Introduction
Amelogenin Y-linked (AMELY) has been
identified to be one of the significantly highly expressed (fold
change ≥2) genes in human hepatocellular carcinoma (HCC).
Systems-computational analysis was used to elucidate the function
and mechanism of AMELY-activated upstream regulation
networks in HCC compared with low or non-tumor hepatitis/cirrhotic
tissues (HBV or HCV infection) from GEO dataset GSE10140-10141
(1).
The possible correlations among immune response,
protein secretion, Wnt signaling and calcium release into the
cytosol with cancer or tumor formation have been investigated in
several studies. Examples include the tetraspanins and the immune
response against cancer (2);
induction of tumor-specific immune response by gene transfer of
Hsp70 cell-penetrating peptide fusion protein and tumors in mice
(3); in situ immune
responses after neoadjuvant chemotherapy for breast cancer
predicting survival (4); human
tumor cells killed by anthracyclines inducing a tumor-specific
immune response (5); keratin 17
promoting epithelial proliferation and tumor growth by polarizing
the immune response in skin (6);
interleukin-10, but not interleukin-18, possibly being associated
with the immune response against well-differentiated thyroid cancer
(7); CXCR7 protein expression
correlating with elevated mmp-3 secretion in breast cancer cells
(8); heat shock cognate 70 protein
secretion as a new growth arrest signal for cancer cells (9); overexpression of p53 protein and local
hGH, IGF-I, IGFBP-3, IGFBP-2 and PRL secretion by human breast
cancer explants (10); restoration
of full-length APC protein in SW480 colon cancer cells inducing
exosome-mediated secretion of DKK-4 (11); cancer cell secretion of the DAMP
protein HMGB1 supporting progression in malignant mesothelioma
(12); targeting Wnt signaling in
colon cancer stem cells (13);
protein cross-talk in CD133+ colon cancer cells
indicating activation of the Wnt pathway and upregulation of SRp20
that is potentially involved in tumorigenicity (14); miRNA-34 intrinsically linking p53
tumor suppressor and Wnt signaling (15); the Wnt/β-catenin pathway regulating
self-renewal of cancer stem-like cells in human gastric cancer
(16); a serrated colorectal cancer
pathway predominating over the classic Wnt pathway in patients with
hyperplastic polyposis syndrome (17); insights from studies with oral cleft
genes suggesting associations between Wnt pathway genes and risk of
oral cancer (18); and hydrogen
sulfide increasing calcium-activated potassium (BK) channel
activity of rat pituitary tumor cells (19). The function and mechanism of the
high expression of the AMELY-activated upstream regulation
network in HCC is not known and remains to be elucidated.
The aim of this study was to compare the biological
processes and occurrence numbers of gene ontology (GO) in non-tumor
hepatitis/cirrhotic tissues (HBV or HCV infection) with low
expression of AMELY upstream regulation networks and the
corresponding HCC tissue with high expression (fold change ≥2) of
AMELY. AMELY-activated upstream regulation molecular
networks in non-tumor hepatitis/cirrhotic tissues and HCC were
constructed. A further aim was to identify the AMELY
upstream regulation molecular network involved in HCC.
Materials and methods
Materials
Microarrays from of 6,144 genes from 25 non-tumor
hepatitis/cirrhotic tissues and 25 HCC patients were used for
analyzing the possible AMELY-activated upstream regulation networks
of HCC based on GEO dataset GSE10140-10141 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE10140
and http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE10141).
The raw microarray data were pre-processed using log base 2.
Methods
A total of 225 molecules with a significantly high
expression (fold change ≥2) in HCC were identified for studying the
function and mechanism of AMELY-activated upstream
regulation networks in HCC by systems-computational analysis
of difference with low or non-tumor hepatitis/cirrhotic tissues
using significant analysis of microarrays (SAM; http://www-stat.stanford.edu/~tibs/SAM/)
(20). We selected the two-class
paired data and a minimum fold change ≥2 under the false-discovery
rate of 0%.
The AMELY-activated upstream regulation
network of HCC was analyzed using the Molecule Annotation System,
MAS (CapitalBio Corporation, Beijing, China). The primary databases
of MAS integrated various well-known biological resources, such as
Gene Ontology (http://www.geneontology.org), KEGG (http://www.genome.jp/kegg/), BioCarta (http://www.biocarta.com/), GenMapp (http://www.genmapp.org/), HPRD (http://www.hprd.org/), MINT (http://mint.bio.uniroma2.it/mint/Welcome.do),
BIND (http://www.blueprint.org/), Intact
(http://www.ebi.ac.uk/intact/), UniGene
(www.ncbi.nlm.nih.gov/UniGene) and OMIM
(http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=OMIM).
