Introduction
Vaccines greatly reduce the spread and mortality of
infectious diseases and cancer. The ability of vaccines to
counteract infections and fight against cancer mainly involves a
three-tiered functionality; humoral and cellular immunity and
regulators of the immune system, such as cytokines (1). Vaccines usually require additional
exogenous adjuvants to improve the immune response to the antigens
following immunization (2). Hence,
one of the most significant challenges in vaccinology is the
selection of suitable adjuvants (3). The aluminum (alum) adjuvant has been
successfully used in millions of humans as the principle adjuvant
in clinical vaccines since 1932 and greatly decreases morbidity and
mortality with minimal toxicity (2). However, alum is a relatively poor
adjuvant in numerous situations, particularly when used with
subunit antigens (4). Furthermore,
alum is not highly effective at stimulating cell-mediated immune
responses (5), including the
vaccination against pathogens that require Th1-cell-mediated
immunity (2). Freund’s complete
adjuvant (6,7) promotes a marked commitment to the Th1
pathway, but has a generally unacceptable level of adverse effects.
The MF59 oil-in-water emulsion adjuvant is the only other adjuvant
approved for human use besides alum, although it has been reported
to favor only the Th2 immune response, similar to alum (8). In addition, alum is not valid for the
induction of mucosal IgA antibody responses and certain studies
have shown that alum is associated with allergic reactions in
certain subjects (3). There is no
controversy over the fact that solving these issues should aid in
improving the effectiveness of alum salts and in the development of
alternative adjuvants, as there is an urgent requirement for
adjuvants capable of boosting Th1-type responses without
unacceptable toxicity.
Carbohydrate structures are renowned for their
ability to boost immune function and have the virtue of maximum
tolerability and safety (9).
Moreover, they are readily biodegradable, with a low risk of
generating toxic metabolites (10,11). A
large number of carbohydrate compounds have emerged as promising
vaccine adjuvant candidates. Among the great range of carbohydrate
structures, polysaccharides stand out as the most promising as they
are less likely to cause adjuvant accumulation and excessive and
chronic immune activation (9).
Polysaccharide-rich fungi and plants have attracted growing
scientific interest for their dietary and medicinal benefits
(12–14), as well as for their ability to exert
marked effects on immune system function, inflammation and cancer
(12,13,15),
Such polysaccharides have also been used as intranasal adjuvants
for the induction of mucosal and systemic immune responses
(16,17). This provides a foundation that may
aid in guiding future research on immune modulation by
well-characterized polysaccharide compounds. However, the
development of vaccines based on polysaccharides must overcome
certain drawbacks and improve the effectiveness of polysaccharides.
For example, children below two years old and the elderly respond
poorly to polysaccharide antigens due to the immaturity or aging of
the immune system (18). Hence,
there is an urgent requirement for the use of polysaccharide
adjuvants, in particular the rational development of new adjuvants
and immunostimulators for vaccines.
Agaricus blazei Murill is a traditional
Chinese fungus that possesses numerous pharmacological properties
(19), including the modulation of
biological homeostasis in order to boost the ability to fight
infection, counteract diseases and prevent cancer (20–22).
Previously, we reported that a polysaccharide from Agaricus
blazei was able to suppress tumor growth and angiogenesis in
vivo(23), inhibit sialyl Lewis
X/E-selectin-mediated metastasis in HT-29 cells (24) and prevent and attenuate hematogenous
metastasis in HT-29 cells (25).
However, to date, no information has been published concerning
Agaricus blazei polysaccharides as potential adjuvants. We
previously identified a new intracellular polysaccharide, ABP-AW1
(Mw, 50 kDa), from Agaricus blazei Murill (26). The structural features of the
purified polysaccharide were successfully characterized by means of
chemical analyses and instrumental spectroscopy. These results
showed that the backbone chains of ABP-AW1 are mainly
(1→6)-linked-β-D-galactopyranose, (1→6)-linked-β-D-glucopyranose
and (1→3,6)-linked-β-D-glucopyranose, terminating with (1→)-linked
Fuc, Ara and Man residues at the O-3 position of
(1→3,6)-linked-β-D-glucopyranose in the proportion of
29:10:10:6:2:2. Hence, in the present study, we hypothesized that
ABP-AW1 is able to act as an adjuvant for the development of
adaptive immunity. To test this hypothesis, the in vivo
adjuvant activity of the ABP-AW1 adjuvant system was evaluated
using ovalbumin (OVA) as a model protein antigen. The aim was to
determine whether ABP-AW1 was able to enhance the cellular
immunity, in addition to the humoral immunity, of mice
subcutaneously immunized with OVA, particularly the Th1-type
responses.
Materials and methods
Mice
Male ICR mice (Grade II, five to six weeks old) were
obtained from Jilin University Animal Research Center (Changchun,
China). The mice were specific pathogen-free and acclimated for one
week prior to use. Rodent laboratory chow and tap water were
provided ad libitum and the mice were exposed to a 12 h/12 h
light/dark cycle at 24±1°C and 50±10% relative humidity. All animal
procedures were performed according to the Guide for the Care and
Use of Laboratory Animals of the National Research Council. This
study was approved by the Animal Care and Use Committee of Qiqihar
Medical University (Qiqihar, China).
