1. Introduction
Stress or heat shock proteins (HSPs) were first
identified in 1962 as highly-conserved proteins, with expression
that is induced by various types of stress (1). Subsequently, it was shown that
intracellular HSPs behave as molecular chaperones for other
cellular proteins (2,3). HSPs perform a variety of chaperone
functions, including the folding and unfolding of nascent
polypeptides and proteins (4), the
degradation of proteins (5), the
transport of proteins and peptides throughout the various cellular
compartments (6–8) and the support of antigen presentation
processes (9,10). Furthermore, HSPs have been
previously shown to have a role in priming immune responses when
released from cells in complex with chaperoned antigenic peptides
(11). In murine systems,
vaccination with purified preparations of selected HSPs (GP96,
HSP70 and HSP90) isolated from tumors, but not from normal tissues,
leads to protective immunity against the tumors used as the source
of the HSPs (12,13). Other than this protection property,
the treatment of mice bearing established or residual tumors with
such vaccines is effective in reducing the tumor burden and
metastasis and prolonging survival (14,15).
In clinical trials, autologous tumor-derived HSP peptide complexes
(HSP.PCs) have been applied to tumor immunotherapy for the
treatment of patients with a variety of advanced malignancies. To
date, results from clinical trials, including those from phase III
trials, have demonstrated that the immunization strategy induces
significant tumor-specific immune responses (16,17).
These results have indicated that HSP.PCs have the qualities
necessary for a tumor vaccine.
However, the efficacy of the HSP.PC vaccine requires
improvement. As a therapeutic tool against established tumors, it
is only marginally effective, particularly with a widely metastatic
disease (18). In rodent models,
autologous tumor-derived HSP.PCs were highly effective in the
treatment of a minimal residual disease setting (14,19).
However, when animals with a widely metastatic disease were
treated, only a small proportion showed any benefit and typically,
only a slowing of the tumor growth rate or stabilization of the
disease was observed (14,20–22).
The success of the clinical trial was also limited, which is
consistent with the observations made in animals (16,17).
These results indicated that novel HSP-based tumor vaccines, with
improved therapeutic potentials, require investigation, and
researchers are thus currently making efforts to identify
alternatives.
2. Location and dual function of HSPs
HSPs are present in all cells in all forms of life
and have a dual function depending on their intracellular or
extracellular location. Intracellular locations include the cytosol
of prokaryotes and the cytosol, nuclei, endoplasmic reticulum (ER),
mitochondria and chloroplasts of eukaryotes (2). HSPs normally constitute 5% of the
total intracellular proteins. However, under various stresses, such
as high temperatures, toxins and oxidative conditions, their levels
may rise to >15% (23).
Intracellular HSPs have a protective function and allow the cells
to exert themselves against environmental stress to survive lethal
conditions. Various mechanisms attribute to this cytoprotective
function.
In addition to their intracellular location, HSPs
have been located on the plasma membrane of malignantly transformed
cells, in the extracellular space and on virally/bacterially
infected cells. The extracellularly located or membrane-bound HSPs
mediate immunological functions through the chaperoning of
antigenic peptides. HSPs elicit immune responses modulated by the
adaptive or innate immune system, and these immunogenic properties
make HSPs good targets for tumor therapy (24).
3. Immune responses by HSPs
HSPs originally came to the attention of
immunologists primarily as besides their chaperone activity, three
additional features characterize HSPs: i) HSPs are efficiently
internalized into antigen-presenting cells (APC) by
receptor-mediated endocytosis (25); ii) once internalized, HSPs traffic
into various cellular compartments where chaperoned peptides are
released, processed and made available for assembly to new major
histocompatibility complex (MHC) molecules (26); and iii) the internalization of
chaperone proteins induces the immune responses, which eventually
activates the adaptive (CD8+ and CD4+
lymphocytes) and innate [natural killer (NK) cell activation and
cytokine secretion and maturation of dendritic cells (DCs)] immune
responses (27). Due to these
properties, HSPs have been described as the ‘Swiss army knife’ fo
the immune system (27), and this
maxim captures the versatility of HSPs. These features allow HSPs
to be exploited to engineer new tumor vaccines and potentially, to
generate a full-fledged immune response overcoming tumor escape and
interfering with growth and metastasis (28).
4. HSP categories
HSPs are classified into >10 families, including
small HSPs, HSP40, HSP47, calreticulin, HSP60, HSP70, HSP90 and
HSP100, which have various locations and functions (23). Each family is composed of members
expressed constitutively or regulated inductively and are targeted
to various subcellular compartments. Each HSP family consists of
between one to five closely related proteins, although there is
little evident amino acid homology between HSP families.
5. History of HSP immunity
Origin of tumor immunotherapy in the
early 20th century
The earliest tumor immunotherapy has been traced
back to the early 20th century, when physicians attempted to treat
tumor patients by vaccination with attenuated tumor cells or their
crude extracts. This pioneering study was the origin of the current
understanding of the versatile properties of HSPs, which was
unclear prior to the 1980s (29).
There has been little basis to determine whether these pioneering
studies had merit, since the controls commonly used today were
absent at that time (30).
Origin of tumor vaccines in the
1940s–1960s
The vaccines against smallpox and polio have been
successfully used, and previous tumor studies have attempted to
design tumor vaccines that are immune against the challenge of
tumor cells, in the same manner as they are against viruses. With
the utilization of inbred strains of mice in the 1940s,
interpretable studies of tumor immunity were initiated and the
fundamental principles gradually became clear. It was demonstrated
that the vaccination of mice with attenuated tumor cells provided
protection from a subsequent challenge with live tumor cells, just
as a vaccination with attenuated virus immunizes against a
subsequent infection with that same virus (31–34).