Biological processes and occurrence numbers of GO in
non-tumor hepatitis/cirrhotic tissues (HBV or HCV infection) with
activated low expression of AMELY upstream regulation networks and
the corresponding HCC tissues with high expression (fold change ≥2)
were identified and computed.
AMELY-activated upstream regulation molecular
networks in non-tumor hepatitis/cirrhotic tissues and HCC were
constructed by GRNInfer (21) and
published studies (22–36), and illustrated by GVedit tool,
respectively.
Results
The biological processes of AMELY-activated upstream
regulation networks in non-tumor hepatitis/cirrhotic tissues and
HCC are presented in Tables I and
II, respectively. The
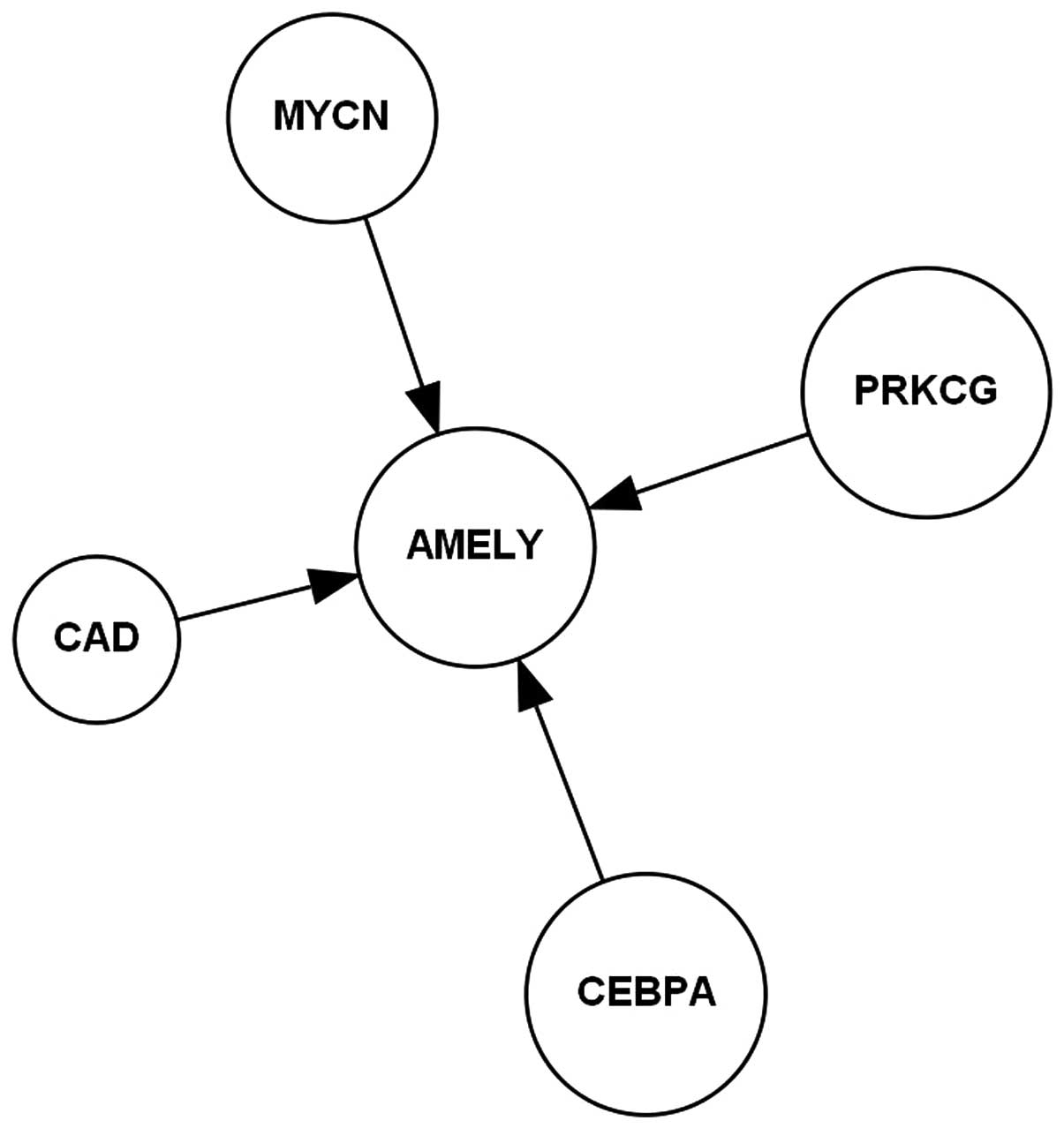
AMELY-activated upstream regulation molecular network
consisted of CAD, CEBPA, MYCN and PRKCG in non-tumor
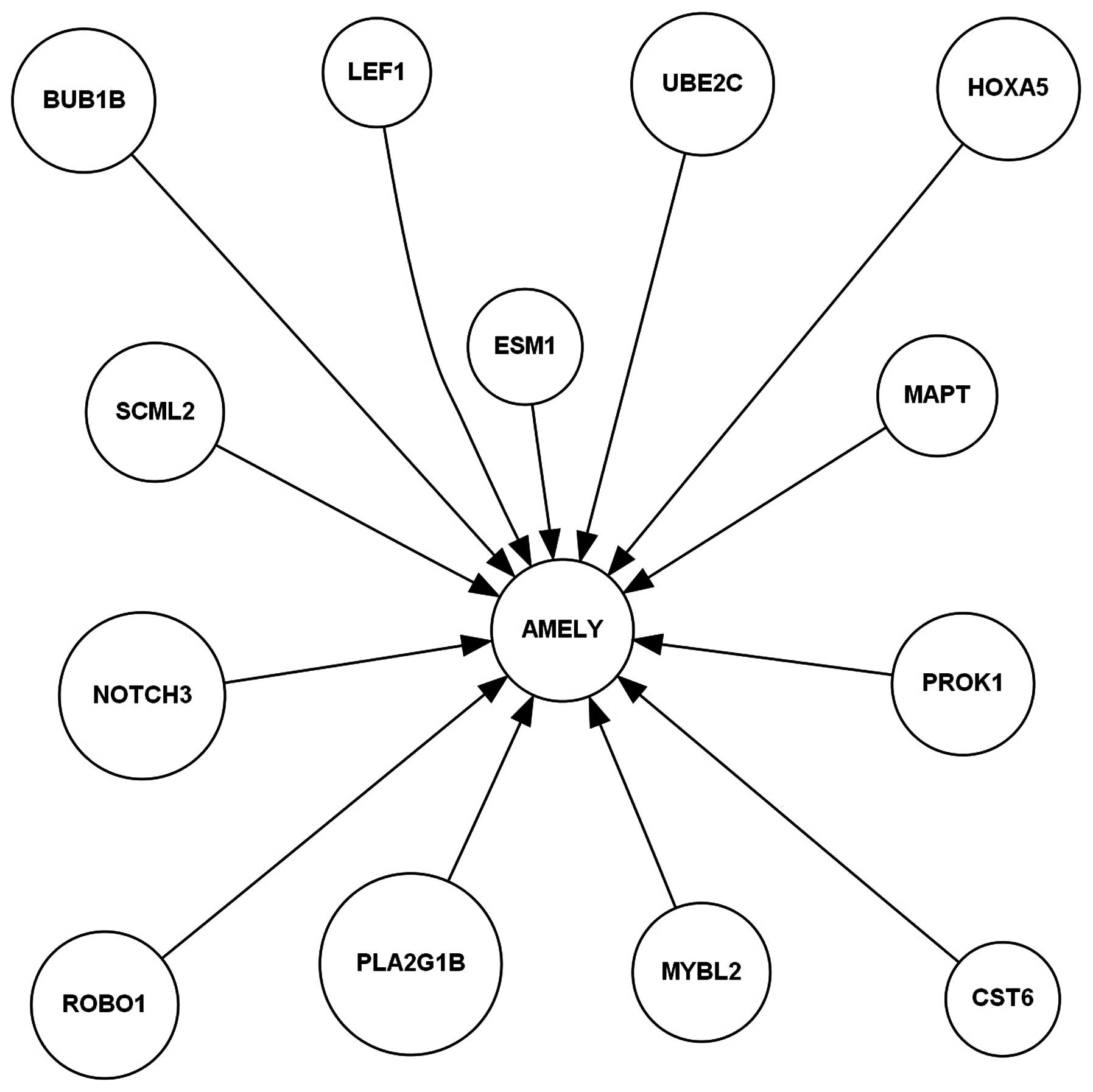
hepatitis/cirrhotic tissues, as shown in Fig. 1. The AMELY-activated upstream
regulation molecular network included BUB1B, CST6, ESM1, HOXA5,
LEF1, MAPT, MYBL2, NOTCH3, PLA2G1B, PROK1, ROBO1, SCML2 and
UBE2C in HCC, as shown in Fig.
