Introduction
Hepatocellular carcinoma (HCC) is the third leading
cause of cancer-related mortality worldwide and is commonly
secondary to chronic hepatitis (1,2).
Despite recent therapeutic advances, HCC malignancy continues to be
a significant cause of cancer-related morbidity and mortality, and
it is generally associated with a poor prognosis. The HCC five-year
survival rate is 25–39% post-surgery, with systemic therapy using
cytotoxic agents providing only marginal benefits (3). Multiple pathogenic factors, including
infection with the hepatitis B and C viruses (HBV and HCV) and the
subsequent multistage pathogenesis of HCC have been studied
extensively. In addition, recent advances in molecular genetics
have identified a large number of activated or suppressed genes
that may have significant involvement in the process of
hepatocarcinogenesis (4); however,
the mechanisms by which these factors may promote progression to
HCC are unclear.
Nuclear factor-κB (NF-κB) is a heterodimeric complex
composed of two subunits of the Rel/NF-κB family, as well as other
factors, including NF-κB1 (p50), NF-κB2 (p52), c-Rel, RelA/p65 and
RelB (5,6). NF-κB exists in the cytoplasm in its
inactive form, in association with the IκB regulatory protein. In
response to a variety of stimuli, including inflammatory cytokines,
oncogenes and viruses (7), the
proteome-dependent degradation of IκB promotes the translocation of
NF-κB to the nucleus, the binding of NF-κB to the decameric DNA
sequences and the transcriptional activation of the target genes
(8–10).
Beclin 1 (BECN1), the mammalian orthologue of
yeast Atg6/Vps30, has been mapped to a tumor susceptibility
locus ∼150 kb centromeric to BRCA1 on human chromosome 17q21
(11). It is a coiled-coil protein
that has been identified to act as a tumor suppressor (12). BECN1 is significant in the process
of vesicle nucleation of autophagy in association with Bcl-2 (an
anti-apoptotic protein) (13),
which has been identified to be monoallelically deleted in 40–70%
of sporadic mammary, ovarian and prostate tumors.
Two study groups have generated BECN1-deficient mice
to investigate whether BECN1 acts as a tumor suppressor, and
whether loss of BECN1 may contribute to an increased cancer
incidence (14,15). The results demonstrated that the
loss of BECN1 is correlated with a reduction in autophagic vacuole
formation, and those animals with reduced levels of BECN1 exhibited
an unpredicted increase in epithelial and hematopoietic
malignancies, including HCC. This data led to the conclusion that
BECN1 is a haploinsufficient tumor suppressor gene (16). Two mechanisms by which BECN1
haploinsufficiency promotes cancer are impaired autophagy and
increased cell proliferation (13).
However, the mechanism by which BECN1 modulates cell death in
cancer cells remains unclear. The present study was designed to
investigate the correlation between NF-κBp65 activation and the
expression of BECN1 in patients with HCC.
Materials and methods
Ethics statement
This study was approved by the Ethics Committee of
The First People’s Hospital of Shunde, Southern Medical University
(Shunde, Guandong, China). All protocols were conducted in
accordance with the Declaration of Helsinki (1964). Written
informed consent was obtained from the patients and their
families.
Human liver samples
All cases were obtained from the Department of
Pathology, The First People’s Hospital of Shunde, and comprised
patients with diagnoses from January, 2003 to December, 2007. HCC
tissue samples were obtained from 50 participants (47 males and 3
females; median age, 56.5 years; age range, 28–71 years). According
to the Edmondson grading system, the histopathological analysis
revealed 24 well- or moderately-differentiated tumors (grades 1 and
2) and 26 poorly differentiated or undifferentiated tumors (grades
3 and 4). Liver cirrhosis tissue samples were obtained from 30
participants (22 males and eight females; median age, 50.6 years;
age range, 25–69 years). Liver tissue samples from patients with
HBV were obtained from 30 participants (28 males and two females;
median age, 34.8 years; age range, 6–51 years). Hepatic tissue
samples were also obtained from deceased, previously healthy,
donors. The Streptavidin Peroxidase (SP) Immune Test kit,
anti-NF-κBp65 (mouse monoclonal antibody) and anti-BECN1 (rabbit
polyclonal antibody) were purchased from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). The in situ hybridization test
kit included probes produced by Boster Biological Technology, Ltd.,
Wuhan, China.
