Introduction
Matrine is an alkaloid isolated from Sophora
flavescens, which has multiple functions, including acting as
an analgesic reagent or against infection by pathogenic
microorganisms (1–6). Matrine may also be used as an
antioxidant that acts by promoting cell metabolism and regulating
immune activities (7–10). It has been demonstrated that matrine
has therapeutic effects on a variety of solid tumors, including
breast, lung, stomach, esophageal, colorectal, cervical and ovarian
cancer, as well as malignant lymphoma (11–13).
However, the molecular mechanism underlying the antitumor function
of matrine remains unclear.
The cellular nuclear factor-κB (NF-κB) signaling
pathway is essential in various cellular processes, including cell
survival, proliferation and apoptosis, which are important for the
development of various types of human cancers (14–16).
Under unstimulated conditions, the human NF-κB transcription
factors are bound by the inhibitor of κB (IκB) proteins (17). However, pathological stimuli or
environmental factors may result in the activation of NF-κB.
Activation of IκB kinases (IKKs), including IKKα and IKKβ, results
in the phosphorylation of IκB and its subsequent
ubiquitin-dependent degradation by the proteasomal pathway
(18,19). The released NF-κB transcription
factors then translocate to the nucleus to regulate the expression
of genes encoding cytokines, cytokine receptors and apoptotic
regulators (20,21).
IKKβ has been demonstrated to be involved in
development of numerous types of human tumors (22,23).
In the present study, the effects of matrine treatment on multiple
breast cancer cell lines, including ER-positive MCF7 cells,
HER2-positive BT-474 cells and the highly metastatic MDA-MB-231
cell line, were determined. It was observed that the matrine
treatment resulted in the death of the three types of cancer cells,
but significantly less toxicity was observed in the control cancer
cells. Our results suggest that matrine may be an effective
approach for treating breast cancer in the future upon further
research.
Materials and methods
Reagents and cell lines
Matrine (chemical formula,
C15H24N2O; molecular weight,
248.36) was purchased from Sigma (cat. no. M5319-100MG; St. Louis,
MO, USA). Matrine was dissolved in RPMI-1640 medium for use (1–4
mM). Three breast cancer cell lines, ER-positive MCF7 cells,
HER2-positive BT-474 cells and the highly metastatic MDA-MB-231
cell line, were provided by the Department of Oncology, Hospital of
Traditional Chinese Medicine (Yantai, China). MCF-7 cells, BT-474
cells and MDA-MB-231 cells were cultured in α-MEM, RPMI and DMEM
(Sigma-Aldrich Co., Ltd., Irvine, CA, USA), respectively. The cells
were cultured at 37°C with 5% CO2 and 100% humidity. The
medium was supplemented with 10% fetal bovine serum (FBS; Hyclone,
Logan, UT, USA), 100 U/ml penicillin and 100 μg/ml
streptomycin.
Cell treatment and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
Briefly, cells were seeded into six-well plates in
medium at a density of 1×105 cells/well and cultured for
24 h. The cells were then treated with matrine (0, 1, 2 and 3 mM).
The untreated cells were used as negative controls. Upon
termination of each experiment (after 48 h), the cells were
incubated with 0.5 mg/ml MTT for 4 h, according to the
manufacturer’s instructions. The viability of the treated cells was
expressed as relative to that of the control cells (relative
viability).
Apoptosis assay
Cells, at a density of 1×105 cells/well,
were cultured in six-well plates in medium supplemented with 10%
calf serum for 24 h, followed by the addition of matrine (0, 1, 2
and 3 mM) or fresh medium (the untreated control). After 48 h, the
cells were pelleted by centrifugation, washed twice with
phosphate-buffered saline (PBS), fixed by incubation in 4%
paraformaldehyde for 30 min at room temperature, and washed again
with PBS to remove the fixative. The fixed cells were resuspended
in PBS containing Hoescht 33258 (5 μg/ml), followed by an
incubation at room temperature for 15 min in the dark. The cells
were placed on glass slides and examined for those with apoptotic
morphology (nuclear condensation and chromatin fragmentation) via
fluorescence microscopy (Labomed LX 400 fluorescence microscope;
Labomed Inc., Culver City, CA, USA). To quantify the apoptosis, 250
nuclei from random microscopic fields were analyzed. Data are
presented as the mean percentages of apoptotic cells.
