Introduction
There are three types of interferons (IFNs): type I,
II and III (1,2). The signaling pathway induced by all
IFNs is the JAK/STAT pathway, in which different STAT proteins play
critical roles (3). IFNγ is the
only type II IFN and is involved in a broad spectrum of immune
regulations, including antiviral and antitumor activities. One of
the dominant mechanisms of these activities is facilitating the
induction of apoptosis in the affected cells (4).
Apoptosis, also known as programmed cell death, is
one of the most important mechanisms for antiviral and antitumor
activities, and is induced by a number of cytokines (5). It is primarily executed by caspases,
the cysteine aspartate-specific proteases (6,7). There
are two major pathways, one mediated by mitochondria (intrinsic
pathway) and another mediated by death receptors (extrinsic
pathway), both of which lead to activation of caspases (5,8,9).
Activated caspases cleave different cellular proteins causing
genomic DNA fragmentation, cell morphology changes and, eventually,
cell death. Other proteins, including p53 and Bcl-XL, have also
been reported to be critical in the apoptosis signaling pathway
(10). Regulations between these
proteins and the β-catenin pathway have been reported in several
cancer cell types, but not in hepatocellular carcinoma (HCC) cells
(11–14).
β-catenin is a key component of the Wnt/β-catenin
signaling pathway, and a mediator for the Ras/phosphatidylinositol
3-kinase (PI3K) pathways (15,16).
Active β-catenin interacts with transcription factors such as T
cell factor/lymphoid enhancer (TCF/LEF), CBP and p300, leading to
target gene transcription. The downstream biological activities
mediated by active β-catenin include differentiation, survival and
proliferation. In addition, active β-catenin also binds to
cadherins in the cell membrane to provide structural support for
adhesion (17,18). The Wnt signaling pathway is also
involved in the carcinogenesis of a number of types of cancer and
is commonly believed to be a survival pathway. There are only a few
reports of its contribution to apoptosis induction (12–14,19,20).
Nothing has been reported concerning the effects of high levels of
β-catenin on IFNγ signaling in HCC cells.
Previously, we studied the regulation of IFNγ and
the β-catenin/Wnt signaling pathway in human astrocytes (21). In the present study, we intended to
investigate the effect of upregulated β-catenin on IFNγ-induced
apoptosis in human liver carcinoma cells, the molecular mechanisms
by which this occurs.
Materials and methods
Reagents and antibodies
FITC-conjugated mouse anti-human caspase 3, caspase
8 and p53 antibodies, APC-conjugated mouse anti-human caspase 9
antibody and mouse anti-human β-catenin antibody were purchased
from BD Biosciences (San Jose, CA, USA). FITC-conjugated goat
anti-mouse antibody was purchased from Jackson ImmunoResearch
Laboratories, Inc. (West Grove, PA, USA). Mouse anti-human Bcl-XL
antibody was purchased from MBL International Corporation (Woburn,
MA, USA). The STAT1 inhibitor fludarabine (FLUD) was purchased from
Sigma-Aldrich (St. Louis, MO, USA. The STAT3 inhibitor S3I was
purchased from Calbiochem/EMD Biosciences, Inc. (Gibbstown, NJ,
USA). The pancaspase inhibitor Z-VAD-FMK was purchased from
Calbiochem/EMD Biosciences.
Cell lines, DNA constructs and
transfection
HepG2 cells, a human HCC cell line (PriCell Research
Institute, Wuhan, China) were maintained in DMEM (Sigma-Aldrich)
with 10% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich).
HepG2 cells were transfected with a constitutively active β-catenin
construct or its cognate vector using TransIT transfection kit
(Mirus Bio LLC, Madison, WI, USA) following manufacturer’s
instruction. The constitutively active β-catenin plasmid contains a
serine-to-tyrosine mutation at position 33 that protects the
protein from proteosomal degradation.
Immunofluorescence staining and flow
cytometry analysis
Flow cytometry was performed as described previously
(21). To detach HepG2 cells
without cleaving surface proteins, they were incubated with 1 mM
EDTA for 5 min and then washed and suspended in 1X PBS. Cells were
stained with appropriate target antibodies and isotype antibodies
using conventional surface- and/or intracellular-staining methods.
When both surface and intracellular staining was desired, cells
were first fixed and made permeable using BD Cytofix/Cytoperm
Fixation and Permeating Solution (BD Pharmingen; San Diego, CA,
USA), followed by staining for intracellular proteins. Cells were
then washed extensively with 1X PBS to remove excess antibodies,
stained for extracellular targets, and fixed with 2% formaldehyde.
