Introduction
Acute promyelocytic leukemia (APL) is a distinct
subtype of acute myeloid leukemia (AML). Cytogenetically, APL is
marked by a balanced reciprocal translocation between chromosomes
15 and 17, which results in the fusion of the promyelocytic
leukemia (PML) and retinoic acid receptor (RAR)α genes (1). The current primary treatment for APL
is anticancer drug-based chemotherapy. With the use of all-trans
retinoid acid (ATRA) as a differentiation induction therapy and
arsenic trioxide (ATO) as a target therapy, the combination of ATRA
and ATO, as well as bone marrow transplantation, has shown
significant progress for the treatment of APL. However, the
long-term use of ATRA often leads to drug resistance of the cancer
cells, causing future treatments on relapsed patients to be
ineffective. ATO has severe side-effects, particularly in patients
with potential heart disease (2).
Therefore, the development of more effective and safe natural
antineoplastic agents is of great interest for potential practical
uses, such as cancer chemotherapy, and for understanding the
mechanisms of tumor development.
Naturally occurring triterpenoids are synthesized
endogenously in various types of plants by the cyclization of
squalene and have been used in traditional medicine in numerous
Asian countries for centuries (3).
Oleanolic acid (OA), a pentacyclic triterpenoid, is the major
component of various plants and a number of medical herbs widely
distributed throughout the world (4,5). OA
exhibits broad biological properties, including protection against
hepatoxicity and nephrotoxicity (6,7),
anti-inflammatory effects (8,9), the
recovery of the hematopoietic system after irradiation (10) and cytotoxicity against several
cancer cell lines (10–12). In a previous study, we showed that
OA induces apoptosis in HL-60 cells through caspase activation and
poly(ADP-ribose) polymerase cleavage (13). The present study was performed to
examine the cytotoxic effects of OA on NB4 cells expressing the
PML/RARα fusion gene and protein.
Materials and methods
Cell line and cell culture
The human leukemia NB4 cell line was purchased from
Shanghai Bioleaf Biotech Co., Ltd. (Shanghai, China). The cells
were cultured in 90% RPMI-1640 and 10% heat-inactivated fetal
bovine serum (Gibco BRL, Gaithersburg, MD, USA), supplemented with
100 IU/ml penicillin and 100 μg/ml streptomycin in a 37ºC,
humidified incubator with 5% CO2.
Cell viability assay
Following treatment with 60, 80 and 100 μmol/l OA
and RPMI-1640 medium (negative control) for 24, 48 and 72 h, NB4
cell viability was determined by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay, as described previously (13).
Relative quantitative PCR
The expression levels of bcl-2 and bax mRNA were
determined by relative quantitative PCR. Total RNA from the NB4
cells was extracted using an RNeasy Mini kit (Qiagen, Hilden,
Germany) following treatment with 80 μmol/l OA for 72 h. RNA
concentrations were determined using a NanoDrop (Thermo Scientific,
Rockford, IL, USA) and 1 μg RNA was reverse transcribed with a
PrimeScript RT reagent kit (Takara, Biotechnology, Co., Ltd.,
Dalian, China). Relative quantification of gene expression was
performed in triplicate. The mRNA expression was determined using
SYBR Premix Ex Taq™ (Takara, Biotechnology, Co., Ltd.). The primers
sequences were synthesized by Takara (Takara, Biotechnology, Co.,
Ltd.) as follows: Bcl-2 forward, 5′-TGA ACC GGC ATC TGC ACA C-3′
and reverse, 5′-CGT CTT CAG AGA CAG CCA GGA G-3′; bax forward,
5′-AGA CAC CTG AGC TGA CCT TGG AG-3′ and reverse, 5′-GTT GAA GTT
GCC ATC AGC AAA CA-3′; and β-actin forward, 5′-AAG AGA GGC ATC CTG
ACC CT-3′ and reverse, 5′-TAC ATG GCT GGG GTG TTG AA-3′. Gene
expression levels were quantified using 7300 Fast Real Time
Sequence detection system software (Applied Biosystems, Foster
City, CA, USA). Relative expression was calculated using the
comparative Ct method.
DNA fragment analysis
The cells (1.5×106) were treated with 80
μmol/l OA for 24, 48 and 72 h, then collected and washed twice with
PBS. DNA was extracted using the Genomic DNA Mini Preparation kit
with Spin Column (Beyotime Institute of Biotechnology, Haimen,
China). DNA (~15 μg) was loaded onto a 1.5% agarose gel. Subsequent
to electrophoresis, the gel was visualized using ethidium bromide
staining under ultraviolet light.
Cell cycle distribution
The NB4 cells were collected and washed twice with
cold PBS following treatment with 80 μmol/l OA for 24, 48 and 72 h,
then cell pellets were suspended in 200 μl propidium iodide (PI)
solution, containing 10 μg/ml PI, 0.1% (w/v) sodium citrate and
0.1% RNase. Cell samples were incubated at 4ºC in darkness for at
least 30 min, then analyzed with a flow cytometer (FACSCalibur;
Becton Dickinson, Sunnyvale, CA, USA) and CellQuest software
(Becton Dickinson).
