Introduction
Hepatocellular carcinoma (HCC) is responsible for
over 600,000 mortalities each year; it is the sixth most common
type of cancer in the world and the third greatest cause of cancer
mortality (1–3). HCC prognosis is generally poor and the
5-year survival rate is <7% (4).
Surgical resection and liver transplantation are still recognized
as effective curative approaches for HCC; however, they are
possible in only a small number of patients. Eventually, the
majority of patients exhibit intrahepatic recurrences that quickly
progress to an advanced disease, with blood vessel invasion and
multiple extrahepatic metastases (5). Angiogenesis is a complex process based
on the activation, proliferation and migration of endothelial
cells. During angiogenesis, endothelial cells are activated by
angiogenic factors. The cells then secrete proteases to dissolve
their basement membrane, allowing their migration toward the
angiogenic signal, where they can proliferate and form new blood
vessels (6). Uncontrolled cell
proliferation and angiogenesis play critical roles in HCC growth,
pathological classification, metastatic spread and prognosis
(7).
Chemotherapy is often the only treatment for
advanced and inoperable HCC. However, its outcomes are often
discouraging due to poor tolerance and low efficacy (8). In the recent decade, natural products
have been a rich source of compounds with numerous applications in
cancer therapy, without the associated side effects. For these
reasons, a number of researchers are trying to screen antitumor
compounds from various natural substances. Cordycepin
(3′-deoxyadenosine), is the major bioactive component of
Cordyceps militaris (9) and
is a natural structural analog of adenosine (10). Its pharmacokinetic profile indicates
that the cordycepin-induced metabolite is suppressed by an
adenosine deaminase inhibitor in vivo, and that it has a
short half-life and high rates of clearance (11). This molecule was shown more than 40
years ago to have antitumor activities in rodent and human in
vivo and in vitro systems (12–14).
However, in previous studies, different sources and various
concentrations of purified cordycepin affect the consistency of
these conclusions. In vitro, cordycepin was tested in
various cancer cells, including breast, prostate, colon, leukemia
and lung carcinoma cells (15–20),
as well as in hepatic cancer cells (21–24).
A number of previous studies have assessed the
effects of cordycepin on HCC cells; however, the effects of
cordycepin on vascular endothelial cell migration and angiogenesis
require investigation. Furthermore, no information is available
regarding the intracellular levels of cordycepin following
treatment in endothelial cells. Therefore, the primary aim of the
present study was to assess the antimigration and anti-angiogenic
effects of cordycepin on vascular endothelial cells, and the
stability of intracellular cordycepin levels following
administration.
Materials and methods
Reagents
Professor Li from the South China Normal University
(Guangzhou, China) developed a novel column chromatography
extraction method for the extraction of cordycepin from solid
rice-based fermentation medium. Using this method, cordycepin is
obtained at a 98% purity, with an overall recovery rate of 90%
(25). Cordycepin was provided by
his laboratory and was freshly prepared as stock solution in
double-distilled water. It was diluted in culture medium at
concentrations of 125, 250, 500, 1,000 and 2,000 μg/ml prior to
experiments. Dulbecco’s modified Eagle’s medium (DMEM) and 4′,
6-Diamidino-2-phenylindole (DAPI) were purchased from Invitrogen
Corporation (Carlsbad, CA, USA). Fetal bovine serum (FBS) was
purchased from Gibco Industries Inc. (Big Cabin, OK, USA) and the
cell apoptosis propidium iodide (PI) detection kit was purchased
from Nanjing KeyGen Biotechnology Co., Ltd. (Nanjing, China).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT)
and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich
(St. Louis, MO, USA). MTT was dissolved in phosphate-buffered
saline (PBS; Gibco, Carlsbad, CA, USA) and stored in the dark.
Transwell chamber and Matrigel were purchased from BD Biosciences
(San Jose, CA, USA). All reagents were of analytical grade, unless
otherwise specified.