2.
 | Table IBiological processes of AMELY
upstream regulation network in non-tumor hepatitis/cirrhotic
tissues using our programming. |
Table I
Biological processes of AMELY
upstream regulation network in non-tumor hepatitis/cirrhotic
tissues using our programming.
| Biological
process | Occurrence of GO
term |
|---|
| Negative regulation
of cell-cell adhesion | 1 |
| Negative regulation
of protein catabolism | 1 |
| Negative regulation
of protein ubiquitination | 1 |
| Positive regulation
of mismatch repair | 1 |
| Positive regulation
of transcription | 1 |
| Regulation of
transcription from RNA polymerase II promoter | 1 |
| Table IIBiological processes of AMELY
upstream regulation network in human HCC using our programming. |
Table II
Biological processes of AMELY
upstream regulation network in human HCC using our programming.
| Biological
process | Occurrence of GO
term |
|---|
| Negative regulation
of endothelial cell differentiation | 1 |
| Negative regulation
of microtubule depolymerization | 1 |
| Negative regulation
of peptidase activity | 1 |
| Negative regulation
of progression through cell cycle | 1 |
| Positive regulation
of calcium ion transport into cytosol | 1 |
| Positive regulation
of cell proliferation | 1 |
| Positive regulation
of DNA replication | 1 |
| Positive regulation
of fibroblast proliferation | 1 |
| Positive regulation
of immune response | 1 |
| Positive regulation
of microtubule polymerization | 1 |
| Positive regulation
of protein secretion | 1 |
| Positive regulation
of specific transcription from RNA polymerase II promoter | 1 |
| Positive regulation
of transcription | 1 |
| Regulation of
angiogenesis | 1 |
| Regulation of cell
growth | 1 |
| Regulation of
protein metabolism | 1 |
| Regulation of Wnt
receptor signaling pathway | 1 |
Discussion
The aim of this study was to elucidate the function
and mechanism of AMELY-activated upstream regulation
networks in HCC using systems-computational analysis of differences
and similarities with non-tumor hepatitis/cirrhotic tissues (HBV or
HCV infection). Biological processes and occurrence numbers of GO
in non-tumor tissues with activated low expression of AMELY
upstream regulation networks and the corresponding HCC tissues with
high expression (fold change ≥2) were identified and computed
(Tables I and II).
We analyzed and compared the biological processes
and occurrence numbers of GO in HCC tissues with high expression
(fold change ≥2) of the AMELY-activated upstream regulation
network and the corresponding non-tumor hepatitis/cirrhotic tissues
with low expression of the activated network. The biological
processes identified solely in non-tumor hepatitis/cirrhotic
tissues included negative regulation of cell-cell adhesion,
transcription from RNA polymerase II promoter, protein
ubiquitination and protein catabolism, as well as positive
regulation of mismatch repair. The processes found only in HCC
consisted of negative regulation of endothelial cell
differentiation, microtubule depolymerization, peptidase activity,
progression through the cell cycle, positive regulation of calcium
ion transport into the cytosol, cell proliferation, DNA
replication, fibroblast proliferation, immune response, microtubule
polymerization, protein secretion, and specific transcription from
RNA polymerase II promoter. There was also regulation of
angiogenesis, cell growth, protein metabolism and the Wnt receptor
signaling pathway. The processes common to both included positive
regulation of transcription. It was therefore postulated that
AMELY-activated coupling upstream positively regulates the
immune response-mediated protein secretion to Wnt signaling and
calcium into cytosol-induced regulation of cell growth and
angiogenesis in HCC.
The correlations between regulation of immune
response, protein secretion, Wnt signaling and calcium movement
into the cytosol with regulation of cell growth or angiogenesis
have been reported. Examples include calcium store sensor
stromal-interaction molecule 1-dependent signaling playing an
important role in cervical cancer growth, migration and
angiogenesis (37); blockade of Wnt
signaling inhibiting angiogenesis and tumor growth in HCC (38); enhancement of the recognized
Wnt/β-catenin signaling activity by HCV core protein promoting cell
growth of HCC cells (39); porous
membrane substrates offering better niches to enhance the Wnt
signaling and promote human embryonic stem cell growth and
differentiation (40); effect of
dietary tea polyphenols on growth performance and cell-mediated
immune response of post-weaning piglets under oxidative stress
(41); Wnt inhibitory factor 1
inducing apoptosis and inhibiting cervical cancer growth, invasion
and angiogenesis in vivo(42); Rspo1/Wnt signaling promoting
angiogenesis via Vegfc/Vegfr3 (43); bone morphogenetic protein 2 inducing
pulmonary angiogenesis via Wnt-β-catenin and Wnt-RhoA-Rac1 pathways
(44); and the correlation between
angiogenesis and the immune response in carcinogenesis and the
progression of malignant disease (45).
