Introduction
Osteosarcoma is the most commonly diagnosed primary
malignant tumor of the bone (1,2). The
contemporary treatment of osteosarcoma requires multidisciplinary
therapy, incorporating surgery and systemic chemotherapy (1,3). The
prognosis of osteosarcoma patients has significantly improved with
the advent of chemotherapy. In the pre-chemotherapy era, when
patients underwent surgery as the only form of treatment, the
survival rate was <20% (4).
Osteoclasts have drawn attention as a therapeutic target in various
bone disorders, including osteosarcoma. The osteoclast is the only
cell that resorbs bone and is central to pathological situations,
where bone destruction is intricately involved. Osteosarcoma cells
are of the osteoblastic lineage. Hence, osteosarcoma is a more
ideal candidate for osteoclast-targeted therapy than other primary
and metastatic bone tumors. The rapid progress that has been made
in understanding the molecular mechanism that regulates osteoclasts
has propelled the development of new therapeutic approaches
(5).
Autophagy is intricately implicated in health and
disease. Autophagy defects play a role in the pathogenesis of
numerous diseases, including myopathy, neuronal degeneration,
microbial infection, inflammatory bowel disease, aging and cancer
(6–11). Studies have demonstrated the
functional role of autophagy in various cellular processes and the
potential of autophagy modulation as a novel therapeutic strategy
for a number of pathological conditions, including cancer (12–14).
Anticancer therapies, including hormonal agents,
chemotherapy and irradiation, frequently induce autophagy, in most
cases as a prosurvival response potentially contributing to
treatment resistance. However, autophagy activation in particular
genetic backgrounds and/or the completion of the autophagic process
beyond the reversibility of cell viability may also lead to cell
death, thus enhancing the efficacy of the treatment (15–18).
The activation of autophagy by the inhibition of the
mammalian target of rapamycin (mTOR) may contribute to anti-tumor
actions. The present study examined the effects of the mTOR
inhibitor, rapamycin, on the activation of autophagy and the
contribution of autophagy to the chemosensitivity effects of
cis-diamminedichloroplatinum (CDDP) on osteosarcoma MG63 cells.
Materials and methods
Reagents
MG63 osteosarcoma cancer cells were purchased from
the Shanghai Institute of Cell Biology, Chinese Academy of Sciences
(Shanghai, China). RPMI-1640 medium was purchased from Gibco
(Rockville, MD, USA) and rapamycin were purchased from Biovision
Technology (Milpitas, CA, USA). Fetal bovine serum was purchased
from Hangzhou Sijiqing Biological Engineering Material Co., Ltd.,
(Hangzhou, Zhejiang, China) and L-glutamine and MTT were purchased
from Sigma (St Louis, MO, USA). Rabbit monoclonal anti-p53, -p62,
-Beclin-1 and -light chain 3 (LC3) antibodies were purchased from
Cell Signaling Technology (Beverly, MA, USA).
Drug preparation
Rapamycin (Biovision Technology) was diluted in DMSO
to create a stock solution that was stored according to the
manufacturer’s instructions. The final concentrations of the
rapamycin and CDDP solutions used were 5 and 2 μmol/l,
respectively. This concentration of rapamycin was selected on the
basis of our experiments on the MG63 cells.
Cell culture and viability assay
The MG63 cells were maintained in RPMI-1640 medium
containing 10% heat-inactivated fetal bovine serum and 0.03%
L-glutamine, and incubated in a 5% CO2 atmosphere at
37ºC. The cells that were in a mid-log phase were used in the
experiments. The cell viability was assessed using an MTT assay. To
determine the time-course of the response of the MG63 cells to
rapamycin and CDDP, the MG63 cells were plated into 96-well
microplates (7×104 cells/well). Rapamycin (5 μmol/l) and
CDDP (2 μmol/l) was added to the culture medium and the cell
viability was assessed using the MTT assay at 24, 48 and 72 h
following the drug treatment. MTT solution (Sigma) was added to the
culture medium (500 μg/ml final concentration) for 4 h prior to the
end of treatment and the reaction was stopped by the addition of
100 μl 10% acidic SDS. The absorbance value (A) at 570 nm was read
using an automatic multiwell spectrophotometer (Bio-Rad, Richmond,
CA, USA). The percentage of cell deaths was calculated as follows:
Cell death (%) = (1 − A of experimental well / A of positive
control well) × 100.
