Introduction
Ovarian cancer is characterized by a high mortality
rate among the gynecological malignancies worldwide (1). To date, although the exact cause of
ovarian cancer remains largely unknown, breast cancer (BRCA)
mutations are the only known hereditary factors. The risk of
ovarian cancer conferred by BRCA mutations has been found to be
regulated by genetic and environmental components (2,3).
Insulin-like growth factor 1 receptor (IGF1R) is a tyrosine kinase
receptor that exerts a direct effect on cell proliferation,
transformation, metastasis, invasion and resistance to anticancer
therapies (4). Recently, IGF1R has
drawn considerable interest in the field of epithelial neoplasms,
in particular, several types of gynecological malignancies,
including ovarian cancer (5).
Notably, emerging evidence has established the following: i) IGF1R
is a potential link between genetic and environmental interactions
(3,6); ii) convergence exists between
IGF1R-related cell proliferation pathways and BRCA1-related tumor
protective pathways (3); and iii)
BRCA1 mutation-related cancer may be regulated by the IGF1R
signaling pathway (3). Therefore,
understanding the complex interrelationship between IGF1R and BRCA1
may improve the current understanding of the basic molecular
mechanism of ovarian cancer. As a result, the present study was
undertaken to investigate IGF1R expression from genetic (BRCA1
mutation) and epigenetic (BRCA1 promoter methylation) aspects in
ovarian cancer and to provide novel insights into the regulatory
mechanism involving IGF1R.
Patients and methods
Patients and tissue collection
The present study was approved by the Institutional
Review Board at the China Medical University (Shenyang, China). In
total, 121 serous ovarian cancer patients (BRCA1 mutation, n=30;
non-BRCA1 mutation, n=32; hypermethylated BRCA1 promoter, n=28; and
non-methylation, n=31) were enrolled between 2010 and 2012, and all
patients provided written informed consent. Fresh tumor samples,
adjacent normal ovarian tissue, ascites and blood samples were
obtained at the time of primary surgery prior to any chemotherapy
or radiotherapy. Hematoxylin and eosin staining of the samples for
histopathological diagnosis and grading were determined by three
pathologists using the World Health Organization criteria (7). All patients were screened for BRCA1
mutations by multiplex polymerase chain reaction (PCR) with
complete sequence analysis as previously reported (8).
Cell culture and lentiviral
infection
Primary ovarian cancer cells were obtained from
ascites of patients undergoing surgery for ovarian cancer and were
cultured in RPMI-1640 with 10% fetal bovine serum (FBS; Invitrogen
Life Technologies, Carlsbad, CA, USA), as described previously
(9). Human 293T and SKOV3 ovarian
cancer cells were maintained in Dulbecco’s modified Eagle’s medium
with 10% FBS (Invitrogen Life Technologies). Lentiviral vectors
expressing short hairpin RNAs against BRCA1 (cat. no. NM_007299)
were obtained from GeneChem Co., Ltd. (Shanghai, China) and
synthesized as follows: Forward, 5′-CcggaaCCTGTCTCCACAAAGTGTGCTCGA
GCACACTTTGTGGAGACAGGTTTTTTTg and reverse,
5′-aattcaaaaaaaCCTGTCTCCACAAAGTGTGCTCGAGCACACTTTGTGGAGACAGGTT. The
non-silencing small interfering RNA sequence (TTCTCCGAACGTGTCACGT)
served as a negative control. For overexpression of BRCA1, the open
reading frame of BRCA1 (cat no. NM_007299) was cloned into the
lentiviral vector GV287 (Ubi-MCS-3FLAG-SV40-EGFP; GeneChem Co.,
Ltd.). Transfections were performed using the polybrene and
enhanced infection solution (GeneChem Co., Ltd.), according to the
manufacturer’s instructions.