However, in contrast to viruses, tumors may be used to vaccinate
against themselves, but not against other tumors. This observation
elicited the subsequent studies that led to the emergence of
HSPs.
Purification of HSPs for tumor vaccines
in the 1980s
The aforementioned research identified that animals
may be specifically vaccinated against their own tumors, but not
other tumors, thus promoting the search for the molecules within
tumor cells that may be responsible for conferring immunity. This
identification predicted that tumors are individually and
antigenically distinct and express tumor-specific antigens. Tumor
cell lysates were fractionated biochemically and individual
fractions were tested for their ability to vaccinate mice against
the subsequent challenge of live tumor cells of the same type used
for fractionation. Finally, it was demonstrated that fractions that
reproducibly protected mice from the tumor challenge contained HSPs
(12,29,35).
HSPs were initially purified from tumor cells and shown to provide
protective immunity in a rat liver cancer model (29). This study was corroborated by two
additional studies in 1986 in a mouse fibrosarcoma model (12,35).
Consequently, researchers hypothesized that tumor-specific antigens
may be heat shock-related proteins. HSP-based tumor vaccines were
rapidly and widely tested in animal models. The results were
unexpected and in almost every model tested the HSPs demonstrated
efficacy; there has been no precedent in tumor immunology (30). This identification inspired
researchers to extensively test HSP immunotherapy in cancer
patients.
HSP.PC is the immunogenic entity
As HSP preparations elicit immunity only against the
tumors from which they were isolated, instead of antigenically
distinct tumors (14), this
established an immunological conundrum, since HSPs are among the
most highly conserved proteins in evolution. Among human beings and
even between humans and mice, HSPs are >95% conserved (2). Therefore, how such conserved
molecules, which exist everywhere, were able to elicit specific
tumor immunity remained unknown. To answer this conundrum, HSP
preparations were isolated from normal tissues and used to immunize
the animals, however, no resistance to any tumors was generated
(36,37). Therefore, it was hypothesized that
HSPs in tumors may harbor mutations that differ among various
tumors. However, this hypothesis was excluded by the subsequent
investigations.
To test the hypothesis, HSP genes among normal
tissues and a variety of tumors were sequenced. Notably, no
differences in nucleotide sequences were detected among any of the
tissues or tumors (38,39). Therefore, this indicated that it is
not HSPs that elicit the tumor antigen-specific immune responses.
Advanced investigations were therefore required.
The breakthrough point for solving how HSPs elicit
specific tumor immunity depended on the normal function of HSPs
inside cells. It is well known that HSPs aid newly synthesized
polypeptides in folding into their functional conformation and also
aid in the transport of proteins and peptides throughout the
various subcellular compartments. Furthermore, it had already been
made clear that tumor immunity is mainly mediated by T cells, which
recognize MHC molecule-peptide complexes. Therefore, it was
indicated that HSP preparations from tumor cells may form complexes
with antigenic peptides (39),
which activate T cell-mediated responses. Several results supported
this indication. Firstly, from the HSP structural aspect, the
peptide-binding domain of bacterial HSP70 was crystallized with
intact peptide in a readily discernible peptide-binding pocket
(40). HSP70 family members possess
a domain in the C-terminus that chaperones unfolded proteins and
peptides and an N-terminal ATPase domain that controls the opening
and closing of the peptide-binding domain. These properties promote
the formation of stable complexes with tumor antigen peptides
(41). Secondly, a large collection
of peptides may be eluted from a homogeneous gp96 preparation, as
it is treated with trifluoroacetic acid (9). Thirdly, much stronger support for this
indication came when the treatment of a tumor-derived HSP70
preparation with ATP had the following two consequences: i) elution
of a wide array of peptide peaks from the HSP70 polypeptide,
leaving the polypeptide intact; and ii) rendering of the HSP70
preparation as non-immunogenic and ineffective in immunizing
against tumor cells, although the amount of HSP70 polypeptide
treated by ATP was equivalent in the untreated HSP70 preparations
(42). This was the first
demonstration that HSP70, isolated from tumors, was associated with
peptides and that the dissociation of peptides from HSP70 resulted
in abrogation of the immunogenicity. It was further demonstrated
that such peptide-free HSP preparations or free, unchaperoned
peptides were not immunogenic (42). Three mammalian HSPs (gp96, HSP90 and
HSP70) were purified from tumor cells or pathogen-infected cells,
and non-covalently associated peptides were eluted off the HSP with
acid or ATP.
Thus, it was confirmed that HSP.PCs are the
immunogenic entities isolated from tumorous or infected cells. HSPs
chaperone the peptide fingerprint, which includes the antigenic
peptides of the cells from which they were isolated. HSP
preparations, which harbor the unique repertoire of antigenic
peptides that exist in individual tumors, elicit the tumor specific
immune responses.
Summary of previous clinical trials
The earliest clinical trial for HSPs began in 1995
with a phase I pilot study in patients with advanced malignancies
that had failed to be treated with previous therapies. This initial
autologous HSP vaccine was HSP-peptide complex 96 (HSPPC-96;
Oncophage; Antigenics Inc., Lexington, MA, USA), produced from
surgically resected tumor tissue and formulated for intradermal or
subcutaneous injection (43). The
purpose was to determine the proper dosage, to address potential
toxicities and side-effects and also to make comparisons with mice
in inducing immune responses. The results demonstrated that the
autologous tumor-derived HSP vaccine elicited powerful T cell
responses against the tumor and did not produce any toxicity, which
was consistent with the results obtained in mice. In addition,
these results demonstrated the feasibility of the preparation of
individual autologous HSP vaccines from each patients’ own tumor.