Fluorescence was evaluated with a FACS Caliber flow cytometer, and
data analyzed using FlowJo software (Tree Star, Inc., Ashland, OR,
USA).
Proliferation and cell viability
assays
Cell viability assays were performed as previously
described (22,23). Briefly, to determine cell viability,
equal amount of cells (105 cells/well) were plated in
6-well plates and transfected and/or treated, as indicated in the
text. Dead cells lost their attachment and were washed away by 1X
PBS. Viable (adherent) cells were released from the wells by
trypsinization prior to cell counting.
TUNEL assay
TUNEL assay to determine DNA fragmentation in
apoptotic cells was performed according to the manufacturer’s
instructions (Promega Corporation, Madison, WI, USA). Briefly,
3–5x106 cells were trypsinized, washed twice with cold
PBS, fixed in 4% paraformaldehyde at 4˚C for 20 min, washed again
with PBS and made permeable with 0.5 ml 0.5% saponin at 22˚C for 5
min. The cells were washed with PBS, incubated with 80 μl
equilibration buffer at 22˚C for 5 min, washed with PBS,
re-suspended in 50 μl Nucleotide Mix and incubated in the dark at
37˚C for 1 h. Cells were washed again with PBS then analyzed by
fluorescence microscopy.
Statistical analysis
Statistical analyses were performed using Prism
software (GraphPad Prism). Untreated and treated groups were
compared using the Student’s t-test when the data were normally
distributed. When the data showed abnormal distribution, the two