Activity analysis of caspase-9 and
caspase-3
The activity levels of caspase-9 and caspase-3 were
measured using the Caspase Activity kit (Beyotime Institute of
Biotechnology). To evaluate the activity levels of caspase-9 and
caspase-3, cell lysates were prepared following treatment with 80
μmol/l OA for 24, 48 and 72 h. Assays were performed on 96-well
microtitre plates by incubating 10 μl cell lysate protein per
sample in 80 μl reaction buffer [1% NP-40, 20 mmol/l Tris-HCL (pH
7.5), 137 mmol/l NaCl and 10% glycerol], containing 10 μl caspase-9
and caspase-3 substrates, respectively (2 mmol/l Ac-DEVD-pNA). The
lysates were incubated at 37ºC for 4 h. The samples were measured
with an ELISA reader (Bio-Tek, Winooski, VT, USA) at an absorbance
of 405 nm. The detailed analysis procedure was performed according
to the manufacturer’s instructions. All the experiments were
performed in triplicate.
Analysis of PML/RARα fusion protein by
western blotting
Subsequent to being treated with 80 μmol/l OA for
24, 48 and 72 h, the NB4 cells were lysed in ice-cold
radioimmunoprecipitation (RIPA) buffer with protease inhibitors.
The protein concentration was determined using the Bradford method
with a Bio-Rad protein assay reagent (Bio-Rad, San Diego, CA, USA).
Proteins (50 μg) were subjected to sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to a nitrocellulose membrane (GE Healthcare, Arlington
Heights, IL, USA). The resulting blots were blocked with 5% skimmed
dry milk in TBST [50 mmol/l Tris-HCl (pH 7.6), 150 mmol/l NaCl,
0.1% Tween-20] for 1.5 h at room temperature. The TBST buffer
solution was used to wash the membrane three times prior to
immunoblotting using antibodies specific for polyclonal rabbit
anti-human PML/RARα (Abcam Inc, Cambridge, MA, USA) and β-actin
(Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA). Following
incubation at 4ºC overnight and a wash with TBST, the membranes
were incubated with horseradish peroxidase-labeled goat anti-rabbit
IgG (Santa Cruz Biotechnology Inc.) for 1.5 h at room temperature.
The membranes were washed with TBST and visualized using
electrochemiluminescence western blotting detection reagents (GE
Healthcare).
Statistical analysis
The statistical analysis was performed using SPSS
13.0 software (SPSS, Inc., Chicago, IL, USA). Data are expressed as
the mean ± SD. Differences were analyzed by ANOVA. P<0.05 was
considered to indicate a statistically significant difference.
Results
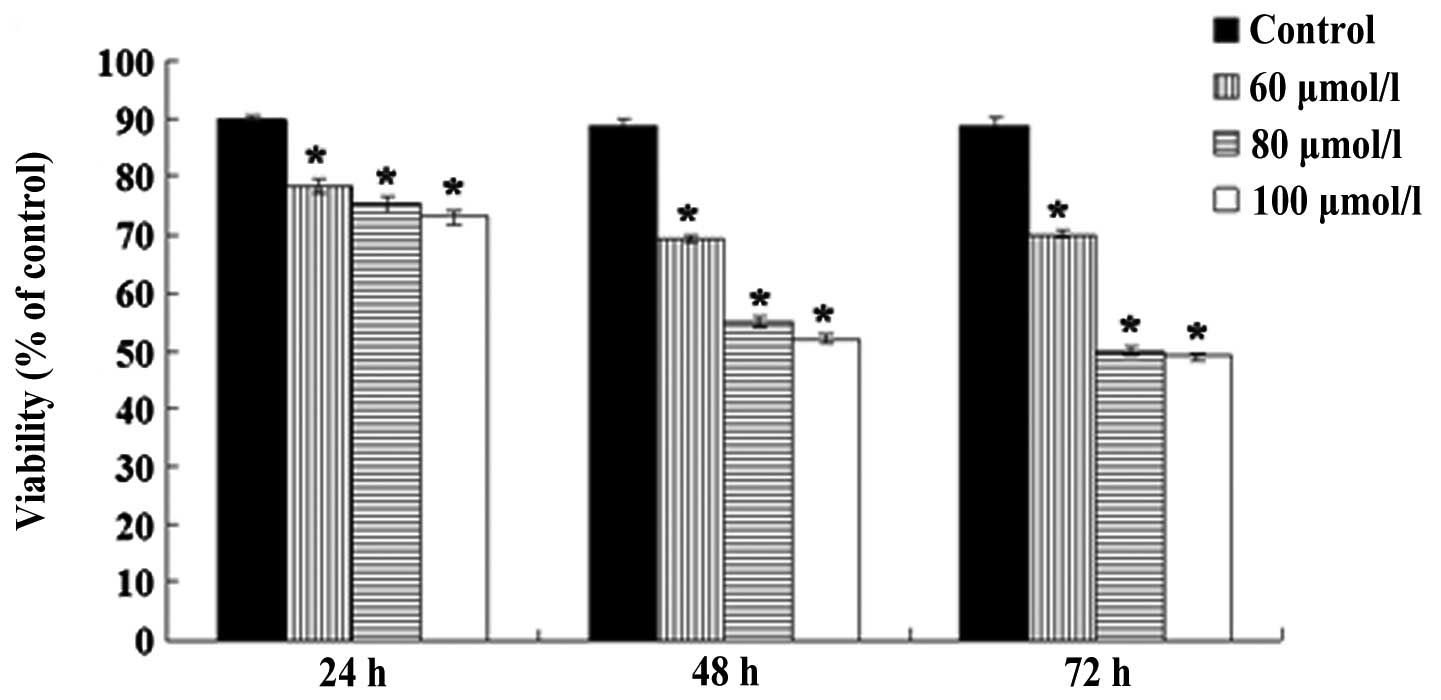
OA inhibits NB4 cell proliferation
To investigate the effect of OA on NB4 cell
proliferation, cell viability was evaluated via the MTT assay. As
shown in Fig. 1, after the cells
had been exposed to 60, 80 and 100 μmol/l OA for 24 h, the
viabilities of the NB4 cells were recorded as 78.5±1.18, 75.2±1.48
and 73±1.06%, respectively, with differences that were significant
compared with the control cells (all P<0.05). After the cells
were exposed to the same concentrations of OA for 48 h, the
viabilities of the NB4 cells were 69.3±0.66, 55±0.85 and 52±0.89%,
respectively, and the differences were significant compared with
the control cells (all P<0.05). Following treatment with the
various concentrations of OA for 72 h, the viabilities of the NB4
cells were 70.2±0.44, 49.9±0.85 and 49±0.5%, respectively, and the
differences were again significant compared with the control cells
(all P<0.05). There were no differences between the effect of 80
and 100 μmol/l OA on the viability of the NB4 cells at the various
time points (P>0.05).
