Introduction
Natural killer (NK) cells and cytolytic T
lymphocytes (CTLs) function as antitumor immune effectors. NK cells
and CTLs are rich in cytoplasmic granules. Upon degranulation,
these cells release cytotoxic substances that act on target cells
(1). Granules in the cytoplasm of
CTL contain perforin, granzyme, granulysin, additional effector
molecules involved in the antitumor response and several
uncharacterized components (2,3).
In our previous study, an antimicrobial polypeptide
was isolated and purified from interleukin (IL)-2-stimulated human
peripheral blood mononuclear leukocytes and the polypeptide was
identified as high mobility group nucleosomal binding domain 2
(HMGN2). Mononuclear leukocytes were stimulated with IL-2 in
vitro and it was confirmed that HMGN2 is expressed in the
cytoplasm and is secreted. Therefore, HMGN2 may represent a
candidate effector molecule involved in CTL and NK cell function
(4). HMGN2 is one of the most
abundant non-histone nuclear proteins of vertebrates and
invertebrates (5). The HMGN2 gene
is highly conserved and is located near several tumor suppressor
genes (6,7). In addition, the α-helical domain of
HMGN2 homes to tumors, particularly to their vascular endothelia
(8).
In the present study, recombinant human HMGN2
protein was prepared and its antitumor activity was examined in the
oral squamous cell carcinoma cell line, Tca8113.
Materials and methods
Production and isolation of recombinant
human HMGN2 protein
Total RNA was isolated from stimulated mononuclear
leukocytes using TRIzol reagent (Gibco-BRL, Carlsbad, CA, USA). The
full-length HMGN2 cDNA was amplified by reverse
transcription-polymerase chain reaction (RT-PCR) and was ligated
into a pGEX-4T-1 expression vector. The following primers were
designed and prepared: P1, 5′-ACG GAT CCC CCA AGA GAA AGG CTG-3′
and P2, 5′-TAG AAT TCC TTG GCA TCC TCC AGC AC-3′, containing
BamHI and XhoI restriction sites, respectively. The
HMGN2 insert in pGEX-HMGN2 was sequence-verified. Escherichia
coli BL21 (State Key Laboratory of Oral Disease, Sichuan
University, Chengdu, China) were transformed with pGEX-HMGN2. Cells
were cultured in Luria-Bertani medium for 12 h in the presence of
isopropylthio-β-D-galactoside (Sigma-Aldrich, St. Louis, MO, USA)
to induce protein expression. Induced E. coli were washed
with phosphate-buffered saline (PBS) and cell lysates were prepared
with five freeze/thaw cycles in the presence of lysozyme. Following
centrifugation at 10,000 × g for 10 min, glutathione
S-transferase (GST)-HMGN2 fusion proteins were purified from
supernatants using a Glutathione Sepharose 4B column (Amersham
Pharmacia Biotech, Amersham, UK). Purified fusion proteins were
cleaved by thrombin digestion. The recombinant HMGN2 protein was
recovered by reverse-phase high-performance liquid chromatography
(RP-HPLC). Protein concentrations were measured using a
bicinchoninic acid (BCA) protein assay kit (Pierce Biotechnology,
Inc., Rockford, IL, USA) with bovine serum albumin (BSA) as the
standard.
Cell culture
Tca8113 cells were obtained from the State Key
Laboratory of Oral Disease (Sichuan University, Chengdu, China).
Cells were cultured in RPMI-1640 medium (Gibco-BRL) supplemented
with 10% fetal bovine serum (FBS; Gibco-BRL), 100 U/ml penicillin
and streptomycin in a humidified incubator at 37°C with 5%
CO2.
MTT assay
Tca8113 cells were plated in 96-well plates at a
density of 1×104 cells/well. Cells were treated with
various concentrations of HMGN2 protein (0, 1, 2, 3, 4 and 5 μg/ml)
for 48 h. Following this, the medium was replaced in each well with
200 μl fresh medium containing MTT [2.5 mg dissolved in 50 μl
dimethylsulfoxide (DMSO)]. Following incubation for 4 h at 37°C,
the MTT medium in each well was replaced with 100 μl DMSO. Viable
cells were detected by measuring the absorbance at 570 nm.
Apoptosis assay
Tca8113 cells were seeded at a density of
5×105 cells/well in six-well plates and incubated
overnight. Medium was replaced with maintenance medium containing
the appointed concentration of HGMN2 protein and cells were
incubated for 24 h. Cells were harvested by trypsin digestion and
stained using the Annexin V-FITC apoptosis detection kit (R&D
Systems, Minneapolis, MN, USA), according to the manufacturer’s
instructions. Briefly, pretreated Tca8113 cells were washed twice
with FCM buffer (PBS with 5% FBS and 0.1% NaN3). Next,
cells were incubated with Annexin V-FITC for 30 min at 4°C.
Propidium iodide (PI; 50 μg/ml) was added and cells were analyzed
by a Beckman Coulter FC500 with submit 5.2 software (Beckman
Coulter, Miami, FL, USA).