groups were compared using the nonparametric Mann-Whitney U test.
All tests were two-tailed. P<0.05 was considered to indicate a
statistically significant difference.
Results
Excess β-catenin promotes IFNγ-induced
apoptosis in HepG2 cells
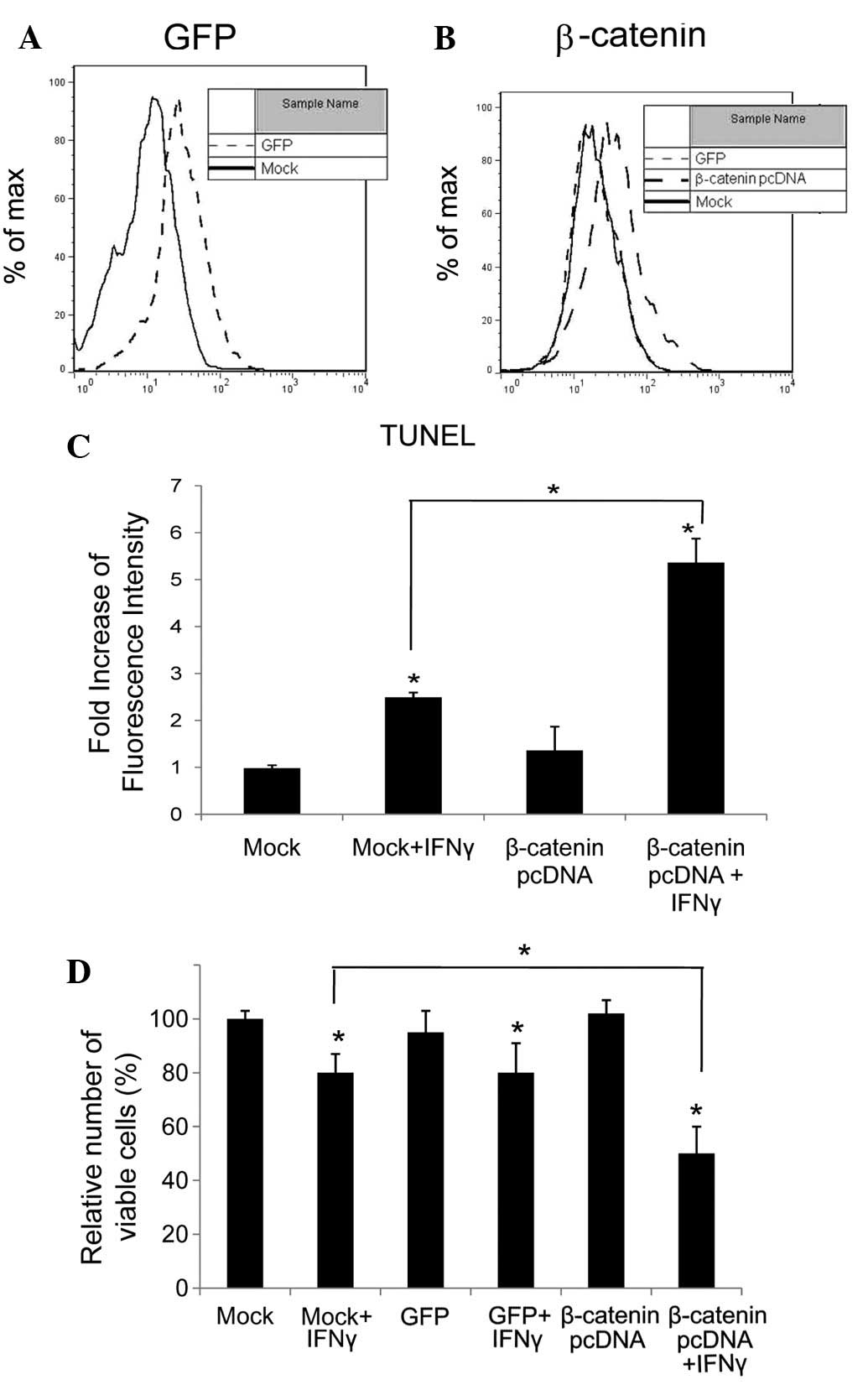
To upregulate β-catenin, we transfected HepG2 HCC
cells with a constitutively active construct of β-catenin
(β-catenin pcDNA). Being controls, equal amount of HepG2 cells were
transfected with cognate vector (Mock) and GFP construct (GFP)
respectively. To test the efficiency of the transfection, the level
of GFP and active β-catenin was determined by flow cytometry, and
the results are shown in Fig. 1A and
B, respectively. Compared with Mock- and GFP-transfected cells,
the levels of β-catenin in β-catenin pcDNA-transfected cells were
significantly elevated (P<0.05).
HepG2 cells transfected with β-catenin pcDNA,
cognate vector (Mock) or GFP construct (GFP) were left untreated
and treated with IFNγ (100 ng/ml) for 72 h, and viable cells were
counted under a microscope. The results demonstrated that, compared
with untreated controls, IFNγ reduced viable cell counts in all
three groups of transfected cells, but most significantly in cells
which expressed excess β-catenin (P<0.05). Upregulated β-catenin
alone in HepG2 cells did not affect cell proliferation (P<0.05;
Fig. 1C).
To determine whether apoptosis was induced by IFNγ
in HepG2 cells, TUNEL assay was used to detect DNA fragmentation in