Detection of mitochondrial membrane
potential (Δψ)
The mitochondrial Δψ was determined using the KeyGen
Mitochondrial Membrane Sensor kit (KeyGen, Nanjing, Jiangsu,
China). The mitosensor dye aggregates in the mitochondria of
healthy cells and emits red fluorescence against a green monomeric
cytoplasmic background staining. However, in cells with a collapsed
mitochondrial Δψ, the dye is not able to accumulate in the
mitochondria and remains as monomers throughout the cells emitting
green fluorescence (19). Briefly,
the MG63 cells were incubated with rapamycin and CDDP in 24-well
plates for the indicated times and then pelleted, washed with PBS
and resuspended in 0.5 ml diluted mitosensor reagent (1 μmol/ml in
incubation buffer). Subsequent to incubating the cells with
mitosensor reagent for 20 min, 0.2 ml incubation buffer was added
and the cells were centrifuged and then resuspended in 40 μl
incubation buffer. Finally, the cells were washed and resuspended
in 1 ml PBS for flow cytometry analysis.
Detection of the cell cycle
To analyze the effects of rapamycin on cell
apoptosis progression, the MG63 cells were incubated with rapamycin
and CDDP. The cells were harvested using 0.25% trypsin, washed with
PBS, counted and adjusted to a concentration of 1×106
cells/ml. The cells were fixed in 70% ethanol, treated with 100
mg/l RNase at 37ºC for 30 min and then stained with 50 mg/l