However, since these studies were non-randomized and were only
compared with historical controls, the results from these trials
were only indicative. Other phase I and II trials in pancreatic or
colon cancers have since been completed with similar outcomes.
Trials in B lymphoma, chronic myelogenous leukemia, lung cancer and
glioma are ongoing (44,45).
In 2001, a study by Castelli et al
demonstrated in a human system that HSP.PCs purified from tumor
cells activate cytotoxic T lymphocyte (CTL) clones specific for
defined tumor antigens (7). This
study further highlighted the reliable theoretical basis for the
use of HSP vaccine in patients.
To further investigate the clinical effect of the
autologous HSP vaccine, two phase III trials followed. The first
study focused on stage IV melanoma patients and ~322 patients were
involved in a randomized, open-label, multicenter phase III trial
(16). The patients in the
treatment group received HSP vaccine derived from autologous
cancers, and the regimen was administration once weekly for the
first 4 weeks and subsequently every other week, for as long as the
vaccine lasted. The patients in the control group received the
physician’s choice of treatment, which consisted of a specific
combination of dacarbazine, temozolomide, interleukin (IL)-2 and
surgery. The general analysis from the survival plots showed no
significant difference between the HSP vaccine treatment and
control groups. However, a specific subset analysis was more
encouraging, informative and significant. Two critical observations
were formed as follows: i) when the vaccine dose increased,
patients treated with vaccine received a greater benefit; and ii)
with an increasing number of immunizations, the hazard ratios
shifted to the left (in favor of vaccine) in M1a and M1b substages,
but not M1c substages. However, the success rate for the production
of vitespen (four injections are the minimal dosage for vitespan
administration) was only 49%, the main reason being the limitated
quantity of resected tumor available for HSP isolation.
The vaccine was effective in the early stage of the
disease instead of the late stage of the disease. When the HSP
regimen was limited to <10 doses, patients with M1a and M1b
stages exhibited improved survival rates compared with the control
group, which was a statistically significant result. However, no
difference was identified between the HSP treatment and control
groups for the patients in the M1c substage.
A second phase III trial, in which 728 patients were
involved, focused on renal cell carcinoma. To date, this is the
largest randomized study for renal cell carcinoma in the adjuvant
setting (17). This trial was also
a randomized, international, multicenter, open-label study. The HSP
vaccine was prepared from surgically removed diseased kidneys. The
patients were at high risk for recurrence following nephrectomy,
therefore, the endpoint was recurrence-free survival. The patients
were randomly distributed in a 1:1 ratio into two groups, the
treatment group, nephrectomy plus HSP vaccine; and the control
group, nephrectomy alone. The results of this phase III trial were
similar to the previously described phase III trial. No difference
was identified in recurrence-free survival between patients who
received vitespen and patients who did not receive treatment.
Specific evidence was identified of an improved recurrence-free
survival with vitespen in patients with an earlier stage of the
disease (AJCC stages 1 and 2), although the difference between the
groups was not statistically significant (P=0.056).
Non-protocol-specified post-hoc analyses confirmed that the
population of patients identified to correlate with the
intermediate-risk category (with stage I/II, high-grade or grade
III and T1/2/3a low-grade disease) had significantly fewer
recurrence events in the vitespen group than in the observation
group (P=0.026). Among the patients at high risk (stage III,
T1/2/3a high-grade, T3b, T3c and stage IV) the differences were
statistically indistinguishable between the vitespen and
observation groups.
From these two randomized phase III clinical trials,
several conclusions may be drawn: i) The HSP vaccine was well
tolerated and any adverse events were generally mild and expected;
ii) the clinical efficacy is associated with the vaccination dose
(increased dose and time of vaccination led to increased
efficiency) and the disease stage (patients with early stage
exhibited apparent benefits in the vaccine treatment group compared
with the control group); and iii) later-stage tumors adopt a number
of mechanisms to subvert the immune response and become resistant
to immunotherapy, offering a potential explanation as to why
vaccine therapy appears to have improved function in earlier-stage
tumors.
6. Mechanisms of immunogenicity of HSP-based
vaccines
The mechanisms by which HSP.PC immunization elicits
potent antitumor effects are becoming clearer. The interaction of
HSP.PC with APCs leads, on the one hand, to the presentation of
antigenic peptides to CD8+ and CD4+ T
lymphocytes (adaptive immunity) and on the other hand, to a cascade
of non-antigen-specific events (innate immunity) that promote
immune responses (Fig. 1) (23).