Isolation and purification of
polysaccharide
The ABP-AW1 polysaccharide was prepared and
characterized as previously described (26). Briefly, Agaricus blazei
Murill were subjected to three treatments with 95% ethanol (5,000
ml) at 75°C for 6 h under reflux to remove any lipids. The residue
was extracted three times with distilled water (8,000 ml) at 75°C
for 3 h. The supernatant was removed and the precipitate was
washed, dried and extracted twice with 0.5 M NaOH solution,
containing 0.3% (w/w) NaBH4 at room temperature, which
was incubated overnight. Subsequent to filtering to remove the
debris, the suspension was neutralized with hydrochloric acid (0.1
M) and filtered. The supernatant containing water-soluble
polysaccharide was dialyzed, concentrated, ethanol precipitated and
dried. The precipitate was obtained by centrifugation (4,000 × g
for 10 min) and deproteinized by the Sevag method (27), followed by exhaustive dialysis with
distilled water for 48 h. The concentrated dialyzate was then
precipitated at 4°C with 95% ethanol (4,000 ml) for 24 h. The
precipitate was washed with absolute ethanol, acetone and ether,
yielding the aqueous crude polysaccharide (CABP-AW).
The CABP-AW was redissolved with distilled water and
centrifuged at 4,000 × g for 10 min, then the supernatant was
applied to a DEAE Sepharose Fast Flow column (Amersham Biosciences,
Uppsala, Sweden) that had been equilibrated with ultrapure water.
After loading the sample, the column was eluted stepwise with NaCl
aqueous solution (0, 0.2, 0.4 and 0.6 M) at a flow rate of 4
ml/min. The fractions (8 ml) were collected using a Frac-950
(Amersham Biosciences) and the polysaccharide was purified further
by gel-permeation chromatography on a Sepharose 6 Fast Flow column
(2.6×100 cm) with 0.15 M NaCl at a flow rate of 1 ml/min. Three
polysaccharide fractions (ABP-AW1, ABP-AWA1 and ABP-AWB1) were
obtained. The eluted ABP-AW1 was applied to a Sephadex G-25 column
(2.6×40 cm; Amersham Biosciences) to remove any salts.
Subsequently, ABP-AW1 was collected, dialyzed and lyophilized to
obtain the purified polysaccharide for use in the subsequent
experiments.
Immunization protocol
For the immunization, groups of male ICR mice were
subcutaneously immunized with 100 μl 0.9% saline containing 100 μg
OVA (Sigma, St. Louis, MO, USA) alone, 100 μg OVA + 200 μg alum
(Sigma), or 100 μg OVA + ABP-AW1 (50, 100 and 200 μg).
Saline-treated mice were included as a control group. Immunizations
were performed twice with a two-week interval and the mice were
sacrificed two weeks after the secondary immunization. Blood was
collected and centrifuged at 3,000 × g for 10 min to generate serum
samples, which were stored at −80°C, and splenocyte suspensions
were collected under aseptic conditions.
Splenocyte proliferation assay in
vitro
Single-cell suspensions in RPMI-1640 were collected
from the OVA-immunized mice under aseptic conditions. Splenocyte
proliferation was assayed as previously described (28). Briefly, single-cell suspensions of
OVA-immunized mouse spleens were created by grinding the spleens
with a syringe plunger against a fine steel mesh. The erythrocytes
were lysed with ammonium chloride (0.8%, w/v) and centrifuged at
300 × g at 4°C for 10 min. Mononuclear cells were washed three
times in PBS and re-suspended in FCS-RPMI. Splenocytes were plated
in triplicate in 96-well plates (Corning Inc., Corning, NY, USA) at
a concentration of 2×106 cells/ml at 37°C in a
humidified 5% CO2 incubator, together with various
concentrations of OVA (10 μg/ml), concanavalin A (ConA; 5 μg/ml;
Sigma) or lipopolysaccharides (LPS; 2 μg/m; Sigma) for 72 h.
Proliferative responses were assessed after 72 h. In the last 4 h
of each culture, the cultures were pulsed with 20 μl 4 mg/ml MTT
(Sigma) and the absorbance was measured using a microplate reader
at 570 nm with a 630 nm reference. The stimulation index (SI) was
calculated based on the following formula: SI = absorbance value
for mitogen-cultures / absorbance value for non-stimulated
cultures. Statistical significance was determined using the
Student’s t-test.
Measurement of OVA-specific IgG and
subclasses
OVA-specific IgG and subclass levels in the serum
were determined with an indirect enzyme-linked immunosorbent assay
(ELISA) according to the previously described method, with slight
modifications (29). In brief,
round-bottom 96-well high-binding microtiter plates were coated
with 100 μl OVA solution (50 μg/ml in 50 mM carbonate-bicarbonate
buffer; pH 9.6) overnight at 4°C. Three washes were performed using
PBS-0.05% Tween (PBST) solution, followed by blocking with 200 μl
5% FCS/PBS at 37°C for 2 h. Subsequent to three washes with PBST,
100 μl of a series of diluted serum samples or 0.5% FCS/PBS as a
control was added to triplicate wells, followed by a 1-h incubation
at 37°C. After being washed, the bound antibody was detected using
horseradish peroxidase-conjugated goat anti-mouse IgG (Southern
Biotech, Birmingham, AL, USA), IgG1 (Southern Biotech) or IgG2b
(Southern Biotech) and the unbound antibodies were removed by
washing five times with PBST after 1 h at 37°C.
3,3′,5,5′-Tetramethylbenzidine (TMB) liquid substrate was added and
incubated for 15 min at 37°C and the reaction was stopped by adding
2 M H2SO4/well. The optical density was
measured using a microplate reader at 450 nm. Statistical
significance was determined using Student’s t-test.
Cytokine determination in cultured
splenocyte supernatants
Splenocytes were isolated from the immunized mice as
described previously (30). All
cell suspensions were aliquoted into 24-well round-bottom plates at
2×106 cells/well. Samples were incubated at 37°C in 5%
CO2 with OVA (final concentration 10 μg/ml). After 48 h,
the plates were centrifuged at 1,400 × g for 5 min and the presence
of IFN-γ was measured in the cultured supernatants of the
splenocytes using the mouse IFN-γ ELISA kits (R&D Systems,
Wiesbaden, Germany). Statistical significance was determined using
Student’s t-test.