propidium iodide (Sigma) for 30 min. The cells were analyzed using
flow cytometry (Epics XL; Beckman Coulter, Fullerton, CA, USA).
Total cell protein extraction and western
blot analysis
For the extraction of the total cell proteins, the
cells were washed with pre-cooled PBS and subsequently lysed in a
pre-cooled RIPA lysis buffer containing 50 mM Tris-HCl (pH 7.4) 150
mM NaCl, 1 mM dithiothreitol (DTT), 0.25% sodium deoxycholate, 0.1%
NP-40, 1 mM phenylmethysulfonyl fluoride (PMSF), 50 mM sodium
pyrophosphate, 1 mM Na3VO4, 1 mM NaF, 5 mM
EDTA, 5 mM EGTA and a protease inhibitor cocktail. Cell lysis was
performed on ice for 30 min. Clear protein extracts were obtained
by centrifugation at 12,000 × g for 30 min at 4ºC. The protein
extraction procedure from the MG63 cells was performed as
previously described. The protein concentration was determined
using a Bradford protein assay kit (KeyGen). The proteins were
resolved on 8.5% polyacrylamide gels and subsequently transferred
onto nitrocellulose membranes. For immunoblotting, the
nitrocellulose membranes were incubated overnight at 4ºC with
specific antibodies recognizing the target proteins. The membranes
were then incubated with a horseradish peroxidase (HRP)-conjugated
secondary antibody (1:3,000) for 1 h at room temperature and
subsequently analyzed using an enhanced chemiluminescence (ECL)
detection system (Amersham Pharmacia Biotech, Amersham, UK)