Cell culture
Human HCC cells (HepG2) and human endothelial-like
immortalized cells (EA.hy926) were obtained from the Cell Bank of
Type Culture Collection of Chinese Academy of Sciences (Shanghai,
China). EA.hy926 and HepG2 cells were cultured in DMEM supplemented
with 10% (v/v) heat-inactivated fetal calf serum, penicillin (100
U/ml) and streptomycin (100 U/ml) (both Sigma-Aldrich). Cultures
were maintained at 37°C in a humidified atmosphere containing 5%
CO2 and 95% air. The medium was changed every two
days.
Cell viability assay
Cell survival changes in response to cordycepin were
evaluated by MTT assay (8,26). Briefly, 2×104 cells in
100 μl DMEM supplemented with 2% (v/v) heat-inactivated FBS,
penicillin (100 U/ml) and streptomycin (100 U/ml) were seeded into
96-well plates. Medium without cells was used as a blank control.
Confluent cells were treated with various concentrations of
cordycepin (125, 250, 500, 1,000 and 2,000 μg/ml) for 1, 2, 3, 4
and 5 days. The same volume of double-distilled water was used as
the negative control (0 μg/ml). At the designed time points, 100 ml
MTT solution in PBS was added to obtain a final concentration of
0.5 g/ml, and the incubation was continued at 37°C for 4 h.
Finally, the medium was removed and replaced with 200 μl DMSO. The
mixture was quantified by determining its absorbance at 540 nm
using a SpectraFluor Plus Reader (Tecan AG, Hombrechtikon,
Switzerland). The relative growth rate was calculated as optical
density (OD)test group/ODnegative
control.
Flow cytometric detection of apoptotic
cells
Assessment of apoptotic cells was performed
according to published methods (27,28).
EA.hy926 cells were exposed to various concentrations of cordycepin
(0, 250, 500, 1,000 and 2,000 μg/ml). They were treated with
trypsin-EDTA (Sigma-Aldrich) and collected by centrifugation at 150
× g for 10 min, then thoroughly rinsed with PBS. Pellets were
resuspended in ice-cold 70% ethanol and fixed at −20°C for 24 h.
Cells were then centrifuged (1,000 rpm for 15 min) and ethanol was
removed by washing thoroughly with PBS. Cell pellets were
resuspended in 1 ml DNA-staining reagent containing 50 μg/ml RNase,
0.1% Triton X-100, 0.1 mmol EDTA (pH 7.4) and 50 μg/ml PI which was
provided with the cell apoptosis PI detection kit. Samples were
stored in the dark at 4°C for 30 min. Red fluorescence (DNA) was
detected through a 563–607 nm band-pass filter using a FC 500
MCL/MPL flow cytometer (Beckman Coulter, Brea, CA, USA). In flow
cytometry histograms, apoptotic cells have a signal in the
sub-diploid regions, which are well-separated from the normal G1
peak. A total of 105 cells in each sample were analyzed
and the percentage of apoptotic cell accumulation in the sub-G1
peak was calculated.
Transwell migration and invasion
assays
A Transwell chamber containing an 8-μm pore
polycarbonate membrane filter was coated either with Matrigel (for
invasion) or without Matrigel (for migration) and inserted in a
24-well culture plate. HepG2 cells were pre-treated with 0, 125,
250, 500 and 1,000 μg/ml cordycepin for 24 h. The cells were then
detached with trypsin-EDTA and resuspended in serum-free DMEM.
After filling the lower chamber with media supplemented with 10%
FBS as a chemoattractant, 105 cells/well in 0.2 ml
serum-free DMEM were loaded in the upper chambers. The apparatus
was incubated at 37°C in a humidified chamber with 5%
CO2 for 12 h (migration assay) or 24 h (invasion assay).
Following incubation, the filter was removed. Cells in the upper
chamber that did not migrate were scraped away with a cotton swab.
The transmembrane cells were fixed in methanol for 30 min, washed
twice with PBS and stained with 300 nM DAPI for 5 min. Migrating or
invading cells were photographed using an inverted microscope (Axio
Observer Z1; Carl Zeiss AG, Oberkochen, Germany) and were counted
in five randomly selected fields per membrane, then the averages
were calculated. Presented data are representative of three
individual wells.
