Introduction
Tumor necrosis factor (TNF)-related
apoptosis-inducing ligand (TRAIL), first identified and cloned by
Wiley et al(1) in 1995, is a
member of the TNF family, and functions by binding to death
receptors and activating the death receptor apoptosis pathway. The
TRAIL gene is located on chromosome 3 at position 3q26, which
consists of 281 amino acids and belongs to a type II membrane
protein (glycoprotein). Its C-terminal extracellular domain shows
clear homology to other TNF family members.
Cumulative evidence from studies demonstrates that
multiple tumor cells are tolerant to TRAIL-induced apoptosis, and
that repeated use of TRAIL may lead to the emergence of resistance
to TRAIL in some sensitive cancer cells (2–5),
limiting the application of TRAIL in the treatment of many tumors.
Reduced expression or mutation of TRAIL death receptor DR4
(6,7), dysfunction of caspase-8 (4), lack of Smac/DIABLO in the cytoplasm or
a reduction in the release of Smac/DIABLO from mitochondria
(8), overexpression of Bcl-2, loss
of Bax or Bak function (3), or
overexpression of the inhibitor of apoptosis (IAP) family of
proteins, including X-linked inhibitor of apoptosis protein (XIAP),
may all lead to the development of resistance of tumor cells to
TRAIL.
Mitochondrial pro-apoptotic protein, as a type of
IAP protein, promotes apoptosis upon stimulation with an apoptotic
signal. The N-terminal Ala-Val-Pro-Ile (AVPI) sequence forms an
important domain to bind to IAPs and exhibit apoptosis, in which
the Ala residue inserts into a pocket of the surface groove on
BIR3, and binds to BIR3 via a hydrogen bond. Smac, on the other
hand, only functions in cytoplasm (9). The AVPI peptide of natural Smac cannot
effectively penetrate the cell membrane, and suffers from problems
of instability, easy degradation, low utilization rate and
inadequate affinity with XIAP. The antennal transcription factor of
Drosophila melanogaster is an important membrane-regulating
protein, in which the protein transduction domain, consisting of
RQIKIWFQNRRMKWKK, has transmembrane transport ability, which may
guide the entry of polypeptides and proteins into target cells
(10).
The present study targeted the AVPI of the protein
Smac. AVPIAQK peptide was synthesized using the Fmoc chemistry
method (solid-phase peptide synthesis), and was then bound to the
protein transduction domain of the antennal transcription factor of
Drosophila melanogaster, with cell membrane penetration via
proline to synthesize the Smac-mimic proapoptotic polypeptide
SmacN7. The effect of exogenous Smac-mimic polypeptide on the
apoptosis of SW1990 cells was assessed at different times following
treatment at various concentrations. In addition, the relationship
between the TRAIL-activated death receptor apoptosis pathway and
the Smac/DIABLO-induced mitochondrial pathway was investigated.
Materials and methods
Design, synthesis and purification of
Smac-mimic polypeptide SmacN7
Based on the four AVPI residues on Smac N-terminus,
an AVPIAQK peptide sequence with a similar KD value to that of the
binding of mature Smac AVPI to XIAP-BIR3 was selected as the
synthesis sequence, whilst the antennal factor sequence
RQIKIWFQNRRMKWKK (10) with cell
membrane penetration served as a vector peptide (AC Scientific,
Inc., Xi’an, China). These were connected via proline. The
synthesis was performed using the standard artificial solid-phase
synthesis protocol on a solid-phase peptide synthesizer. Single
amino acids were sequentially added into the C-terminus of AVPI and
the terminal was labeled with biotin. Resin was used for support of
the solid-phase synthesis, and the flow phase contained 0.1%
trifluoroacetic acid (TFA; Tedia, Pudong, Shanghai, China) aqueous
solution/0.1% trifluoroacetic acid-CH3CN aqueous solution. The
target polypeptide was synthesized through multiple-step reactions.