SP immunohistochemical staining
SP-immunohistochemistry (SP-IHC) was performed
according to the manufacturer’s instructions. The liver tissue
samples were formalin-fixed, paraffin-embedded and serially
sectioned (4-μm thickness). Following deparaffinization and
rehydration with graded ethanol, immunohistochemistry was
performed. Endogenous peroxidase was quenched with 3%
H2O2 in deionized water for 10 min and then
washed with phosphate-buffered saline (PBS) for 5 min. Antigen
retrieval was performed using ethyl-enediaminetetraacetic acid
(EDTA; pH 8.0) for 3 min in an autoclave at 118°C, followed by
cooling to room temperature. Incubation of the sections in 10%
normal goat serum for 15 min blocked the non-specific binding
sites. The sections were subsequently treated with primary antibody
overnight at 4°C and secondary antibody at 37°C for 30 min. This
was then followed by 3,3′-diaminobenzidine (DAB) visualization.
Following a number of washes, the sections were counter-stained
with hematoxylin. The negative control slides were treated with
PBS.
In situ hybridization
In situ hybridization was performed according
to the manufacturer’s instructions. All equipment and buffers used
were treated with diethylpyrocarbonate (DEPC; Sigma-Aldrich, St
Louis, MO, USA). The liver tissue samples were formalin-fixed,
paraffin-embedded and serially sectioned (6-μm thickness).
Following deparaffinization and rehydration with graded ethanol,
the tissues were digested with 2 μg/ml pepsin for 15 min at
37°C, washed in PBS for 5 min and post-fixed with 4%
paraformaldehyde in PBS for 10 min. Subsequent to being washed with
PBS, the slides were incubated with pre-hybridization solution at
55°C for ≥2 h in a humid chamber. The probes were added to each
tissue section and hybridized at 55°C for 16 h. The slides were
washed twice in 2X saline sodium citrate (SSC) for 10 min at 37°C,
twice in 0.5X SSC for 10 min and twice in 0.2X SSC for 10 min at
55°C. The blocking liquid was then added to the sections, which
were allowed to set at room temperature for 30 min to block out the
non-specific antigen. Subsequently, anti-mouse digoxin was added to
the sections and incubated at 42°C for 4 h, then washed with PBS
three times for 5 min. Streptavidin-biotin complex (SABC;
Sigma-Aldrich) was added to the sections and maintained at room
temperature for 30 min. Horseradish peroxidase (HRP; Sigma-Aldrich)
was then added to the sections and maintained at room temperature
for 30 min, followed by 3-amino-9-ethylcarbazole (AEC)
visualization. Subsequent to a number of washes, the sections were
counterstained with hematoxylin. Placebo probes were added as a
control.
Semi-quantitative method
The total BECN1 and NF-κBp65 staining scores were
calculated as the sum of the percentage of positively stained tumor
cells and the staining intensity scores. Two pathologists
incorporated a double-blind method to quantify the number of
stained cells. The percentage of positively stained cells was then
scored as 1 (<5%, negative), 2 (5–25%, sporadic), 3 (25–50%,
focal) or 4 (>50%, diffuse). The staining intensity was scored
as 1 (no staining), 2 (weakly stained), 3 (moderately stained) or 4
(strongly stained). The percentage of positively stained cells and
the staining intensity were determined utilizing the double-blind
design. The final BECN1 and NF-κBp65 expression scores were
calculated by multiplying the values of the percentage of
positively stained cells and the staining intensity scores; these
values ranged between 1 and 16. The expression level was defined as
follows: − (score of <4), + (score of 4–8), ++ (score of 9–12)
or +++ (score of 13–16).
Pathway analysis
An online search for the pathways related to BECN1
and NF-κBp65 was performed based on the GeneGo database (http://www.genego.com/). The terms used during our
search were “BECN1” and “NF-κB”.