Wound healing assay
EA.hy926 cells were cultured as confluent
monolayers, synchronized in 1% FBS for 24 h and wounded by removing
a 300- to 500 μm-wide strip of cells across the well with a
standard 200 μl pipette tip. Wounded monolayers were washed twice
with PBS to remove non-adherent cells and then treated with 0, 125,
500 and 2,000 μg/ml cordycepin for 6 h. EA.hy926 cell migration was
recorded under inverted microscope (Axio Observer Z1; Carl Zeiss
AG). Wound healing was quantified, using Image J software (National
Institutes of Health, Bethesda, MD, USA), as follows: Wound healing
area (%) = [cell-free area (0 h) - cell-free area (6 h)] /
cell-free area (0 h) ×100 (29).
Tube formation assay for
angiogenesis
To investigate the effect of cordycepin on
angiogenic activity of EA.hy926 cells in vitro, a tube
formation assay was performed following the procedure by Oikawa
et al (30).
Twenty-four-well cluster tissue culture dishes were coated with 500
μg/ml Matrigel and incubated for 30 min at 37°C. EA.hy926 cells
were pre-treated with 0, 125, 250, 500 and 1,000 μg/ml cordycepin
for 12 h and were then seeded onto solidified gels at a density of
105 cells/well in 1 ml culture medium. After 24 h of
incubation, the total lengths of tube-like structures in five
randomly selected microscopic fields per well were determined by
phase-contrast microscopy and quantified using Image J
software.
High-performance liquid chromatography
(HPLC) assay of intracellular cordycepin levels
Intracellular cordycepin levels were measured
according to a previously published method (31). EA.hy926 cells were seeded into
six-well plates at a density of 1.5–2×106 cells/well.
After reaching confluence, cells were pretreated with 125 μg/ml
cordycepin for 0.5–3 h. In order to investigate intracellular
cordycepin levels, the culture medium was removed, the cells were
rinsed three times with PBS and were submitted to two
freeze-and-thaw cycles, then homogenized on ice. The cell
homogenate was centrifuged at 12,000 × g for 15 min at 4°C. The
supernatant was stored on ice and was filtered through a 0.22-μm
filter. The supernatant was finally assayed by HPLC (Dalian Elite
Analytical Instruments Co., Ltd., Dalian, China) with dual P230
pumps, an UV230+ detector and analytical software. Samples were
processed on an YMC-packed C18 column (5 μm, 250×4.6 mm). The
mobile phase consisted of methanol:water (20:80 v/v), with a flow
rate of 1.0 ml/min. The UV detector was set at 260 nm and the
amount of injected sample was 10 μl. Quantitative analysis of
cordycepin was determined by its peak area based on a standard
curve built using 100 μg cordycepin. Cordycepin peaks in the
samples were identified by the retention time and co-injection
tests with the corresponding standard compound. The peak for
cordycepin was shown at a retention time of 8.96 min.