Apoptosis was measured using Hoechst 33258 (Promega
Corporation, Madison, WI, USA), according to the manufacturer’s
instructions. Cells were cultured overnight in six-well plates and
treated with HMGN2 protein for 24 h. Subsequently, cells were
washed with PBS and fixed with a solution of 4% methanol-free
formaldehyde in PBS for 25 min. Staining was performed according to
the manufacturer’s instructions. Fluorescence was visualized using
an Olympus BX60 microscope (Olympus Corporation, Tokyo, Japan).
Cell cycle analysis
Tca8113 cells were seeded at a density of
5×105 cells/well in six-well plates. Cells were
incubated overnight and the medium was replaced with maintenance
medium containing the appointed concentration of HGMN2 protein.
After 24 h, floating and trypsin-harvested cells were combined and
cell cycles were analyzed using PI staining. Briefly, cells were
washed with cold PBS, fixed with cold 70% ethanol and maintained at
4°C overnight. Cells were then washed once with PBS, digested with
200 μl RNase (1 mg/ml) at 37°C for 30 min, and stained with 800 μl
PI (50 μg/ml) at room temperature for 30 min. Cells were analyzed
using a Beckman Coulter FC500 with submit 5.2 software. Cell cycle
histograms were analyzed using MultiCycle for Windows software
(Beckman Coulter).
Western blotting
Total proteins were isolated from cultured cells
using a total protein extraction kit (Nanjing KeyGen Biotech. Co.
Ltd., Nanjing, China) and protein concentrations were measured
using a BCA protein assay kit (Pierce Biotechnology, Inc.).
Proteins were separated by 15% SDS-PAGE and transferred
electrophoretically to polyvinylidene difluoride membranes
(Millipore, Billerica, MA, USA). Membranes were blocked with 2% BSA
in TBS containing 0.1% Tween 20 (TBST) for 2 h at 37°C. Next,
membranes were incubated for 2 h in anti-p53 (polyclonal/rabbit),
anti-Bax (polyclonal/rabbit), anti-Bcl-2 (polyclonal/rabbit),
anti-caspase-3 (polyclonal/rabbit) and anti-GAPDH
(monoclonal/mouse) (all 1:500; Cell Signaling Technology, Inc.,
Beverly, MA, USA). Membranes were subsequently exposed to
horseradish peroxidase-conjugated anti-mouse or -rabbit IgG
secondary antibodies (1:5,000 in TBST with 2% BSA) for 1 h at 37°C.