The synthesized peptide was purified using a high-performance
liquid chromatography system (Waters 600E; Ledon Technologies,
Suzhou, Jiangsu, China) with a column of C18 (20×50 mm). When the
purity was more than 95%, a rotary evaporator was used for vacuum
pumping. After being frozen at a low temperature and dried, the
crystalline solid was collected, weighed using an electronic
balance, packaged separately at 1 mg per bottle, and stored at
−20°C for the subsequent experiment.
Plotting of cell growth curve
The grown cells that were almost fused were
harvested, digested with pancreatin, prepared into suspensions
using new medium and counted. Cells were then transferred into 21
bottles for passage at a concentration of 5×104/ml per
bottle. After 24 h, the cells were counted, and then counted again,
once every 24 h. The number of cells in 3 bottles was counted,
respectively, and the mean number was calculated. The cells were
counted for 7 successive days in total. Based on the cell counts,
the cell growth curve was plotted based on the cell number per unit
(cells/ml, vertical coordinate) and time (horizontal
coordinate).
Drug preparation
The Smac-mimic polypeptide SmacN7 was formulated
into a storage solution at a concentration of 500 μg/ml with
phosphate-buffered saline (PBS); TRAIL (Chemicon, Temecula, CA,
USA) was formulated into a storage solution of 5 μg/ml with
PBS; gemcitabine (Lilly France S.A., Lilly, Fegersheim, France) was
formulated into a storage solution of 60 μmol/l using
RPMI-1640 medium, which was then stored at −20°C, and the working
concentration was prepared using RPMI-1640 medium. The TRAIL was
then diluted into concentrations of 200, 500, 1,000 and 2,500
ng/ml, and gemcitabine was prepared into 10, 20, 40 and 60 mol/l
for the subsequent experiment.
Apoptotic rate detected by flow
cytometry (FCM)
SW1990 cells (Shandong Cancer Institute, Shandong
Province Key Laboratory of Radiation Oncology), at a concentration
of 5×106/ml, were seeded in a 25-ml culture bottle.
SW1990 cells at a concentration of 5×106/ml, were seeded
in a 25-ml culture flask. Then the cells were repeatedly cultured
in five flasks. Cells were placed in an incubator at 37°C
containing 5% CO2 for 24 h. When the majority of the
cells were adherent to the wall of the bottle, the cells were
cultured at 4°C for a further 2 h to promote synchronous cell
growth. Some cells were harvested to prepare single-cell
suspensions, and then treated with the Smac-mimic polypeptide
SmacN7 at a concentration of 500 μg/ml. After 24 h of
treatment, the cells were harvested, transferred into a 5-ml tube,
centrifuged at 1,500 × g for 15 min and then washed in PBS twice.
Cells were then re-suspended in 0.5 ml of pre-cooled 1X binding
buffer, supplemented with 1.25 μl of Annexin V-FITC (200
μg/ml; Alexis Biochemicals, San Diego, CA, USA), incubated
in the dark, at room temperature for 15 min and centrifuged at
1,000 × g at room temperature for 5 min to remove the supernatant.
Cells were then re-suspended in 0.5 ml of pre-cooled 1X binding
buffer, supplemented with 10 μl of propidium iodide (PI),
and detected on a flow cytometer. In the two-parameter scatter plot
diagram of FCM, the Annexin V−/PI cells in the left
lower quadrant served as controls, the Annexin
V+/PI− cells in the right lower quadrant were
defined as apoptotic cells, and the Annexin
V+/PI+ cells in the right upper quadrant were
defined as necrotic cells. The apoptotic rate was then
calculated.