Cell surface and intracellular cytokine
staining
The cell surface and intracellular cytokine staining
was evaluated using flow cytometry (31,32).
Briefly, cell suspensions from immunized mice were harvested from
the spleen as described previously, then splenocytes were aliquoted
into six-well round-bottom plates at 1×106 cells/well
and stimulated with 10 μg/ml OVA in the presence of 5 μM brefeldin
A (10 μg/ml; Sigma) for 24 h. The splenocytes were subsequently
surface stained with 20 μl anti-mouse CD4-FITC Ab (BD Pharmingen,
San Diego, CA, USA) and 20 μl CD69-PE/Cy5.5 Ab (BD Pharmingen)
diluted in FACS buffer (0.5% bovine serum albumin in
phosphate-buffered saline) for 30 min at 4°C. The cells were then
washed with FACS, fixed with 1 ml 4% paraformaldehyde and incubated
for 10 min at 4°C. The cells were washed three times with 1 ml PBS,
then permeabilized with 0.1% saponin for 2–3 min at 4°C. After
centrifugation, intracellular staining was performed with
anti-mouse IFN-γ-Alexa Fluor 647 (20 μl; BD Pharmingen), followed
by incubation for 30 min at 4°C and flow cytometry analysis, which
was performed using the FlowJo software (Tree Star, Ashland, OR,
USA).
Statistical analysis
The data are expressed as the mean ± SD. Student’s
t-tests were conducted to analyze the differences among the means
using SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
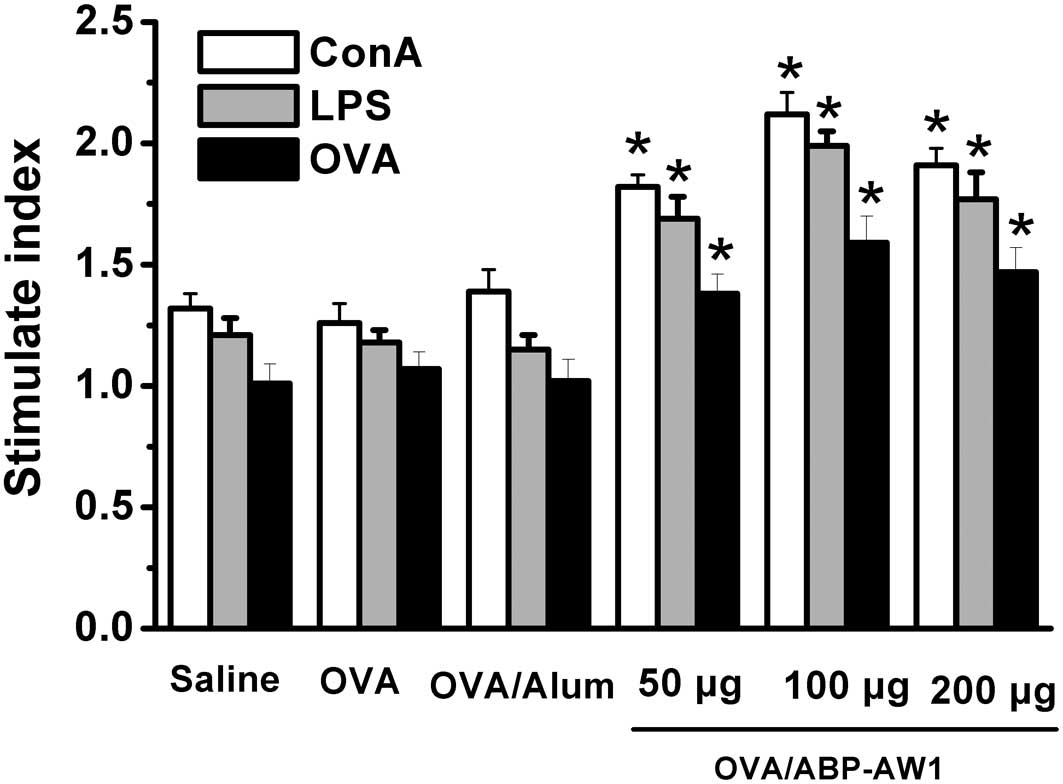
Effect of ABP-AW1 on the proliferation of
splenocytes from OVA-immunized mice
To investigate the effect of ABP-AW1 on
OVA-immunized mice, the proliferation of these cells was analyzed
in the spleens of OVA-immunized mice. It was observed that the mice
immunized with OVA/ABP-AW1 had significant immunological responses
(Fig. 1). ConA, LPS and OVA
stimulated the proliferation of splenic lymphocytes in the mice
immunized with OVA/ABP-AW1, and the response was significantly
larger than in the OVA or OVA/alum groups (P<0.05), particularly
with ABP-AW1 at a dose of 100 μg. However, no significant
differences were detected among the saline, OVA alone and OVA/alum
groups (P>0.05).