Proteins were quantified by band densitometry (GS-700; Bio-Rad,
Hercules, CA, USA) using Quantity One 4.4.0 software (Bio-Rad).
In vivo tumor formation assay
Nude mice were randomly divided into three groups of
seven mice each. Tca8113 cells (1×107) were injected
subcutaneously to construct a xenotransplantation tumor model.
HMGN2 (50 ng/g weight) was injected around the tumor tissue on days
21, 25, 29 and 33 following tumor cell transplantation. Identical
doses of cisplatin and PBS were used as positive and negative
controls, respectively. All nude mice were sacrificed at
post-transplantation day 37 and images were captured (Nikon J1,
Wuxi, China). Tumor tissue was removed and the tumor volume was
calculated as follows: Tumor volume = 1/2 × (longer diameter) ×
(shorter diameter)2.
Hematoxylin and eosin (H&E)
staining
Tca8113 xenograft specimens were fixed in 10%
buffered formalin, processed and embedded in paraffin. Sections
(3-μm thick) were cut and stained with H&E. Slides were
visualized using an Imager Z1 microscope equipped with an AxioCam
MRc5 camera (Carl Zeiss AG, Oberkochen, Germany).
Statistical analysis
One-tailed unpaired Student’s t-tests were used to
detect significant differences among treatment groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Purification and characterization of
recombinant human HMGN2 protein
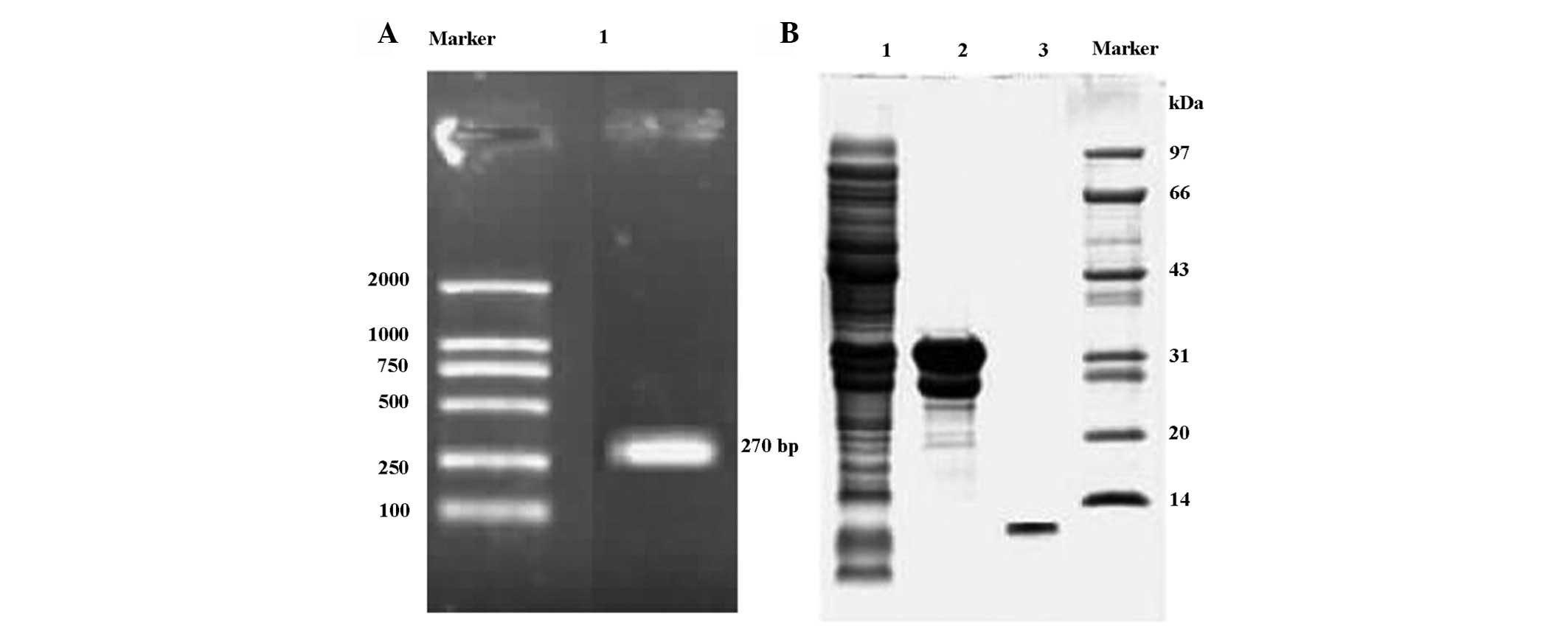
Holo-HMGN2 cDNA was prepared by RT-PCR (Fig. 1A) and used to construct the
prokaryotic expression vector, pGEX-HMGN2. The insert sequence and
orientation of the recombinant vector were confirmed by direct
sequencing. E. coli BL21, transformed with the pGEX-HMGN2
construct, generated HMGN2 fusion proteins in bulk that were
purified by GST affinity chromatography. Purified recombinant HMGN2
was obtained using RP-HPLC (Fig.
1B).
HMGN2 inhibits the growth and colony
formation of Tca8113 cells
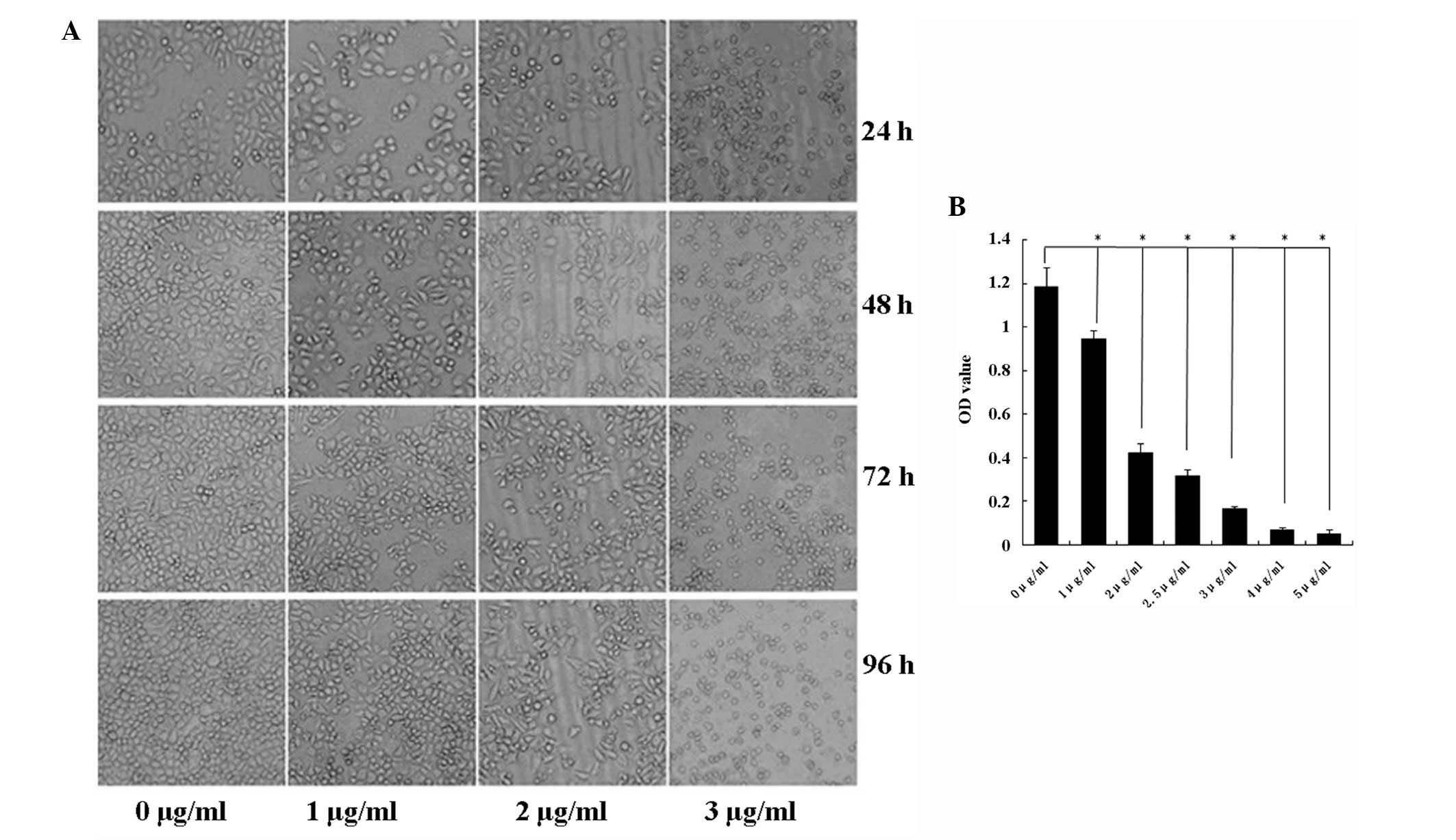
Tca8113 cell growth was suppressed in response to
HMGN2 treatment (Fig. 2A). The MTT
assay was used to assess the toxicity of HMGN2 expression in
Tca8113 cells. At HMGN2 protein concentrations of 1, 2, 3, 4 and 5
μg/ml, Tca8113 cell growth decreased by ~20, 70, 80, 90 and 95%,
respectively (Fig. 2B).