AMELY-activated upstream regulation molecular
networks in non-tumor hepatitis/cirrhotic tissues and HCC were
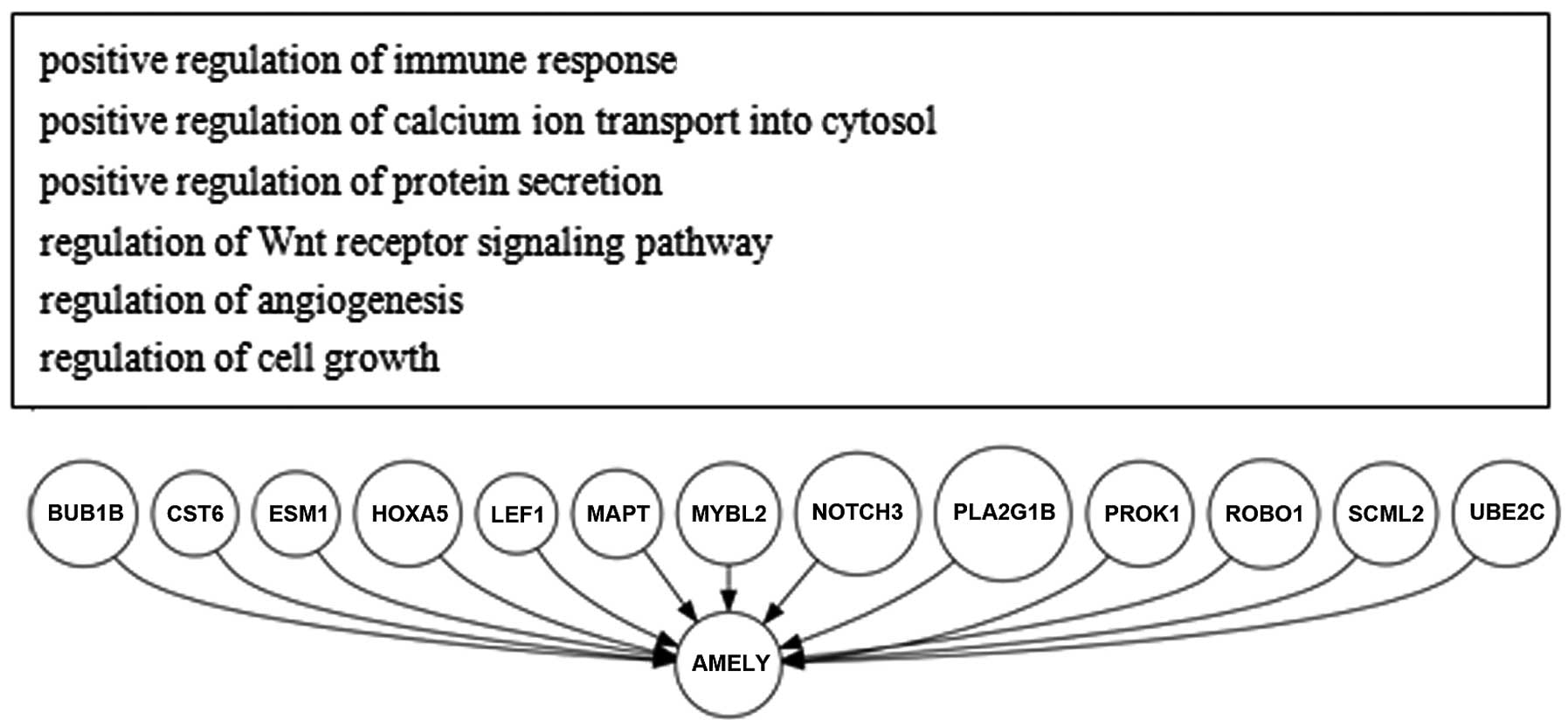
constructed (Figs. 1 and 2). We further constructed a model of the
AMELY upstream regulation molecular network in HCC only,
which included BUB1B, CST6, ESM1,
HOXA5, LEF1, MAPT, MYBL2,
NOTCH3, PLA2G1B, PROK1, ROBO1,
SCML2 and UBE2C, by comparison with the corresponding
activated GO molecular network of non-tumor hepatitis/cirrhotic
tissues (Fig. 3).