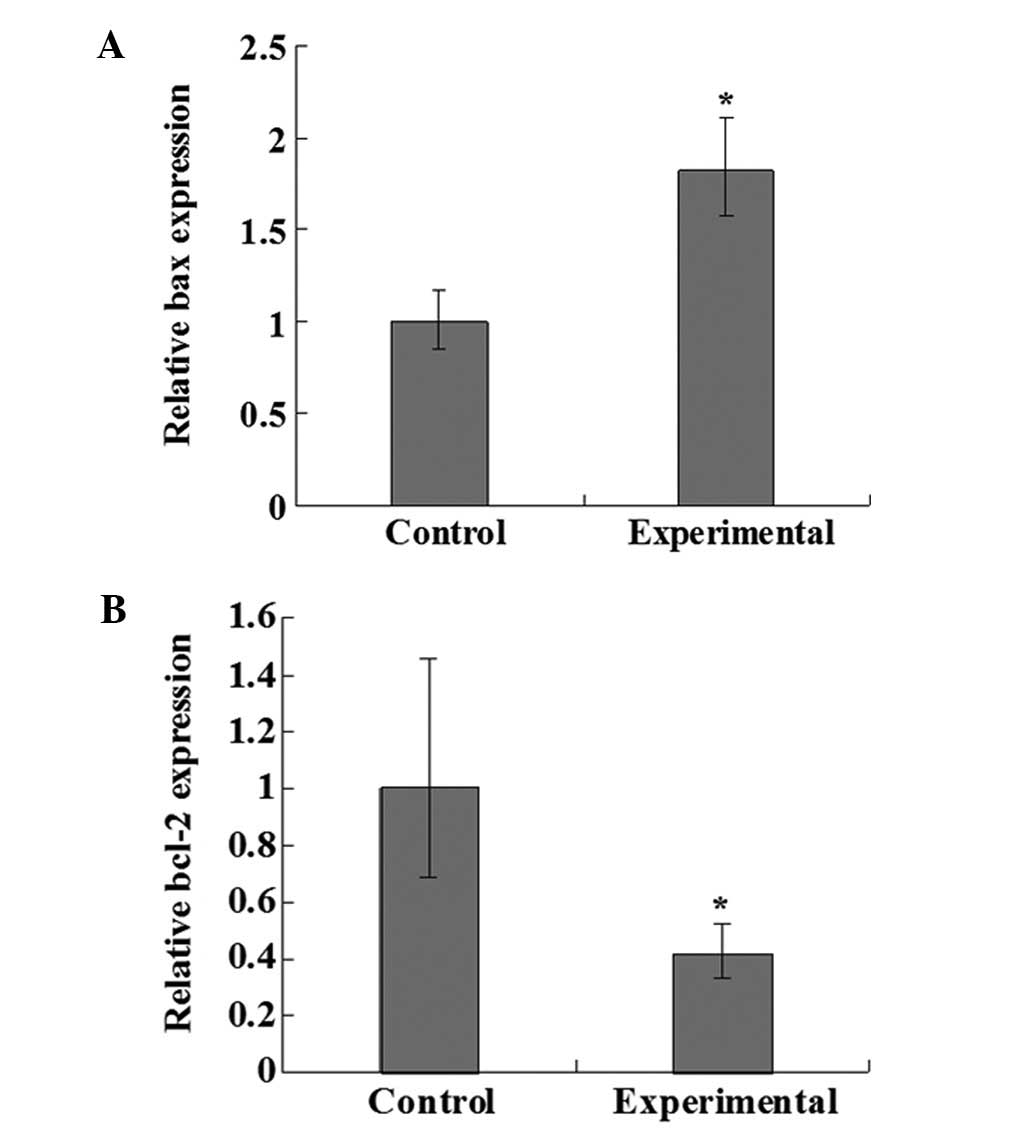
OA upregulates bax and downregulates
bcl-2 mRNA expression in NB4 cells
The relative expression levels of bax and bcl-2 mRNA
in the NB4 cells were tested following treatment with 80 μmol/l OA
for 72 h. As shown in Fig. 2, the
mRNA expression level of pro-apoptotic bax was increased by 81%
(Fig. 2A) compared with the control
group, and the difference was significant (P<0.05). The mRNA
expression level of anti-apoptotic bcl-2 was decreased by 59%
(Fig. 2B) compared with the control
group, and the difference was also significant (P<0.05).