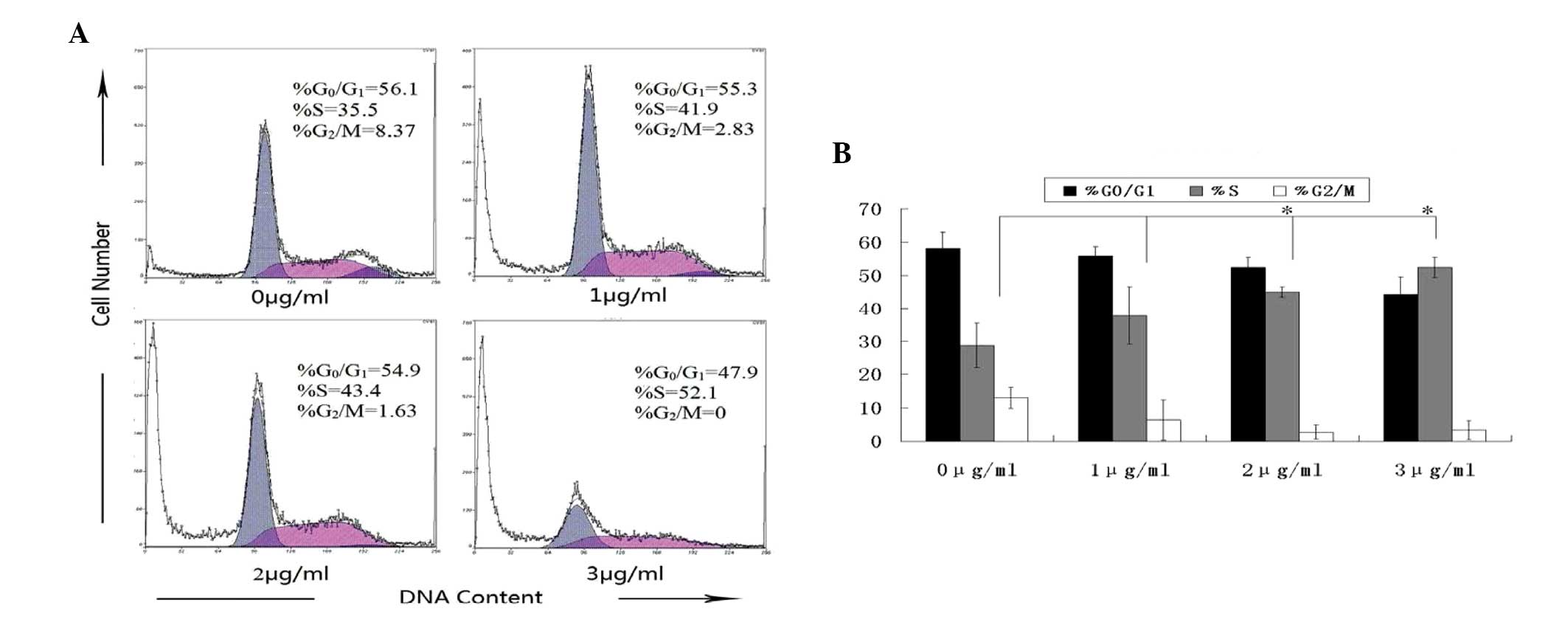
HMGN2 induces S phase cell cycle arrest
in Tca8113 cells
To investigate the potential mechanisms by which
HMGN2 inhibits Tca8113 cell growth, the effect of HMGN2 on the cell
cycle was evaluated by flow cytometry. At 24-h post-treatment, the
percentage of untreated Tca8113 cells in S phase was 35.5%, whereas
the percentage of Tca8113 cells exposed to 3 μg/ml HMGN2 in S phase
was 52.1% (Fig. 3). In addition,
10–15% of untreated Tca8113 cells were in the G2/M phase compared
with 5% of cells treated with 3 μg/ml HMGN2. These results indicate
that HMGN2 treatment may arrest Tca8113 cells in S phase by
inhibiting the S-G2 transition.