In summary, the biological processes and occurrence
numbers of GO in HCC tissue with high expression (fold change ≥2)
of AMELY-activated upstream regulation network were compared
with those of the corresponding non-tumor hepatitis/cirrhotic
tissues with low expression of the activated network. The
biological processes in the non-tumor hepatitis/cirrhotic tissues
included positive regulation of mismatch repair, regulation of
transcription from RNA polymerase II promoter, negative regulation
of cell-cell adhesion, protein ubiquitination and protein
catabolism. The processes in HCC consisted of positive regulation
of calcium ion transport into the cytosol, cell proliferation, DNA
replication, fibroblast proliferation, immune response, microtubule
polymerization, protein secretion and specific transcription from
RNA polymerase II promoter; regulation of angiogenesis, cell
growth, protein metabolism and the Wnt receptor signaling pathway;
and negative regulation of endothelial cell differentiation,
microtubule depolymerization, peptidase activity and progression
through the cell cycle. A common factor included positive
regulation of transcription. Activated AMELY coupling
upstream positive regulation of immune response-mediated protein
secretion to Wnt signaling and calcium into cytosol-induced
regulation of cell growth and angiogenesis network in HCC was
proposed. The AMELY upstream regulation molecular network
model was constructed, including BUB1B, CST6, ESM1, HOXA5, LEF1,
MAPT, MYBL2, NOTCH3, PLA2G1B, PROK1, ROBO1, SCML2 and
UBE2C, in HCC from a GEO dataset by gene regulation network
inference methods and computational programming.
Acknowledgements
This study was supported by the
National Natural Science Foundation of China (No. 61171114), the
Returned Overseas Chinese Scholars for Scientific Research
Foundation of State Education Ministry, the Significant Science and
Technology Project for New Transgenic Biological Species
(2009ZX08012-001B), Automatical Scientific Planning of Tsinghua
University (20111081023 and 20111081010) and the State Key
Laboratory of Pattern Recognition Open Foundation.
References
|
1
|
Hoshida Y, Villanueva A, Kobayashi M, et
al: Gene expression in fixed tissues and outcome in hepatocellular
carcinoma. N Engl J Med. 359:1995–2004. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Veenbergen S and van Spriel AB:
Tetraspanins in the immune response against cancer. Immunol Lett.
138:129–136. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nishikawa M, Otsuki T, Ota A, et al:
Induction of tumor-specific immune response by gene transfer of
Hsp70-cell-penetrating peptide fusion protein to tumors in mice.
Mol Ther. 18:421–428. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ladoire S, Mignot G, Dabakuyo S, et al: In
situ immune response after neoadjuvant chemotherapy for breast
cancer predicts survival. J Pathol. 224:389–400. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fucikova J, Kralikova P, Fialova A, et al:
Human tumor cells killed by anthracyclines induce a tumor-specific
immune response. Cancer Res. 71:4821–4833. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Depianto D, Kerns ML, Dlugosz AA and
Coulombe PA: Keratin 17 promotes epithelial proliferation and tumor
growth by polarizing the immune response in skin. Nat Genet.
42:910–914. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cunha LL, Tincani AJ, Assumpcao LV, Soares
FA, Vassallo J and Ward LS: Interleukin-10 but not interleukin-18
may be associated with the immune response against
well-differentiated thyroid cancer. Clinics (Sao Paulo).
66:1203–1208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zabel BA, Miao Z, Lai NL, et al: CXCR7
protein expression correlates with elevated mmp-3 secretion in
breast cancer cells. Oncol Lett. 1:845–847. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nirde P, Derocq D, Maynadier M, et al:
Heat shock cognate 70 protein secretion as a new growth arrest
signal for cancer cells. Oncogene. 29:117–127. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Milewicz T, Rýs J, Wójtowicz A, et al:
Overexpression of P53 protein and local hGH, IGF-I, IGFBP-3,
IGFBP-2 and PRL secretion by human breast cancer explants. Neuro
Endocrinol Lett. 32:328–333. 2011.PubMed/NCBI
|
|
11
|
Lim JW, Mathias RA, Kapp EA, et al:
Restoration of full-length APC protein in SW480 colon cancer cells
induces exosome-mediated secretion of DKK-4. Electrophoresis.