Statistical analysis
The statistical analysis was conducted with SPSS
software, version 10.1 (SPSS, Inc., Chicago, IL, USA). Categorical
variables were analyzed using the χ2 contingency test
and the exact probability test. The Spearman’s rank correlation
test was utilized to reveal the correlation between BECN1 and
NF-κBp65 expression. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of BECN1 in hepatic
tissues
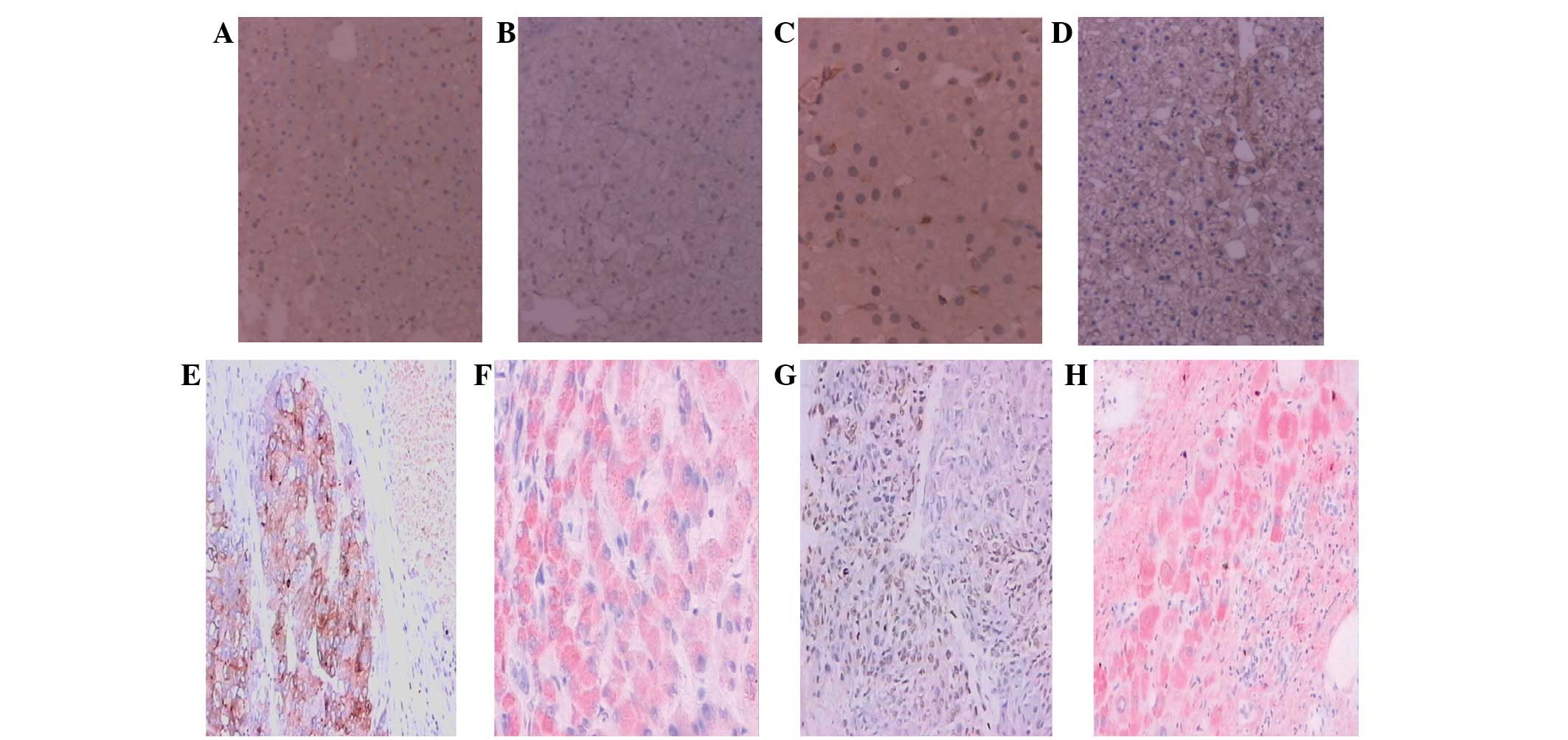
The BECN1 protein was localized in the cytoplasm of
the hepatocytes, stained as brown granules or dots (Fig. 1A, C and E), while BECN1 mRNA
was localized primarily in the cytoplasm of the hepatocytes,
appearing as intense red dots distributed in sheets (Fig 1F). The expression rates of BECN1
protein in the HCC, cirrhotic, hepatitis and normal tissues were
78.00, 26.67, 53.33 and 10.00%, respectively (P<0.05;
χ2=28.34; Table I). The
expression of the BECN1 protein in the HCC tissue was significantly
higher than that of the cirrhotic, hepatitis and normal tissues
(P<0.05; χ2=20.39, 5.31 and 14.42, respectively). The
expression of the BECN1 protein in the hepatitis tissue was
significantly higher than that of the cirrhotic and normal tissues
(P<0.05; χ2=4.44 and 4.12, respectively).