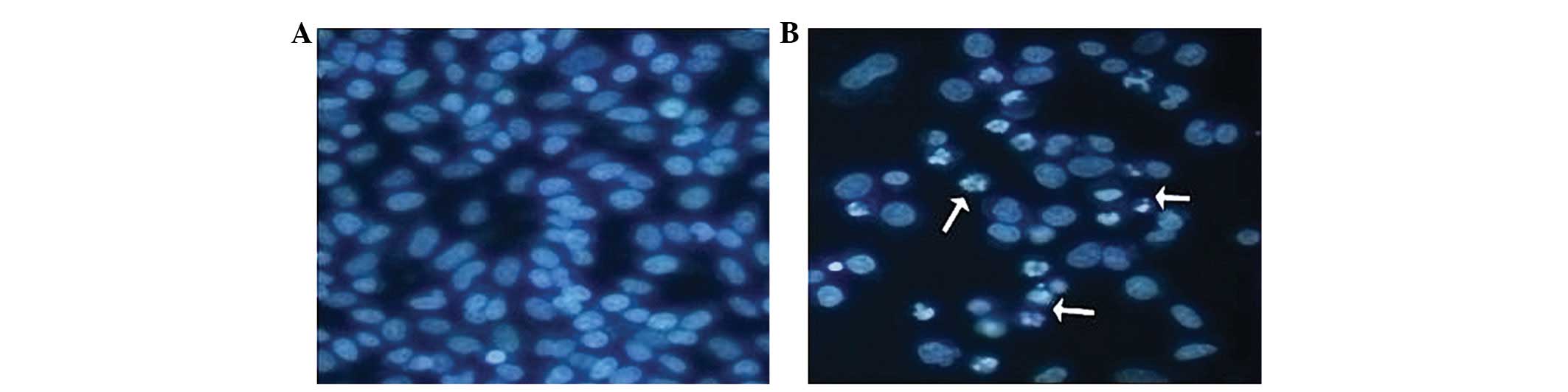
Apoptosis detected by Hoechst 33342
staining
The pancreatic cancer SW1990 cells were seeded on
12-well plates, and cultured routinely until ∼80% were fused.
Physiological saline and 500 μg/ml of SmacN7 were added,
and, after a further culture of 24 h, Hoechst 33342 staining
solution (Invitrogen Life Technologies, Carlsbad, CA, USA) was
added to yield a concentration of 10 μg/ml. Cells were then
cultured in the dark at 37°C for 15–20 min. Under a fluorescence
microscope, the ultraviolet of the Hoechst 33342, excited by a
Krypton laser, produced a blue fluorescence. Although the normal
cells and the middle- and early-stage apoptotic cells were all
stained by Hoechst 33342, the nuclei of the normal cells stained
with Hoechst 33342 appeared round and light blue, whereas the
nuclei of the apoptotic cells appeared bright blue, with a
debris-like shape.
MTT assay detects inhibition of SmacN7 on
growth of pancreatic cancer cell lines
Cell culture. The pancreatic cancer cells were
cultured routinely in 25-ml bottles, and, when 80% of the cells
were fused, were digested using 1 ml of 0.25% trypsin. When
microscopy revealed that the cells had turned round and that some
had dropped from the bottle wall, 4 ml of medium-containing serum
was added to terminate the digestion. The cells were then prepared
into a single-cell suspension in the culture bottle. The cell
concentration and number were counted using the blood count plate
under a microscope. The single-cell suspension was diluted into
solution of 1×105 cells/l with the medium. The
pancreatic cancer cells were seeded on 96-well plates, and 100
μl of the aforementioned single-cell suspension was added to
each well, with the exception of the blank control well. The
pancreatic cancer cells seeded onto the 96-well plates were
routinely cultured until 80% of fusion.