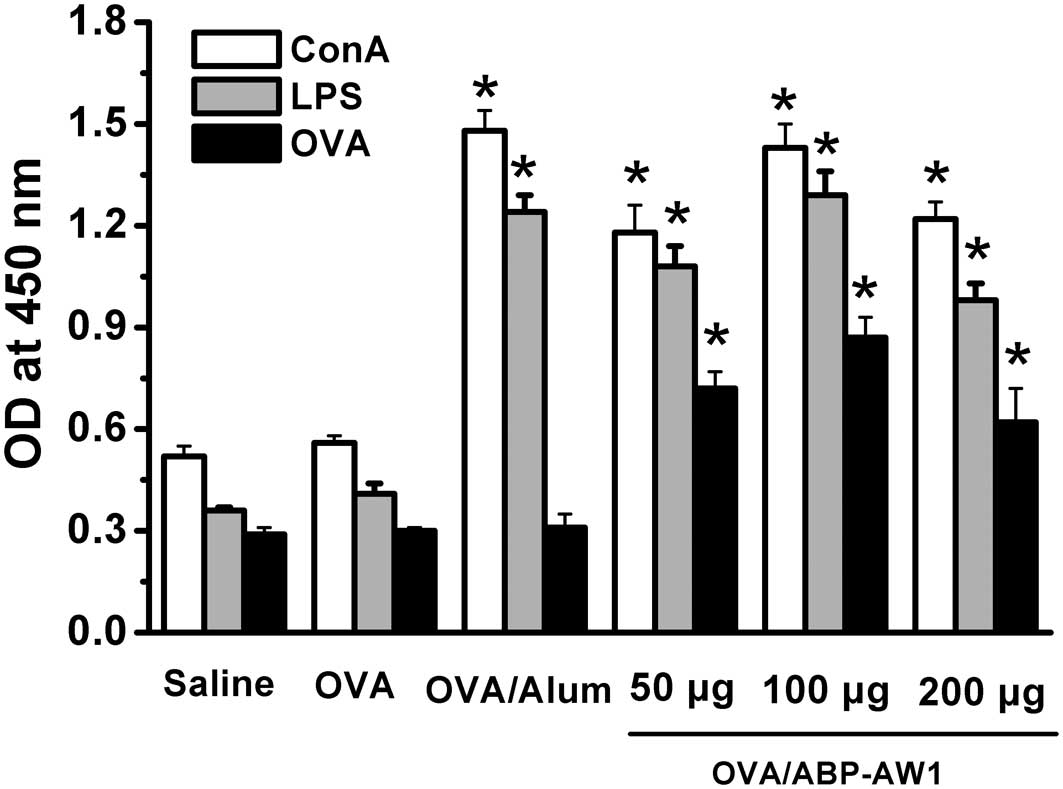
Effect of ABP-AW1 on OVA-specific serum
antibody response
The effects of ABP-AW1 on OVA-specific IgG, IgG1 and
IgG2b antibody production were examined using the methods described
previously. The total levels of IgG antibody (Fig. 2) were elevated significantly in the
OVA/alum and OVA/ABP-AW1 groups compared with the OVA alone group.
The IgG1 subtype is reported to be associated with Th2-dominated
immune responses, whereas IgG2b is considered to be a mediator of
Th1-type immunity (33). Fig. 2 shows that the administration of
OVA/ABP-AW1 markedly augmented the production of the OVA-specific
IgG1 and IgG2b isotypes. Moreover, considerable enhancements were
detected in OVA-specific IgG2b levels in mice immunized with
OVA/ABP-AW1 compared with the OVA group (P<0.05), and
significant differences were observed among the ABP-AW1 groups at
all doses (P<0.05). As a positive control, alum elicited the
highest IgG and IgG1 isotype levels, but did not function at the
IgG2b level. These data indicate that the Th1 and Th2 immune
responses were induced by the OVA/ABP-AW1 vaccine in vivo
and that the ABP-AW1 adjuvant is required to induce Th1-biased IgG
isotype switching.
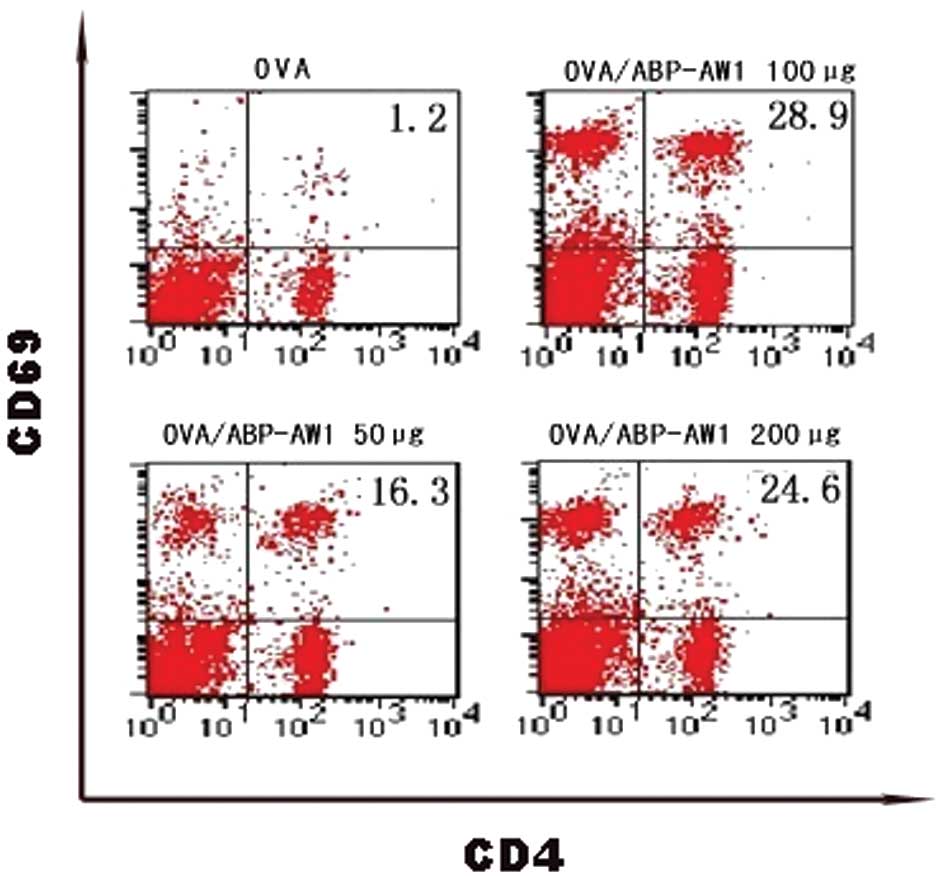
Splenic CD4+ activation
assay
The recall response of T helper cells was assayed
two weeks after the final booster vaccination. The phenotype of the
CD4+ T cells was characterized in the immunized mice in
order to determine whether CD4+ T cells express
activation markers. The early activation marker CD69 was detected
on the splenic CD4+ T cells. It was observed that the
ratio of activated T cells (CD4+CD69+) to
CD4+ T cells in the OVA/ABP-AW1 immunized group was
higher compared with the OVA alone group (Fig. 3), indicating that OVA/ABP-AW1
induced marked antigen-specific induction of splenic
CD4+CD69+ T cells in comparison with OVA
stimulation.