 | Table IExpression of BECN1 in various types
of liver tissue. |
Table I
Expression of BECN1 in various types
of liver tissue.
| | BECN1 protein | | BECN1
mRNA | |
|---|
|
|
|---|
| Tissue type | No. of samples | Negative, n | Positive, n | Positive rate
(%) | Negative, n | Positive, n | Positive rate
(%) |
|---|
| Normal liver | 10 | 9 | 1 | 10.00 | 9 | 1 | 10.00 |
| Hepatitis | 30 | 14 | 16 | 53.33 | 12 | 18 | 60.00 |
| Cirrhosis | 30 | 22 | 8 | 26.67 | 23 | 7 | 23.33 |
| HCC | 50 | 11 | 39 | 78.00 | 16 | 34 | 68.00 |
The positive expression rates of BECN1 mRNA
in the HCC, cirrhotic, hepatitis and normal tissues were 68.00,
23.33, 60.00 and 10.00%, respectively (P<0.05;
χ2=22.61; Table I). The
expression of BECN1 mRNA in the HCC tissue was significantly
higher than that of the cirrhotic and normal tissues (P<0.05;
χ2=14.97 and 9.27, respectively). The expression of
BECN1 mRNA in the hepatitis tissue was significantly higher
than that of the cirrhotic and normal tissues (P<0.05;
χ2=8.30 and 5.65, respectively).
Expression of NF-κBp65 in hepatic
tissues
The NF-κBp65 protein was distributed in the nucleus
and/or cytoplasm of the hepatic cells, stained as brown granules or
dots (Fig. 1B, D and G), while the
NF-κBp65 mRNA was localized mainly in the cytoplasm
of the liver cells, resembling intense red dots distributed in
sheets (Fig. 1H). The positive
expression rates of the NF-κBp65 protein in the HCC, cirrhotic,
hepatitis and normal tissues were 74.00, 36.67, 30.00 and 20.00%,
respectively (P<0.05; χ2=21.42; Table II). The expression of the NF-κBp65
protein in HCC tissue was significantly higher than that of the
cirrhotic, hepatitis and normal tissues (P<0.05;
χ2=10.89, 13.93 and 8.44, respectively).
| Table IIExpression of NF-κBp65 in different
types of liver tissue. |
Table II
Expression of NF-κBp65 in different
types of liver tissue.
| | NF-κBp65
protein | | NF-κBp65
mRNA | |
|---|
|
|
|---|
| Tissue type | No. of samples | Negative, n | Positive, n | Positive rate
(%) | Negative, n | Positive, n | Positive rate
(%) |
|---|
| Normal liver | 10 | 8 | 2 | 20.00 | 9 | 1 | 10.00 |
| Hepatitis | 30 | 20 | 9 | 30.00 | 26 | 4 | 13.33 |
| Cirrhosis | 30 | 19 | 11 | 36.67 | 20 | 10 | 33.33 |
| HCC | 50 | 13 | 37 | 74.00 | 11 | 39 | 78.00 |
The positive expression rates of the
NF-κBp65 mRNA in the HCC, cirrhotic, hepatitis and
normal tissues were 78.00, 33.33, 13.33 and 10.00%, respectively
(P<0.05; χ2=40.75; Table
II). The expression of the NF-κBp65 mRNA in the HCC
tissues was significantly higher than that of the cirrhotic,
hepatitis and normal tissues (P<0.05; χ2=15.76, 31.54
and 14.42, respectively).