SmacN7 treatment and MTT assay
The SW1990 cells were seeded onto 96-well plates,
supplemented with 50, 100, 200 and 500 μg/ml of SmacN7 and
cultured for 4 h. Mitomycin C (0.05 mg/ml) was then added, leaving
200 μl of total reaction system in each well. Five replicate
wells were set for each concentration, and the cells without drug
treatment served as controls. Following a further culture for 24,
48 and 72 h, 20 μl of MTT solution (5 mg/ml, formulated with
PBS) was added to each well, and the cells were cultured for a
further 4 h. The supernatant was then removed, and 200 μl of
dimethyl sulfoxide (DMSO) was added to each well, prior to 10 min
oscillation. The absorption values A1 and A2 on 550
nm and 630 nm of each well were then measured using the microplate
reader. The A value of each well was defined as
A1-A2, and in the current study, the mean of the
A values in 5 replicate wells was described as the A
value of the well. The cell growth inhibition rate (CGIR) was
calculated using the following formula. CGIR (%)=(1−A value
in the experiment group)/A value ×100. All experiments were
repeated 3 times, and the mean value was calculated.
Combined treatment of SmacN7 and
different concentrations of TRAIL and MTT assay
Cells were cultured as described above. The
inhibition of 500 μg/ml of SmacN7 in combination with TRAIL
at concentrations of 200, 500, 1,000 and 2,500 ng/ml on the growth
of SW1990 cells was assessed.
Combined treatment of SmacN7 and
different concentrations of gemcitabine and MTT assay
Cells were cultured as described above. The
inhibition of 500 μg/ml of SmacN7 in combination with
gemcitabine at concentrations of 10, 20, 40 and 60 μmol/l on
the growth of SW1990 cells was assessed.
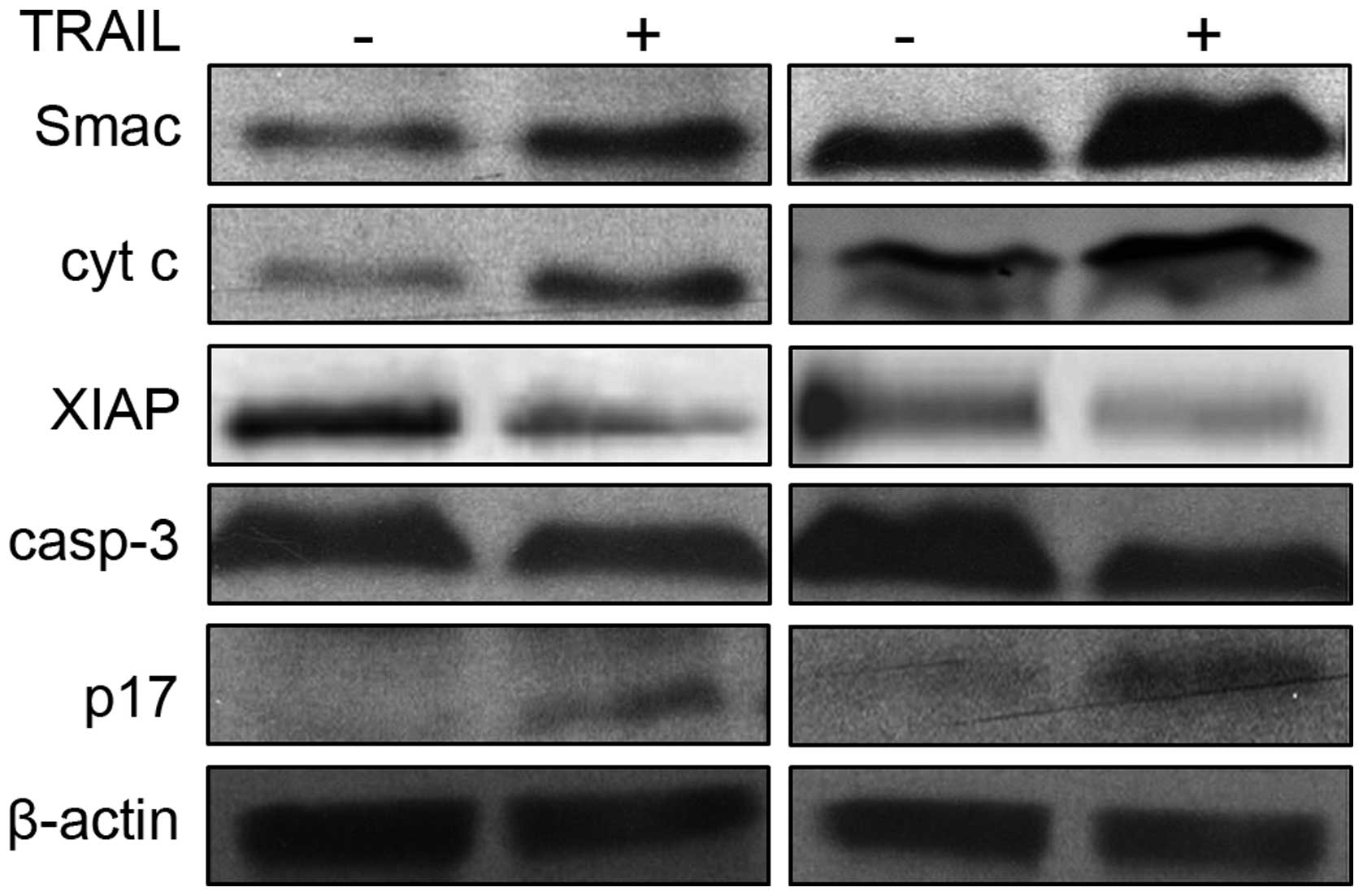
Western blot determines expression of
XIAP, cytochrome C and caspase-3 proteins
After 24 h of combined treatment of SmacN7, TRAIL
(500 ng/ml) and gemcitabine (20 μmol/l), the SW1990 cells
were digested with 0.25% pancreatin. The cell sedimentation was
harvested, combined with 400 μl of lysis buffer,
centrifuged, and then sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) was performed. The electrophoresed
product was transferred onto a nitrocellulose (NC) membrane, which