Real-time PCR (qPCR) and
immunohistochemistry analysis
qPCR and immunohistochemistry were performed as
previously described (8). The
specific primer sequences used for real-time PCR were as follows:
Forward, 5′-TGATCCTGGATGCGGTGTCCAATA-3′ and reverse,
5′-TGGTCTTCTCACACATCGGCTTCT-3′ for IGF1R; forward,
5′-GGCTATCCTCTCAGAGTGACATTT-3′ and reverse,
5′-GCTTTATCAGGTTATGTTGCATGG-3′ for BRCA1; and forward,
5′-AGGTGAAGGTCGGAGTCA-3′ and reverse, 5′-GGTCATTGATGGCAACAA-3′ for
GAPDH. The primary antibody used for immunohistochemistry was
rabbit anti-IGF1R of human origin (1:200; Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA). Immunostaining was evaluated by two
independent pathologists, blinded to the identity of the subject
groups. Area quantification was performed using a light microscope
(Nikon Eclipse 80i, Nikon Corporation, Tokyo, Japan; magnification,
×400) and analyzed by Image-Pro Plus 6.0 (Media Cybernetics, Inc.,
Rockville, MD, USA). The intensity of the staining was divided into
10 units.
Bisulfite sequencing
Genomic DNA extracted from ovarian cancer and normal
ovarian tissue using a TIANamp Genomic DNA kit (Tiangen Biotech
Co., Ltd., Beijing, China) was subjected to bisulfite conversion
using the EZ DNA Methylation-Direct kit (Zymo Research Corporation,
Irvine, CA, USA) according to the manufacturer’s instructions; the
conversion efficiency was estimated to be ≥99.6%. The DNA was
subsequently amplified by nested PCR and following gel
purification, cloning and transformation into JM109 competent