Western blot analysis
Cells, at a density of 1×105 cells/well,
were cultured in six-well plates in medium supplemented with 10%
calf serum for 24 h, followed by the addition of matrine (0, 1, 2
and 3 mM) or fresh medium (the untreated control). After 48 h, the
cells were pelleted by centrifugation and washed twice with PBS.
Total proteins were harvested from the cells, then separated on 10%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) gels and subjected to immunoblot analyses. The primary
antibodies against IKKβ (∼90 kDa) and β-actin were purchased from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA; anti-IKKβ,
cat. no. sc-8014, 1:200; anti-β-actin, cat. no. sc-130301,
1:10,000). The secondary antibody used in the present study was
goat anti-mouse IgG-horseradish peroxidase (HRP) (cat. no. sc-2005,
1:10,000; Santa Cruz Biotechnology, Inc.). Bound antibodies were
detected using the ECL system (Cat No. 32134; Pierce Biotechnology,
Inc., Rockford, IL, USA). The immunoblot experiments were repeated
at least three times. The mean normalized optical densities (ODs)
of the IKKβ protein bands relative to the ODs of the β-actin bands
from the same condition were calculated.
Statistical analysis
The experimental data are presented as the mean ±
standard error (SEM). Statistical software (SPSS 12.0; SPSS, Inc.,
Chicago, IL, USA) was used to perform independent sample t-tests,
followed by one-way analysis of variance. P<0.05 was considered
to indicate a statistically significant difference.
Results
Matrine is toxic to breast cancer cell
lines
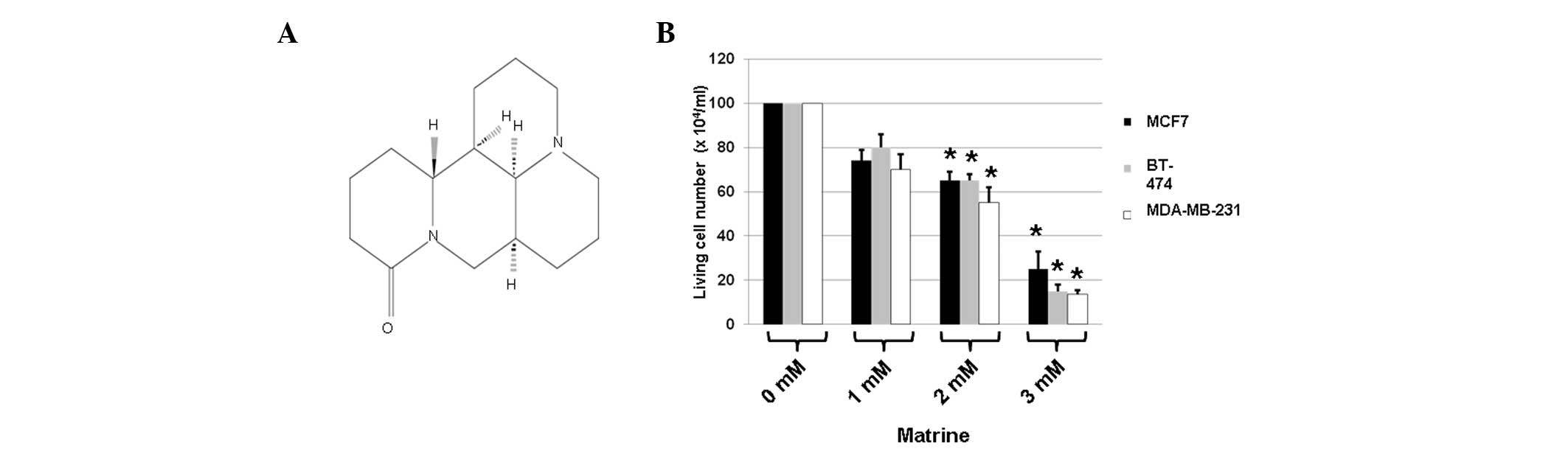
To determine whether matrine (Fig. 1A) is toxic to breast cancer cell
lines, ER-positive MCF7 cells, HER2-positive BT-474 cells and the
highly metastatic MDA-MB-231 cell line were treated with medium
only (matrine, 0 mM) or matrine (1, 2 or 3 mM). The cell viability
was measured using the MTT assay immediately following 48 h of
incubation with matrine. The treatment with medium alone served as
a control, as the matrine used in the remaining groups was
dissolved in medium. The cells were analyzed for differences in
cell death following the various matrine treatments by counting the
number of living cells in the presence or absence of the
aforementioned compounds, using the MTT assay.