33:1873–1880. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jube S, Rivera ZS, Bianchi ME, et al:
Cancer cell secretion of the DAMP protein HMGB1 supports
progression in malignant mesothelioma. Cancer Res. 72:3290–3301.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
de Sousa EM, Vermeulen L, Richel D and
Medema JP: Targeting Wnt signaling in colon cancer stem cells. Clin
Cancer Res. 17:647–653. 2011.PubMed/NCBI
|
|
14
|
Corbo C, Orru S, Gemei M, et al: Protein
cross-talk in CD133+ colon cancer cells indicates activation of the
Wnt pathway and upregulation of SRp20 that is potentially involved
in tumorigenicity. Proteomics. 12:2045–2059. 2012.
|
|
15
|
Cha YH, Kim NH, Park C, Lee I, Kim HS and
Yook JI: MiRNA-34 intrinsically links p53 tumor suppressor and Wnt
signaling. Cell Cycle. 11:1273–1281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cai C and Zhu X: The Wnt/β-catenin pathway
regulates self-renewal of cancer stem-like cells in human gastric
cancer. Mol Med Rep. 5:1191–1196. 2012.
|
|
17
|
Boparai KS, Dekker E, Polak MM, Musler AR,
van Eeden S and van Noesel CJ: A serrated colorectal cancer pathway
predominates over the classic WNT pathway in patients with
hyperplastic polyposis syndrome. Am J Pathol. 178:2700–2707. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Andrade Filho PA, Letra A, Cramer A, et
al: Insights from studies with oral cleft genes suggest
associations between WNT-pathway genes and risk of oral cancer. J
Dent Res. 90:740–746. 2011.PubMed/NCBI
|
|
19
|
Sitdikova GF, Weiger TM and Hermann A:
Hydrogen sulfide increases calcium-activated potassium (BK) channel
activity of rat pituitary tumor cells. Pflugers Arch. 459:389–397.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Storey JD: A direct approach to false
discovery rates. J Roy Stat Soc, Ser B. 64:479–498. 2002.
View Article : Google Scholar
|
|
21
|
Wang Y, Joshi T, Zhang XS, Xu D and Chen
L: Inferring gene regulatory networks from multiple microarray
datasets. Bioinformatics. 22:2413–2420. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang J, Wang L, Jiang M and Zheng X:
Interferon α-inducible protein 27 computational network
construction and comparison between the frontal cortex of HIV
encephalitis (HIVE) and HIVE-control patients. Open Genomics J.
3:1–8. 2010.
|
|
23
|
Huang JX, Wang L and Jiang MH: TNFRSF11B
computational development network construction and analysis between
frontal cortex of HIV encephalitis (HIVE) and HIVE-control
patients. J Inflamm (Lond). 7:502010. View Article : Google Scholar
|
|
24
|
Sun L, Wang L, Jiang M, Huang J and Lin H:
Glycogen debranching enzyme 6 (AGL), enolase 1 (ENOSF1),
ectonucleotide pyrophosphatase 2 (ENPP2_1), glutathione
S-transferase 3 (GSTM3_3) and mannosidase (MAN2B2) metabolism
computational network analysis between chimpanzee and human left
cerebrum. Cell Biochem Biophys. 61:493–505. 2011. View Article : Google Scholar
|
|
25
|
Sun Y, Wang L, Jiang M, Huang J, Liu Z and
Wolfl S: Secreted phosphoprotein 1 upstream invasive network
construction and analysis of lung adenocarcinoma compared with
human normal adjacent tissues by integrative biocomputation. Cell
Biochem Biophys. 56:59–71. 2010. View Article : Google Scholar
|
|
26
|
Sun Y, Wang L and Lui L: Integrative
decomposition procedure and Kappa statistics set up ATF2 ion
binding module in Malignant Pleural Mesothelioma (MPM). Frontiers
of Electrical and Electronic Engineering in China. 3:381–387. 2008.
View Article : Google Scholar
|
|
27
|
Wang L, Huang J and Jiang M: CREB5
computational regulation network construction and analysis between
frontal cortex of HIV encephalitis (HIVE) and HIVE-control
patients. Cell Biochem Biophys. 60:199–207. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang L, Huang J and Jiang M: RRM2
computational phospho-protein network construction and analysis
between no-tumor hepatitis/cirrhotic liver tissues and human
hepatocellular carcinoma (HCC). Cell Physiol Biochem. 26:303–310.
