human cancer. Cell Res.
15:237–246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Iwata N, Yamamoto H, Sasaki S, et al:
Frequent hypermethylation of CpG islands and loss of expression of
the 14-3-3 sigma gene in human hepatocellular carcinoma. Oncogene.
19:5298–5302. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gasco M, Bell AK, Heath V, et al:
Epigenetic inactivation of 14-3-3 sigma in oral carcinoma:
association with p16 (INK4a) silencing and human papillomavirus
negativity. Cancer Res. 62:2072–2076. 2002.PubMed/NCBI
|
|
11
|
Gasco M, Sullivan A, Repellin C, et al:
Coincident inactivation of 14-3-3sigma and p16INK4a is an early
event in vulval squamous neoplasia. Oncogene. 21:1876–1881. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Suzuki H, Itoh F, Toyota M, Kikuchi T,
Kakiuchi H and Imai K: Inactivation of the 14-3-3 sigma gene is
associated with 5V CpG island hypermethylation in human cancers.
Cancer Res. 60:4353–4357. 2000.PubMed/NCBI
|
|
13
|
Ikeda K and Inoue S: Estrogen receptors
and their downstream targets in cancer. Arch Histol Cytol.
67:435–442. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang H, Zhao R and Lee MH: 14-3-3sigma, a
p53 regulator, suppresses tumor growth of nasopharyngeal carcinoma.
Mol Cancer Ther. 5:253–260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hermeking H, Lengauer C, Polyak K, et al:
14-3-3 σ is a p53-regulated inhibitor of G2/M progression. Mol
Cell. 1:3–11. 1997.
|
|
16
|
Taylor WR and Stark GR: Regulation of the
G2/M transition by p53. Oncogene. 20:1803–1815. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Laronga C, Yang HY, Neel C and Lee MH:
Association of the cyclin-dependent kinases and 14-3-3sigma
negatively regulates cell cycle progression. J Biol Chem.
275:23106–23112. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang HY, Wen YY, Chen CH, Lozano G and Lee
MH: 14-3-3 σ positively regulates p53 and suppresses tumor growth.
Mol Cell Biol. 23:7096–7107. 2003.
|
|
19
|
Ikawa Z, Nakagawara A and Ikawa Y: p53
family genes: structural comparison, expression and mutation. Cell
Death Differ. 6:1154–1161. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cattoretti G, Rilke F, Andreola S, D’
Amato L and Domenico D: p53 in breast cancer. Int J Cancer.
41:178–183. 1988. View Article : Google Scholar
|
|
21
|
Moll UM, Riou G and Levine AJ: Two
distinct mechanisms alter p53 in breast cancer: mutation and
nuclear exclusion. Proc Natl Acad Sci USA. 89:7262–7266. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sang M, Li Y, Ozaki T, et al:
p73-dependent induction of 14-3-3 σ increases the chemo-sensitivity
of drug-resistant human breast cancers. Biochem Biophys Res Commun.
347:327–333. 2006.PubMed/NCBI
|
|
23
|
Vayssade M, Haddada H, Faridoni-Laurens L,
Tourpin S, Valent A, Bénard J and Ahomadegbe JC: p73 functionally
replaces p53 in Adriamycin-treated, p53-deficient breast cancer
cells. Int J Cancer. 116:860–869. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Prasad GL, Valverius EM, McDuffie E and
Cooper HL: Complementary DNA cloning of a novel epithelial cell
marker protein, Hmel, that may be down-regulated in neoplastic
mammary cells. Cell Growth Differ. 3:507–513. 1992.PubMed/NCBI
|
|
25
|
Vercoutter-Edouart AS, Lemoine J, Le
Bourhis X, et al: Proteomic analysis reveals that 14-3-3σ is
down-regulated in human breast cancer cells. Cancer Res. 61:76–80.
2001.
|