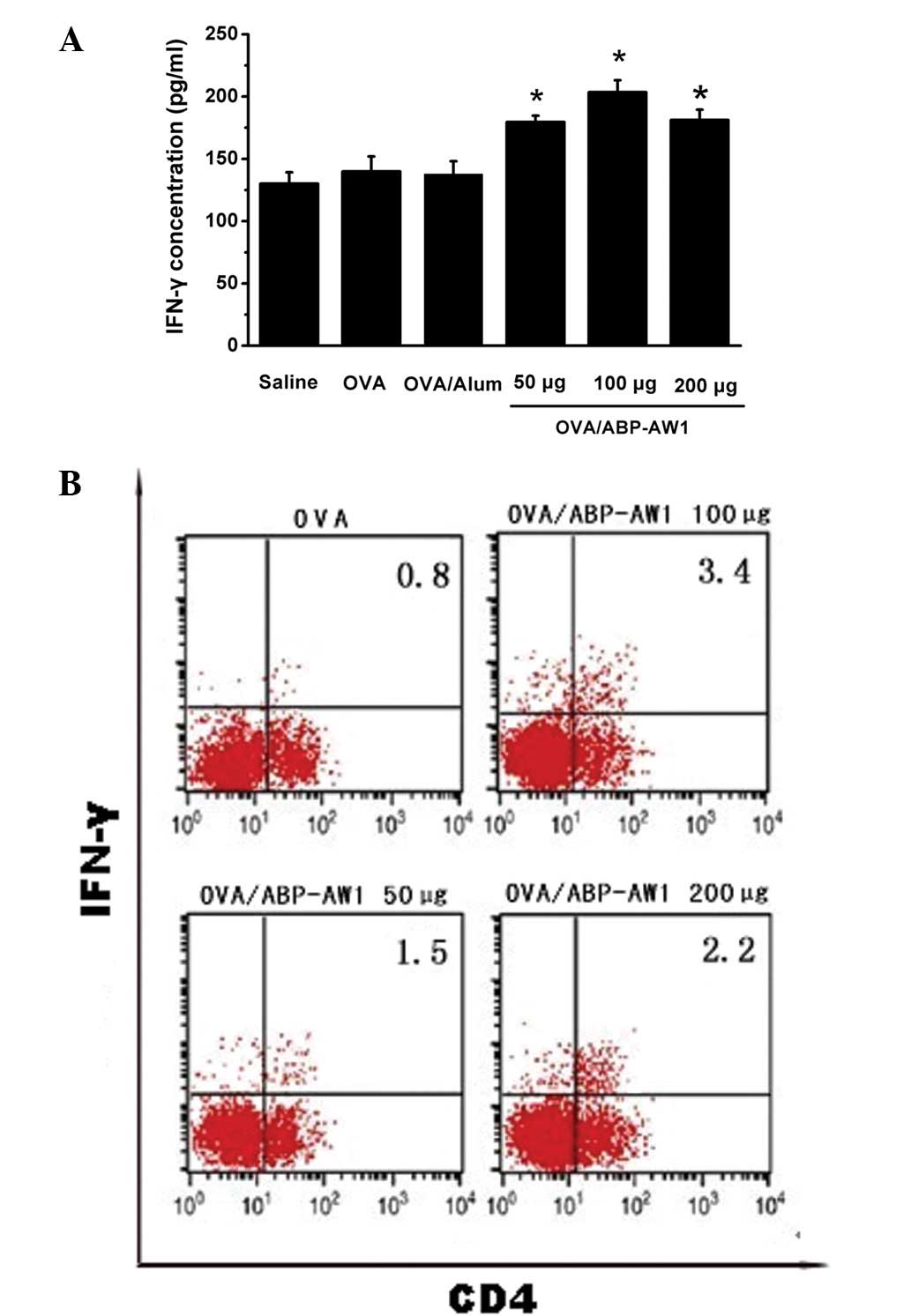
Effect of ABP-AW1 on cytokine levels in
splenocytes from OVA-immunized mice
The previously mentioned findings suggested that
ABP-AW1 has immunomodulation activity as an adjuvant. To
investigate this further, ELISA analysis was performed to determine
the expression patterns of the immune response in the immunized
mice. Compared with the levels of cytokines in the culture
supernatant of splenocytes from OVA-immunized mice, it was observed
that the OVA/ABP-AW1 vaccine had differential cytokine expression
profiles. The culture supernatant of splenocytes from the
OVA/ABP-AW1-immunized mice showed high levels of IFN-γ (Th1 type
immune response; Fig. 4A) in
comparison to the OVA/alum or OVA alone groups. Consistent with the
ELISA results, the intracellular FACS analysis showed that the
CD4+ T cell populations from the OVA/ABP-AW1 group
expressed significantly higher levels of IFN-γ compared with the
OVA control group (Fig. 4B). The
proportion of IFN-γ+CD4+ (double-positive) T
cells from the mice immunized with OVA/ABP-AW1 was markedly
increased. The CD4+ T cells obtained from the mice
immunized with OVA/alum showed negligible IFN-γ production (data
not shown). Thus, it may be suggested that OVA/ABP-AW1 markedly
induces Th1 cells, as determined by IFN-γ production. These results
showed noticeable correlations, indicating that the addition of
ABP-AW1 to OVA enhances Th1 polarization.