and visualized by autoradiograpy. β-actin proteins (1:5,000; Sigma)
were used as loading controls.
Statistical analysis
All data are presented as the mean ± SD. The
statistical analysis was performed using an ANOVA followed by
Dunnett’s t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Rapamycin inhibits cell viability and
enhances the effects of CDDP-induced tumor cell growth
inhibition
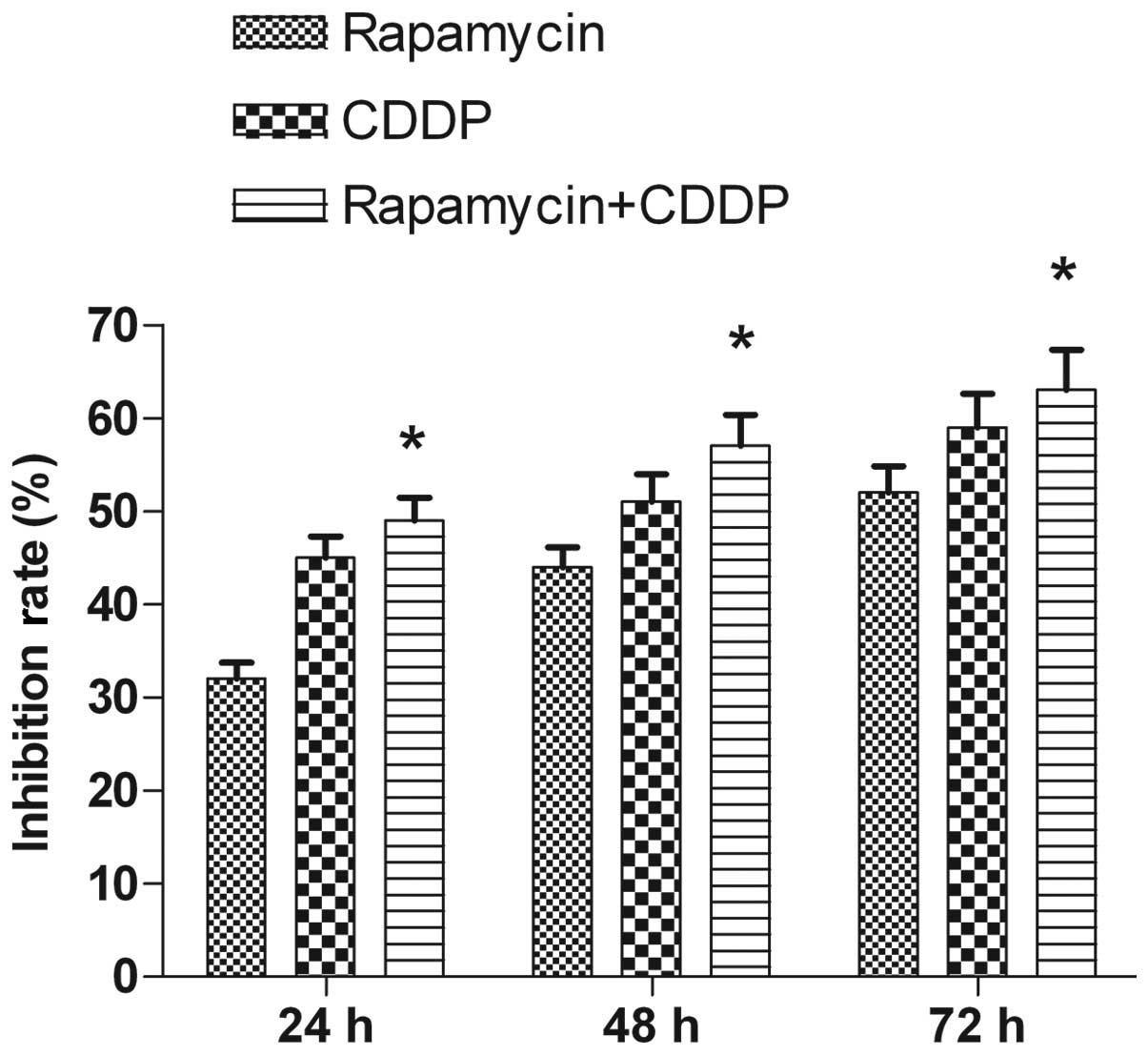
In the present study, rapamycin reduced MG63 cell
viability in a time-dependent manner. The MTT assays revealed that
following 24 h of treatment, the rate of inhibition reached
32±1.76% at the dose (5 μmol/l) used. The rate of inhibition
increased when the incubation time was prolonged, reaching 44±2.09%
at 48 h and 52±2.87% at 72 h following the treatment (Fig. 1). CDDP (2 μmol/l) was used to assess
the clinical value of the mTOR inhibitor in the treatment of the
tumor and to test the synergistic inhibitory effect of the mTOR
inhibitor on the growth of the cells in combination with a
chemotherapy drug. Rapamycin was shown to have an increased effect
when used in combination with CDDP compared with when used alone
(Fig. 1). Thus, rapamycin inhibited
the proliferation of the MG63 cells and enhanced the
chemosensitivity of CDDP.
Rapamycin induces mitochondrial
dysfunction and enhances the effects of CDDP-induced mitochondrial
dysfunction
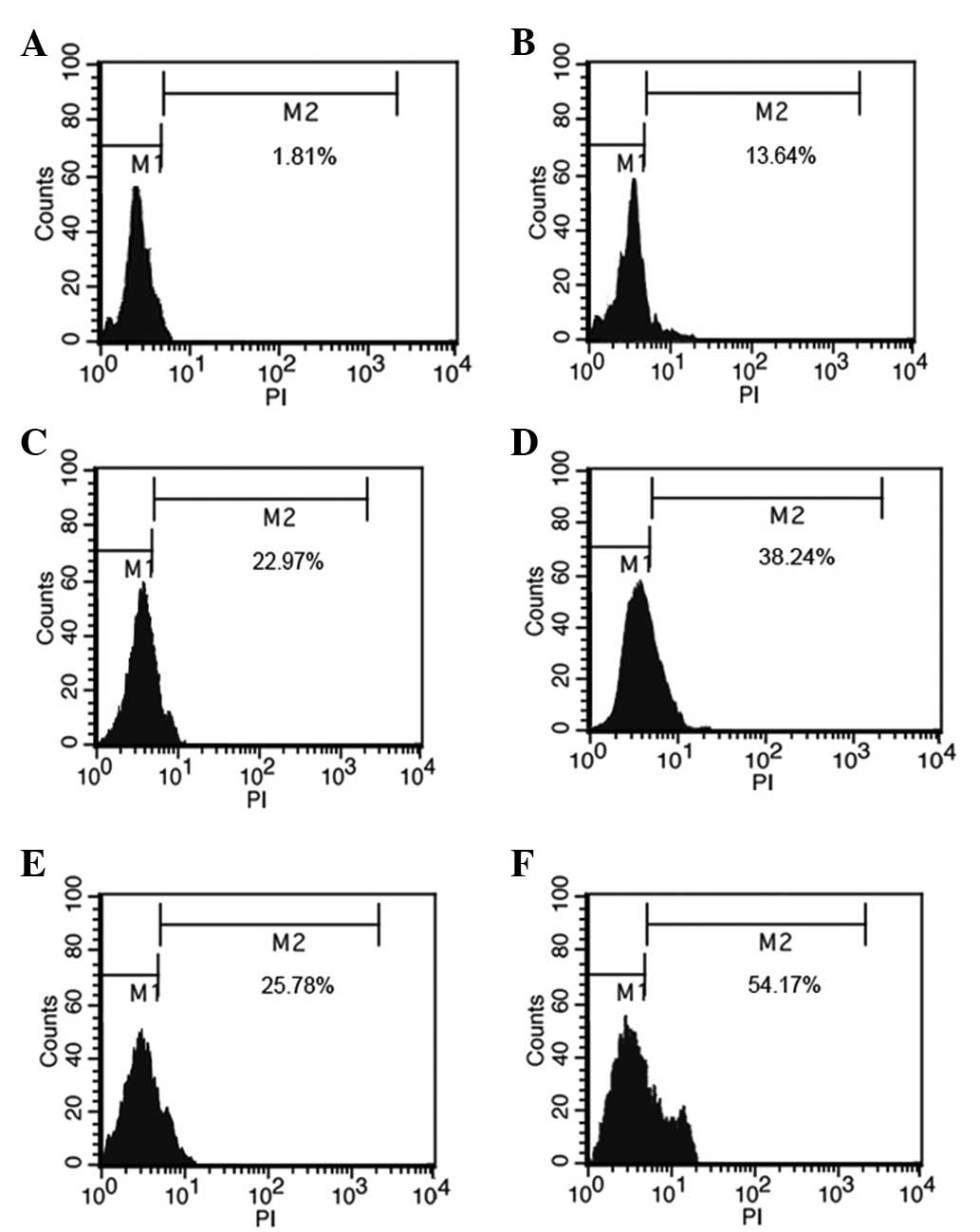