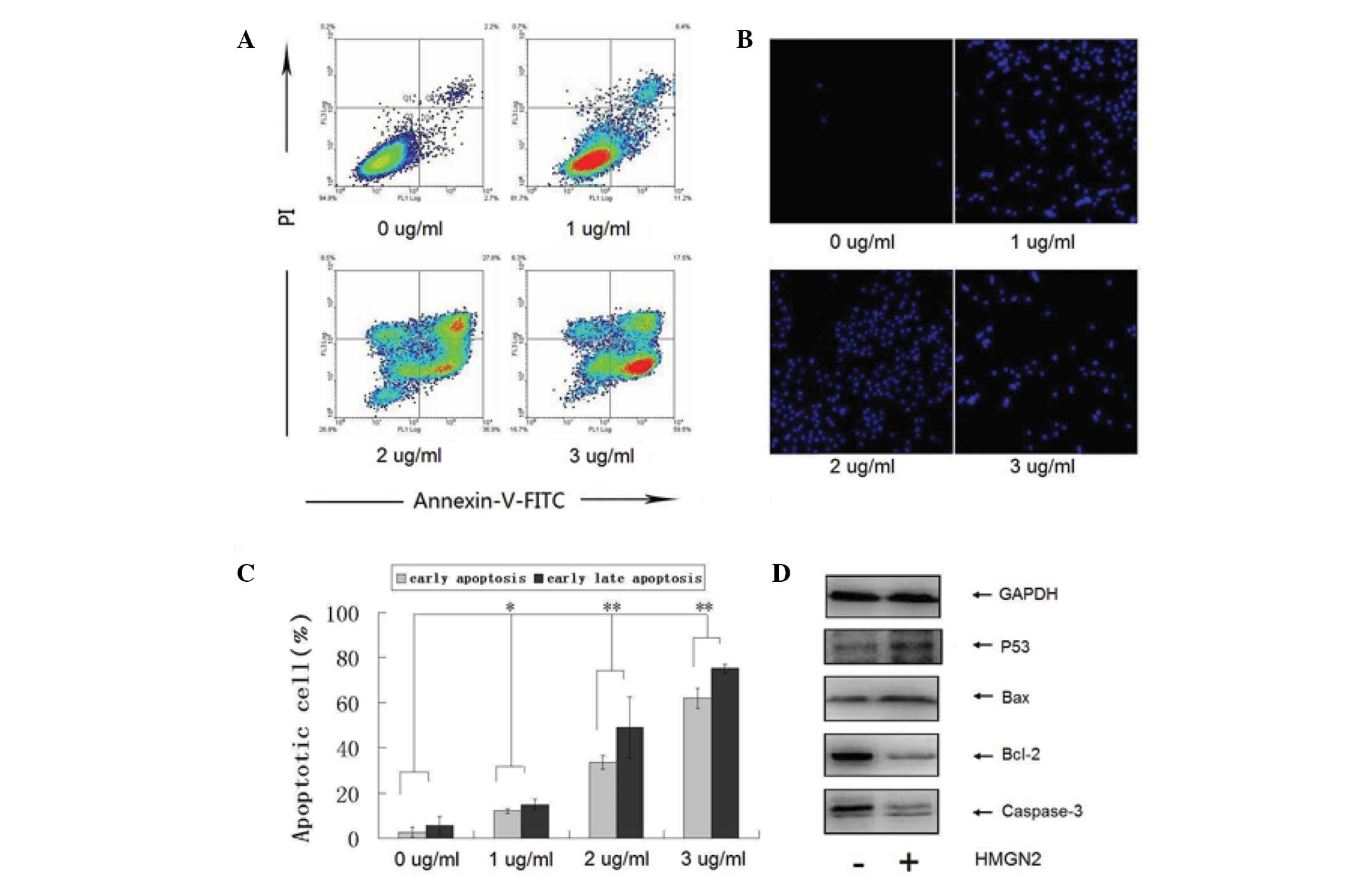
HMGN2 induces apoptosis in Tca8113
cells
To investigate whether the HMGN2-induced growth
inhibition of Tca8113 cells is associated with apoptosis,
HMGN2-exposed cells were analyzed by flow cytometry and fluorescent
microscopy following staining with Annexin V/PI or Hoechst. The
results indicated that HMGN2 induced Tca8113 cell apoptosis in a
dose-dependent manner (Figs. 4A and
B). Compared with untreated cells, the percentage of apoptotic
cells (Annexin V+/PI− and Annexin
V+/PI+) was significantly increased following
exposure to >1 μg/ml HMGN2. The percentage of apoptotic cells
exposed to concentrations of 0, 1, 2 and 3 μg/ml HMGN2 protein were
5, 18, 65 and 77%, respectively. Consistent with the Annexin V/PI
double staining results, the number of Hoechst-positive cells
examined by fluorescence microscopy was also significantly
increased following treatment with >1 μg/ml HMGN2 (Fig. 4C). Next, the effects of HMGN2
treatment on the expression of p53, Bcl-2, Bax and caspase-3 were
examined. When Tca8113 cells were exposed to 2 μg/ml HMGN2 protein
for 24 h, the levels of p53 and Bax proteins were upregulated,
whereas Bcl-2 was significantly downregulated. In addition,
caspase-3 was found to be activated (Fig. 4D).