HCC gene expression and clinical
features
The correlations between the immunohistochemistry
results and the clinical and pathological findings in the HCC
tissues were evaluated. BECN1/NF-κBp65 gene expression was observed
to be correlated with HCC tumor size (P<0.05), but not with
patient age, Edmondson tumor type, hepatitis B surface antigen
(HBsAg) or tumor metastasis (P>0.05; Table III).
| Table IIICorrelation between BECN1/NF-κBp65
protein expression and the clinical features of patients with
HCC. |
Table III
Correlation between BECN1/NF-κBp65
protein expression and the clinical features of patients with
HCC.
| BECN1
expression | | | NF-κBp65
expression | | |
|---|
|
|
|---|
|
Characteristics | Negative, n | Positive, n | χ2 | P-value | Negative, n | Positive, n | χ2 | P-value |
|---|
| Age (years) | | | | | | | | |
| ≥60 (n=16) | 4 | 12 | 0.00 | P>0.05 | 3 | 13 | 0.21 | P>0.05 |
| <60
(n=34) | 7 | 27 | | | 10 | 24 | | |
| Tumor size
(cm) | | | | | | | | |
| ≤5 (n=19) | 8 | 11 | 5.45 | P<0.05 | 9 | 10 | 5.60 | P<0.05 |
| >5 (n=31) | 3 | 28 | | | 4 | 27 | | |
| Edmondson type | | | | | | | | |
| I/II (n=24) | 3 | 21 | 2.43 | P>0.05 | 2 | 12 | 0.67 | P>0.05 |
| III/IV
(n=26) | 8 | 18 | | | 11 | 25 | | |
| HBsAg | | | | | | | | |
| Positive
(n=42) | 7 | 35 | 2.63 | P>0.05 | 12 | 30 | 0.26 | P>0.05 |
| Negative
(n=8) | 4 | 4 | | | 1 | 7 | | |
| Metastasis | | | | | | | | |
| Positive
(n=14) | 1 | 13 | 1.44 | P>0.05 | 2 | 12 | 0.67 | P>0.05 |
| Negative
(n=36) | 10 | 26 | | | 11 | 25 | | |
Correlation between BECN 1 and NF-κBp65
expression in HCC
As shown in Table
IV, a positive correlation was identified between the
BECN1 and NF-κBp65 mRNA expression levels,
with increases in either one promoting increases in the other
(Spearman’s correlation rank analysis; P<0.05, r=0.676).
| Table IVCorrelation between BECN1 and
NF-κBp65 mRNA expression in HCC. |
Table IV
Correlation between BECN1 and
NF-κBp65 mRNA expression in HCC.
| | NF-κBp65, n
|
|---|
| BECN1, n | No. of samples | − | + | ++ | +++ |
|---|
| − | 16 | 10 | 3 | 2 | 1 |
| + | 8 | 1 | 4 | 2 | 1 |
| ++ | 9 | 0 | 1 | 5 | 3 |
| +++ | 17 | 0 | 1 | 8 | 8 |
| Total | 50 | 11 | 9 | 17 | 13 |
Pathways for BECN1 and NF-κBp65
A pathway search at the GeneGo website found a
comprehensive pathway map for BECN1 and NF-κBp65. It appeared that
the myeloid differentiation primary response gene 88 (MYD88) played
an important role in the pathway, which linked both proteins
through multiple proteins, such as interleukin-1
receptor-associated kinase 1/2 (IRAK1/2), tumor necrosis factor
receptor-associated factor 6 (TRAF6), transforming growth factor-β
activated kinase 1 (TAK1), c-Jun, toll-like receptor 2 (TLR2) and
TLR4.
Discussion
The present study performed immunohistochemical
analyses to determine the expression of the BECN1 and NF-κBp65
proteins in pathogenic and normal hepatic tissues, and to evaluate
a potential correlation between BECN1 and NF-κBp65 expression. The
expression of the BECN1 and NF-κBp65 proteins in the HCC tissue was
significantly higher than that of the cirrhotic, hepatitis and
normal tissues. The expression of BECN1 protein in hepatitis tissue
was significantly higher than that of cirrhotic and normal tissues,
and the BECN1 protein expression was positively correlated with the
NF-κBp65 protein expression in the HCC tissue.
An in situ hybridization analysis was also
performed to detect the expression levels and potential
correlations, the results of which were consistent with the
immunohistochemical analysis. These results suggested that BECN1
and NF-κBp65 may be important in HCC development. A decreased
expression of BECN1 has been identified in human breast
carcinoma, and BECN1 has been suggested to be a mammalian
autophagy gene that may inhibit tumorigenesis (17). This gene has been considered to be a
tumor suppressor gene in breast cancer (17,18).