was then placed into the buffer containing DAB at room temperature.
When the protein color reached the reaction requirement, the
membrane was transferred to another dish to cease the reaction. The
protein expression was analyzed using the digital gel imaging
system.
Statistical analysis
All data were expressed as mean ± standard deviation
(SD), and all statistical analyses were performed using the
statistical software SPSS version 11.5. The difference between the
means of two samples was tested for statistical significance using
a t-test, while one-way analysis of variance in combination with
the least significant difference (LSD) was used to compare multiple
samples. P<0.05 was considered to indicate a statistically
significant result.
Results
Purification and mass spectrometric
identification of SmacN7
SmacN7 is a penetrating fusion polypeptide
synthesized by seven residues AVPIAQK and RQIKIWFQNRRMKWKK on the
protein transduction domain of the antennal transcription factor of
Drosophila melanogaster, with a molecular weight of 3278.08.
Following solid-phase polypeptide purification and reverse-phase
HPLC purification, the purity of the peak of the product reached
more than 95%. Mass spectrometric identification demonstrated that
the molecular ion peak of SmacN7 was fully consistent with the
expected result.
Effect of SmacN7 on morphology and
growth of SW1990 cells
Following treatment with SmacN7 and Hoechst 33342
staining, SW1990 cells showed enlarged cell bodies and nuclei, mild
intracellular swelling and a short-shuttle shape appearance. FCM
detected cell cycle arrest during the G0/G1 phase, reduction in
cell percentage at S phase and slow cell growth (Fig. 1).
Inhibition of SmacN7 on growth of
SW1990 cells
The CGIRs of SW1990 cells caused by treatment of
SmacN7 at concentrations of 50, 100, 200 and 500 μg/ml for
24, 48 and 72 h were determined by MTT assay, and the results
showed that the CGIRs appeared in a time- and
concentration-dependent manner (P<0.05) (Table I).
 | Table IInhibition of SW1990 cell growth 24–72
h after treatment with different concentrations of SmacN7. |
Table I
Inhibition of SW1990 cell growth 24–72
h after treatment with different concentrations of SmacN7.