Escherichia coli cells (Takara Bio, Inc., Shiga, Japan), 10
positive clones of each sample were sequenced to ascertain the
methylation patterns of each CpG locus. The following primers were
used: Forward, 5′-TTGTAGTTTTTTTAAAGAGT-3′ and reverse,
5′-TACTACCTTTACCCAAAACAAAA-3′ for round I; and forward,
5′-GTAGTTTTTTTAAAGAGTTGTA-3′ and reverse,
5′-ACCTTTACCCAAAACAAAAA-3′ for round II. The conditions used were
as follows: 95°C for 2 min; 40 cycles of 30 sec at 95°C, 30 sec at
56°C and 45 sec at 72°C; and 72°C for 7 min.
Statistical analysis
Data are presented as means ± standard deviation.
Statistical differences in the data were evaluated by Student’s
t-test or one-way analysis of variance as appropriate. P<0.05
was considered to indicate a statistically significant
difference.
Results
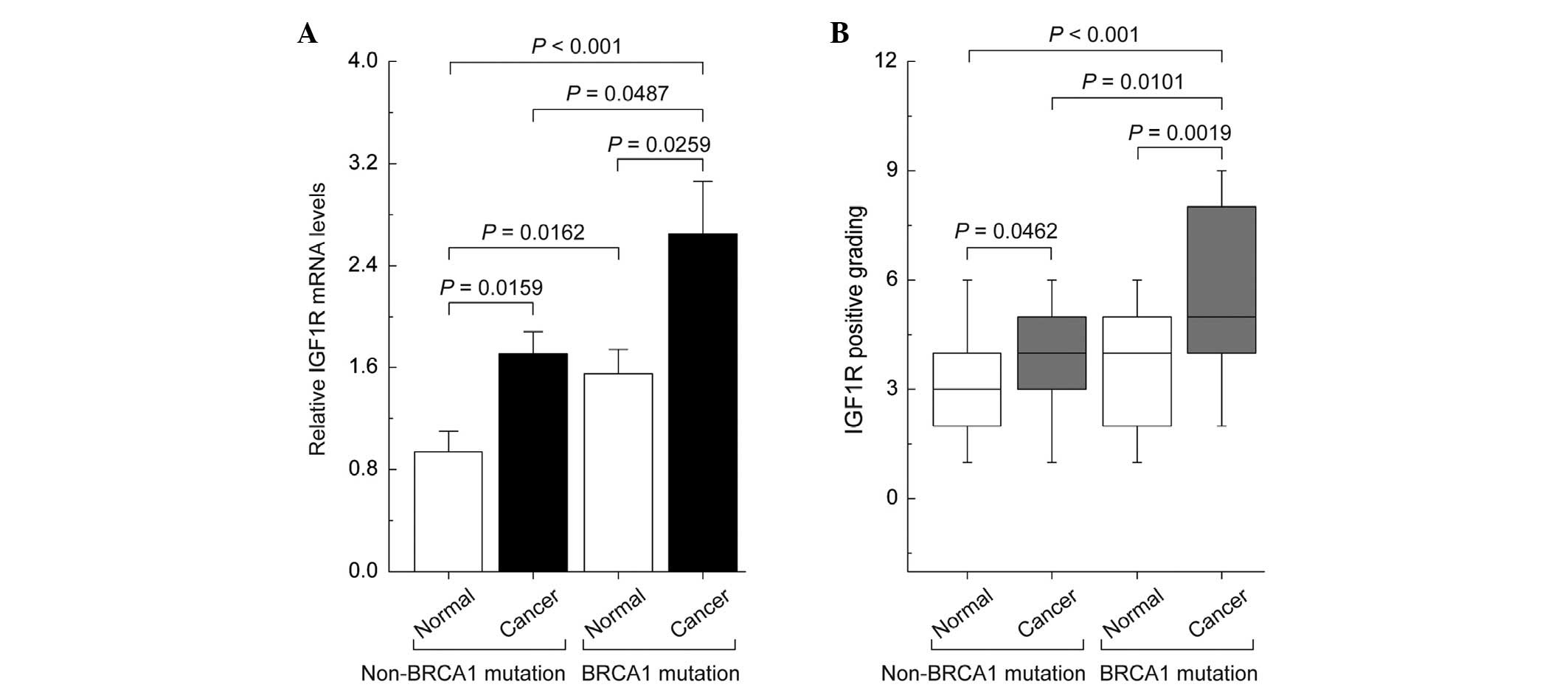
Differences in the expression patterns of
IGF1R in non-mutated and BRCA1-mutated ovarian cancer
qPCR and immunohistochemical analysis showed that
the levels of IGF1R mRNA and protein were increased in non-mutated
and BRCA1-mutated ovarian cancer tissue compared with their
adjacent normal tissue. However, BRCA1-mutated ovarian cancer
markedly increased the expression of IGF1R compared with the
remaining three groups (Fig.
1).
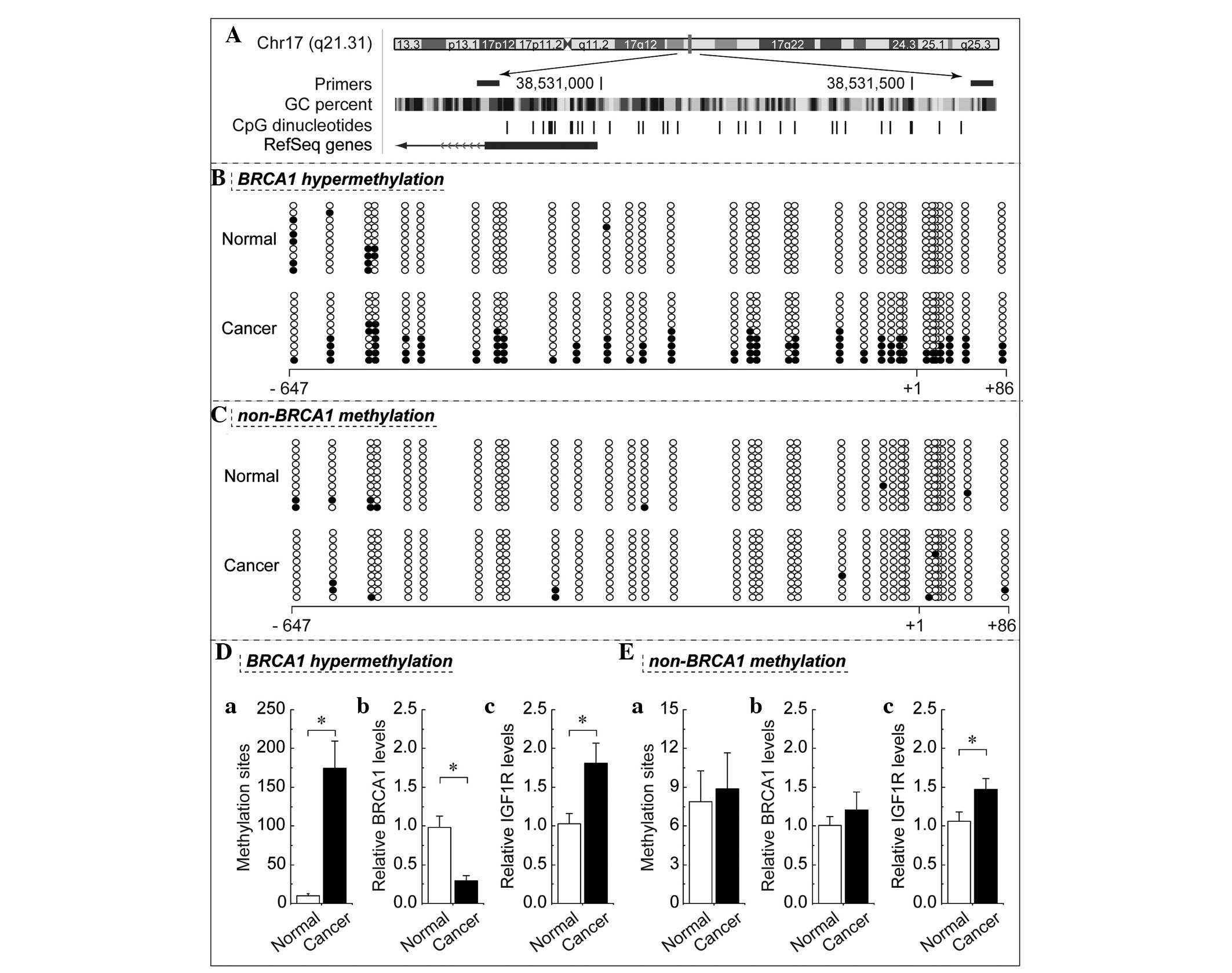
Decreased expression of BRCA1 mediated by
BRCA1 promoter hypermethylation is inversely correlated with IGF1R
levels
In mammals, promoter methylation is an epigenetic