The present results demonstrated that the expression of BECN1 mRNA
and protein were increased in hepatitis and HCC tissues.
The function of BECN1 in HCC pathogenesis is
unclear. The overexpression of BECN1 has been demonstrated to
inhibit Sindbis virus replication, reduce central nervous system
apoptosis and provide an initial protection against a fatal Sindbis
virus infection. BECN1 may therefore be involved in the host
defences against a Sindbis virus infection (19). The high expression of BECN1 in
hepatitis and HCC tissues may be promoted by viral
infection-induced interferon-γ (20). The overexpression of BECN1 may
prevent hepatocyte apoptosis, in that HBV infection has been
demonstrated to be significant in the development and prognosis of
hepatitis, cirrhosis and HCC (21).
For cirrhotic tissue, BECN1 (functioning as a tumor suppressor
gene) may prevent hepatocyte apoptosis and protect against HBV
infection. As autophagy may be involved in either cell death or
survival, depending on the cellular context (19,22–24),
the increased expression of BECN1 in the hepatitis and HCC tissues
may have implications for its unknown biological role. The present
study identified that the levels of BECN1 and NF-κBp65 expression
in the HCC tissues were not correlated with the clinical and
pathological features, including age, Edmondson type, HBsAg or
metastasis. However, their expression was enhanced in tumors of a
greater size (P<0.05), indicating a potential association with
the pathology of HCC.
In numerous cancer cells, the constitutive
activation of NF-κB lowers cell sensitivity to apoptotic signaling
and consequently to apoptosis, thus favoring neoplastic cell
survival (25). In the present
study, the expression of NF-κBp65 in cancer tissue was
significantly higher than that of the cirrhotic, hepatitis and
normal tissues. A significant correlation was demonstrated between
BECN1 and NF-κBp65 expression in the HCC tissue, suggesting
interactions between the two signaling pathways, which may be
mediated via the Bcl-2 family, with apoptosis as the intersection
of these two pathways. Under certain conditions, apoptosis and
autophagy are able to occur concurrently in the same cell,
indicating the involvement of common regulatory mechanisms
(26). NF-κB (one of the key
regulators of apoptosis) may interact with autophagy. In the
present study, BECN1 and NF-κBp65 expression was also observed in
the endothelial cells. NF-κB plays a key role in inflammatory
disease (10) and may be involved
in autophagy, while autophagy itself may also participate in the
pathogenesis of inflammation and inflammatory disease.
A comprehensive search from GeneGo pathway database
observed a clear link between BECN1 and NF-κBp65 through multiple
proteins, including MYD88, IRAK1/2, TRAF6, TAK1, c-Jun, TLR2 and
TLR4. Many of these proteins are important regulators of cell
proliferation, apoptosis and metabolism and changes in these
proteins due to mutations or exposure to risky factors may
contribute to the pathogenesis of liver cancer.
In conclusion, the results of the present study
indicated that BECN1 and NF-κBp65 are upregulated in primary HCC
and may serve as effective biomarkers for the diagnosis of this
disease. A search from GeneGo pathway database observed a link
between BECN1 and NF-κBp65 through multiple proteins. BECN1 and
NF-κBp65 may interact, contributing to the pathogenesis of HCC,
however, the precise network that controls the crosstalk between
BECN1 and NF-κBp65 is largely unknown. Further studies are required
to delineate the functions of BECN1 and its potential correlation
with NF-κBp65; this may promote a better understanding of the
underlying mechanisms of carcinogenesis and tumor progression in
HCC.
Acknowledgements
This project was supported by a
research fund from The First People’s Hospital, Southern Medical
University, Shunde, Guandong, China.
References
|
1
|
Block TM, Mehta AS, Fimmel CJ and Jordan
R: Molecular viral oncology of hepatocellular carcinoma. Oncogene.