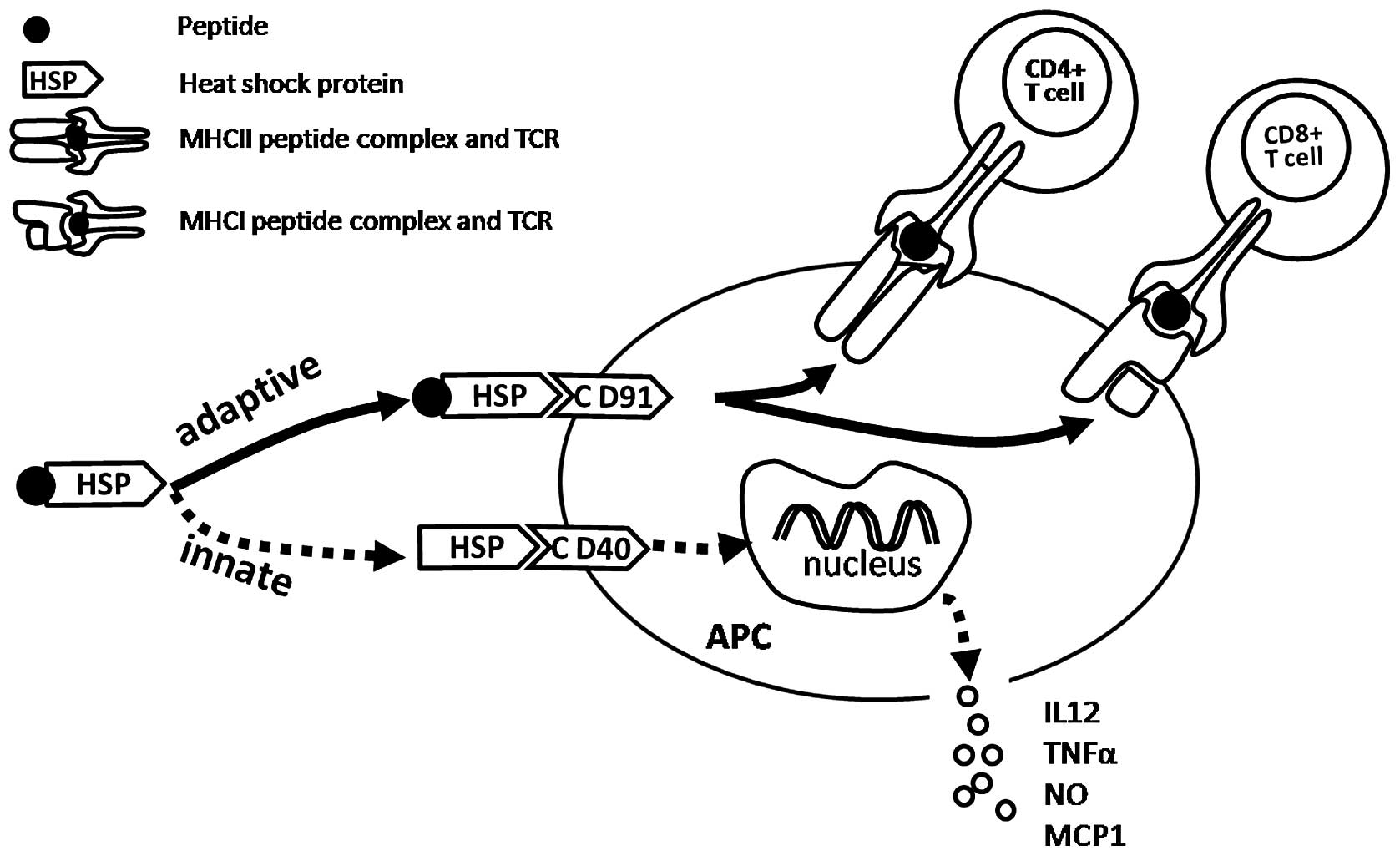 | Figure 1HSP-APC interaction activates
adaptive and innate immune responses. The interaction between the
HSP-peptide complex and CD91 receptor on the APC leads to
stimulation of peptide-specific CD8+ and CD4+
T lymphocyte responses. The interaction of HSP (no peptide complex)
with CD40, CD36, etc., on APCs leads to the non-antigen-specific
innate immune responses, including cytokine and chemokine release
and DC maturation. HSP, heat shock proteins; APC,
antigen-presenting cell; MHC, major histocompatibility complex;
IL-12, interleukin 12; MCP-1, monocyte chemoattractant protein-1;
NO, nitric oxide; TNFα, tumor necrosis factor α; DC, dendritic
cell. |
Activation of adaptive immunity
The fact that immunization with femtomole quantities
of antigenic peptides chaperoned by HSPs is effective in eliciting
such potent T cell responses is noteworthy (46). By way of comparison, tens to
hundreds of micrograms (or tens of nanomoles) of peptides (in a
conventional microbial adjuvant) are typically used to immunize and
elicit a similar response (47,48).
Further investigation was required to analyze how immunization with
femtomoles of peptides is effective. There is a general biological
principle that extraordinary efficiencies are always achieved
through specific receptors. Therefore, researchers hypothesized
that HSPs interact with APCs through specific receptors and that
such interactions result in the endocytosis of HSP.PC, followed by
the processing of peptides and their presentation by MHC molecules
(49). The subsequent
investigations demonstrated this hypothesis and the answer to this
conundrum was identified as CD91. It has been previously confirmed
that HSP90, HSP70, calreticulin and gp96 interact with macrophages
and DCs through a common receptor, CD91 (50,51).
Once HSP.PC is taken up through CD91, it may enter
one or more of several trafficking and processing pathways, which
are likely to lead to the stimulation of various T cell
responses.
The interaction of HSP.PC with CD91 leads to the
internalization of complexes into a non-acidic endosomal
compartment and then the complex or the peptide alone is
transferred to the cytosol (51–53) by
an unknown mechanism. Next, the peptides are processed by the
proteasomes and are transported into the ER by the transporters
associated with antigen processing (TAP) (51). The peptides are then loaded onto MHC
class I molecules. The occupied MHC molecules then pass through the
secretory pathway to the cell surface where they interact with the
receptors of CD8 T cells.