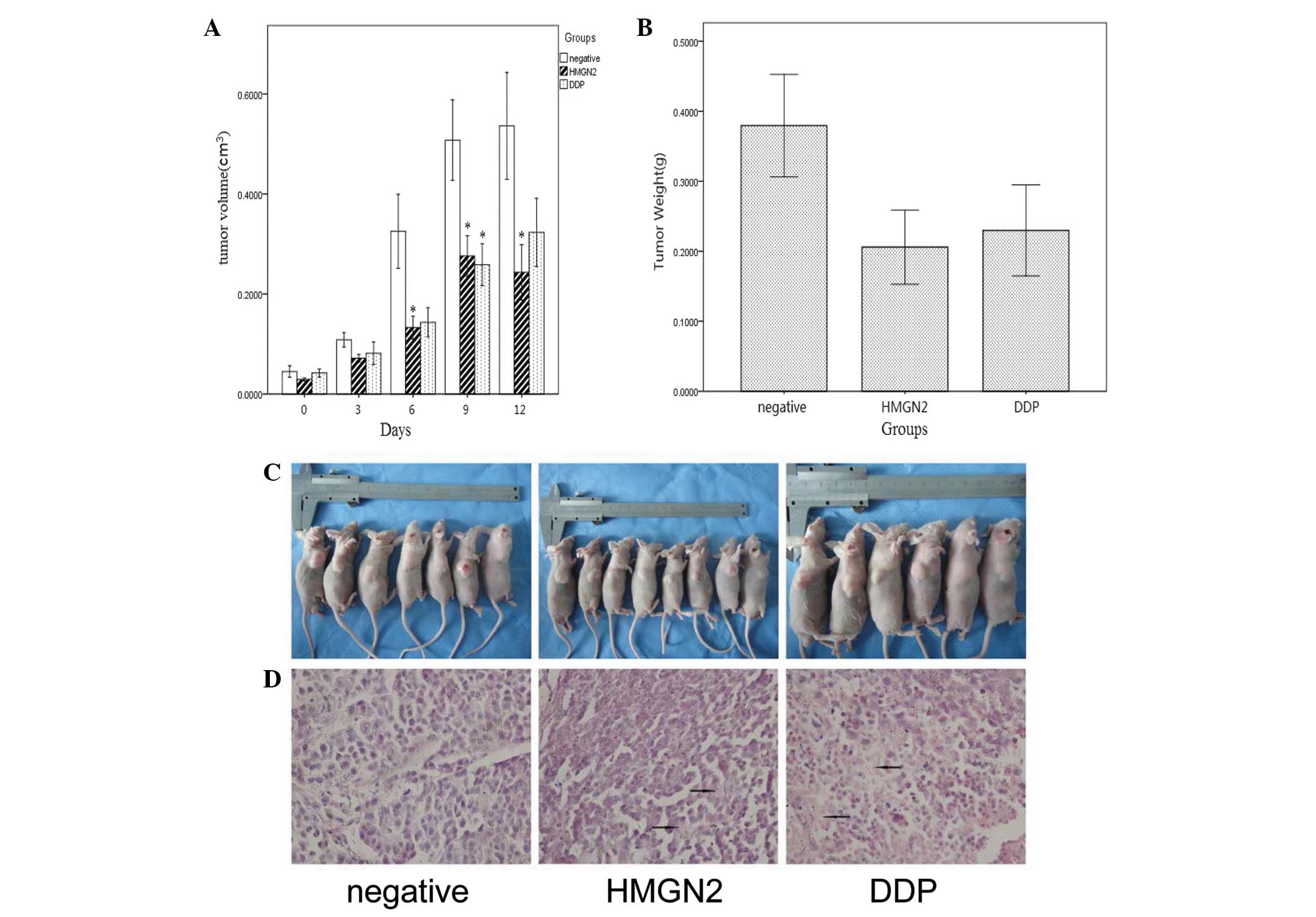
HMGN2 suppresses the growth of Tca8113
cells in vivo
A tumor formation assay was performed to determine
whether HMGN2 is able to affect the growth of Tca8113 cells in
vivo. The growth rate of xenografts in HMGN2-treated groups was
slower compared with that of untreated controls, particularly
during the initial 20 days. A 50% reduction in average tumor volume
was observed in HMGN2-treated tumors compared with controls. Upon
completion of the experiment, the average weight of tumors excised
from HMGN2-treated animals was ~40% of the average control weight
(Fig. 5). H&E staining
indicated that necrosis occurred in the majority of the
HMGN2-treated xenografts during tumor formation.