modification involved in regulating gene expression (10). Consistent with this theory, the
present study showed that ovarian cancer tissue with
hypermethylated BRCA1 promoter (Fig. 2B
and Da) exhibited decreased expression of BRCA1 (Fig. 2D) when compared with adjacent normal
tissue. However, no significant differences in BRCA1 expression
(Fig. 2Eb) were observed in ovarian
cancer with unmethylated BRCA1 promoter (Fig. 2C and Ea) as compared with their
adjacent normal tissue. Based on these considerations, the low
levels of BRCA1 appeared to be mediated by promoter
hypermethylation, establishing this an appropriate model to
investigate the physiological correlation between BRCA1 and IGF1R.
Notably, the expression levels of IGF1R were markedly increased
alongside hypermethylated promoter-mediated BRCA1 deficiency in
ovarian cancer (Fig. 2Dc). In
addition, IGF1R expression was also increased in ovarian cancer
tissue (Fig. 2Ec), with no
significant difference identified between BRCA1 promoter
methylation and expression (Fig. 2Ea
and b); however, the increase was not significant when compared
with that observed in ovarian cancer with BRCA1 deficiency. BRCA1
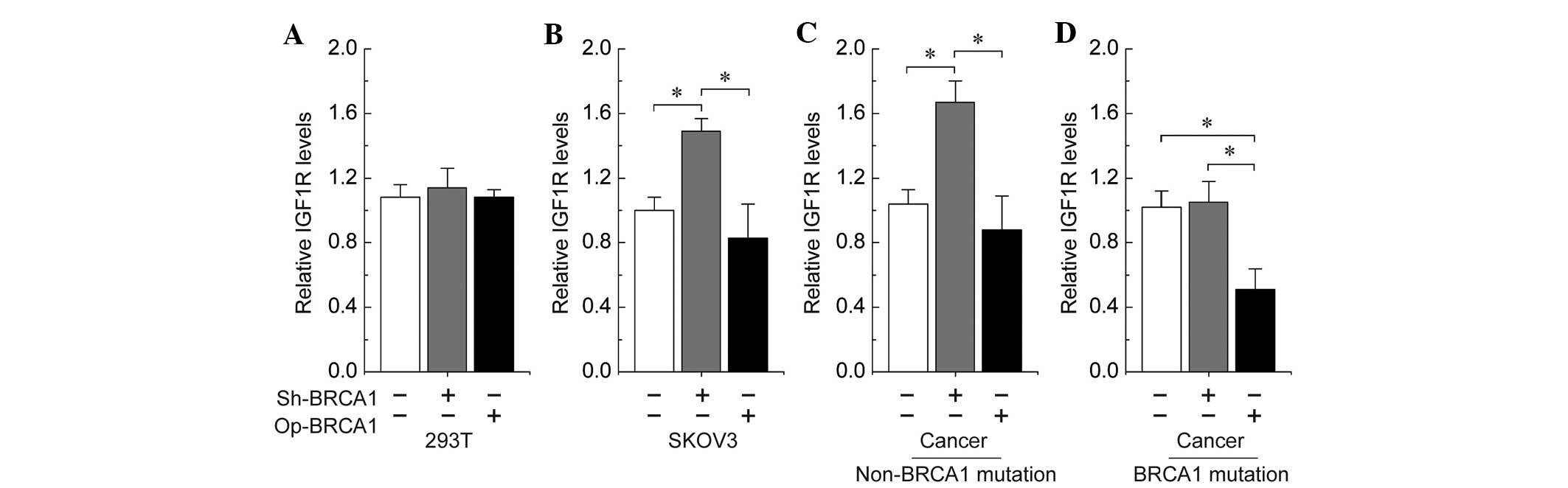
regulates IGF1R expression in ovarian cancer cells. To further
confirm the role of BRCA1 in the regulation of IGF1R, the effects
of overexpression or knockdown of BRCA1 were observed in 293T
cells, the SKOV3 human ovarian cancer cell line, and primary
ovarian cancer cells with and without identified BRCA1 mutations.