2011. View Article : Google Scholar
|
|
29
|
Wang L, Huang J, Jiang M and Lin H:
Tissue-specific transplantation antigen P35B (TSTA3) immune
response-mediated metabolism coupling cell cycle to postreplication
repair network in no-tumor hepatitis/cirrhotic tissues (HBV or HCV
infection) by biocomputation. Immunol Res. 52:258–268. 2012.
View Article : Google Scholar
|
|
30
|
Wang L, Huang J, Jiang M and Lin H: Signal
transducer and activator of transcription 2 (STAT2) metabolism
coupling postmitotic outgrowth to visual and sound perception
network in human left cerebrum by biocomputation. J Mol Neurosci.
47:649–658. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang L, Huang J, Jiang M and Sun L:
Survivin (BIRC5) cell cycle computational network in human no-tumor
hepatitis/cirrhosis and hepatocellular carcinoma transformation. J
Cell Biochem. 112:1286–1294. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang L, Huang J, Jiang M and Sun L: MYBPC1
computational phosphoprotein network construction and analysis
between frontal cortex of HIV encephalitis (HIVE) and HIVE-control
patients. Cell Mol Neurobiol. 31:233–241. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang L, Huang J, Jiang M and Zheng X: AFP
computational secreted network construction and analysis between
human hepatocellular carcinoma (HCC) and no-tumor
hepatitis/cirrhotic liver tissues. Tumour Biol. 31:417–425. 2010.
View Article : Google Scholar
|
|
34
|
Wang L, Sun L, Huang J and Jiang M:
Cyclin-dependent kinase inhibitor 3 (CDKN3) novel cell cycle
computational network between human non-malignancy associated
hepatitis/cirrhosis and hepatocellular carcinoma (HCC)
transformation. Cell Prolif. 44:291–299. 2011. View Article : Google Scholar
|
|
35
|
Gao P, Sun X, Chen X, Subjeck J and Wang
XY: Secretion of stress protein grp170 promotes immune-mediated
inhibition of murine prostate tumor. Cancer Immunol Immunother.
58:1319–1328. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang L, Sun Y, Jiang M and Zheng X:
Integrative decomposition procedure and Kappa statistics for the
distinguished single molecular network construction and analysis. J
Biomed Biotechnol. 2009:7267282009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen YF, Chiu WT, Chen YT, et al: Calcium
store sensor stromal-interaction molecule 1-dependent signaling
plays an important role in cervical cancer growth, migration, and
angiogenesis. Proc Natl Acad Sci USA. 108:15225–15230. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu J, Dong A, Fernandez-Ruiz V, et al:
Blockade of Wnt signaling inhibits angiogenesis and tumor growth in
hepato-cellular carcinoma. Cancer Res. 69:6951–6959. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu J, Ding X, Tang J, et al: Enhancement
of canonical Wnt/β-catenin signaling activity by HCV core protein
promotes cell growth of hepatocellular carcinoma cells. PLoS One.
6:e274962011.
|
|
40
|
Jin S, Yao H, Krisanarungson P, Haukas A
and Ye K: Porous membrane substrates offer better niches to enhance
the wnt signaling and promote human embryonic stem cell growth and
differentiation. Tissue Eng Part A. 18:1419–1430. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Deng Q, Xu J, Yu B, et al: Effect of
dietary tea polyphenols on growth performance and cell-mediated
immune response of post-weaning piglets under oxidative stress.
Arch Anim Nutr. 64:12–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ramachandran I, Thavathiru E, Ramalingam
S, et al: Wnt inhibitory factor 1 induces apoptosis and inhibits
cervical cancer growth, invasion and angiogenesis in vivo.
Oncogene. 31:2725–2737. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gore AV, Swift MR, Cha YR, et al:
Rspo1/Wnt signaling promotes angiogenesis via Vegfc/Vegfr3.
Development. 138:4875–4886. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
de Jesus Perez VA, Alastalo TP, Wu JC, et
al: Bone morphogenetic protein 2 induces pulmonary angiogenesis via
Wnt-β-catenin and Wnt-RhoA-Rac1 pathways. J Cell Biol. 184:83–99.
2009.PubMed/NCBI
|
|
45
|
O’Byrne KJ, Dalgleish AG, Browning MJ,
Steward WP and Harris AL: The relationship between angiogenesis and
the immune response in carcinogenesis and the progression of
malignant disease. Eur J Cancer. 36:151–169. 2000.PubMed/NCBI
|