Discussion
HMG proteins have been described as an abundant
family of non-histone proteins in the cell nucleus of vertebrate
and invertebrate organisms (9). The
HMG protein family is subdivided into three subfamilies: HMGB, HMGA
and HMGN. Each subfamily appears to exert a single characteristic
nuclear function (9), however,
peptides in the HMG protein family also exhibit adjunct roles. For
example, HMG box1 (HMGB1) is an abundant, highly conserved cellular
protein, widely known as a nuclear DNA-binding protein, which
stabilizes nucleosome formation, facilitates gene transcription and
regulates the activity of a steroid hormone receptor (10,11). A
decade-long search has culminated in HMGB1 as a late toxic cytokine
of endotoxemia. HMGB1, released by macrophages upon exposure to
endotoxins, activates a number of other proinflammatory mediators
and is lethal to otherwise healthy animals (10,11).
In addition, HMGB1 exhibits potent bactericidal activity (12). Fernandes et al(13) identified an HMG family peptide in
the mucus secretions of fish skin that also exhibits potent
antimicrobial activity.
The HMGN family includes five chromatin
architectural proteins that are present in higher vertebrates
(14). Of these proteins, HMGN1, 2
and 4 are expressed ubiquitously (15,16),
whereas HMGN3 and 5 are expressed in specific tissues (17,18).
Initially, HMGNs were regarded as transcription coregulators,
however, their roles in DNA repair and cancer progression have been
determined using HMGN1 knockout mice (19). These studies indicate that the
archetype of HMGN1 exhibits characteristics of a tumor suppressor
gene. In addition to HMGN1, the expression of HMGN5 (formerly
NSBP1) (20) was found to be
elevated four-fold in highly metastatic breast cancer cells
compared with that in low metastatic cells (21). In mice, overexpression of HMGN5 in
the uterus was associated with the development of uterine
adenocarcinoma (22). These studies
are consistent with the involvement of HMGN5 in cancer
progression.
The HMGN2 gene is located on chromosome 1p36.1 and
contains six exons (23) with an
extremely high GC content and an ‘HpaII tiny fragment’
island. These hallmarks are indicative of a housekeeping gene that
may be crucial to the basal functioning of cells (7,8).
However, biological roles of this protein have been poorly defined.
HMGN2 is preferentially associated with chromatin subunits
(9), and abnormal HMGN2 gene or
protein expression is associated with neoplasms and autoimmune
diseases (24,25). Porkka et al(8) examined phage-displayed cDNA libraries
in vivo to identify phages capable of homing to the vascular
endothelia of tumors. The screen revealed a markedly potent homing
peptide, F3, which corresponded to a 17- to 48-aa fragment in
HMGN2. The 31-residue peptide selectively bound tumor cells in
vitro and in vivo.
CTL and NK cells are rich in cytoplasmic granules.
Following degranulation, the cells release specific biologically
active substances that are cytotoxic to target cells (1). The granules in the cytoplasm of CTL
contain perforin, granzyme, granulysin and other effector molecules
involved in the antitumor effect, as well as certain unidentified
components (2,3). In our previous study (4), HMGN2 was found to be released by human
peripheral blood mononuclear leukocytes in the presence of IL-2.
HMGN2 may represent an effector molecule for CTL or NK cells.
The present study investigated the activity of the
HMGN2 protein in the oral squamous cell carcinoma line, Tca8113.
HMGN2 protein was demonstrated to inhibit the growth of Tca8113
cells and partially induce apoptosis. Western blotting indicated
the upregulation of p53 and Bax proteins, whereas Bcl-2 was
significantly downregulated. In addition, caspase-3 was found to be
activated and HMGN2 protein is likely to suppress the growth of
Tca8113 cells in vivo. The results indicate that HMGN2
protein exhibits antitumor activity against oral squamous cell
carcinoma and that HMGN2 may represent a candidate effector
molecule for CTL or NK cells. Studies are underway to delineate the
role of HMGN2 as an effector molecule for the antitumor activity of
CTL or NK cells.
Acknowledgements
The present study was supported by grants from the
National Science Funds for Talented Professionals of China (no.
30725041), the National Natural Science Foundation of China (nos.
81372892 and 30972764) and the Changjiang Professorship Support
Program of MOE.
References
|
1
|
Edwards KM, Davis JE, Browne KA, Sutton VR
and Trapani JA: Anti-viral strategies of cytotoxic T lymphocytes
are manifested through a variety of granule-bound pathways of
apoptosis induction. Immunol Cell Biol. 77:76–89. 1999. View Article : Google Scholar
|
|
2
|
Anderson DH, Sawaya MR, Cascio D, Ernst W,
Modlin R, Krensky A and Eisenberg D: Granulysin crystal structure
and a structure-derived lytic mechanism. J Mol Biol. 325:355–365.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kumar J, Okada S, Clayberger C and Krensky
AM: Granulysin: a novel antimicrobial. Expert Opin Investig Drugs.
10:321–329. 2001. View Article : Google Scholar
|
|
4
|
Feng Y, Huang N, Wu Q and Wang B: HMGN2: a
novel antimicrobial effector molecule of human mononuclear
leukocytes? J Leukoc Biol. 78:1136–1141. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Postnikov YV, Herrera JE, Hock R, Scheer U
and Bustin M: Clusters of nucleosomes containing chromosomal
protein HMG-17 in chromatin1. J Mol Biol. 274:454–465. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Srikantha T, Landsman D and Bustin M:
Retropseudogenes for human chromosomal protein HMG-17. J Mol Biol.