The results indicated that no significant changes were identified
in the expression of IGF1R following overexpression or knockdown of
BRCA1 in 293T cells (Fig. 3A).
Notably, the knockdown of BRCA1 was observed to be an effective
method of inducing an increase of IGF1R levels in SKOV3 and
non-BRCA1-mutated ovarian cancer cells (Fig. 3B and C). In addition, the
overexpression of BRCA1 effectively decreased the expression of
IGF1R in BRCA1-mutated ovarian cancer cells (Fig. 3D).
Discussion
The present study reports the following associations
between BRCA1 and IGF1R status in ovarian cancer cells: i) IGF1R
expression is increased in non-BRCA1-mutated ovarian cancer; ii)
BRCA1 inactivation (BRCA1 mutation and promoter hypermethylation)
markedly increases the expression of IGF1R; and iii) BRCA1
knockdown is an effective method to activate the IGF1R gene. These
results indicated that BRCA1 may be a potential regulator of IGF1R
in ovarian cancer, although, a comparable phenomenon has been
observed in breast cancer (11,12).
Notably, the activation effect of BRCA1 was primarily observed in
cells originating from ovarian cancer, however, 293T cells were
insensitive to the overexpression or knockdown of BRCA1. Therefore,
the induced expression of IGF1R may be the result of a complex
interaction of specific factors in ovarian cancer cells. Notably,
numerous previous studies have hypothesized that BRCA1
haploinsufficiency is more likely to result in cancer, due to an
extraordinary ability for clonal growth and proliferation (4). IGF1R is also key in regulating cell
growth and proliferation during cancer development (13). Therefore, it may be predicted that
BRCA1 inactivation-related high levels of IGF1R may be involved in
promoting ovarian cancer progression. An increasing quantity of
data indicates that there is extensive crosstalk between
BRCA1-associated signaling pathways and hormone receptor signaling
pathways. For example, BRCA1 degrades the progesterone receptor
(PR) by counteracting the action of progesterone (14) and inhibits PR activity by preventing
the PR from binding to the progesterone response elements (15). By contrast, progesterone has
previously been reported to induce the downregulation of BRCA1
(16). In addition, BRCA1 represses
estrogen receptor (ER) activity by regulating the phosphoinositide
3-kinase/Akt pathway and the acetylation versus ubiquitination of
ER (17,18). Conversely, zinc finger protein 423
and cathepsin O are involved in estrogen-dependent BRCA1 expression
(19); Nelson et al
(20) observed that IGF1R promotes
the expression and regulates the stability of BRCA1, while the
present study endorsed the theory that the IGF1R gene is a
physiologically relevant downstream target for BRCA1.
In conclusion, the present study emphasizes the
convergence of the IGF1R-mediated cell proliferation pathway and a
BRCA1-mediated antitumor mechanism. However, further clarification
of the complex interactions between the BRCA1 and IGF1R signaling
pathways, at the transcriptional, post-transcriptional and
epigenetic levels, may improve the current understanding of the
basic molecular mechanism of ovarian cancer.
References
|
1
|
Lech A, Daneva T, Pashova S, et al:
Ovarian cancer as a genetic disease. Front Biosci (Landmark Ed).