Discussion
The present study demonstrated the in vivo
efficacy of ABP-AW1 as an adjuvant for OVA-based vaccines. Ideally,
adjuvants should promote an appropriate immune response, (Th1 or
Th2), be biodegradable and not be immunogenic themselves. Adjuvants
should also offer excellent safety, tolerability, ease of
manufacture and formulation. Therefore, modern adjuvants should
have a profound effect on the nature of the immune response and it
is often desirable to induce specific types of immunity (34), which may bias the immune system
toward either a Th1 or a Th2 type response (35). Among the T lymphocytes, T cell
responses have been divided in two subclasses, Th1 and Th2. Th1 and
Th2 immune response profiles correspond to the activation of two
distinct major subsets of T cells characterized by their pattern of
cytokine production (29,33), i.e. IL-2, TNF-β and IFN-γ vs. IL-4,
IL-5 and IL-10, respectively. Th1 cells protect against
intracellular pathogens, activate phagocytosis, induce IgG2a, IgG2b
and IgG3 antibodies and promote delayed-type hypersensitivity
responses, whereas Th2 cells protect against extracellular
pathogens, activate eosinophils, induce IgE-mediated allergic
responses and promote other humoral responses in which IgG1, IgGE
and IgA predominate (35).
Polarized Th1 and Th2 phenotypes are critical in the course of
immune responses, and efficient Th1 and Th2 immunity-inducing
adjuvants, particularly those for Th1, are highly in demand
(2). Such adjuvants promote good
cell-mediated immunity. The development of such adjuvants would
benefit from increased knowledge of the molecular mechanisms and
factors controlling these responses.
The present study evaluated whether ABP-AW1 was able
to enhance immunity to OVA in mice, providing insight into the
OVA/ABP-AW1 adjuvant system and in vivo evidence. The
results showed that ABP-AW1 was sufficient to enhance the
activation potential of T and B lymphocytes in OVA-immunized mice.
Furthermore, antigen-specific CD4+ helper T cells are
also regarded as an important attribute of ABP-AW1 responses, which
are directly associated with immunostimulation. To the best of our
knowledge, CD69 is a C-type lectin that is expressed on the surface
of all leukocytes during activation, and the engagement of CD69
maintains the high expression of membrane-bound TGF-β1 on T cells
(36). The early activation marker
CD69 was detected on CD4+ T cells, suggesting that their
generation requires T cell activation. Adaptive major
histocompatibility complex (MHC)-II-mediated CD4+ T cell
activation has been considered to be strictly restricted to protein
antigens (37). When presented to T
cells by MHC-II, antigens generally trigger a T cell-dependent
immune response typified by the production of Th1 or Th2 cytokines,
as well as IgG and the induction of immunological memory. However,
the T-lymphocyte-independent nature of a polysaccharide antigen may
be overcome by conjugating polysaccharides to protein carriers
(38,39). Such conjugates have been
demonstrated to be efficient in inducing T-lymphocyte-dependent
immunity, as well as protecting infants and the elderly from
infection (40). The present study
demonstrated that OVA/ABP-AW1 activated T cells. This result was
consistent with the increased numbers of splenic
CD4+CD69+ T cells observed in the
OVA-immunized mice (Fig. 3).
ABP-AW1 induced marked antigen-specific activation of the splenic
CD4+CD69+ T cells in response to OVA
stimulation, indicating that the OVA/ABP-AW1 vaccine was efficient
in inducing T-lymphocyte-dependent immunity.
Evidence is accumulating that ABP-AW1 is a potential
new adjuvant. Cytokines have a central role during Th1 immune
adjustment and IFN-γ is a critical cytokine that coordinates the
immune response through the transcriptional regulation of
immunologically relevant genes, inducing a wide variety of
antigen-presenting cells to express MHC-I and MHC-II molecules,
promoting the efficacy of antigen-specific T cell activation and
macrophage phagocytosis and stimulating B cells to produce
antibodies (41). IFN-γ is involved
in a wide range of infectious diseases and cancer immunotherapy. As
shown in Fig. 4, ABP-AW1 elicited a
marked Th1-polarized immune response to OVA in the immunized mice.
The level of the cytokine, IFN-γ, in the cultured supernatants from
the splenocytes was significantly enhanced by OVA/ABP-AW1 compared
with OVA alone. Th1 polarization was also characterized by
CD4+ T-cell cytokine release profiles, as demonstrated
by the high levels of IFN-γ produced in response to antigen
restimulation. The humoral immune response induced by the
OVA/ABP-AW1 vaccine was also determined in accordance with Th1
polarization. The key finding of the present study was that the
OVA/ABP-AW1 vaccine was able to induce high levels of IgG (total),
IgG1 and IgG2b antibodies (Fig. 2),
with the addition of ABP-AW1 to OVA at a suitable dose being
effective on Th1 and Th2 cells, resulting in a mixed Th1/Th2 immune
response. However, as a positive control group OVA/alum generated
lower levels of IgG2b compared with OVA/ABP-AW1. ABP-AW1 had the
benefit over alum in that it also stimulated cellular immunity as
reflected by Th1 antibody IgG2b isotype induction. Collectively,
this cascade of immune events provided evidence that ABP-AW1
adjuvant has a key role in inducing a marked Th1 immune response to
OVA antigen.