Statistical analysis
All investigations were conducted with at least
three independent experiments, each performed in triplicates. Data
are expressed as the mean ± standard deviation and were evaluated
for statistical significance using one-way analysis of variance
followed by Duncan’s multiple range tests. GraphPad Prism 5.0
(GraphPad Software Inc., San Diego, CA, USA) was used to perform
statistical analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
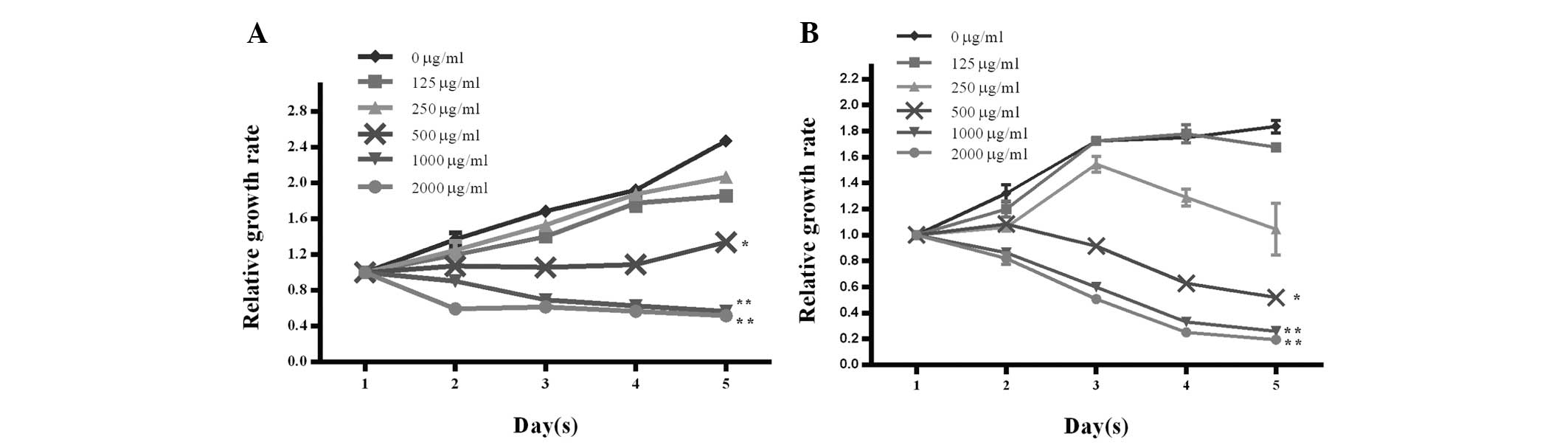
Cordycepin inhibits EA.hy926 and HepG2
cell proliferation
To investigate whether cordycepin affects cell
proliferation in HCC cells, we performed MTT assays in EA.hy926 and
HepG2 cells. As shown in Fig. 1A and
B, the relative growth rates were markedly decreased in the
presence of high doses of cordycepin exceeding 500 μg/ml,