18:543–563. 2013. View
Article : Google Scholar
|
|
2
|
Pruthi S, Gostout BS and Lindor NM:
Identification and management of women with BRCA mutations or
hereditary predisposition for breast and ovarian cancer. Mayo Clin
Proc. 85:1111–1120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Werner H and Bruchim I: IGF-1 and BRCA1
signalling pathways in familial cancer. Lancet Oncol. 13:e537–e544.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Samani AA, Yakar S, LeRoith D and Brodt P:
The role of the IGF system in cancer growth and metastasis:
overview and recent insights. Endocr Rev. 28:20–47. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bruchim I and Werner H: Targeting IGF-1
signaling pathways in gynecologic malignancies. Expert Opin Ther
Targets. 17:307–320. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pfäffle R, Kiess W and Klammt J:
Downstream insulin-like growth factor. Endocr Dev. 23:42–51.
2012.
|
|
7
|
Scully R: Histological Typing of Ovarian
Tumours. 9. 2nd edition. Springer; Berlin, Germany: 1999,
View Article : Google Scholar
|
|
8
|
Bi FF, Li D and Yang Q: Promoter
hypomethylation, especially around the E26 transformation-specific
motif, and increased expression of poly (ADP-ribose) polymerase 1
in BRCA-mutated serous ovarian cancer. BMC Cancer. 13:902013.
View Article : Google Scholar
|
|
9
|
Szlosarek PW, Grimshaw MJ, Kulbe H, Wilson
JL, Wilbanks GD, Burke F and Balkwill FR: Expression and regulation
of tumor necrosis factor alpha in normal and malignant ovarian
epithelium. Mol Cancer Ther. 5:382–390. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Varley KE, Gertz J, Bowling KM, et al:
Dynamic DNA methylation across diverse human cell lines and
tissues. Genome Res. 23:555–567. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hudelist G, Wagner T, Rosner M, et al:
Intratumoral IGF-I protein expression is selectively upregulated in
breast cancer patients with BRCA1/2 mutations. Endocr Relat Cancer.
14:1053–1062. 2007. View Article : Google Scholar
|
|
12
|
Maor S, Yosepovich A, Papa MZ, Yarden RI,
Mayer D, Friedman E and Werner H: Elevated insulin-like growth
factor-I receptor (IGF-IR) levels in primary breast tumors
associated with BRCA1 mutations. Cancer Lett. 257:236–243. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen X, Meng Q, Zhao Y, et al: Angiotensin
II type 1 receptor antagonists inhibit cell proliferation and
angiogenesis in breast cancer. Cancer Lett. 328:318–324. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calvo V and Beato M: BRCA1 counteracts
progesterone action by ubiquitination leading to progesterone
receptor degradation and epigenetic silencing of target promoters.
Cancer Res. 71:3422–3431. 2011. View Article : Google Scholar
|
|
15
|
Katiyar P, Ma Y, Riegel A, Fan S and Rosen
EM: Mechanism of BRCA1-mediated inhibition of progesterone receptor
transcriptional activity. Mol Endocrinol. 23:1135–1146. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ansquer Y, Legrand A, Bringuier AF, Vadrot
N, Lardeux B, Mandelbrot L and Feldmann G: Progesterone induces
BRCA1 mRNA decrease, cell cycle alterations and apoptosis in the
MCF7 breast cancer cell line. Anticancer Res. 25:243–248.
2005.PubMed/NCBI
|
|
17
|
Ma Y, Fan S, Hu C, et al: BRCA1 regulates
acetylation and ubiquitination of estrogen receptor-alpha. Mol
Endocrinol. 24:76–90. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ma Y, Hu C, Riegel AT, Fan S and Rosen EM:
Growth factor signaling pathways modulate BRCA1 repression of
estrogen receptor-alpha activity. Mol Endocrinol. 21:1905–1923.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ingle JN, Liu M, Wickerham DL, et al:
Selective estrogen receptor modulators and pharmacogenomic
variation in ZNF423 regulation of BRCA1 expression: individualized
breast cancer prevention. Cancer Discov. 3:812–825. 2013.
View Article : Google Scholar
|
|
20
|
Nelson AC, Lyons TR, Young CD, Hansen KC,
Anderson SM and Holt JT: AKT regulates BRCA1 stability in response
to hormone signaling. Mol Cell Endocrinol. 319:129–142. 2010.
View Article : Google Scholar : PubMed/NCBI
|