In the present study, the mitochondrial Δψ was
examined using the fluorescent dye, JC-1. A collapse in the
mitochondrial Δψ was detected as early as 6 h after rapamycin or
CDDP treatment, as indicated by an increased emission of green
fluorescence. This change reached a maximum level following 24 h of
rapamycin treatment or 12 h of CDDP treatment (Fig. 2). A collapse in mitochondrial Δψ
indicates cell apoptosis or necrosis. Rapamycin used in combination
with CDDP, rather than used alone, induced mitochondrial
dysfunction and activated cell apoptosis in the MG63 cells. The
present results demonstrate that rapamycin enhanced the effects of
CDDP-induced mitochondrial dysfunction in the MG63 cells.
Rapamycin induces apoptosis and enhances
the effects of the CDDP-induced apoptosis of MG63 cells
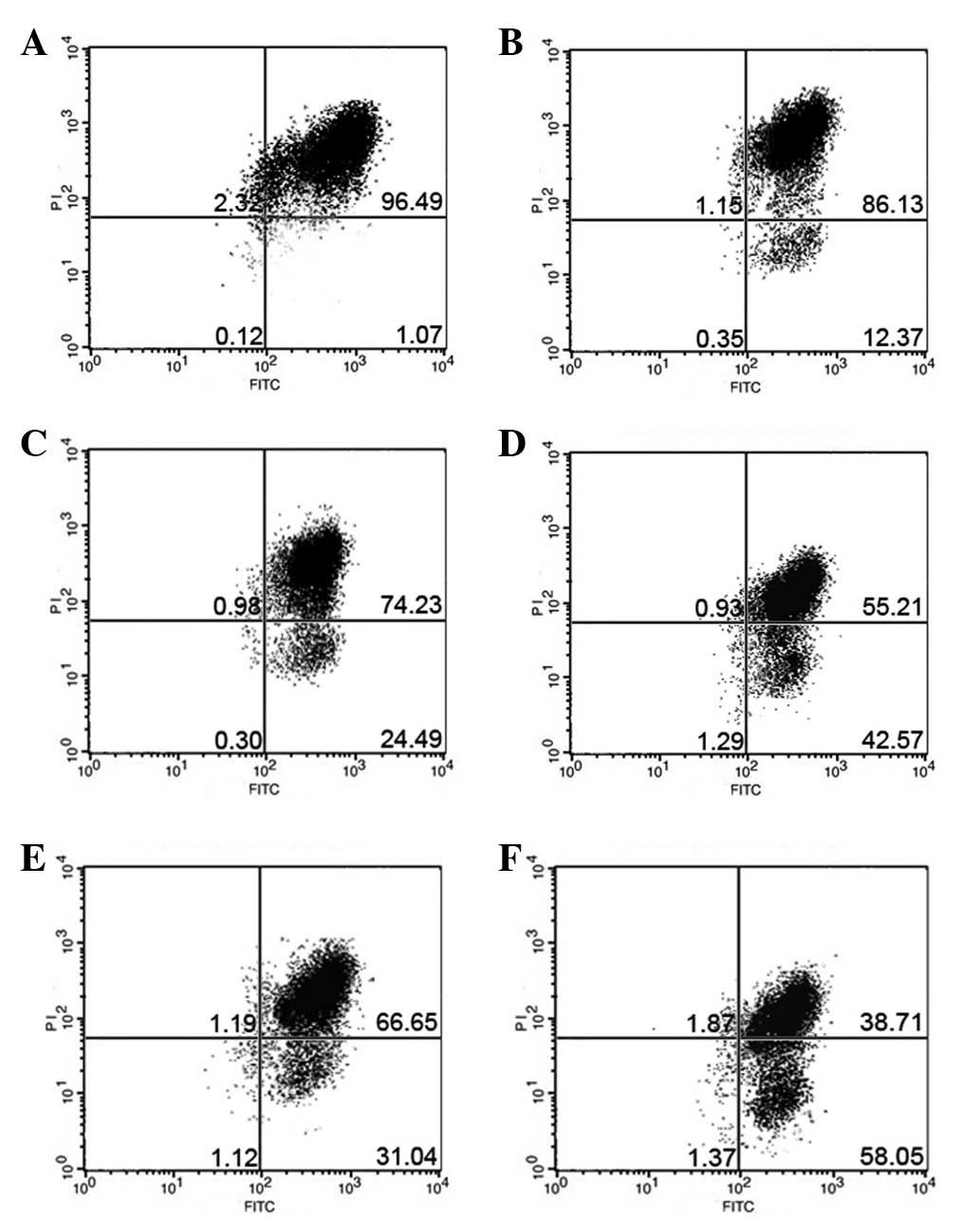
The effect of rapamycin on the cell apoptosis
progression of the MG63 cells was studied following 6, 12 and 24 h
of exposure to 5 μmol/l rapamycin. The flow cytometry analysis
indicated that rapamycin induced cell apoptosis following 6, 12 and
24 h of treatment. There was a significant difference between the
5μmol/l rapamycin and CDDP groups and the control group at 12 h.
When the cells were treated with rapamycin and CDDP at 12 h, the
apoptosis of the MG63 cells was significantly increased than when
rapamycin was used alone (Fig. 3).
The results indicated that rapamycin enhanced the effects of