However, following internalization through the CD91
receptor, a small proportion of HSP.PCs enter an acidic
compartment, where the peptide is loaded onto MHC class II
molecules, leading to the stimulation of CD4+ T cells
(54).
Thus, HSP.PC internalized by APCs through CD91
receptors are presented by MHC class I and II molecules, which
stimulate the CD8 and CD4 T cells, respectively. CD91 is the main
receptor involved in this HSP-based immune response, however, there
may be other specific receptors involved, which are currently being
investigated (51).
Activation of innate immunity
It is generally accepted that HSPs, functioning as
chaperones for tumor antigens, elicit tumor-specific adaptive
immune responses. HSPs also appear to induce innate immune
responses in an antigen-independent fashion. Innate responses
generated by HSPs may contribute to antitumor immunity. These
responses include the cytokine and chemokine release by APCs and T
cells and the maturation of DCs. The innate immune responses by the
HSP-APC interaction may be summarized as follows (23): i) secretion of inflammatory
cytokines, tumor necrosis factor α (TNFα), IL-1β, IL-12 and
granulocyte-macrophage colony-stimulating factor (GM-CSF) by
macrophages (55) and NK cells,
which are stimulated by IL-12, were shown to be required for
therapeutic antitumor activity mediated by HSPs (14); ii) secretion of chemokines,
including monocyte chemoattractant protein-1 (MCP-1), macrophage
inflammatory protein-2 (MIP-2) and regulated upon activation,
normal T cells expressed and secreted (RANTES) by T cells (56,57);
iii) induction of nitric oxide (NO) production by macrophages and
DCs and synthesis of inducible NO (58). The production of NO by HSP-activated
APC is likely to have a consequence for the innate control of
tumors and infectious diseases. NO released by HSP-activated APC
may also provide a layer of immunomodulation of Th cells by
necrosis-released HSP. Whereas lower levels of NO are
cytoprotective, higher levels are cytotoxic to T cells,
particularly Th1 cells (58); iv)
maturation of DCs with increased expression of MHC II, B7-2 and
CD40 molecules (59); v) migration
of DCs from the site of injection of HSPs to the draining lymph
nodes. Binder et al(60)
previously confirmed that the immunization of mice with the HSP,
gp96, but not the control proteins, leads to a 5–7 fold enlargement
of the draining lymph nodes. This observation uncovered a novel
aspect of the HSP-APC interaction and adds to the mechanistic
explanation for the unusually high immunogenicity of HSP.PC; and
vi) translocation of nuclear factor-κB (NF-κB) into the nuclei of
macrophages and DCs, which is a key transcription factor involved
in the expression of genes encoding a number of the aforementioned
molecules (55).
The previously described immune responses associated
with innate immune responses induced by HSPs have independent
peptides and play a role in antitumor responses. Previous studies
by Udono and Srivastava and Ciupitu et al confirmed that
prophylactic vaccination with HSPs isolated from normal tissues
cause complete tumor rejection in a small proportion of animals, in
this case, tumor-specific T cells were not expected to be primed
(37,61). Baker-LePain et al
demonstrated that the rate of tumor growth was slowed without
causing complete tumor rejection by HSPs secreted from various
irradiated tumor cells and fibroblast lines, in an
antigen-non-specific manner (62).
In therapeutic settings, tumor-unrelated HSP preparations have been
observed to reduce the metastatic burden and prolong the survival
rate of mice, albeit in a significantly smaller percentage of mice
compared with those treated with tumor-derived HSP preparations
(14).
Collectively, while the innate immune response
elicited by HSPs contributes to tumor immunity, the adaptive immune
responses elicited by HSP-chaperoned tumor-specific peptides are
more important in the antitumor immunity.
7. Limitations of HSP-based vaccines and
alternatives of improvement
Currently, there are >150 medical institutions
undertaking basic and clinical research on HSPs. The two largest
randomized, open-label, multicenter phase III clinical trials
reported in 2008 further confirmed that HSP-based vaccines are
safe, effective and clinically feasible, which inspires further
research. However, these phase III clinical trials also indicated
the limitations of HSP-based vaccines. Firstly, the immunogenicity
is not strong enough, since the efficacy was usually observed in
early-stage disease or with high-dose vaccination, which is
consistent with the results obtained in mice. Secondly, clinical
use of HSP vaccines is limited by the yield of tumor tissue from
the patients and ~50% of patients do not have adequate tumor cells
for the isolation of enough vaccine (16). Therefore, the enhancement of the
clinical effect is urgently required.
Certain alternatives have been tested, including
HSP-pulsed DCs (18), tumor-derived
chaperone-rich cell lysate (CRCL) (63) or a combination with GM-CSF (21,64),
which showed improved immunogenicity. To enhance the
immunogenicity, specific new methods to prepare the vaccine have
been developed, and improved antitumor immune responses have been
observed (65,66). A more powerful HSP-based tumor
vaccine has been developed from DC/tumor fusion cells, which
greatly enhances the immunogenicity of HSP-based vaccines compared
with that derived from the tumor. This new vaccine may present an
improved treatment against malignant cancer (67,68).