The results showed that, in comparison with the
untreated cells, the 48 h-treatment with matrine decreased the cell
viability of all three types of cancer cells (Fig. 1B). Treatment with 1 mM matrine for
48 h had inhibitory effects on the cell viability of all three
types of cells, leading to reductions in such cell numbers (to
71.5–80.2%) compared with the controls (Fig. 1B). Treatment with 2 mM matrine for
48 h had clear inhibitory effects on the cell viability of all
three types of cells, leading to reductions in such cell numbers
(to 57.6–63.2%) compared with the controls (Fig. 1B). Finally, treatment with 3 mM
matrine resulted in reductions in such cell numbers (to 15.5–23.6%)
compared with the controls (Fig.
1B). Among the three types of cells, MDA-MB-231 cells were the
most sensitive to treatment with matrine (Fig. 1B). These results suggested that
matrine exerted significant toxic effects on the breast cancer
cells.
Matrine induces apoptosis in breast
cancer cells
As matrine exerted toxic effects on ER-positive MCF7
cells, HER2-positive BT-474 cells and highly metastatic MDA-MB-231
cells, the effects of the compound on apoptosis were determined in
all of three types of cells. The cells were treated with medium
only (matrine, 0 mM) or matrine (1, 2 or 3 mM) for 48 h. To
quantify the apoptotic incidence, a fluorescence microscopic assay
was used following staining of the drug-treated cells with Hoescht
33258.
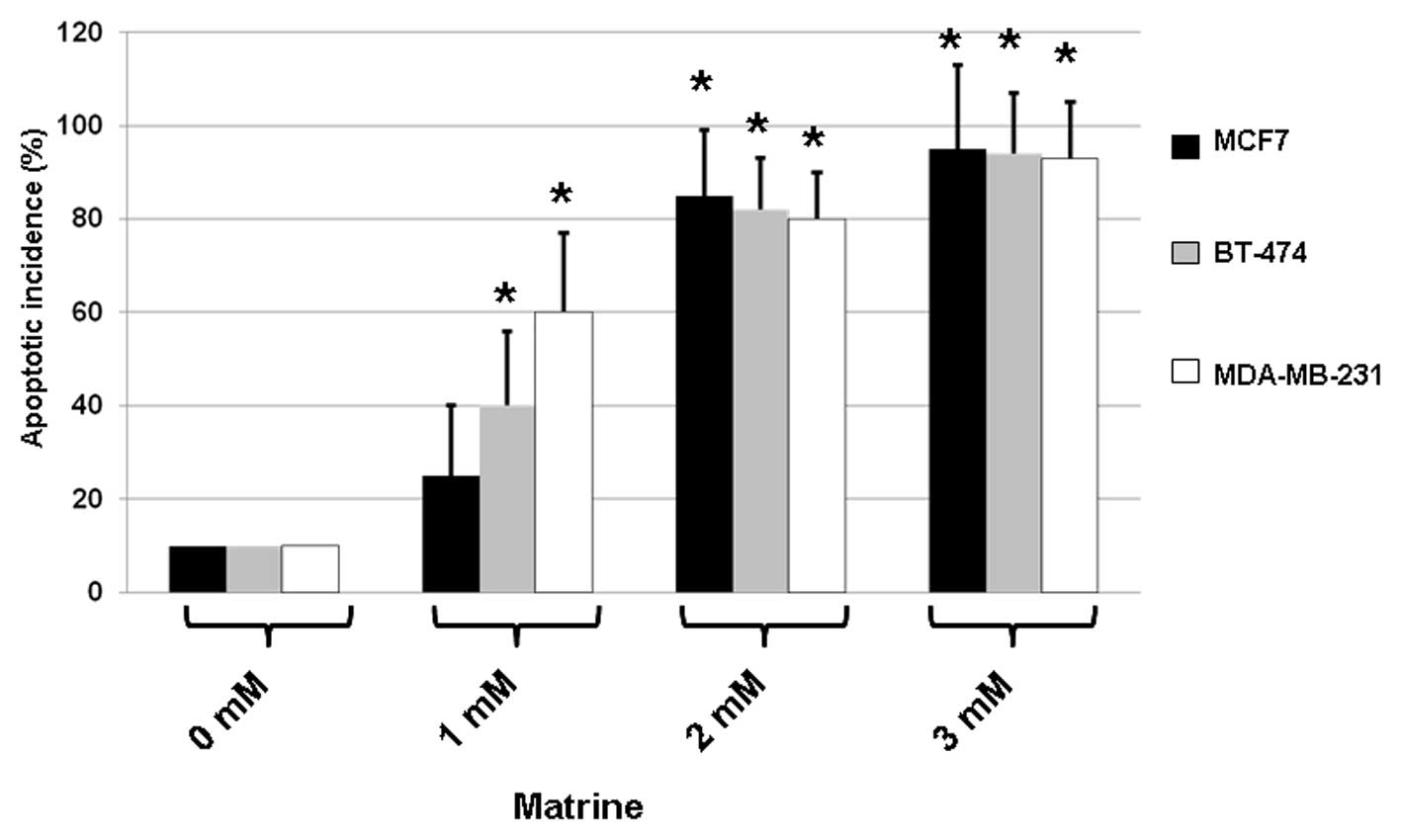
As shown in Fig. 2,
treatment with matrine resulted in increases in the apoptosis of
all three types of cells. When compared with the untreated control,
matrine (3 mM) caused the apoptosis of MCF7, BT-474 and MDA-MB-231
cells with incidences of ∼90%. These results indicate that matrine
significantly elevated apoptosis in treated cells.
Matrine treatment leads to the
degradation of IKKβ
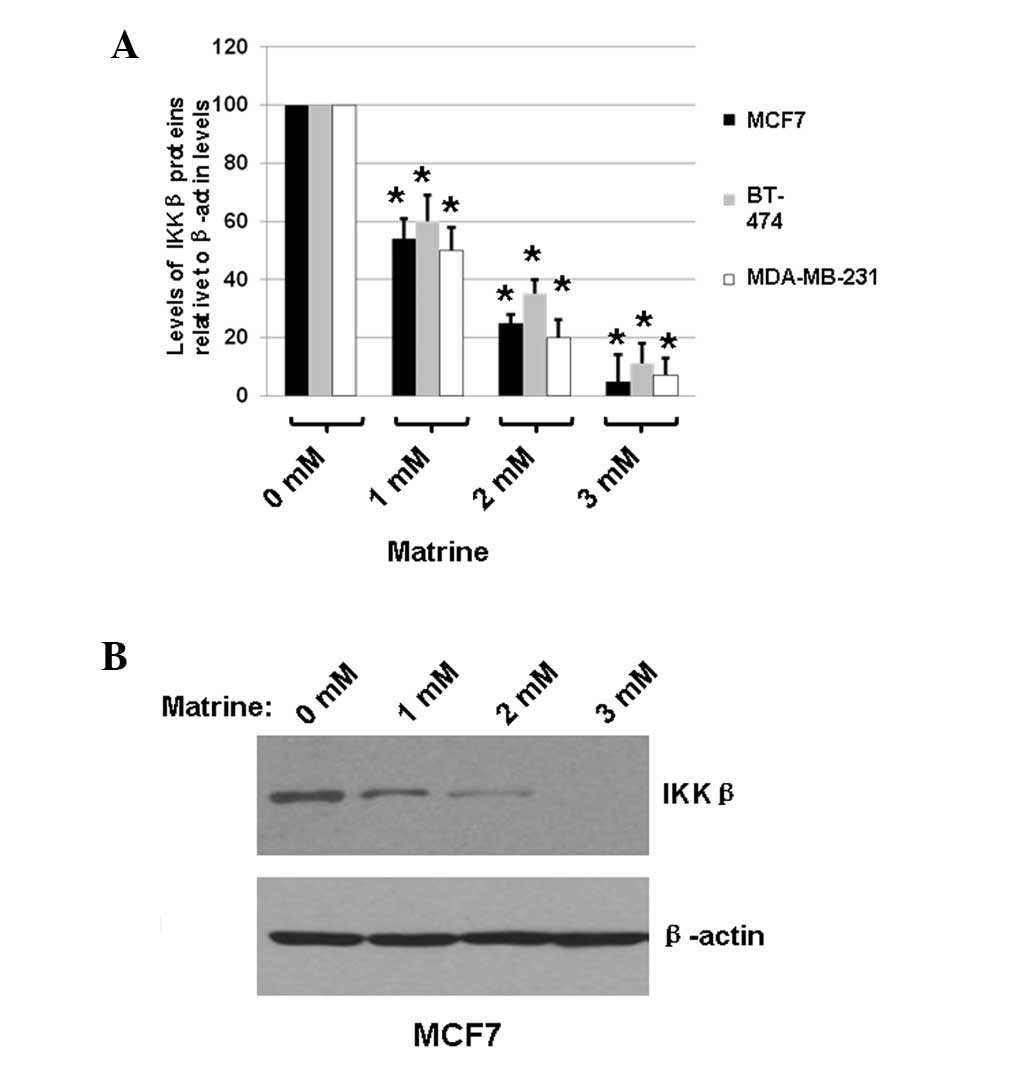
To determine whether matrine inhibited the
expression of IKKβ in MCF7, BT-474 and MDA-MB-231 cells, the cells
were treated with medium only (matrine, 0 mM) or matrine (1, 2 or 3
mM) for 48 h. The total proteins were extracted and the expression
levels of IKKβ were determined using immunoblot analysis, with the
cellular β-actin protein serving as a loading control. The mean
normalized ODs of the IKKβ protein bands relative to the ODs of the
β-actin bands from the same condition were calculated and subjected
to statistical analyses. The calculated ratios of the levels of
IKKβ proteins relative to the β-actin levels are shown in Fig. 3A. A representative blot is shown in
Fig. 3B.
As shown in Fig. 3,
treating MCF7, BT-474 and MDA-MB-231 cells with matrine decreased
the expression of IKKβ by ≤95%, according to the calculated OD
values of the IKKβ bands relative to the β-actin bands. These
results indicated that matrine significantly decreased IKKβ
expression in the treated breast cancer cells, suggesting that
matrine effectively inhibited the proliferation of breast cancer
cells by a mechanism associated with the NF-κB signaling
pathway.