inducing MG63 cell apoptosis by CDDP.
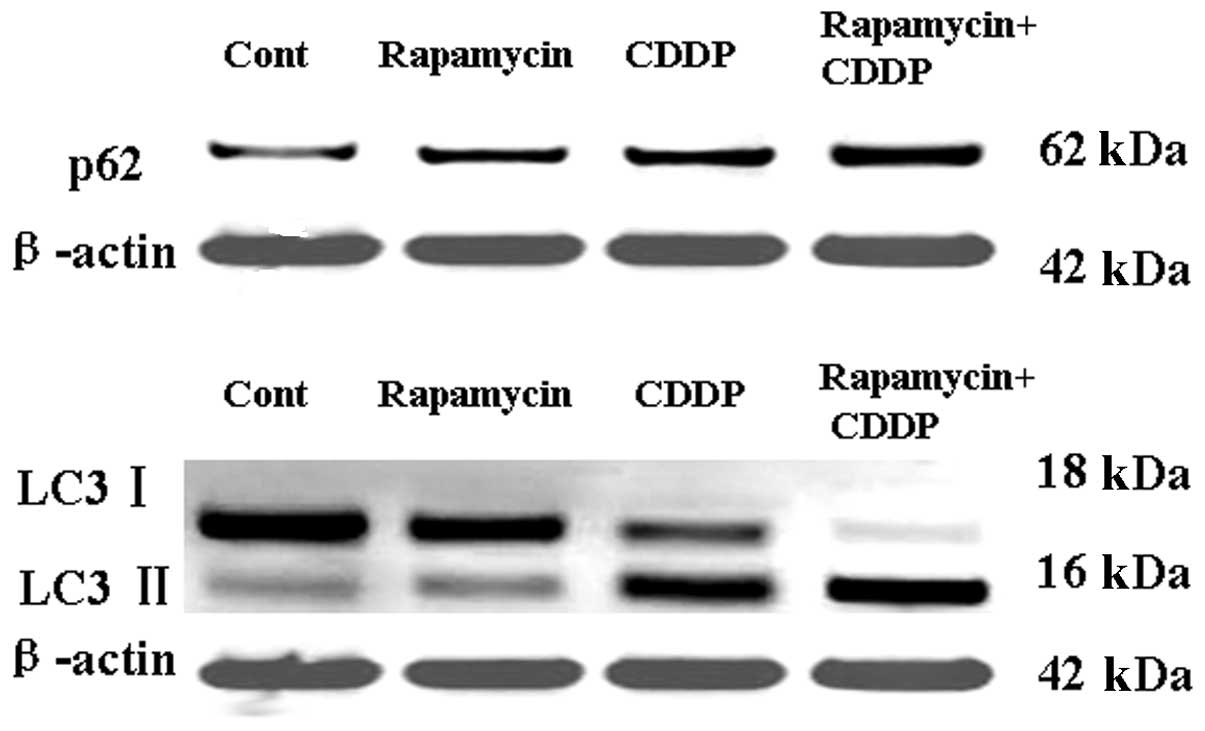
Rapamycin increases the expression of p62
and LC3 and enhances the effects of CDDP-activated autophagy
To distinguish the specific inhibition of
mTOR-mediated cell proliferation from autophagy, the expression of
the autophagic proteins, LC3 and p62, was measured following
treatment with rapamycin or CDDP. As shown in Fig. 4A, the expression of p62 in the MG63
cell line was activated by rapamycin or CDDP treatment, and
rapamycin may have enhanced the effects of CDDP-activated
autophagy. As shown in Fig. 4B,
western blotting analysis was used to detect the protein levels of
LC3-I and LC3-II. The results revealed that the levels of LC3,
particularly LC3-II, had increased, leading to an increased ratio
of LC3-II/LC3-I following rapamycin or CDDP treatment. Rapamycin
may have thus enhanced the effects of CDDP-activated autophagy.
These results suggest that rapamycin may induce tumor cell
apoptosis by regulating p62 and LC3II.
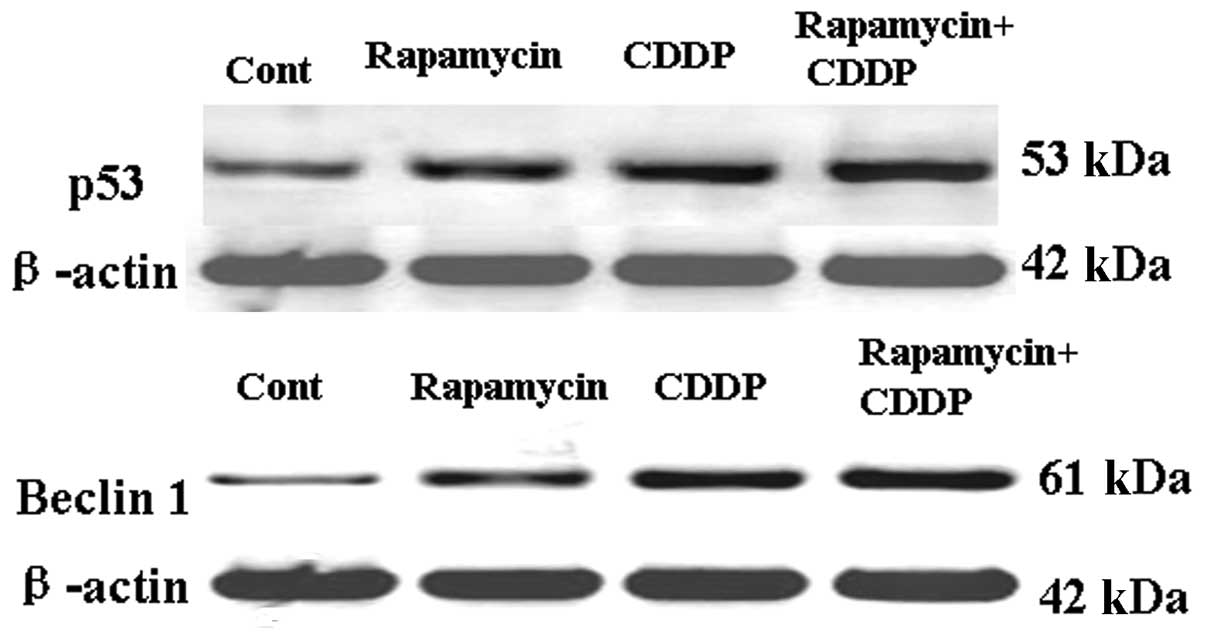
Rapamycin upregulates p53 and Beclin-1
and enhances the effects of CDDP in the upregulation of p53 and
Beclin-1
To distinguish the specific inhibition of
mTOR-mediated cell proliferation from autophagy and apoptosis, the
expression of the autophagic and apoptotic proteins, p53 and
Beclin-1, was measured following treatment with rapamycin or CDDP.