Conclusions
Tumor-derived HSP-based vaccines have shown great
prospect in tumor immune therapy. Various animal studies and
clinical trials have demonstrated the efficacy, safety and
feasibility of this vaccine. It is now clear that the
immunogenicity entity was HSP-peptide complexes derived from tumors
and the mechanism was due to activation of adaptive and innate
immunity following the interaction of HSP.PC with APCs through
receptor mediated endocytosis. However, the limitations are also
apparent: i) the immunogenicity is not strong enough and ii) the
yield of tumor vaccine was limited. Certain alternatives to improve
the vaccine have been made and enhanced responses have been
observed. In the future, focus should be on how to improve the
immunogenicity and how to increase the bioavailable efficiency of
this vaccine. Tumor-derived HSP.PC-based vaccines are a promising
vaccination strategy and are likely to provide great help to tumor
patients.
Acknowledgements
The authors would like to thank Dr Jianlin Gong of
Boston University for her assistance and the grant support from the
Nature Science Foundation of China (no. 81000918/H1604).
Abbreviations:
|
HSPs
|
heat shock proteins
|
|
HSP.PC
|
heat shock proteins peptide
complex
|
|
ER
|
endoplasmic reticulum
|
|
APC
|
antigen-presenting cells
|
|
MHC
|
major histocompatibility complex
|
|
NK
|
natural killer
|
|
DC
|
dendritic cell
|
|
CTL
|
cytotoxic T lymphocyte
|
|
TAP
|
transporters associated with antigen
processing
|
|
TNF-α
|
tumor necrosis factor α
|
|
IL-1β
|
interleukin-1β
|
|
GM-CSF
|
granulocyte-macrophage
colony-stimulating factor
|
|
MCP-1
|
monocyte chemoattractant protein-1
|
|
MIP-2
|
macrophage inflammatory protein-2
|
|
RANTES
|
regulated upon activation, normal T
cells expressed and secreted
|
|
NO
|
nitric oxide
|
|
iNOS
|
inducible nitric oxide
|
|
NF-κB
|
nuclear factor-κB
|
|
CRCL
|
chaperone-rich cell lysate
|
References
|
1
|
Ritossa P: Problems of prophylactic
vaccinations of infants. Riv Ist Sieroter Ital. 37:79–108. 1962.(In
Italian).
|
|
2
|
Lindquist S and Craig EA: The heat-shock
proteins. Annu Rev Genet. 22:631–677. 1988. View Article : Google Scholar
|
|
3
|
Georgopoulos C and Welch WJ: Role of the
major heat shock proteins as molecular chaperones. Annu Rev Cell
Biol. 9:601–634. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gething MJ and Sambrook J: Protein folding
in the cell. Nature. 355:33–45. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Parsell DA and Lindquist S: The function
of heat-shock proteins in stress tolerance: degradation and
reactivation of damaged proteins. Annu Rev Genet. 27:437–496. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bukau B, Deuerling E, Pfund C and Craig
EA: Getting newly synthesized proteins into shape. Cell.
101:119–122. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Castelli C, Ciupitu AM, Rini F, et al:
Human heat shock protein 70 peptide complexes specifically activate
antimelanoma T cells. Cancer Res. 61:222–227. 2001.PubMed/NCBI
|
|
8
|
Craig EA, Weissman JS and Horwich AL: Heat
shock proteins and molecular chaperones: mediators of protein
conformation and turnover in the cell. Cell. 78:365–372. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Z and Srivastava PK: Tumor rejection
antigen gp96/grp94 is an ATPase: implications for protein folding
and antigen presentation. EMBO J. 12:3143–3151. 1993.PubMed/NCBI
|
|
10
|
Sherman M and Multhoff G: Heat shock
proteins in cancer. Ann N Y Acad Sci. 1113:192–201. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Milani V, Noessner E, Ghose S, et al: Heat
shock protein 70: role in antigen presentation and immune
stimulation. Int J Hyperthermia. 18:563–575. 2002.PubMed/NCBI
|
|
12
|
Srivastava PK, DeLeo AB and Old LJ: Tumor
rejection antigens of chemically induced sarcomas of inbred mice.
Proc Natl Acad Sci USA. 83:3407–3411. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vanaja DK, Grossmann ME, Celis E and Young
CY: Tumor prevention and antitumor immunity with heat shock protein
70 induced by 15-deoxy-delta12,14-prostaglandin J2 in transgenic
adenocarcinoma of mouse prostate cells. Cancer Res. 60:4714–4718.
2000.PubMed/NCBI
|
|
14
|
Tamura Y, Peng P, Liu K, Daou M and
Srivastava PK: Immunotherapy of tumors with autologous
tumor-derived heat shock protein preparations. Science.
278:117–120. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nicchitta CV: Biochemical, cell biological
and immunological issues surrounding the endoplasmic reticulum
chaperone GRP94/gp96. Curr Opin Immunol. 10:103–109. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Testori A, Richards J, Whitman E, et al:
Phase III comparison of vitespen, an autologous tumor-derived heat
shock protein gp96 peptide complex vaccine, with physician’s choice
of treatment for stage IV melanoma: the C-100-21 Study Group. J
Clin Oncol. 26:955–962. 2008.PubMed/NCBI
|
|
17
|
Wood C, Srivastava P, Bukowski R, et al:
An adjuvant autologous therapeutic vaccine (HSPPC-96; vitespen)
versus observation alone for patients at high risk of recurrence
after nephrectomy for renal cell carcinoma: a multicentre,
open-label, randomised phase III trial. Lancet. 372:145–154. 2008.