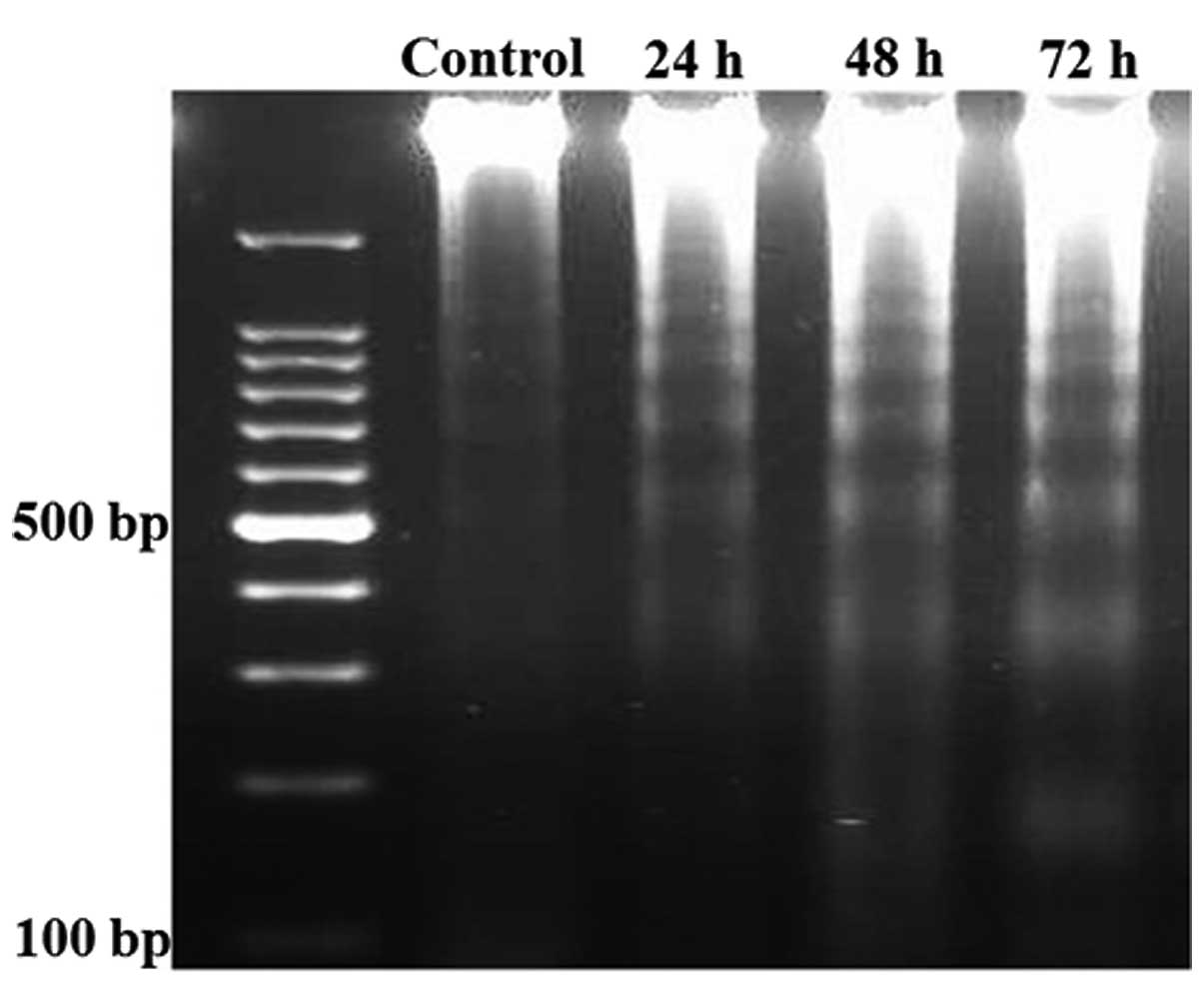
OA induces apoptosis in NB4 cells
Since the presence of the genomic DNA ladder has
been used extensively as a marker for apoptotic cell death,
apoptosis was tested by the DNA ladder formation assay. As shown in
Fig. 3, after the NB4 cells were
treated with OA for 24, 48 and 72 h, DNA laddering was
observed.