apoptosis, and was performed on β-catenin pcDNA- and cognate vector
(Mock)-transfected HepG2 cells, treated with or without IFNγ for 72
h. The fluorescence was elevated by 2.5-fold in the IFNγ-treated
Mock-transfected cells and 5.5-fold in cells expressing excess
β-catenin (Fig. 1D). IFNγ-induced
apoptosis was promoted in HepG2 cells with high levels of
β-catenin.
β-catenin upregulation leads to changes
in signaling components in apoptosis pathway
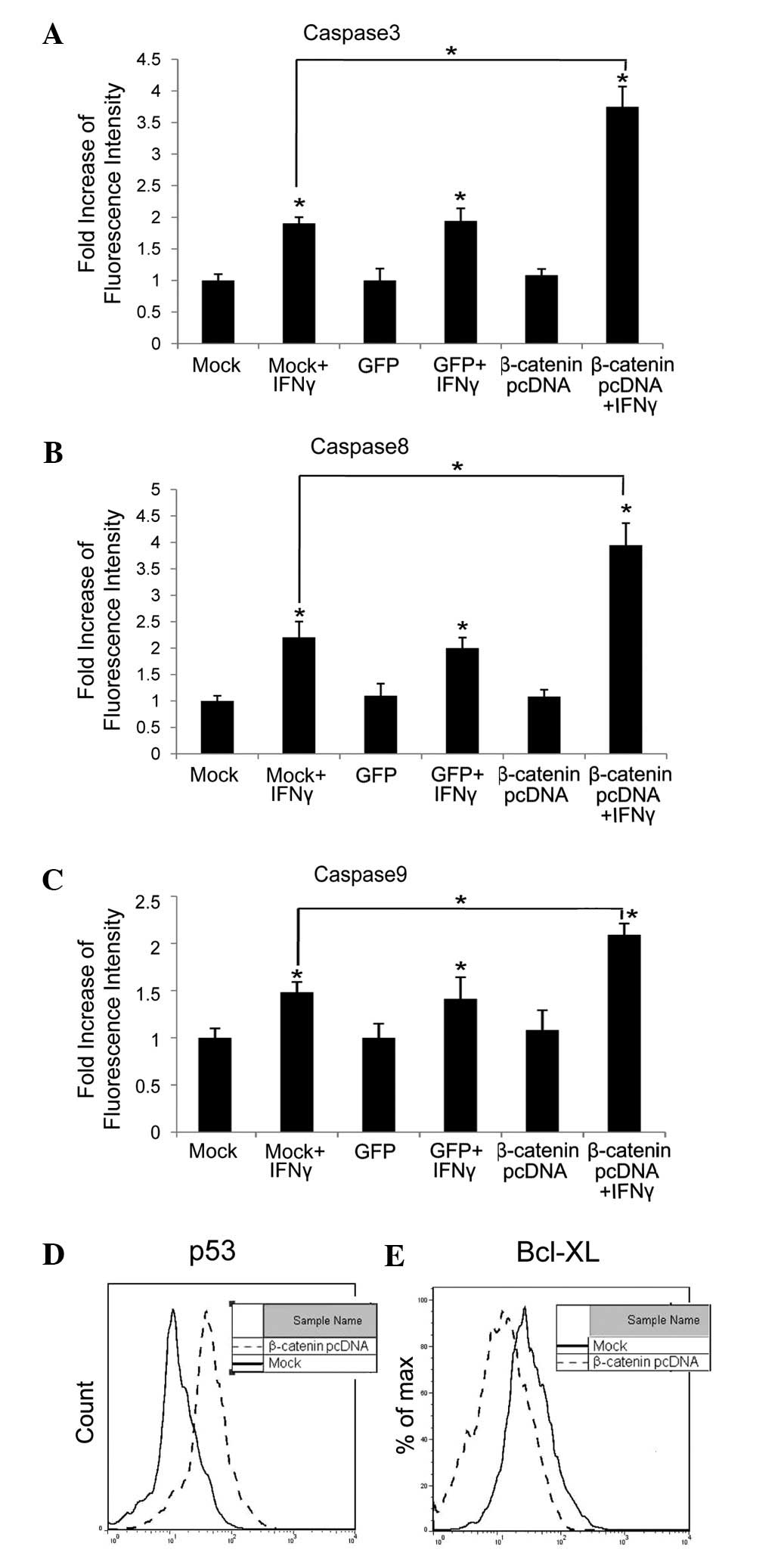
We next investigated whether levels of signaling
components in the apoptosis pathway were changed due to β-catenin
upregulation. Levels of activated caspase 3 (Fig. 2A), 8 (Fig. 2B) and 9 (Fig. 2C) were tested, and found to be
increased 2-, 2- and 1.5-fold in HepG2 cells transfected with
cognate vector (Mock) or GFP and 4-, 4- and 2-fold in HepG2 cells
with excess β-catenin, when treated with IFNγ (100 ng/ml) for 72 h,
compared with untreated cells (P<0.05), respectively.
Previous studies have reported that, due to high
levels of β-catenin, p53 was accumulated and the Bcl-XL level was
decreased (11–14). p53 is a proapoptotic cellular
protein, while Bcl-XL is antiapoptotic. Both are vital components
in carcinogenesis (10).
In the present study we tested p53 and Bcl-XL levels
in HepG2 cells transfected with β-catenin pcDNA or its cognate
vector (Mock). The results indicated that, when β-catenin was
upregulated, the p53 level was elevated and the Bcl-XL level was
reduced (Fig. 2D and E).