View Article : Google Scholar
|
|
18
|
Shinagawa N, Yamazaki K, Tamura Y, et al:
Immunotherapy with dendritic cells pulsed with tumor-derived gp96
against murine lung cancer is effective through immune response of
CD8+ cytotoxic T lymphocytes and natural killer cells. Cancer
Immunol Immunother. 57:165–174. 2008.PubMed/NCBI
|
|
19
|
Sato K, Torimoto Y, Tamura Y, et al:
Immunotherapy using heat-shock protein preparations of leukemia
cells after syngeneic bone marrow transplantation in mice. Blood.
98:1852–1857. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Srivastava PK: Immunotherapy of human
cancer: lessons from mice. Nat Immunol. 1:363–366. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kojima T, Yamazaki K, Tamura Y, et al:
Granulocyte-macrophage colony-stimulating factor gene-transduced
tumor cells combined with tumor-derived gp96 inhibit tumor growth
in mice. Hum Gene Ther. 14:715–728. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kovalchin JT, Murthy AS, Horattas MC,
Guyton DP and Chandawarkar RY: Determinants of efficacy of
immunotherapy with tumor-derived heat shock protein gp96. Cancer
Immun. 1:72001.PubMed/NCBI
|
|
23
|
Srivastava P: Roles of heat-shock proteins
in innate and adaptive immunity. Nat Rev Immunol. 2:185–194. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schmitt E, Gehrmann M, Brunet M, Multhoff
G and Garrido C: Intracellular and extracellular functions of heat
shock proteins: repercussions in cancer therapy. J Leukoc Biol.
81:15–27. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Srivastava P: Interaction of heat shock
proteins with peptides and antigen presenting cells: chaperoning of
the innate and adaptive immune responses. Annu Rev Immunol.
20:395–425. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Berwin B and Nicchitta CV: To find the
road traveled to tumor immunity: the trafficking itineraries of
molecular chaperones in antigen-presenting cells. Traffic.
2:690–697. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schild H and Rammensee HG: gp96 - the
immune system’s Swiss army knife. Nat Immunol. 1:100–101. 2000.
|
|
28
|
Castelli C, Rivoltini L, Rini F, et al:
Heat shock proteins: biological functions and clinical application
as personalized vaccines for human cancer. Cancer Immunol
Immunother. 53:227–233. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Srivastava PK and Das MR: The
serologically unique cell surface antigen of Zajdela ascitic
hepatoma is also its tumor-associated transplantation antigen. Int
J Cancer. 33:417–422. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hoos A and Levey DL: Vaccination with heat
shock protein-peptide complexes: from basic science to clinical
applications. Expert Rev Vaccines. 2:369–379. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gross L: Intradermal immunization of C3H
mice against a sarcoma that originated in an animal of the same
line. Cancer Res. 323–326. 1943.
|
|
32
|
Klein G, Sjogren HO, Klein E and Hellstrom
KE: Demonstration of resistance against methylcholanthrene-induced
sarcomas in the primary autochthonous host. Cancer Res.
20:1561–1572. 1960.PubMed/NCBI
|
|
33
|
Prehn RT and Main JM: Immunity to
methylcholanthrene-induced sarcomas. J Natl Cancer Inst.
18:769–778. 1957.PubMed/NCBI
|
|
34
|
Old LJBE, Clarke DA and Carswell EA:
Antigenic properties of chemically induced tumors. Ann NY Acad Sci.
80–106. 1962.
|
|
35
|
Ullrich SJ, Robinson EA, Law LW,
Willingham M and Appella E: A mouse tumor-specific transplantation
antigen is a heat shock-related protein. Proc Natl Acad Sci USA.
83:3121–3125. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Srivastava PK, Menoret A, Basu S, Binder
RJ and McQuade KL: Heat shock proteins come of age: primitive
functions acquire new roles in an adaptive world. Immunity.
8:657–665. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Udono H and Srivastava PK: Comparison of
tumor-specific immunogenicities of stress-induced proteins gp96,
hsp90 and hsp70. J Immunol. 152:5398–5403. 1994.PubMed/NCBI
|
|
38
|
Srivastava PK, Chen YT and Old LJ:
5′-structural analysis of genes encoding polymorphic antigens of
chemically induced tumors. Proc Natl Acad Sci USA. 84:3807–3811.
1987.
|
|
39
|
Srivastava PK and Maki RG: Stress-induced
proteins in immune response to cancer. Curr Top Microbiol Immunol.
167:109–123. 1991.PubMed/NCBI
|
|
40
|
Zhu X, Zhao X, Burkholder WF, et al:
Structural analysis of substrate binding by the molecular chaperone
DnaK. Science. 272:1606–1614. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bukau B and Horwich AL: The Hsp70 and
Hsp60 chaperone machines. Cell. 92:351–366. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Udono H and Srivastava PK: Heat shock
protein 70-associated peptides elicit specific cancer immunity. J
Exp Med. 178:1391–1396. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Janetzki S, Palla D, Rosenhauer V, Lochs
H, Lewis JJ and Srivastava PK: Immunization of cancer patients with
autologous cancer-derived heat shock protein gp96 preparations: a
pilot study. Int J Cancer. 88:232–238. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rivoltini L, Castelli C, Carrabba M, et
al: Human tumor-derived heat shock protein 96 mediates in vitro
activation and in vivo expansion of melanoma- and colon
carcinoma-specific T cells. J Immunol. 171:3467–3474. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li Z, Qiao Y, Liu B, et al: Combination of
imatinib mesylate with autologous leukocyte-derived heat shock
protein and chronic myelogenous leukemia. Clin Cancer Res.