Discussion
Matrine has been demonstrated to possess multiple
functions, including acting as an analgesic reagent or against
infection by pathogenic microorganisms (1–6). It
may also be used as an antioxidant, as it promotes cell metabolism
and regulates immune activity (7–9). As
matrine has therapeutic effects on various solid tumors, including
liver, lung, stomach, esophageal, colorectal, cervical and ovarian
cancer, as well as malignant lymphoma (11–13),
the present study investigated whether matrine has antitumor
effects on three breast cancer cell lines, ER-positive MCF7 cells,
HER2-positive BT-474 cells and highly metastatic MDA-MB-231
cells.
In the present study, cell viability was measured
using the MTT assay immediately following two days of incubation
with matrine. Treatment with 1 mM matrine for 48 h exerted
inhibitory effects on the cell viability of all three types of
cells, leading to 19.8–28.5% reductions in cell numbers.
Furthermore, treatment with 3 mM matrine resulted in 76.4–84.5%
reductions in cell numbers. Of the three types of cells, MDA-MB-231
cells were the most sensitive to treatment. The results indicated
that matrine reduced the cell viability in a
concentration-dependent manner. Furthermore, treatment with matrine
resulted in apoptosis. Treatment with matrine also resulted in
increases in the apoptotic index of all three types of cells.
Compared with the untreated control, matrine (3 mM) caused the
apoptosis of MCF7, BT-474 and MDA-MB-231 cells with incidences of
∼90%, indicating that matrine significantly increased the levels of
apoptosis in the treated cells.
Treatment of MCF7, BT-474 and MDA-MB-231 cells with
matrine decreased the expression of IKKβ by ≤95%, according to the
calculated OD values of the IKKβ bands relative to the cellular
protein β-actin bands. These results indicated that matrine
significantly decreased IKKβ expression in the treated breast
cancer cells, suggesting that matrine effectively inhibited the
proliferation of breast cancer cells by a mechanism associated with
IKKβ (24). In conclusion, the
present results suggested that matrine may be a promising reagent
for treating breast cancer in the future, following further
research.
Acknowledgements
The present study was supported by a
Shandong research grant (No. 6231).
References
|
1.
|
Chen F and Huang K: Effects of the Chinese
medicine matrine on experimental C. parvum infection in
BALB/c mice and MDBK cells. Parasitol Res. 111:1827–1832. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Wan XY, Luo M, Li XD and He P:
Hepatoprotective and anti-hepatocarcinogenic effects of
glycyrrhizin and matrine. Chem Biol Interact. 181:15–19. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Jong TT, Lee MR, Chiang YC and Chiang ST:
Using LC/MS/MS to determine matrine, oxymatrine, ferulic acid,
mangiferin, and glycyrrhizin in the Chinese medicinal preparations
Shiau-feng-saan and Dang-guei-nian-tong-tang. J Pharm Biomed Anal.
40:472–477. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Yin LL and Zhu XZ: The involvement of
central cholinergic system in (+)-matrine-induced antinociception
in mice. Pharmacol Biochem Behav. 80:419–425. 2005.
|
|
5.
|
Kobashi S, Takizawa M, Kubo H, Yamauchi T
and Higashiyama K: Antinociceptive effects of
N-acyloctahydropyrido[3,2,1-ij] [1,6]naphthyridine in mice:
structure-activity relation study of matrine-type alkaloids part
II. Biol Pharm Bull. 26:375–379. 2003.PubMed/NCBI
|
|
6.
|
Yang Y, Xiu J, Zhang X, Zhang L, Yan K,
Qin C and Liu J: Antiviral effect of matrine against human
enterovirus 71. Molecules. 17:10370–10376. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Zhang S, Zhang Y, Zhuang Y, et al: Matrine
induces apoptosis in human acute myeloid leukemia cells via the
mitochondrial pathway and akt inactivation. PLoS One. 7:e468532012.
View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Fu S, Sun C, Tao X and Ren Y:
Anti-inflammatory effects of active constituents extracted from
Chinese medicinal herbs against Propionibacterium acnes. Nat
Prod Res. 26:1746–1749. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Zhao X, Kan Q, Zhu L and Zhang GX: Matrine
suppresses production of IL-23/IL-17 and ameliorates experimental
auto-immune encephalomyelitis. Am J Chin Med. 39:933–941. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Xu M, Yang L, Hong LZ, Zhao XY and Zhang
HL: Direct protection of neurons and astrocytes by matrine via
inhibition of the NF-κB signaling pathway contributes to
neuroprotection against focal cerebral ischemia. Brain Res.