indicating that cordycepin inhibited EA.hy926 and HepG2 cell
proliferation in a dose- and time-dependent manner.
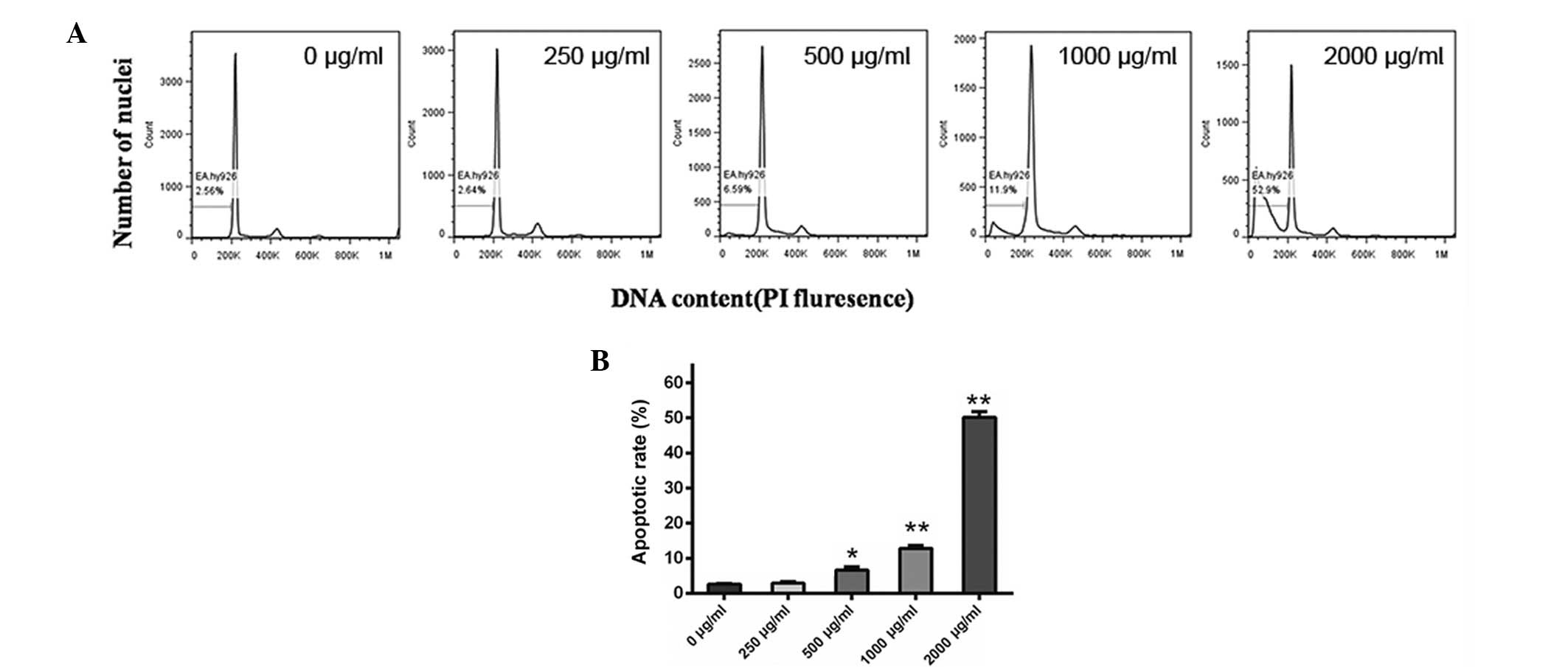
Cordycepin induces EA.hy926 cell
apoptosis
To assess whether cordycepin affects apoptosis of
endothelial cells, we incubated EA.hy926 cells with 0, 250, 500,
1,000 and 2,000 μg/ml cordycepin for 24 h and performed a flow
cytometry assay. As shown in Fig. 2A
and B, 250 μg/ml cordycepin had no detectable effect on cell
apoptosis, while 500, 1,000 and particularly 2,000 μg/ml cordycepin
caused a marked increase in the percentage of apoptotic cells,
compared with the negative control (0 μg/ml) (all P<0.05).
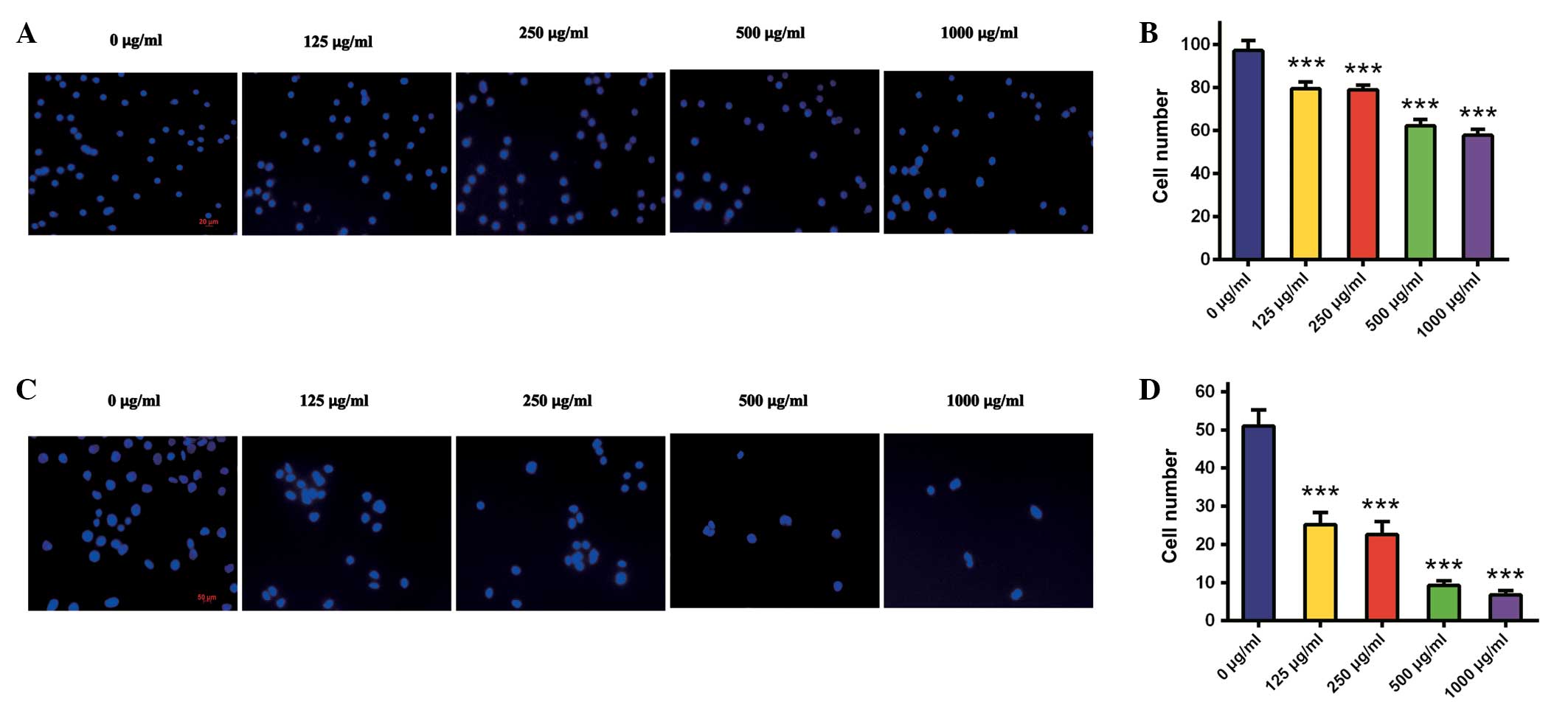
Corcydepin inhibits HepG2 cell migration
and invasion
In HCC development and metastatic spread, cell
migration and invasion are essential processes. To detect antitumor
activities of cordycepin on HepG2 cells, Transwell migration and
invasion assays either without or with precoated Matrigel were
performed following treatment with cordycepin at doses of 0, 125,
250, 500 and 1,000 μg/ml. HepG2 migration was significantly
suppressed by cordycepin in a dose-dependent manner (all P<0.05;
Fig. 3A and B). The numbers of
migrating cells following treatment with cordycepin at doses of 0,
125, 250, 500 and 1,000 μg/ml were 97±5, 80±3, 79±2, 62±3 and 58±3,
respectively.
Similarly, HepG2 cell invasion was markedly
inhibited in a dose-dependent manner (all P<0.05; Fig. 3C and D). The numbers of invading
cells following incubation with 0, 125, 250, 500 and 1,000 μg/ml
cordycepin were 51±4, 25±3, 23±3, 9±1 and 7±1, respectively. These
results demonstrate that cordycepin significantly reduced the
potential of HepG2 cells to migrate and invade.