Roles of STATs and caspases in
IFNγ-induced apoptosis in HepG2 cells with excess β-catenin
To further identify the key signaling components for
IFNγ to induce apoptosis in HepG2 cells that express excess
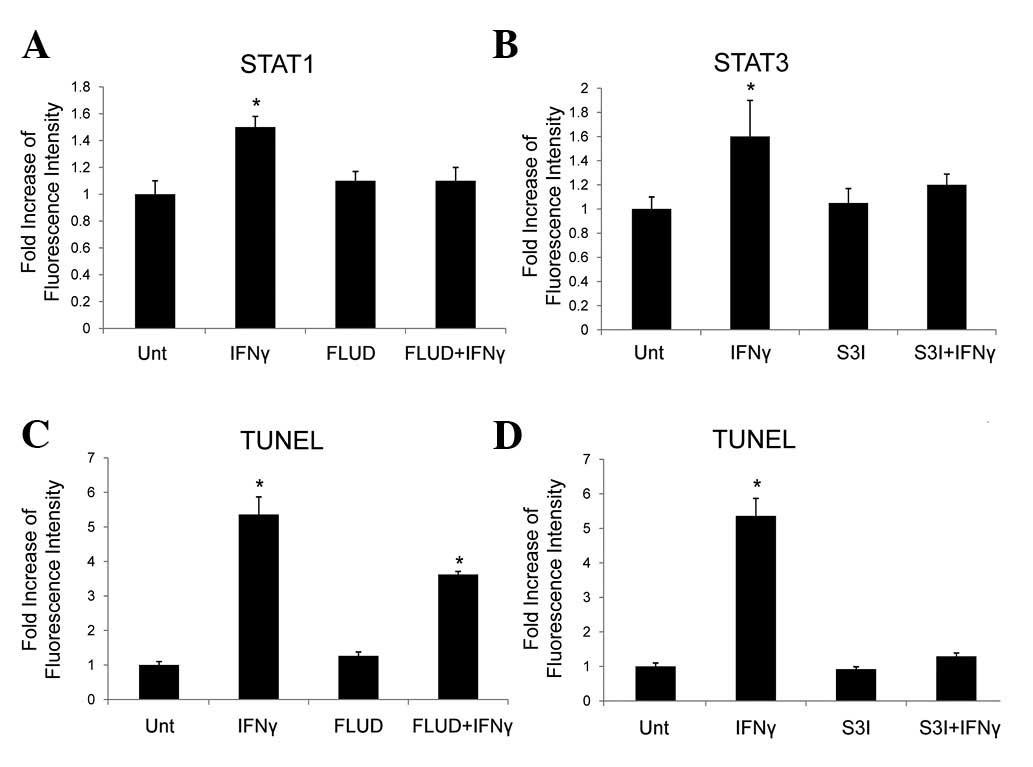
β-catenin, we used inhibitors for STAT1, STAT3 and caspases. In a
previous study, we demonstrated that STAT1 and STAT3 were induced
by IFNγ in human astroglioma cells, in which STAT3 played a key
role in the regulation of the β-catenin pathway (21). In the present study, we confirmed
the STAT1 and STAT3 activation induced by IFNγ, and the effects of
STAT1 inhibitor, FLUD, and STAT3 inhibitor, S3I, in HepG2 cells
with high levels of β-catenin by flow cytometry (Fig. 3A and B). Apoptosis induction in the
presence or absence of FLUD or S3I was then tested by TUNEL assay,
and results showed that FLUD partially blocked IFNγ-induced
apoptosis, while S3I completely suppressed it (Fig. 3C and D). These results demonstrated
that STAT3 is critical for IFNγ-induced apoptosis in HepG2 cells
with high levels of β-catenin.
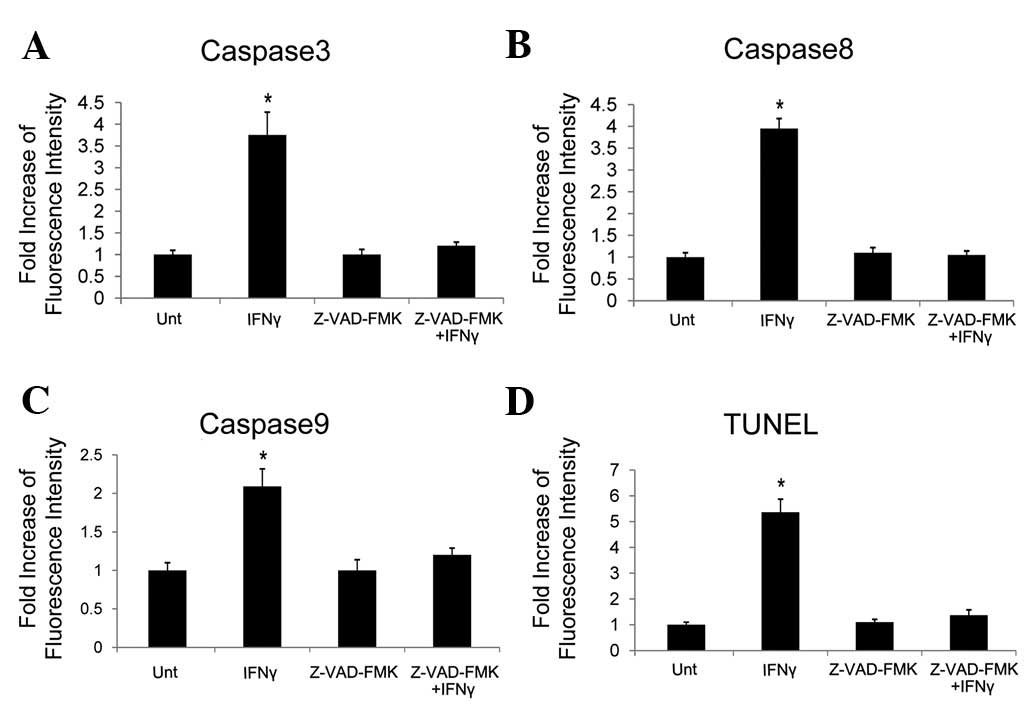
We have demonstrated that caspases were induced by
IFNγ in HepG2 cells with upregulated β-catenin. We then tested
their necessity. The pancaspase inhibitor Z-VAD-FMK was used.