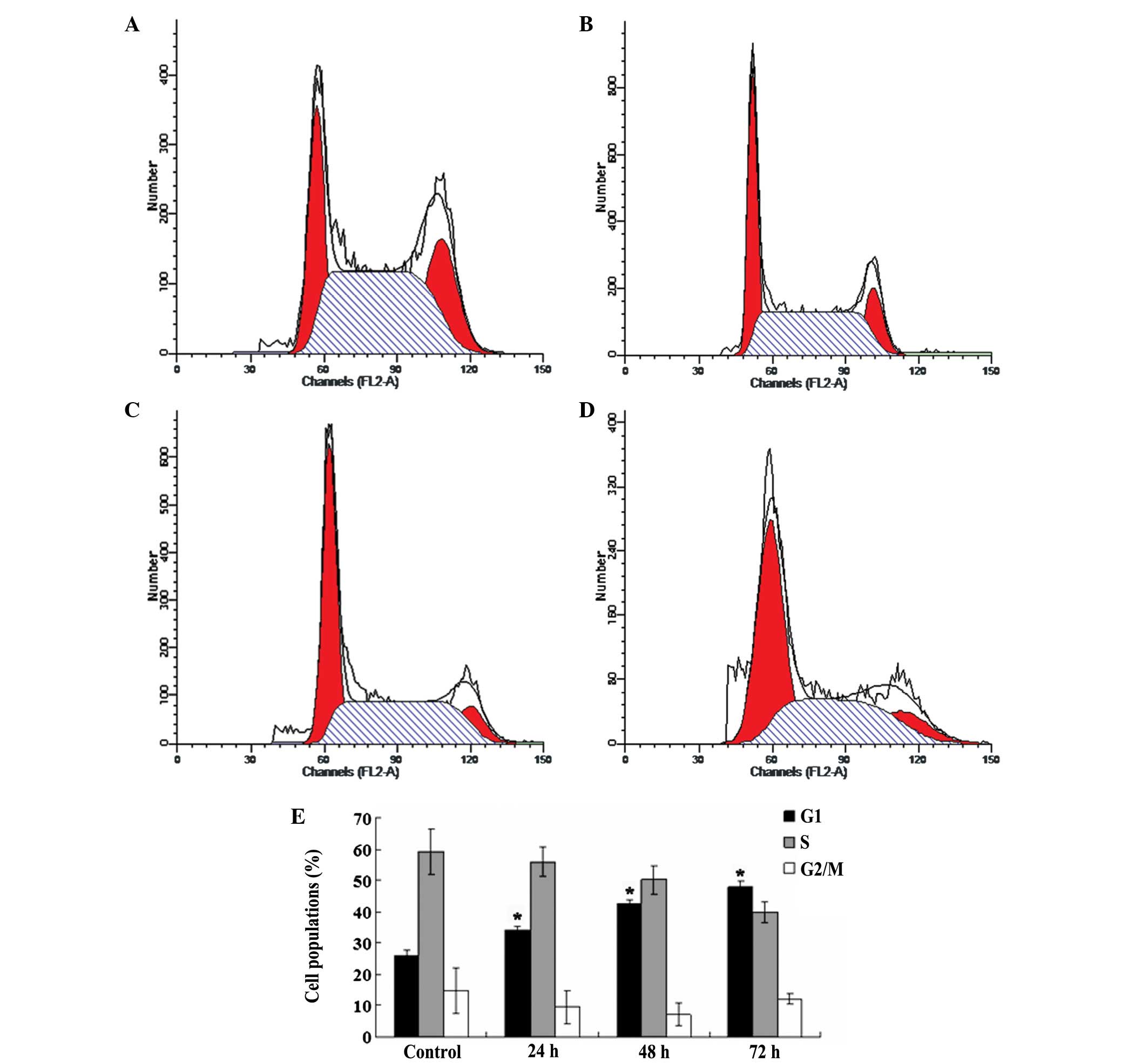
OA induces G1 phase arrest in
NB4 cells
To show in more detail that the inhibition of cell
growth by OA is closely associated with cell cycle control and
apoptosis, the cell cycle distributions of the OA treated tumor
cells and the control cells were analyzed with a flow cytometer
(Fig. 4A–D). As shown in Fig. 4E, compared with the control cells,
following the treatment of the NB4 cells with 80 μmol/l OA for 24,
48 and 72 h, the G1 subpopulation of the NB4 cells was
increased from 25.88±1.7% to 34.24±1.33, 42.51±1.38 and
47.94±1.66%, respectively, and the differences were significant
(all P<0.05).
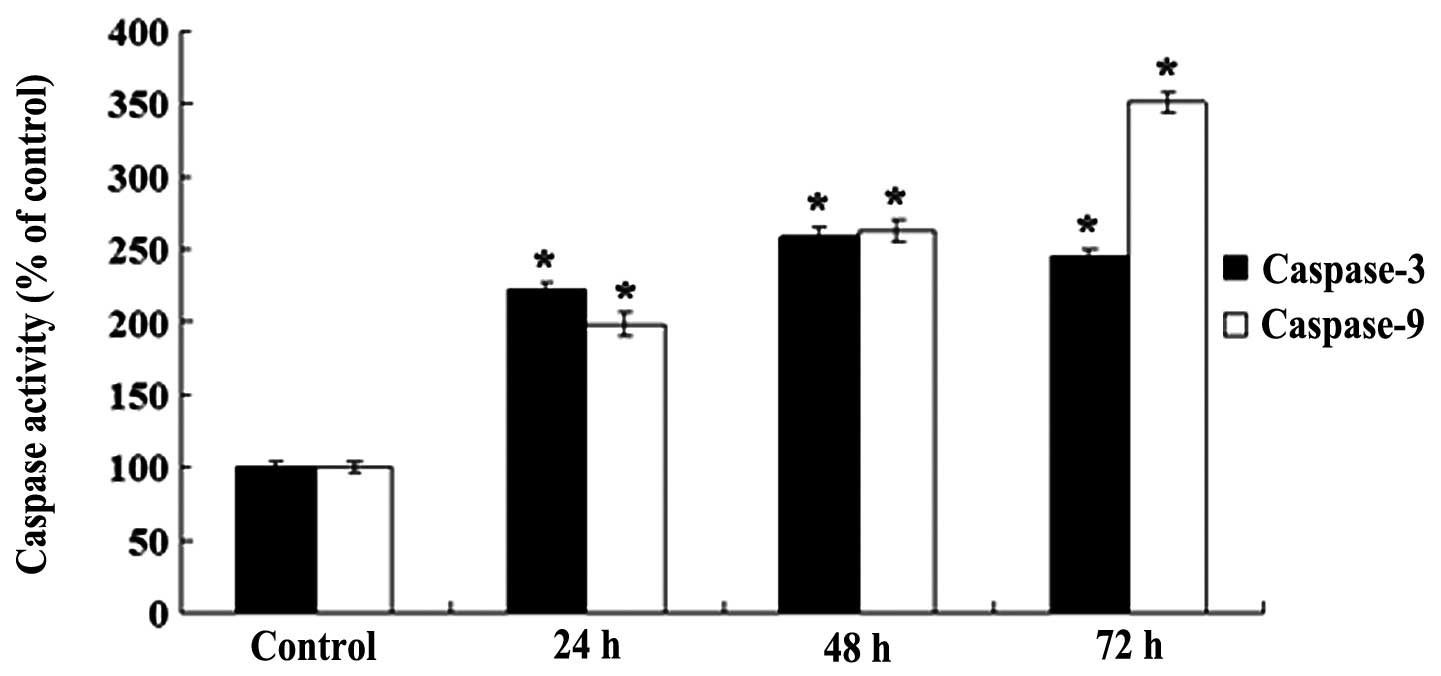
OA increases the activity levels of
caspase-3 and caspase-9
To investigate the effect of OA on caspase-3 and
caspase-9, the activities of caspase-3 and caspase-9 were evaluated
by colorimetric assays. As shown in Fig. 5, subsequent to treating the NB4
cells with 80 μmol/l OA for 24, 48 and 72 h, the activity of
caspase-3 was increased by 2.2-, 2.6- and 2.5-fold, respectively,
compared with the control cells. The activity of caspase-9 was
increased by 2.0-, 2.6- and 3.5-fold compared with the control