| Concentration of
SmacN7 (μg/ml) |
|---|
|
|---|
| Time following
treatment (h) | 50 | 100 | 200 | 500 |
|---|
| 24 | 18.11±0.96 | 26.03±1.33 | 37.08±1.26 | 46.19±1.41 |
| 48 | 20.23±1.11 | 38.35±1.13 | 48.96±1.14 | 72.55±1.38 |
| 72 | 22.64±0.80 | 48.01±0.92 | 61.82±1.33 | 77.18±1.17 |
Combined treatment of SmacN7 and
various concentrations of TRAIL or gemcitabine inhibit growth of
SW1990 cells
Combined treatment of SmacN7 at a concentration of
500 μg/ml and TRAIL at concentrations of 200, 500, 1,000 and
2,500 ng/ml for 24 h achieved CGIRs of 318.11%, 37.67%, 42.63% and
67.6% respectively, in comparison to 17.65%, 31.85%, 40.11%, 74.99%
following treatment with SmacN7 in combination with gemcitabine at
concentrations of 10, 20, 40 and 60 μmol/l respectively.
Combined treatment of SmacN7, TRAIL
and gemcitabine on expression of apoptosis-related proteins in
SW1990 cells
Combined treatment of SmacN7 and TRAIL (500 ng/ml)
for 24 h led to elevated expression of Smac/DIABLO, cytochrome C,
and caspase-3 cleavage fragment, p17, and a reduction in XIAP
protein expression. However, the changes in protein expression in
TRAIL-treated SW1990 cells were greater than those in the control
cells (Fig. 2). Following combined
treatment of SmacN7 and gemcitabine (20 μmol/l) for 24 h,
the expression of Smac/DIABLO, cytochrome C and p17 was elevated,
while the XIAP protein expression was reduced, which was similar to
those post-treatment with TRAIL (500 ng/ml). In addition, the
changes in protein expression in TRAIL-treated SW1990 cells were
greater than those in the control cells.
Discussion
Currently, it is widely recognized that there are
two pathways of apoptosis (11–13):
the death receptor pathway and the mitochondrial pathway. In the
death receptor apoptosis pathway, death ligand-like TRAIL binds to
the death receptor (mainly TNF family receptors including TNFR1,
TNFR2, Fas, DR4 and DR5) on the cell surface, then the death domain
in the intracellular fragment of the death receptor attracts an
adaptor protein such as Fas-associated protein with death domain
(FADD) or tumor necrosis factor receptor type 1-associated death
domain protein (TRADD), and recruits caspase-8 and caspase-10
precursors. This leads to the formation of a death-induced signal
complex (DISC) and the production of active initiator caspases,
followed by the activation of effector caspases such as caspase-3,
caspase-6 and caspase-7, thereby inducing apoptosis. In the
mitochondrial apoptosis pathway, many apoptotic signals, such as
DNA damage, external stimuli and some chemotherapeutic drugs, can
induce the release of cytochrome C and Smac/DIABLO from
mitochondria into the cytoplasm. Cytochrome C binds to apoptotic
protease-activating factor (Apaf-1), recruits caspase-9 precursor
to form an apoptosome, produces active caspase-9, and then
activates downstream effector caspases to induce apoptosis.
Smac/DIABLO does not induce apoptosis in normal
cells, but only functions in damaged cells (14). Under normal circumstances,
Smac/DIABLO is predominantly present in the mitochondria, but is
released from the mitochondria into the cytoplasm when cells
undergo apoptotic stimuli such as anti-cancer drugs, or chemical or
physical apoptotic signals. These apoptotic stimuli promote
apoptosis by interacting with IAPs, thus leading to a relief of the
IAP-mediated inhibition of caspases (14,15).