1454:48–64. 2012.PubMed/NCBI
|
|
11.
|
Zhang J, Li Y, Chen X, et al: Autophagy is
involved in anticancer effects of matrine on SGC-7901 human gastric
cancer cells. Oncol Rep. 26:115–124. 2011.PubMed/NCBI
|
|
12.
|
Liu XY, Ruan LM, Mao WW, Wang JQ, Shen YQ
and Sui MH: Preparation of RGD-modified long circulating liposome
loading matrine, and its in vitro anti-cancer effects. Int J Med
Sci. 7:197–208. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Yu P, Liu Q, Liu K, Yagasaki K, Wu E and
Zhang G: Matrine suppresses breast cancer cell proliferation and
invasion via VEGF-Akt-NF-kappaB signaling. Cytotechnology.
59:219–229. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Kelliher MA, Grimm S, Ishida Y, Kuo F,
Stanger BZ and Leder P: The death domain kinase RIP mediates the
TNF-induced NF-κB signal. Immunity. 8:297–303. 1998.
|
|
15.
|
Ting AT, Pimentel-Muiños FX and Seed B:
RIP mediates tumor necrosis factor receptor 1 activation of NF-κB
but not Fas/APO-1-initiated apoptosis. EMBO J. 15:6189–6196.
1996.
|
|
16.
|
Zhang J and Winoto A: A mouse
Fas-associated protein with homology to the human Mort1/FADD
protein is essential for Fas-induced apoptosis. Mol Cell Biol.
16:2756–2763. 1996.PubMed/NCBI
|
|
17.
|
Karin M, Yamamoto Y and Wang QM: The IKK
NF-κB system: a treasure trove for drug development. Nat Rev Drug
Discov. 3:17–26. 2004.
|
|
18.
|
Yang J, Lin Y, Guo Z, et al: The essential
role of MEKK3 in TNF-induced NFκB activation. Nat Immunol.
2:620–624. 2001.PubMed/NCBI
|
|
19.
|
Wu MX, Ao Z, Prasad KV, Wu R and
Schlossman SF: IEX-1L, an apoptosis inhibitor involved in
NF-κB-mediated cell survival. Science. 281:998–1001. 1998.
|
|
20.
|
Wang CY, Mayo MW, Korneluk RG, Goeddel DV
and Baldwin AS Jr: NFκB antiapoptosis: induction of TRAF1 and TRAF2
and c-IAP1 and c-IAP2 to suppress caspase-8 activation. Science.
281:1680–1683. 1998.
|
|
21.
|
Kamata H, Honda S, Maeda S, Chang L,
Hirata H and Karin M: Reactive oxygen species promote
TNFalpha-induced death and sustained JNK activation by inhibiting
MAP kinase phosphatases. Cell. 120:649–661. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Broemer M, Krappmann D and Scheidereit C:
Requirement of Hsp90 activity for IkappaB kinase (IKK) biosynthesis
and for constitutive and inducible IKK and NF-kappaB activation.
Oncogene. 23:5378–5386. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Lewis J, Devin A, Miller A, et al:
Disruption of hsp90 function results in degradation of the death
domain kinase, receptor-interacting protein (RIP), and blockage of
tumor necrosis factor-induced nuclear factor-kappaB activation. J
Biol Chem. 275:10519–10526. 2000. View Article : Google Scholar
|
|
24.
|
Park DH, De Xu H, Shim J, et al:
Stephania delavayi Diels inhibits breast carcinoma
proliferation through the p38 MAPK/NF-κB/COX-2 pathway. Oncol Rep.
26:833–841. 2011.
|