cells (Fig. 5). The differences
were significant (all P<0.05).
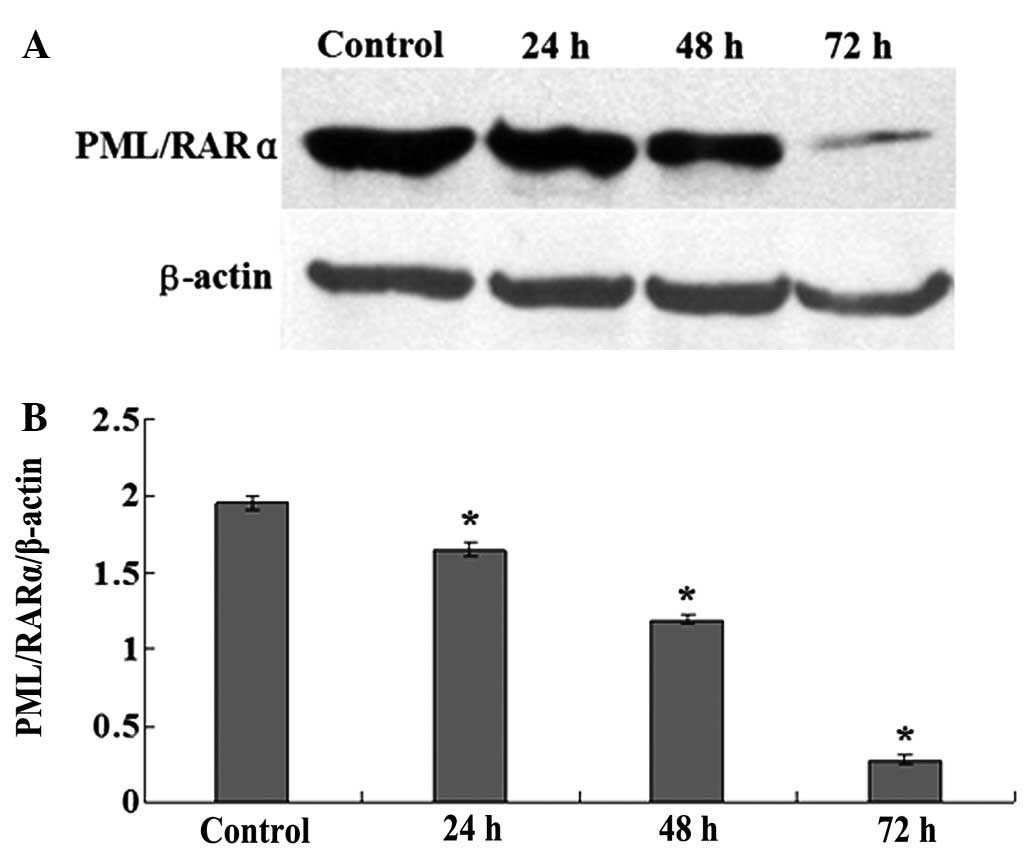
OA downregulates the expression of
PML/RARα fusion protein
To investigate the effect of OA on the expression of
the PML/RARα fusion protein, the expression of PML/RARα was
detected by western blotting (Fig.
6A). As shown in Fig. 6B,
following the treatment of the NB4 cells with 80 μmol/l OA for 24,
48 and 72 h, the expression levels of the PML/RARα fusion protein
were decreased by 16, 39 and 86%, respectively, compared with
control cells. The differences were significant (all
P<0.05).
Discussion
The suppression of apoptosis may contribute to tumor
development by means of the accumulation of continuously
proliferating cells, and the disruption or elimination of
genetically altered cells may decrease the tumor potential
(14–16). It is well known that one of the
biological properties possessed by leukemia is the deregulation of
apoptosis (1). Consequently, the
induction of apoptosis in leukemia cells may be an effective
therapy. The present study showed that OA inhibited the
proliferation of NB4 cells (Fig. 1)
and induced NB4 cell apoptosis (Fig.