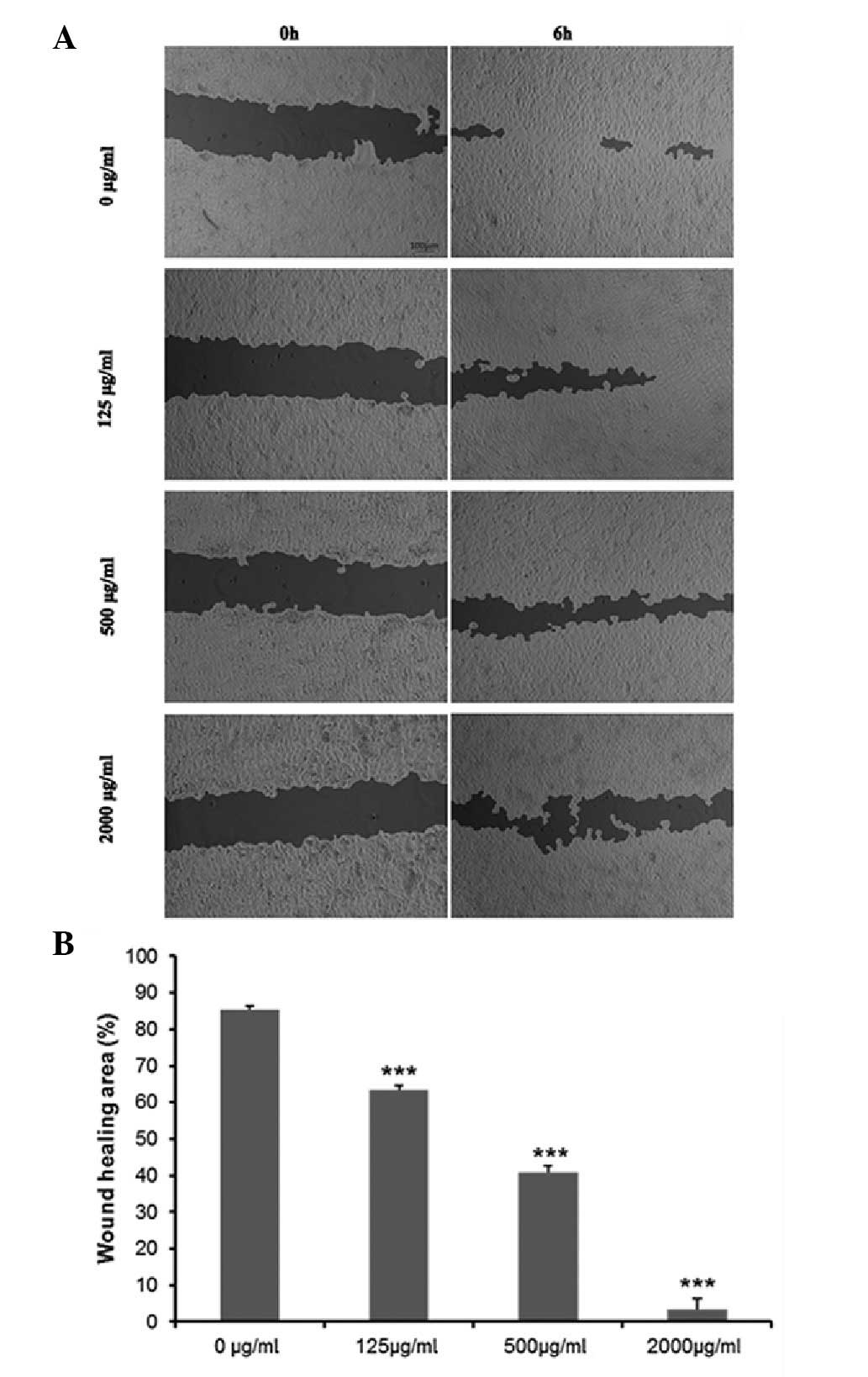
Cordycepin inhibits EA.hy926 cell
migration and angiogenesis
To explore whether cordycepin affects the angiogenic
potential of HCC cells, we examined cell migration and angiogenesis
by wound healing and tube formation assays in EA.hy926 cells. The
wound healing assay demonstrated that the migration of EA.hy926
cells incubated with cordycepin was significantly decreased
compared with the negative control (0 μg/ml) (all P<0.05). After
treatment with various concentrations of cordycepin for 24 h, the
percentage of wound healing area was 85.48±0.84% in the negative
control cells, 63.50±1.08% in 125 μg/ml-treated cells, 40.81±1.76
in 500 μg/ml-treated cells and 3.45±0.29% in 2,000 μg/ml-treated
cells (Fig. 4A and B).