Caspases 3, 8 and 9 were blocked by Z-VAD-FMK (Fig. 4A–C) and IFNγ-induced apoptosis was
also inhibited, as demonstrated by the results of the TUNEL assay
(Fig. 4D). These data illustrate
that the apoptosis induced by IFNγ in HepG2 cells with high levels
of β-catenin is caspase-dependent.
Discussion
Although β-catenin pathway is recognized as a
well-known enhancer of proliferation and survival in tumor cells,
its over-expression or accumulation has also been reported to
induce apoptosis in fibroblasts and multiple myeloma cells, as well
as several other tumor cell lines (12–14,19,20,24).
Raab et al demonstrated that inhibition of PKC led to
accumulation of active β-catenin, which contributes to
enzastaurin-induced cell death in multiple myeloma cells (20). Other studies have shown that
constitutively active β-catenin triggered p53-dependent growth
arrest in fibroblasts and endometrial carcinoma cells (12,13).
However, Kim et al reported the induction of apoptosis
independent of p53 status and LEF-1 activation by β-catenin, when
it was overexpressed in colon cancer or HeLa cells (14). Overexpression of a stable form of
β-catenin or inhibited endogenous β-catenin degradation has been
reported to lead to G2 cell cycle arrest and apoptosis in epidermal
keratinocytes (24). Nevertheless,
the ability of β-catenin to induce apoptosis has been discovered
but not well characterized. In the present study, we found that
overexpression of β-catenin alone did not promote apoptosis in
liver carcinoma cells. However, when combined with IFNγ
stimulation, apoptosis was markedly induced compared with
Mock-transfected liver carcinoma cells. In addition, we aimed to
identify key modulators in this regulation. Studies concerning the
regulation of the β-catenin pathway by IFNγ have been published
(21,25). We have shown that the β-catenin
pathway regulates IFNγ signaling.
In this study, we showed the proapoptotic effect of
accumulated β-catenin in IFNγ-treated liver carcinoma cells. The
active β-catenin was upregulated by transfection of a plasmid
containing sequence of a constitutively active β-catenin (β-catenin
pcDNA). High levels of β-catenin alone did not affect the
proliferation of transfected HepG2 cells, but promoted IFNγ-induced
apoptosis compared with data of Mock-transfected cells, confirmed
by TUNEL assay (Fig. 1). In other
studies, upregulated β-catenin alone led to apoptosis in specific
cell lines (11,12,14),
which is different from the results of the present study in HepG2
cells.
We next found that excess β-catenin further promoted
the IFNγ-induced activation of caspases 3, 8 and 9, upregulated the
p53 level and downregulated Bcl-XL, compared with Mock-transfected
cells (Fig. 2). It is known that
IFNγ induces caspases 3, 8 and 9 in certain cell lines, including
glioblastoma and conjunctival epithelial cells (26,27).
In the present study, we demonstrated that IFNγ induced these
caspases in HCC cells, and that their activation was enhanced by
β-catenin overexpression. It has been reported that excess
β-catenin results in p53 accumulation (11–13),
which is consistent with our findings. We also showed that excess
β-catenin down-regulated Bcl-XL in HCC cells, which is in
accordance with the study by Kim et al, where Bcl-XL
inhibited the apoptotic effects of excess β-catenin (14).