As shown in Fig. 5A, the expression
of Beclin-1 in the MG63 cell line was activated by rapamycin or
CDDP treatment and rapamycin may have enhanced the effects of
CDDP-induced autophagy and apoptosis. As shown in Fig. 5B, the expression of p53 in the MG63
cell line was also upregulated and rapamycin may have enhanced the
effects of CDDP-induced apoptosis. These results suggest that
rapamycin may induce tumor cell apoptosis by regulating Beclin-1
and p53.
Discussion
The tumor suppressor, p53, plays a central role in
sensing various genotoxic stresses. p53 is known to play
significant roles in apoptosis by regulating the expression of
proapoptotic proteins (20,21). The present study demonstrated that
as an inhibitor of autophagy, rapamycin significantly upregulated
the levels of p53, indicating that apoptosis may be triggered by
CDDP. The mitochondrial Δψ was shown to collapse following
rapamycin treatment. The study revealed that rapamycin induced
mitochondrial dysfunction in the MG63 cells. Mitochondria play a
central role in regulating cell death and survival. Diverse
proapoptotic stimuli act on mitochondria, triggering mitochondrial
Δψ collapse, cytochrome c release and caspase activation. The
mitochondrial permeability transition (MPT) represents a
significant event in initiating apoptotic cell death (22).
Increasing evidence suggests that autophagy plays
significant roles in tumor cell growth, differentiation and the
response to anti-tumor drugs (23).
Numerous classical anti-tumor drugs have now been identified to
exert their cytotoxic actions by autophagic mechanisms (24–26).
In the present study, the inhibition of mTOR by rapamycin resulted
in a significant increase in the levels of p62, LC3 and Beclin-1,
and a particularly increased production of LC3-II. LC3 is an
autophagosomal ortholog of yeast Atg8. LC3 has been best
characterized as an autophagosomal marker in mammalian autophagy
and the levels of LC3 may also reflect the levels of autophagy
(27). Beclin-1 is the mammalian
ortholog of the yeast ATG6-Vps30 gene. p62 is also present in
protein aggregates that are positive for ubiquitin and
microtubule-associated protein-1 (MAP1)LC3, a well-characterized
marker of a cellular process known as autophagy. p62 is believed to
be the link between polyubiquitinated proteins and autophagy
(28). The present results suggest
that autophagy inhibited by rapamycin may contribute to cell
apoptosis.
In summary, the present study revealed a new
mechanism associated with mTOR inhibition that triggered the
impairment of cell proliferation and the induction of the cell
death of cancer cells. The inhibition of mTOR increases the
expression of p53 and induces the expression of the proapoptotic
and autophagic proteins, p62, LC3 and Beclin-1. Rapamycin induced
the death of cancer cells through apoptotic and autophagic
mechanisms and may have enhanced the effects of CDDP on activating
autophagy and inducing apoptosis. Further investigation of the
association between autophagy activation and the anti-tumor effects
of mTOR inhibitors may unveil new strategies for tumor therapy.
References
|
1
|
Raymond AK, Ayala AG and Knuutila S:
Conventional osteosarcoma. World Health Organization Classification
of Tumors. Fletcher CDM, Unni KK and Mertens F: 4th edition. IARC
Press; Lyon, France: pp. 264–270. 2002
|
|
2
|
Tan JZ, Schlicht SM, Powell GJ, et al:
Multidisciplinary approach to diagnosis and management of
osteosarcoma-a review of the St Vincent’s Hospital experience. Int
Semin Surg Oncol. 3:382006.
|
|
3
|
Ferrari S and Palmerini E: Adjuvant and
neoadjuvant combination chemotherapy for osteogenic sarcoma. Curr
Opin Oncol. 19:341–346. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marcove RC, Miké V, Hajek JV, Levin AG and
Hutter RV: Osteogenic sarcoma under the age of twenty-one. A review
of one hundred and forty-five operative cases. J Bone Joint Surg
Am. 52:411–423. 1970.PubMed/NCBI
|
|
5
|
Akiyama T, Dass CR and Choong PF: Novel
therapeutic strategy for osteosarcoma targeting osteoclast
differentiation, bone-resorbing activity, and apoptosis pathway.