The efficacy of OVA antigen was greatly enhanced by
formulation with ABP-AW1. This may be explained by investigating
the various types of pathways by which ABP-AW1 may stimulate the
immune system. For example, the ABP-AW1 response may be derived
from NK cells stimulating IFN-γ production at the injection site
(13,42), which may result in the increased
activation of dendritic cells (DCs) that are taking up antigen. In
turn, DCs stimulate IFN-γ-producing cells, potentially prolonging
the release from the NK cells and T cells, while IFN-γ further
substantially enhances IgG2b and IgG2a production by effecting the
isotype switching of the B-lymphocytes (33,35).
Another possible pathway is the spread of ABP-AW1 into the
circulation, whereby ABP-AW1 likely acts systemically on the NK or
T cells to stimulate IFN-γ production or the clonal expansion of
the T cells. In addition, a low incidence of adverse events is
critical for the advancement of vaccine adjuvant candidates;
ABP-AW1 is extracted from Agaricus blazei Murill, which is
edible to humans without ill effects (25). While further studies are required to
clarify the relative contributions of these pathways, the present
data showed that ABP-AW1 could be selected and used as an adjuvant
for the modulation of immune responses.
In conclusion, the present study evaluated the
adjuvant activity of ABP-AW1 extracted from Agaricus blazei
Murill. The present data indicate that ABP-AW1 is a candidate
immunity-stimulating adjuvant and that ABP-AW1 particularly
promotes the development of Th1 polarization. Future studies should
aim to evaluate this adjuvant for use in vaccines instead of alum.
Moreover, the mechanism of the effects of ABP-AW1 as an adjuvant
must be elucidated more specifically.
Acknowledgements
The present study was supported by the Natural
Science Foundation of China (grant no. 81173609) and Heilongjiang
Province Foundation for Traditional Chinese Medicine Scientific,
China (grant no. ZHY-ZB12).
References
|
1
|
Kubena KS and McMurray DN: Nutrition and
the immune system: a review of nutrient-nutrient interactions. J Am
Diet Assoc. 96:1156–1164. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Marrack P, McKee AS and Munks MW: Towards
an understanding of the adjuvant action of aluminium. Nat Rev
Immunol. 9:287–293. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Singh M, Chakrapani A and O’Hagan D:
Nanoparticles and microparticles as vaccine-delivery systems.
Expert Rev Vaccines. 6:797–808. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
HogenEsch H: Mechanisms of stimulation of
the immune response by aluminum adjuvants. Vaccine. 20(Suppl 3):
S34–S39. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schirmbeck R, Melber K, Kuhröber A,
Janowicz ZA and Reimann J: Immunization with soluble hepatitis B
virus surface protein elicits murine H-2 class I-restricted CD8+
cytotoxic T lymphocyte responses in vivo. J Immunol. 152:1110–1119.
1994.PubMed/NCBI
|
|
6
|
Aguilar JC and Rodríguez EG: Vaccine
adjuvants revisited. Vaccine. 25:3752–3762. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guy B: The perfect mix: recent progress in
adjuvant research. Nat Rev Microbiol. 5:505–517. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pascual DM, Morales RD, Gil ED, Muñoz LM,
López JE and Casanueva OL: Adjuvants: Present regulatory
challenges. Vaccine. 24(Suppl 2): S88–S89. 2006. View Article : Google Scholar
|
|
9
|
Petrovsky N and Cooper PD:
Carbohydrate-based immune adjuvants. Expert Rev Vaccines.
10:523–537. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rivas E, Gómez-Arnáiz M, Ricoy JR, et al:
Macrophagic myofasciitis in childhood: a controversial entity.
Pediatr Neurol. 33:350–356. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gherardi RK, Coquet M, Cherin P, et al:
Macrophagic myofasciitis lesions assess long-term persistence of
vaccine-derived aluminium hydroxide in muscle. Brain.
124:1821–1831. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ramberg JE, Nelson ED and Sinnott RA:
Immunomodulatory dietary polysaccharides: a systematic review of
the literature. Nutr J. 9:542010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Paulsen BS: Plant polysaccharides with
immunostimulatory activities. Curr Org Chem. 5:939–950. 2001.
View Article : Google Scholar
|
|
14
|
Kusaykin M, Bakunina I, Sova V, et al:
Structure, biological activity, and enzymatic transformation of
fucoidans from the brown seaweeds. Biotechnol J. 3:904–915. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paulsen BS: Biologically active
polysaccharides as possible lead compounds. Phytochem Rev.
1:379–387. 2002. View Article : Google Scholar
|
|
16
|
Bacon A, Makin J, Sizer PJ, et al:
Carbohydrate biopolymers enhance antibody responses to mucosally
delivered vaccine antigens. Infect Immun. 68:5764–5770. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stambas J, Pietersz G, McKenzie I and
Cheers C: Oxidised mannan as a novel adjuvant inducing mucosal IgA
production. Vaccine. 20:1068–1078. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mond JJ, Vos Q, Lees A and Snapper CM: T
cell independent antigens. Curr Opin Immunol. 7:349–354. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hetland G, Johnson E, Lyberg T,
Bernardshaw S, Tryggestad AM and Grinde B: Effects of the medicinal
mushroom Agaricus blazei Murill on immunity, infection and
cancer. Scand J Immunol. 68:363–370. 2008.
|
|
20
|
Endo M, Beppu H, Akiyama H, et al:
Agaritine purified from Agaricus blazei Murill exerts
anti-tumor activity against leukemic cells. Biochim Biophy Acta.