3) and G1 phase arrest (Fig. 4).
As a naturally occurring triterpenoid, OA has been
identified in >120 plants (17),
where it exerts cytotoxic effects through multiple mechanisms. OA
has been reported to inhibit the proliferation of human colon
carcinoma HCT15 cells through G0/G1 phase
arrest (12). Additionally, OA has
been shown to induce apoptosis in the non-small cell lung cancer
cell lines A549 and H460 by inhibiting the activity of multidrug
resistance-associated protein 1 (MRP1) (18). Furthermore, OA has been shown to
inhibit the growth and induce the apoptosis of K562, an
erythroleukemia cell line. Notably, OA also inhibited the
proliferation of Lucena 1, a vincristine-resistant derivative of
K562 that possesses several multidrug resistance (MDR)
characteristics (19). OA has
effectively inhibited the tumor promotion induced by
12-O-tetradecanoylphorbol-13-acetate (TPA) in mouse skin and the
activity levels were shown to be comparable to retinoic acid (RA),
a known inhibitor of tumor promotion (20). Furthermore, OA also has cytotoxicity
against SK-OV-3 (ovary), SK-MEL-2 (melanoma) and XF498 (central
nerve system) cells in vitro(11).
It is well established that apoptosis may occur by
either the death-receptor or mitochondrial pathways. The two
pathways are executed by cysteine proteases (caspases) that are
activated specifically in apoptotic cells (14,15).
The death-receptor pathway is triggered by members of the
death-receptor superfamily and involves caspase-8 activation
(14). The mitochondrial pathway is
mobilized in response to extracellular cues and internal insults,
such as DNA damage, often through the activation of a pro-apoptotic
member of the Bcl-2 family, such as Bax or Bid, followed by the
alteration of mitochondrial membrane permeability and the release
of mitochondrial cytochrome c into the cytosol. Cytochrome c
associates with Apaf-1 then procaspase-9 to form the apoptosome.
The death-receptor and mitochondrial pathways converge at the level
of caspase-3 activation, which is followed by caspase-8 activation
or apoptosome formation (14). With
the purpose of identifying the apoptotic pathway that causes the
OA-induced apoptosis of NB4 cells, the mRNA expression levels of
bax and bcl-2 were evaluated in the present study, as well as the
activity levels of caspase-9 and caspase-3. The results indicated
that OA upregulated the expression level of the pro-apoptotic bax
gene, while downregulating the expression level of the
anti-apoptotic bcl-2 gene in the NB4 cells (Fig. 2). Furthermore, OA increased the
activity of caspase-9 and caspase-3 (Fig. 5). These results suggest that OA
induces apoptosis in NB4 cells via the mitochondria-dependent
pathway. However, other mechanisms, such as the death-receptor
pathway, oxidative damage or direct killing by T cells, may not be
excluded, as OA has been shown to stimulate nitric oxide (NO) and
tumor necrosis factor-α (TNFα) release and also to upregulate
inducible nitric oxide synthase (iNOS) and TNFα gene expression
through NF-κB transactivation (21).
In addition to anti-tumor promotion and the
induction of tumor cell apoptosis, the induction of tumor cell
differentiation is also a significant mechanism through which OA
elicits its biological effects. In cultured F9 teratocarcinoma stem
cells, OA was shown to induce differentiation through the
regulation of differentiation-specific genes, including laminin B1,
type IV collagen and RARβ (22). OA
caused the morphological changes of F9 cells into endoderm cells,
as did RA. Furthermore, OA has been demonstrated to possess
antimetastatic activity in vivo(18), as well as anti-angiogenesis activity
(23). All these studies show that
OA affects various stages of tumor development.
As mentioned previously, APL has a unique and
specific chromosomic aberration, t(15;17), which results in the
formation of a fusion gene and protein, PML/RARα, which is not only
necessary for the diagnosis of APL, but is also critical for APL
pathogenesis (1). Studies in
transgenic mice have demonstrated that the PML/RARα fusion protein
blocks granulocytic differentiation, resulting in the accumulation
of abnormal promyelocytes within the bone marrow (24). A common pharmacological activity
shared by ATRA and ATO is the modulation and/or degradation of the
PML/RARα fusion protein (1). The
present study showed that OA significantly decreased the expression
of the PML/RARα fusion protein in the NB4 cells (Fig. 6), suggesting that OA may inhibit
proliferation and induce apoptosis in NB4 cells through the
downregulation of the PML/RARα fusion protein. Notably, the most
effective concentrations of OA on HL-60 and NB4 cells are
different. In our previous study, 4.57×10−2 mg/ml
(equivalent 100 μmol/l) OA had the greatest inhibitory effect on
HL-60 cell proliferation in vitro(13). However, in the present study, the
most effective concentration of OA on the NB4 cells was 80 μmol/l
(equivalent 3.65×10−2 mg/ml). This difference may result
from the presence or absence of the PML/RARα fusion protein, since
the HL-60 cell line lacks the PML/RARα fusion gene (25); this may be the main reason why NB4
cells are more sensitive to OA than HL-60 cells.
In conclusion, the present data suggest that OA
inhibits proliferation and induces apoptosis in NB4 cells in
vitro through the downregulation of the PML/RARα fusion
protein. These results provide new insights for the use of OA in
the therapy for leukemia and indicates that the PML/RARα fusion
protein may be the target of OA.