Smac/DIABLO can disrupt the interaction between
XIAP-BIR3 and caspase-9, and Linker-BIR2 and caspase-3 or
caspase-7, leading to a relief of the XIAP-mediated caspase
inhibition. The four N-terminal AVPI residues (Smac-4) of
Smac/DIABLO play a critical role in Smac/DIABLO functions. Numerous
studies have demonstrated that the AVPI sequence is capable of
binding to the BIR domain, which is the functional basis of
Smac/DIABLO (16). It has been
demonstrated that the four N-terminal residues of caspase-9 linker
peptide share significant homology with the N-terminal
tetra-peptide in mature Smac. Therefore, removal of the inhibition
of caspase-9 by XIAP promotes activation of caspase-9 and the
subsequent activation and apoptosis of caspase-3.
There is competition between Smac/DIABLO and IAP.
Therefore, an increase in Smac/DIABLO concentration leads to the
promotion of apoptosis. In addition, Smac/DIABLO may regulate IAP
functions through other mechanisms. It has been indicated that
Smac/DIABLO increases ubiquitin ligase activity of IAP, which
degrades IAP proteins, leading to a reduced amount of IAP. If
Smac/DIABLO can selectively lead to ubiquitination of c-IAP1 and
c-IAP2, resulting in reduction in concentrations of both proteins
(17), a further increase in
relative concentration of Smac/DIABLO may promote apoptosis. In
addition, it has been demonstrated that the co-presence of
Smac/DIABLO and other molecules reduces degradation through the
inhibition of Smac ubiquitination, for example, the interaction
between NADE, previously known as p75NTR-associated cell death
executor, and Smac/DIABLO decreases Smac ubiquitination (18), which is beneficial for Smac/DIABLO
functioning. Based on the TRAIL-regulated apoptosis pathway, it is
considered that Smac/DIABLO plays a critical role in TRAIL-induced
apoptosis (19). The present study
investigated the effect of SmacN7 on cell growth and TRAIL
sensitivity, and found that exogenous Smac-mimic polypeptide
affected the growth of pancreatic cancer SW1990 cells. The
inhibition of treatment with SmacN7 at various concentrations on
SW1990 cells appeared in a time- and concentration-dependent
manner. The TRAIL- or gemcitabine-induced apoptosis of pancreatic
cancer cells, enhanced by Smac-mimic polypeptide, may be associated
with the activity of intracellular pro-apoptotic proteins such as
Smac/DIABLO, cytochrome C, XIAP and caspase-3.
Acknowledgements
This study was financially supported
by Grant No. 30940087 from the Natural Science Foundation of China,
Grant No. 2011YD18047 from Shandong Province Science and Technology
Research Program and Grant No. 2009HZ090 from Shandong Provincial
Medical Health Plan. The authors appreciate the valuable comments
from other members of their laboratories.
References
|
1
|
Wiley SR, Schooley K, Smolak PJ, Din WS,
Huang CP, Nicholl JK, et al: Identification and characterization of
a new member of the TNF family that induces apoptosis. Immunity.
3:673–682. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jin Z, McDonald ER III, Dicker DT and
El-Deiry WS: Deficient tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL) death receptor transport to the
cell surface in human colon cancer cells selected for resistance to
TRAIL-induced apoptosis. J Biol Chem. 279:35829–35839. 2004.
View Article : Google Scholar
|
|
3
|
Zhang L and Fang B: Mechanisms of
resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther.