22:5093–5107. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thomas MB and Zhu AX: Hepatocellular
carcinoma: the need for progress. J Clin Oncol. 23:2892–2899. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Frenette C and Gish RG: Hepatocellular
carcinoma: molecular and genomic guideline for the clinician. Clin
Liver Dis. 15:307–321. vii–x. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baeuerle PA and Baltimore D: NF-kappa B:
ten years after. Cell. 87:13–20. 1996.PubMed/NCBI
|
|
6
|
Siebenlist U, Franzoso G and Brown K:
Structure, regulation and function of NF-kappa B. Annu Rev Cell
Biol. 10:405–455. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pikarsky E, Porat RM, Stein I, Abramovitch
R, Amit S, Kasem S, et al: NF-kappaB functions as a tumour promoter
in inflammation-associated cancer. Nature. 431:461–466. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chan CF, Yau TO, Jin DY, Wong CM, Fan ST
and Ng IO: Evaluation of nuclear factor-kappaB, urokinase-type
plasminogen activator, and HBx and their clinicopathological
significance in hepatocellular carcinoma. Clin Cancer Res.
10:4140–4149. 2004. View Article : Google Scholar
|
|
9
|
Karin M and Lin A: NF-kappaB at the
crossroads of life and death. Nat Immunol. 3:221–227. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tak PP and Firestein GS: NF-kappaB: a key
role in inflammatory diseases. J Clin Invest. 107:7–11. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Aita VM, Liang XH, Murty VV, Pincus DL, Yu
W, Cayanis E, et al: Cloning and genomic organization of beclin 1,
a candidate tumor suppressor gene on chromosome 17q21. Genomics.
59:59–65. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Friedman LS, Ostermeyer EA, Lynch ED,
Szabo CI, Anderson LA, Dowd P, et al: The search for BRCA1. Cancer
Res. 54:6374–6382. 1994.PubMed/NCBI
|
|
13
|
Liang XH, Jackson S, Seaman M, Brown K,
Kempkes B, Hibshoosh H and Levine B: Induction of autophagy and
inhibition of tumorigenesis by beclin 1. Nature. 402:672–676. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qu X, Yu J, Bhagat G, Furuya N, Hibshoosh
H, Troxel A, et al: Promotion of tumorigenesis by heterozygous
disruption of the beclin 1 autophagy gene. J Clin Invest.
112:1809–1820. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yue Z, Jin S, Yang C, Levine AJ and Heintz
N: Beclin 1, an autophagy gene essential for early embryonic
development, is a haploinsufficient tumor suppressor. Proc Natl
Acad Sci USA. 100:15077–15082. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Edinger AL and Thompson CB: Defective
autophagy leads to cancer. Cancer Cell. 4:422–424. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liang XH, Yu J, Brown K and Levine B:
Beclin 1 contains a leucine-rich nuclear export signal that is
required for its autophagy and tumor suppressor function. Cancer
Res. 61:3443–3449. 2001.PubMed/NCBI
|
|
18
|
Liang XH, Kleeman LK, Jiang HH, Gordon G,
Goldman JE, Berry G, et al: Protection against fatal Sindbis virus
encephalitis by beclin, a novel Bcl-2-interacting protein. J Virol.
72:8586–8596. 1998.PubMed/NCBI
|
|
19
|
Qian YW, Wang YC, Hollingsworth RE Jr,
Jones D, Ling N and Lee EY: A retinoblastoma-binding protein
related to a negative regulator of Ras in yeast. Nature.
364:648–652. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li P, Du Q, Cao Z, Guo Z, Evankovich J,
Yan W, et al: Interferon-γ induces autophagy with growth inhibition
and cell death in human hepatocellular carcinoma (HCC) cells
through interferon-regulatory factor-1 (IRF-1). Cancer Lett.
314:213–222. 2012.
|
|
21
|
Alison MR, Nicholson LJ and Lin WR:
Chronic inflammation and hepatocellular carcinoma. Recent Results
Cancer Res. 185:135–148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alva AS, Gultekin SH and Baehrecke EH:
Autophagy in human tumors: cell survival or death? Cell Death
Differ. 11:1046–1048. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Baehrecke EH: Autophagy: dual roles in
life and death? Nat Rev Mol Cell Biol. 6:505–510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kroemer G and Jäättelä M: Lysosomes and
autophagy in cell death control. Nat Rev Cancer. 5:886–897. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bours V, Dejardin E, Goujon-Letawe F,
Merville MP and Castronovo V: The NF-kappa B transcription factor
and cancer: high expression of NF-kappa B- and I kappa B-related
proteins in tumor cell lines. Biochem Pharmacol. 47:145–149. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jäättelä M: Multiple cell death pathways
as regulators of tumour initiation and progression. Oncogene.
23:2746–2756. 2004.PubMed/NCBI
|