Acknowledgements
The present study was supported by the Education
Department of Heilongjiang Province (no. 12521631).
References
|
1
|
Wang ZY and Chen Z: Acute promyelocytic
leukemia: from highly fatal to highly curable. Blood.
111:2505–2515. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kwong YL: Pathogenesis and treatment of
leukemia: an Asian perspective. Expert Opin Ther Targets. 16(Suppl
1): S37–S43. 2012. View Article : Google Scholar
|
|
3
|
Li M, Sun K, Redelman D, Welniak LA and
Murphy WJ: The triterpenoid CDDO-Me delays murine acute
graft-versus-host disease with the preservation of
graft-versus-tumor effects after allogeneic bone marrow
transplantation. Biol Blood Marrow Transplant. 16:739–750. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sultana N and Ata A: Oleanolic acid and
related derivatives as medicinally important compounds. J Enzyme
Inhib Med Chem. 23:739–756. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ovesná Z, Vachálková A, Horváthová K and
Tóthová D: Pentacyclic triterpenoic acids: new chemoprotective
compounds. Minireview Neoplasma. 51:327–333. 2004.PubMed/NCBI
|
|
6
|
Kim NY, Lee MK, Park MJ, et al: Momordin
Ic and oleanolic acid from Kochiae Fructus reduce carbon
tetrachloride-induced hepatotoxicity in rats. J Med Food.
8:177–183. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Patil CR, Jadhav RB, Singh PK, Mundada S
and Patil PR: Protective effect of oleanolic acid on gentamicin
induced nephrotoxicity in rats. Phytother Res. 24:33–37. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Singh GB, Singh S, Bani S, Gupta BD and
Banerjee SK: Anti-inflammatory activity of oleanolic acid in rats
and mice. J Pharm Pharmacol. 44:456–458. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Giner-Larza EM, Máñez S, Recio MC, Giner
RM, Prieto JM, Cerdá-Nicolás M and Ríos JL: Oleanonic acid, a
3-oxotriterpene from Pistacia, inhibits leukotriene synthesis and
has anti-inflammatory activity. Eur J Pharmacol. 428:137–143. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hsu HY, Yang JJ and Lin CC: Effects of
oleanolic acid and ursolic acid on inhibiting tumor growth and
enhancing the recovery of hematopoietic system postirradiation in
mice. Cancer Lett. 111:7–13. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim YK, Yoon SK and Ryu SY: Cytotoxic
triterpenes from stem bark of Physocarpus intermedius.
Planta Med. 66:485–486. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li J, Guo WJ and Yang QY: Effects of
ursolic and oleanolic acid on human colon carcinoma cell line
HCT15. World J Gastroenterol. 8:493–495. 2002.PubMed/NCBI
|
|
13
|
Zhang P, Li H, Chen D, Ni J, Kang Y and
Wang S: Oleanolic acid induces apoptosis in human leukemia cells
through caspase activation and poly(ADP-ribose) polymerase
cleavage. Acta Biochim Biophys Sin (Shanghai). 39:803–809. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Okada H and Mak TW: Pathways of apoptotic
and non-apoptotic death in tumor cells. Nat Rev Cancer. 4:592–603.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Roninson IB, Broude EV and Chang BD: If
not apoptosis, then what? Treatment-induced senescence and mitotic
catastrophe in tumor cells. Drug Resist Updat. 4:303–313. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang B and Jiang ZH: Studies of oleanolic
acid. Chinese Pharmaceutical Journal. 27:393–397. 1992.
|
|
18
|
Lúcio KA, da Rocha GG, Monção-Ribeiro LC,
Fernandes J, Takiya CM and Gattass CR: Oleanolic acid initiates
apoptosis in non-small cell lung cancer cell lines and reduces
metastasis of a B16F10 melanoma model in vivo. PLoS One.
6:e285962011.PubMed/NCBI
|
|
19
|
Fernandes J, Castilho RO, da Costa MR,
Wagner-Souza K, Coelho Kaplan MA and Gattass CR: Pentacyclic
triterpenes from Chrysobalanaceae species: cytotoxicity on
multidrug resistant and sensitive leukemia cell lines. Cancer Lett.
190:165–169. 2003.
|
|
20
|
Tokuda H, Ohigashi H, Koshimizu K and Ito
Y: Inhibitory effects of ursolic and oleanolic acid on skin tumor
promotion by 12-O-tetradecanoylphorbol-13-acetate. Cancer Lett.
33:279–285. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Choi CY, You HJ and Jeong HG: Nitric oxide
and tumor necrosis factor-alpha production by oleanolic acid via
nuclear factor-kappaB activation in macrophages. Biochem Biophys
Res Commun. 288:49–55. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee HY, Chung HY, Kim KH, Lee JJ and Kim
KW: Induction of differentiation in the cultured F9 teratocarcinoma
stem cells by triterpene acids. J Cancer Res Clin Oncol.
120:513–518. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sohn KH, Lee HY, Chung HY, Young HS, Yi SY
and Kim KW: Anti-angiogenic activity of triterpene acids. Cancer
Lett. 94:213–218. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rego EM and Pandolfi PP: Analysis of the
molecular genetics of acute promyelocytic leukemia in mouse models.
Semin Hematol. 38:54–70. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dalton WT Jr, Ahearn MJ, McCredie KB,
Freireich EJ, Stass SA and Trujillo JM: HL-60 cell line was derived
from a patient with FAB-M2 and not FAB-M3. Blood. 71:242–247.
1988.PubMed/NCBI
|