Molecular Cancer Ther. 7:3461–3469. 2008. View Article : Google Scholar
|
|
6
|
Malicdan MC, Noguchi S, Nonaka I, Saftig P
and Nishino I: Lysosomal myopathies: an excessive build-up in
autophagosomes is too much to handle. Neuromuscul Disord.
18:521–529. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Winslow AR and Rubinsztein DC: Autophagy
in neurodegeneration and development. Biochim Biophys Acta.
1782:723–729. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Orvedahl A and Levine B: Eating the enemy
within: autophagy in infectious diseases. Cell Death Differ.
16:57–69. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cadwell K, Liu JY, Brown SL, Miyoshi H,
Loh J, Lennerz JK, et al: A key role for autophagy and the
autophagy gene Atg16l1 in mouse and human intestinal Paneth cells.
Nature. 456:259–263. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Saitoh T, Fujita N, Jang MH, Uematsu S,
Yang BG, Satoh T, et al: Loss of the autophagy protein Atg16L1
enhances endotoxin-induced IL-1beta production. Nature.
456:264–268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yen WL and Klionsky DJ: How to live long
and prosper: autophagy, mitochondria and aging. Physiology
(Bethesda). 23:248–262. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mathew R, Karantza-Wadsworth V and White
E: Role of autophagy in cancer. Nat Rev Cancer. 7:961–967. 2007.
View Article : Google Scholar
|
|
13
|
Carew JS, Nawrocki ST and Cleveland JL:
Modulating autophagy for therapeutic benefit. Autophagy. 3:464–467.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen N and Karantza-Wadsworth V: Role and
regulation of autophagy in cancer. Biochim Biophys Acta.
1793:1516–1523. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Al-Ejeh F, Kumar R, Wiegmans A, Lakhani
SR, Brown MP and Khanna KK: Harnessing the complexity of DNA-damage
response pathways to improve cancer treatment outcomes. Oncogene.
29:6085–6098. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gewirtz DA: Autophagy as a mechanism of
radiation sensitization in breast tumor cells. Autophagy.
3:249–250. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
John S, Nayvelt I, Hsu HC, Yang P, Liu W,
Das GM, et al: Regulation of estrogenic effects by beclin 1 in
breast cancer cells. Cancer Res. 68:7855–7863. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Buytaert E, Callewaert G, Vandenheede JR
and Agostinis P: Deficiency in apoptotic effectors Bax and Bak
reveals an autophagic cell death pathway initiated by photodamage
to the endoplasmic reticulum. Autophagy. 2:238–240. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Petronilli V, Miotto G, Canton M, et al:
Imaging the mitochondrial permeability transition pore in intact
cells. Biofactors. 8:263–272. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moll UM and Zaika A: Nuclear and
mitochondrial apoptotic pathways of p53. FEBS Lett. 493:65–69.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Erster S, Mihara M, Kim RH, Petrenko O and
Moll UM: In vivo mitochondrial p53 translocation triggers a rapid
first wave of cell death in response to DNA damage that can precede
p53 target gene activation. Mol Cell Biol. 24:6728–6741. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hirsch T, Marzo I and Kroemer G: Role of
the mitochondrial permeability transition pore in apoptosis. Biosci
Rep. 17:67–76. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gozuacik D and Kimchi A: Autophagy as a
cell death and tumor suppressor mechanism. Oncogene. 23:2891–2906.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bursch W, Ellinger A, Kienzl H, Török L,
Pandey S, Sikorska M, et al: Active cell death induced by the
anti-estrogens tamoxifen and ICI 164 384 in human mammary carcinoma
cells (MCF-7) in culture: the role of autophagy. Carcinogenesis.
17:1595–1607. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kanzawa T, Germano IM, Komata T, Ito H,
Kondo Y and Kondo S: Role of autophagy in temozolomide-induced
cytotoxicity for malignant glioma cells. Cell Death Differ.
11:448–457. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Paglin S, Hollister T, Delohery T, Hackett
N, McMahill M, Sphicas E, et al: A novel response of cancer cells
to radiation involves autophagy and formation of acidic vesicles.
Cancer Res. 61:439–444. 2001.PubMed/NCBI
|
|
27
|
Kabeya Y, Mizushima N, Ueno T, et al: LC3,
a mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kirkin V, McEwan DG, Novak I and Dikic I:
A role for ubiquitin in selective autophagy. Mol Cell. 34:259–269.
2009. View Article : Google Scholar : PubMed/NCBI
|