11:4460–4468. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Blachere NE, Li Z, Chandawarkar RY, et al:
Heat shock protein-peptide complexes, reconstituted in vitro,
elicit peptide-specific cytotoxic T lymphocyte response and tumor
immunity. J Exp Med. 186:1315–1322. 1997. View Article : Google Scholar
|
|
47
|
Martin S, Lappin MB, Kohler J, et al:
Peptide immunization indicates that CD8+ T cells are the dominant
effector cells in trinitrophenyl-specific contact hypersensitivity.
J Invest Dermatol. 115:260–266. 2000.
|
|
48
|
Abiru N, Maniatis AK, Yu L, et al: Peptide
and major histocompatibility complex-specific breaking of humoral
tolerance to native insulin with the B9-23 peptide in
diabetes-prone and normal mice. Diabetes. 50:1274–1281. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Srivastava PK, Udono H, Blachere NE and Li
Z: Heat shock proteins transfer peptides during antigen processing
and CTL priming. Immunogenetics. 39:93–98. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Binder RJ, Han DK and Srivastava PK: CD91:
a receptor for heat shock protein gp96. Nat Immunol. 1:151–155.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Basu S, Binder RJ, Ramalingam T and
Srivastava PK: CD91 is a common receptor for heat shock proteins
gp96, hsp90, hsp70 and calreticulin. Immunity. 14:303–313. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Suto R and Srivastava PK: A mechanism for
the specific immunogenicity of heat shock protein-chaperoned
peptides. Science. 269:1585–1588. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Singh-Jasuja H, Toes RE, Spee P, et al:
Cross-presentation of glycoprotein 96-associated antigens on major
histocompatibility complex class I molecules requires
receptor-mediated endocytosis. J Exp Med. 191:1965–1974. 2000.
View Article : Google Scholar
|
|
54
|
Matsutake T and Srivastava PK: CD91 is
involved in MHC class II presentation of gp96-chaperoned peptides.
Cell Stress Chaperones. 3782000.
|
|
55
|
Basu S, Binder RJ, Suto R, Anderson KM and
Srivastava PK: Necrotic but not apoptotic cell death releases heat
shock proteins, which deliver a partial maturation signal to
dendritic cells and activate the NF-kappa B pathway. Int Immunol.
12:1539–1546. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Moré SH, Breloer M and von Bonin A:
Eukaryotic heat shock proteins as molecular links in innate and
adaptive immune responses: Hsp60-mediated activation of cytotoxic T
cells. Int Immunol. 13:1121–1127. 2001.PubMed/NCBI
|
|
57
|
Lehner T, Bergmeier LA, Wang Y, et al:
Heat shock proteins generate beta-chemokines which function as
innate adjuvants enhancing adaptive immunity. Eur J Immunol.
30:594–603. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Panjwani NN, Popova L and Srivastava PK:
Heat shock proteins gp96 and hsp70 activate the release of nitric
oxide by APCs. J Immunol. 168:2997–3003. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Singh-Jasuja H, Scherer HU, Hilf N, et al:
The heat shock protein gp96 induces maturation of dendritic cells
and down-regulation of its receptor. Eur J Immunol. 30:2211–2215.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Binder RJ, Anderson KM, Basu S and
Srivastava PK: Cutting edge: heat shock protein gp96 induces
maturation and migration of CD11c+ cells in vivo. J Immunol.
165:6029–6035. 2000.PubMed/NCBI
|
|
61
|
Ciupitu AM, Petersson M, Kono K, Charo J
and Kiessling R: Immunization with heat shock protein 70 from
methylcholanthrene-induced sarcomas induces tumor protection
correlating with in vitro T cell responses. Cancer Immunol
Immunother. 51:163–170. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Baker-LePain JC, Sarzotti M, Fields TA, Li
CY and Nicchitta CV: GRP94 (gp96) and GRP94 N-terminal geldanamycin
binding domain elicit tissue nonrestricted tumor suppression. J Exp
Med. 196:1447–1459. 2002. View Article : Google Scholar
|
|
63
|
Li G, Zeng Y, Chen X, et al: Human ovarian
tumour-derived chaperone-rich cell lysate (CRCL) elicits T cell
responses in vitro. Clin Exp Immunol. 148:136–145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Pilla L, Patuzzo R, Rivoltini L, et al: A
phase II trial of vaccination with autologous, tumor-derived
heat-shock protein peptide complexes Gp96, in combination with
GM-CSF and interferon-alpha in metastatic melanoma patients. Cancer
Immunol Immunother. 55:958–968. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Murshid A, Gong J and Calderwood SK:
Purification, preparation and use of chaperone-peptide complexes
for tumor immunotherapy. Methods Mol Biol. 960:209–217. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gao Y, Chen X, Gao W, Yang Y, Ma H and Ren
X: A new purification method for enhancing the immunogenicity of
heat shock protein 70-peptide complexes. Oncol Rep. 28:1977–1983.
2012.PubMed/NCBI
|
|
67
|
Enomoto Y, Bharti A, Khaleque AA, et al:
Enhanced immunogenicity of heat shock protein 70 peptide complexes
from dendritic cell-tumor fusion cells. J Immunol. 177:5946–5955.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Gong J, Zhang Y, Durfee J, et al: A heat
shock protein 70-based vaccine with enhanced immunogenicity for
clinical use. J Immunol. 184:488–496. 2010. View Article : Google Scholar : PubMed/NCBI
|