12:228–237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Platzbecker U, Kurre P, Guardiola P, Ward
JL, Radich JP, Kiem HP and Deeg HJ: Fanconi anemia type C-deficient
hematopoietic cells are resistant to TRAIL (TNF-related
apoptosis-inducing ligand)-induced cleavage of pro-caspase-8. Exp
Hematol. 32:815–821. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Poulaki V, Mitsiades CS, McMullan C,
Fanourakis G, Negri J, Goudopoulou A, et al: Human retinoblastoma
cells are resistant to apoptosis induced by death receptors: role
of caspase-8 gene silencing. Invest Ophthalmol Vis Sci. 46:358–366.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim K, Fisher MJ, Xu SQ and el-Deiry WS:
Molecular determinants of response to TRAIL in killing of normal
and cancer cells. Clin Cancer Res. 6:335–346. 2000.PubMed/NCBI
|
|
7
|
Ozören N, Fisher MJ, Kim K, Liu CX, Genin
A, Shifman Y, et al: Homozygous deletion of the death receptor DR4
gene in a nasopharyngeal cancer cell line is associated with TRAIL
resistance. Int J Oncol. 16:917–925. 2000.PubMed/NCBI
|
|
8
|
Kandasamy K, Srinivasula SM, Alnemri ES,
Thompson CB, Korsmeyer SJ, Bryant JL and Srivastava RK: Involvement
of proapoptotic molecules Bax and Bak in tumor necrosis
factor-related apoptosis-inducing ligand (TRAIL)-induced
mitochondrial disruption and apoptosis: differential regulation of
cytochrome c and Smac/DIABLO release. Cancer Res. 63:1712–1721.
2003.
|
|
9
|
Hasenjäger A, Gillissen B, Müller A,
Normand G, Hemmati PG, Schuler M, et al: Smac induces cytochrome c
release and apoptosis independently from Bax/Bcl-x(L) in a strictly
caspase-3-dependent manner in human carcinoma cells. Oncogene.
23:4523–4535. 2004.PubMed/NCBI
|
|
10
|
Thorén PE, Persson D, Karlsson M and
Nordén B: The antennapedia peptide penetratin translocates across
lipid bilayers - the first direct observation. FEBS Lett.
482:265–268. 2000.PubMed/NCBI
|
|
11
|
Suliman A, Lam A, Datta R and Srivastava
RK: Intracellular mechanisms of TRAIL: apoptosis through
mitochondrial-dependent and -independent pathways. Oncogene.
20:2122–2133. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Saelens X, Festjens N, Vande Walle L, van
Gurp M, van Loo G and Vandenabeele P: Toxic proteins released from
mitochondria in cell death. Oncogene. 23:2861–2874. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vaux DL and Silke J: Mammalian
mitochondrial IAP binding proteins. Biochem Biophys Res Commun.
304:499–504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Verhagen AM, Ekert PG, Pakusch M, Silke J,
Connolly LM, Reid GE, et al: Identification of DIABLO, a mammalian
protein that promotes apoptosis by binding to and antagonizing IAP
proteins. Cell. 102:43–53. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Du C, Fang M, Li Y, Li L and Wang X: Smac,
a mitochondrial protein that promotes cytochrome c-dependent
caspase activation by eliminating IAP inhibition. Cell. 102:33–42.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo F, Nimmanapalli R, Paranawithana S,
Wittman S, Griffin D, Bali P, et al: Ectopic overexpression of
second mitochondria-derived activator of caspases (Smac/DIABLO) or
cotreatment with N-terminus of Smac/DIABLO peptide potentiates
epothilone B derivative-(BMS 247550) and Apo-2L/TRAIL-induced
apoptosis. Blood. 99:3419–3426. 2002. View Article : Google Scholar
|
|
17
|
Creagh EM, Murphy BM, Duriez PJ, Duckett
CS and Martin SJ: Smac/Diablo antagonizes ubiquitin ligase activity
of inhibitor of apoptosis proteins. J Biol Chem. 279:26906–26914.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yoon K, Jang HD and Lee SY: Direct
interaction of Smac with NADE promotes TRAIL-induced apoptosis.
Biochem Biophys Res Commun. 319:649–654. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ng CP and Bonavida B: X-linked inhibitor
of apoptosis (XIAP) blocks Apo2 ligand/tumor necrosis
factor-related apoptosis-inducing ligand-mediated apoptosis of
prostate cancer cells in the presence of mitochondrial activation:
sensitization by overexpression of second mitochondria-derived
activator of caspase/direct IAP-binding protein with low pl
(Smac/DIABLO). Mol Cancer Ther. 1:1051–1058. 2002.
|