We further investigated the importance of several
key signaling components in IFNγ-induced apoptosis. We used STAT1,
STAT3 and caspase inhibitors (FLUD, S3I and Z-VAD-FMK,
respectively) to inhibit specific signaling proteins, and observed
their effects on IFNγ-induced apoptosis in cells expressing stable
β-catenin. STAT1 and STAT3 are induced by all IFNs, including IFNγ,
and are critical signaling components in the JAK/STAT pathway
(3). Z-VAD-FMK has been reported to
be able inhibit most caspases to block IFNγ-induced apoptosis in
HT29 colorectal carcinoma cells (28). We found that STAT3 and caspases, but
not STAT1, were indispensible for apoptosis induction (Figs. 3 and 4). This is consistent with the results of
our previous study in human astroglioma cells, in which IFNγ
regulates the β-catenin pathway in a STAT3-dependent manner, in
which STAT1 it is not necessarily involved (21).
The β-catenin pathway is generally considered a
survival signaling pathway, but the results of the present study,
along with several others, clearly describe its positive roles in
apoptosis induction (11–14,20).
It remains unclear as to which mechanisms it employs to trigger
apoptosis. There may be a molecular ‘detector’ to monitor β-catenin
levels, which may be extremely high in cancer cells. When the level
of β-catenin reaches a certain threshold level, the detector
triggers apoptosis, with or without additional stimulation, for
example, by IFNγ. This hypothesis requires further investigation.
IFNγ is a strong immune modulator, and has a broad effect on the
immune system (4). New findings on
the interaction between the β-catenin and IFNγ pathways may aid the
understanding of the cellular signaling network, the identification
of the potentials of β-catenin and IFNγ signaling and the
development of approaches to manage different types of cancer.
We have identified the potential of the β-catenin
pathway in promoting apoptosis induction. It is possible that the
upregulation of β-catenin in cancer cells may induce apoptosis and
eliminate cancer cells. Further studies are required to test this
hypothesis. There are chemicals, such as DKK1 neutralizing
antibody, that upregulate β-catenin (21) and which may be used to promote
IFNγ-induced apoptosis in liver cancer cells.
In conclusion, we have revealed the regulation of
the IFNγ signaling pathway by the β-catenin pathway in liver cancer
cells. We have shown in this study that the overexpression of
β-catenin in HCC cells promoted IFNγ-induced apoptosis, possibly
via the regulation of p53 and Bcl-XL levels. The apoptosis was
STAT3- and caspase-dependent. These findings extend our knowledge
of the Wnt/β-catenin pathway and its interaction with the IFN
signaling pathway, which may aid the development of new strategies
to manage liver cancer.
Reference
|
1.
|
S ParmarLC PlataniasInterferonsCancer
Treat Res1264568200510.1007/0-387-24361-5_3
|
|
2.
|
C KellyP KlenermanE BarnesInterferon
lambdas: the next cytokine
stormGut6012841293201110.1136/gut.2010.22297621303914
|
|
3.
|
C SchindlerC PlumleeInteferons pen the
JAK-STAT pathwaySemin Cell Dev
Biol19311318200810.1016/j.semcdb.2008.08.01018765289
|
|
4.
|
U BoehmT KlampM GrootJC HowardCellular
responses to interferon-gammaAnnu Rev
Immunol15749795199710.1146/annurev.immunol.15.1.7499143706
|
|
5.
|
MJ ClemensInterferons and apoptosisJ
Interferon Cytokine
Res23277292200310.1089/10799900376662812412859854
|
|
6.
|
NA ThornberryCaspases: key mediators of
apoptosisChem
Biol5R97R103199810.1016/S1074-5521(98)90615-99578633
|
|
7.
|
HR StennickeGS SalvesenCaspase
assaysMethods
Enzymol32291100200010.1016/S0076-6879(00)22010-710914007
|
|
8.
|
DV KalvakolanuThe GRIMs: a new interface
between cell death regulation and interferon/retinoid induced
growth suppressionCytokine Growth Factor
Rev15169194200410.1016/j.cytogfr.2004.01.00215110800
|
|
9.
|
MO HengartnerThe biochemistry of
apoptosisNature407770776200010.1038/3503771011048727
|
|
10.
|
S ElmoreApoptosis: a review of programmed
cell deathToxicol
Pathol35495516200710.1080/0192623070132033717562483
|
|
11.
|
A DamalasA Ben-Ze’evI SimchaExcess
beta-catenin promotes accumulation of transcriptionally active
p53EMBO J1830543063199910.1093/emboj/18.11.305410357817
|
|
12.