1800:669–673. 2010.
|
|
21
|
Takaku T, Kimura Y and Okuda H: Isolation
of an antitumor compound from Agaricus blazei Murill and its
mechanism of action. J Nutr. 131:1409–1413. 2001.PubMed/NCBI
|
|
22
|
Oh TW, Kim YA, Jang WJ, Byeon JI, Ryu CH,
Kim JO and Ha YL: Semipurified fractions from the submerged-culture
broth of Agaricus blazei Murill reduce blood glucose levels
in streptozotocin-induced diabetic rats. J Agr Food Chem.
58:4113–4119. 2010.PubMed/NCBI
|
|
23
|
Niu YC, Liu JC, Zhao XM and Cao J: A low
molecular weight polysaccharide isolated from Agaricus
blazei Murill (LMPAB) exhibits its anti-metastatic effect by
down-regulating metalloproteinase-9 and up-regulating Nm23-H1. Am J
Chin Med. 37:909–921. 2009.PubMed/NCBI
|
|
24
|
Liu JC, Yue LL, Zhang C, et al: A
polysaccharide isolated from Agaricus blazei Murill inhibits
sialyl Lewis X/E-selectin-mediated metastatic potential in HT-29
cells through downregulating α-1,3-fucosyltransferase-VII
(FucT-VII). Carbohydr Polym. 79:921–926. 2010.
|
|
25
|
Yue LL, Cui HX, Li CC, Lin Y, Niu YC, Sun
YX, Niu YC, Wen XC and Liu JC: A polysaccharide from Agaricus
blazei attenuates tumor cell adhesion via inhibiting E-selectin
expression. Carbohydr Polym. 88:1326–1333. 2012.
|
|
26
|
Liu JC and Sun YX: Structural analysis of
an alkali-extractable and water-soluble polysaccharide (ABP-AW1)
from the fruiting bodies of Agaricus blazei Murill.
Carbohydr Polym. 86:429–432. 2011. View Article : Google Scholar
|
|
27
|
Sun Y, Wang S, Li T, Li X, Jiao L and
Zhang L: Purification, structure and immunobiological activity of a
new water-soluble polysaccharide from the mycelium of Polyporus
albicans (Imaz.) Teng. Bioresour Technol. 99:900–904. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Concha C, Hu S and Holmberg O: The
proliferative responses of cow stripping milk and blood lymphocytes
to pokeweed mitogen and ginseng in vitro. Vet Res. 27:107–115.
1996.PubMed/NCBI
|
|
29
|
Sjölander A, van’t Land B and Lövgren
Bengtsson K: Iscoms containing purified Quillaja saponins
upregulate both Th1-like and Th2-like immune responses. Cell
Immunol. 177:69–76. 1997.
|
|
30
|
Estrada A, Katselis GS, Laarveld B and
Barl B: Isolation and evaluation of immunological adjuvant
activities of saponins from Polygala senega L. Comp Immunol
Microbiol Infect Dis. 23:27–43. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hartmann G, Marschner A, Viveros PR, et
al: CpG oligonucleotides induce strong humoral but only weak CD4+ T
cell responses to protein antigens in rhesus macaques in vivo.
Vaccine. 23:3310–3317. 2005.PubMed/NCBI
|
|
32
|
Sobol PT, Boudreau JE, Stephenson K, Wan
Y, Lichty BD and Mossman KL: Adaptive antiviral immunity is a
determinant of the therapeutic success of oncolytic virotherapy.
Mol Ther. 19:335–344. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kawase O, Goo YK, Jujo H, Nishikawa Y and
Xuan X: Starfish, Asterias amurensis and Asterina
pectinifera, as potential sources of Th1 immunity-stimulating
adjuvants. J Vet Med Sci. 73:227–229. 2011.
|
|
34
|
Pulendran B and Ahmed R: Translating
innate immunity into immunological memory: implications for vaccine
development. Cell. 124:849–863. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun Y, Liu J, Yu H and Gong C: Isolation
and evaluation of immunological adjuvant activities of saponins
from the roots of Pulsatilla chinensis with less adverse
reactions. Int Immunopharmacol. 10:584–590. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han Y, Guo Q, Zhang M, Chen Z and Cao X:
CD69+ CD4+ CD25- T cells, a new subset of regulatory T cells,
suppress T cell proliferation through membrane-bound TGF-beta 1. J
Immunol. 182:111–120. 2009.
|
|
37
|
Watts C and Powis S: Pathways of antigen
processing and presentation. Rev Immunogenet. 1:60–74.
1999.PubMed/NCBI
|
|
38
|
Insel RA and Anderson PW:
Oligosaccharide-protein conjugate vaccines induce and prime for
oligoclonal IgG antibody responses to the Haemophilus
influenzae b capsular polysaccharide in human infants. J Exp
Med. 163:262–269. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shelly MA, Jacoby H, Riley GJ, Graves BT,
Pichichero M and Treanor JJ: Comparison of pneumococcal
polysaccharide and CRM197 conjugated pneumococcal oligosaccharide
vaccines in young and elderly adults. Infect Immun. 65:242–247.
1997.PubMed/NCBI
|
|
40
|
Kasper DL, Paoletti LC, Wessels MR,
Guttormsen HK, Carey VJ, Jennings HJ and Baker CJ: Immune response
to type III group B streptococcal polysaccharide-tetanus toxoid
conjugate vaccine. J Clin Invest. 98:2308–2314. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shiomi H, Masuda A, Nishiumi S, et al:
Gamma interferon produced by antigen-specific CD4+ T
cells regulates the mucosal immune responses to Citrobacter
rodentium infection. Infect Immun. 78:2653–2666. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Meng C, Peng X, Shi X, Wang H and Guo Y:
Effects of a chemically derived homo zwitterionic polysaccharide on
immune activation in mice. Acta Biochim Biophys Sin (Shanghai).
41:737–744. 2009. View Article : Google Scholar : PubMed/NCBI
|