197:405–413. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Spieker N, Beitsma M, van Sluis P, Roobeek
I, den Dunnen JT, Speleman F, Caron H and Versteeg R: An integrated
5-Mb physical, genetic, and radiation hybrid map of a 1p36.1 region
implicated in neuroblastoma pathogenesis. Gene Chromosomes Cancer.
27:143–152. 2000. View Article : Google Scholar
|
|
8
|
Porkka K, Laakkonen P, Hoffman JA,
Bernasconi M and Ruoslahti E: A fragment of the HMGN2 protein homes
to the nuclei of tumor cells and tumor endothelial cells in vivo.
Proc Natl Acad Sci USA. 99:7444–7449. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bustin M: Regulation of DNA-dependent
activities by the functional motifs of the high-mobility-group
chromosomal proteins. Mol Cell Biol. 19:5237–5246. 1999.PubMed/NCBI
|
|
10
|
Czura CJ, Wang H and Tracey KJ: Dual roles
for HMGB1: DNA binding and cytokine. J Endotoxin Res. 7:315–321.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang H, Wang H and Tracey KJ: HMG-1
rediscovered as a cytokine. Shock. 15:247–253. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zetterström CK, Strand ML and Söder O: The
high mobility group box chromosomal protein 1 is expressed in the
human and rat testis where it may function as an antibacterial
factor. Hum Reprod. 21:2801–2809. 2006.PubMed/NCBI
|
|
13
|
Fernandes JM, Saint N, Kemp GD and Smith
VJ: Oncorhyncin III: a potent antimicrobial peptide derived from
the non-histone chromosomal protein H6 of rainbow trout,
Oncorhynchus mykiss. Biochem J. 373:621–628. 2003.
View Article : Google Scholar
|
|
14
|
Gerlitz G: HMGNs, DNA repair and cancer.
Biochim Biophys Acta. 1799:80–85. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Birger Y, Ito Y, West KL, Landsman D and
Bustin M: HMGN4, a newly discovered nucleosome-binding protein
encoded by an intronless gene. DNA Cell Biol. 20:257–264. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bustin M and Reeves R: High-mobility-group
chromosomal proteins: architectural components that facilitate
chromatin function. Prog Nucleic Acid Res Mol Biol. 54:35–100.
1996. View Article : Google Scholar
|
|
17
|
West KL, Ito Y, Birger Y, Postnikov Y,
Shirakawa H and Bustin M: HMGN3a and HMGN3b, two protein isoforms
with a tissue-specific expression pattern, expand the cellular
repertoire of nucleosome-binding proteins. J Biol Chem.
276:25959–25969. 2001. View Article : Google Scholar
|
|
18
|
Shirakawa H, Landsman D, Postnikov YV and
Bustin M: NBP-45, a novel nucleosomal binding protein with a
tissue-specific and developmentally regulated expression. J Biol
Chem. 275:6368–6374. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Birger Y, Catez F, Furusawa T, Lim JH,
Prymakowska-Bosak M, West KL, Postnikov YV, Haines DC and Bustin M:
Increased tumorigenicity and sensitivity to ionizing radiation upon
loss of chromosomal protein HMGN1. Cancer Res. 65:6711–6718. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rochman M, Malicet C and Bustin M:
HMGN5/NSBP1: a new member of the HMGN protein family that affects
chromatin structure and function. Biochim Biophys Acta. 1799:86–92.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li DQ, Hou YF, Wu J, Chen Y, Lu JS, Di GH,
Ou ZL, Shen ZZ, Ding J and Shao ZM: Gene expression profile
analysis of an isogenic tumour metastasis model reveals a
functional role for oncogene AF1Q in breast cancer metastasis. Eur
J Cancer. 42:3274–3286. 2006. View Article : Google Scholar
|
|
22
|
Tang WY, Newbold R, Mardilovich K,
Jefferson W, Cheng RY, Medvedovic M and Ho SM: Persistent
hypomethylation in the promoter of nucleosomal binding protein 1
(Nsbp1) correlates with overexpression of Nsbp1 in mouse uteri
neonatally exposed to diethylstilbestrol or genistein.
Endocrinology. 149:5922–5931. 2008. View Article : Google Scholar
|
|
23
|
Popescu N, Landsman D and Bustin M:
Mapping the human gene coding for chromosomal protein HMG-17. Human
Genet. 85:376–378. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Okamura S, Ng CC, Koyama K, Takei Y,
Arakawa H, Monden M and Nakamura Y: Identification of seven genes
regulated by wild-type p53 in a colon cancer cell line carrying a
well-controlled wild-type p53 expression system. Oncol Res.
11:281–285. 1999.
|
|
25
|
Bustin M, Reisch J, Einck L and Klippel
JH: Autoantibodies to nucleosomal proteins: antibodies to HMG-17 in
autoimmune diseases. Science. 215:1245–1247. 1982. View Article : Google Scholar : PubMed/NCBI
|