|
A DamalasS KahanM ShtutmanA Ben-Ze’evM
OrenDeregulated beta-catenin induces a p53- and ARF-dependent
growth arrest and cooperates with Ras in transformationEMBO
J2049124922200110.1093/emboj/20.17.491211532955
|
|
13.
|
M SaegusaM HashimuraT KuwataM HamanoI
OkayasuBeta-catenin simultaneously induces activation of the
p53-p21WAF1 pathway and overexpression of cyclin D1 during squamous
differentiation of endometrial carcinoma cellsAm J
Pathol16417391749200410.1016/S0002-9440(10)63732-7
|
|
14.
|
K KimKM PangM EvansED HayOverexpression of
beta-catenin induces apoptosis independent of its transactivation
function with LEF-1 or the involvement of major G1 cell cycle
regulatorsMol Biol
Cell1135093523200010.1091/mbc.11.10.350911029052
|
|
15.
|
J EspadaM Pérez-MorenoVM BragaP
Rodriguez-VicianaA CanoH-Ras activation promotes cytoplasmic
accumulation and phosphoinositide 3-OH kinase association of
beta-catenin in epidermal keratinocytesJ Cell
Biol146967980199910.1083/jcb.146.5.96710477752
|
|
16.
|
K WillertS ShibamotoR NusseWnt-induced
dephosphorylation of axin releases beta-catenin from the axin
complexGenes Dev1317681773199910.1101/gad.13.14.176810421629
|
|
17.
|
RT MoonJD BrownM TorresWNTs modulate cell
fate and behavior during vertebrate developmentTrends
Genet13157162199710.1016/S0168-9525(97)01093-79097727
|
|
18.
|
JR MillerRT MoonSignal transduction
through beta-catenin and specification of cell fate during
embryogenesisGenes
Dev1025272539199610.1101/gad.10.20.25278895655
|
|
19.
|
JC GhoshDC AltieriActivation of
p53-dependent apoptosis by acute ablation of glycogen synthase
kinase-3beta in colorectal cancer cellsClin Cancer
Res1145804588200510.1158/1078-0432.CCR-04-262415958644
|
|
20.
|
MS RaabI BreitkreutzG TononTargeting PKC:
a novel role for beta-catenin in ER stress and apoptotic
signalingBlood11315131521200910.1182/blood-2008-05-15704019018094
|
|
21.
|
W LiLJ HendersonEO MajorL
Al-HarthiIFN-gamma mediates enhancement of HIV replication in
astrocytes by inducing an antagonist of the beta-catenin pathway
(DKK1) in a STAT 3-dependent mannerJ
Immunol18667716778201110.4049/jimmunol.110009921562161
|
|
22.
|
W LiA Lewis-AntesJ HuangM BalanSV
KotenkoRegulation of apoptosis by type III interferonsCell
Prolif41960979200810.1111/j.1365-2184.2008.00558.x19040572
|
|
23.
|
W LiX HuangZ LiuY WangH ZhangH TongH WuS
LinType III interferon induces apoptosis in human lung cancer
cellsOncol Rep2811171125201222766785
|
|
24.
|
D OlmedaS CastelS VilaroA CanoBeta-catenin
regulation during the cell cycle: implications in G2/M and
apoptosisMol Biol
Cell1428442860200310.1091/mbc.E03-01-086512857869
|
|
25.
|
P NavaS KochMG LaukoetterInterferon-gamma
regulates intestinal epithelial homeostasis through converging
beta-catenin signaling
pathwaysImmunity32392402201010.1016/j.immuni.2010.03.001
|
|
26.
|
R JanardhananNL BanikSK
RayN-(4-Hydroxyphenyl) retinamide induced differentiation with
repression of telomerase and cell cycle to increase
interferon-gamma sensitivity for apoptosis in human glioblastoma
cellsCancer Lett2612636200810.1016/j.canlet.2007.11.016
|
|
27.
|
X ZhangW ChenCS De PaivaInterferon-gamma
exacerbates dry eye-induced apoptosis in conjunctiva through dual
apoptotic pathwaysInvest Ophthalmol Vis
Sci5262796285201110.1167/iovs.10-708121474767
|
|
28.
|
MC ZhangHP LiuLL DemchikYF ZhaiDJ
YangLIGHT sensitizes IFN-gamma-mediated apoptosis of HT-29 human
carcinoma cells through both death receptor and mitochondria
pathwaysCell Res14117124200410.1038/sj.cr.729021015115612
|