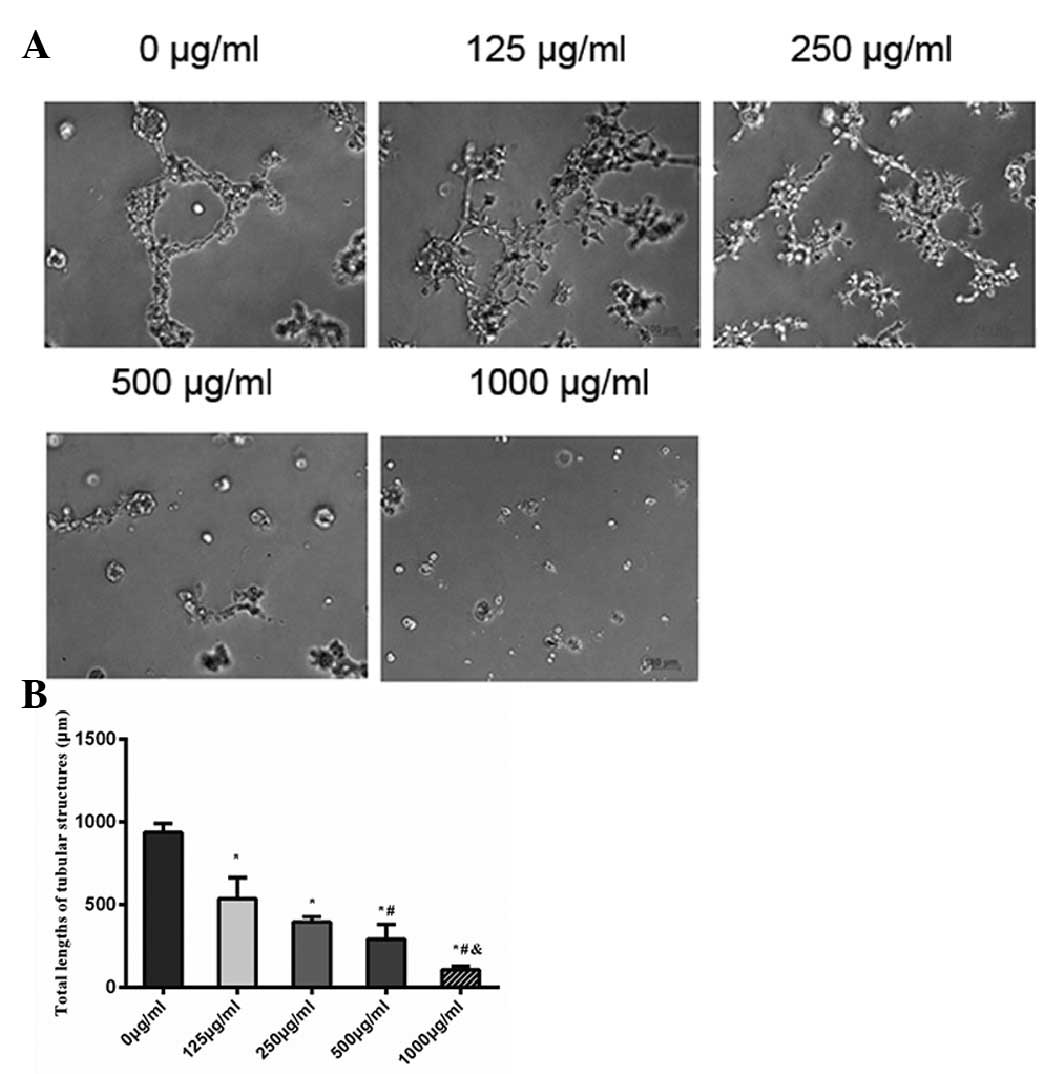
As shown in Fig. 5A,
EA.hy926 cells in the negative control aligned to form tube-like
structures and crossing tubes with multicentric junctions. EA.hy926
cells treated with various concentrations of cordycepin tended to
form fewer tubes, as well as fewer and weaker junctions. The total
lengths of tubular structure after incubation with 0, 125, 250, 500
and 1,000 μg/ml cordycepin for 24 h were 936±56, 536±126, 395±31,
292±88 and 107±39 μm (Fig. 5B). The
results revealed that cordycepin significantly inhibited migration
and angiogenesis of endothelial cells.
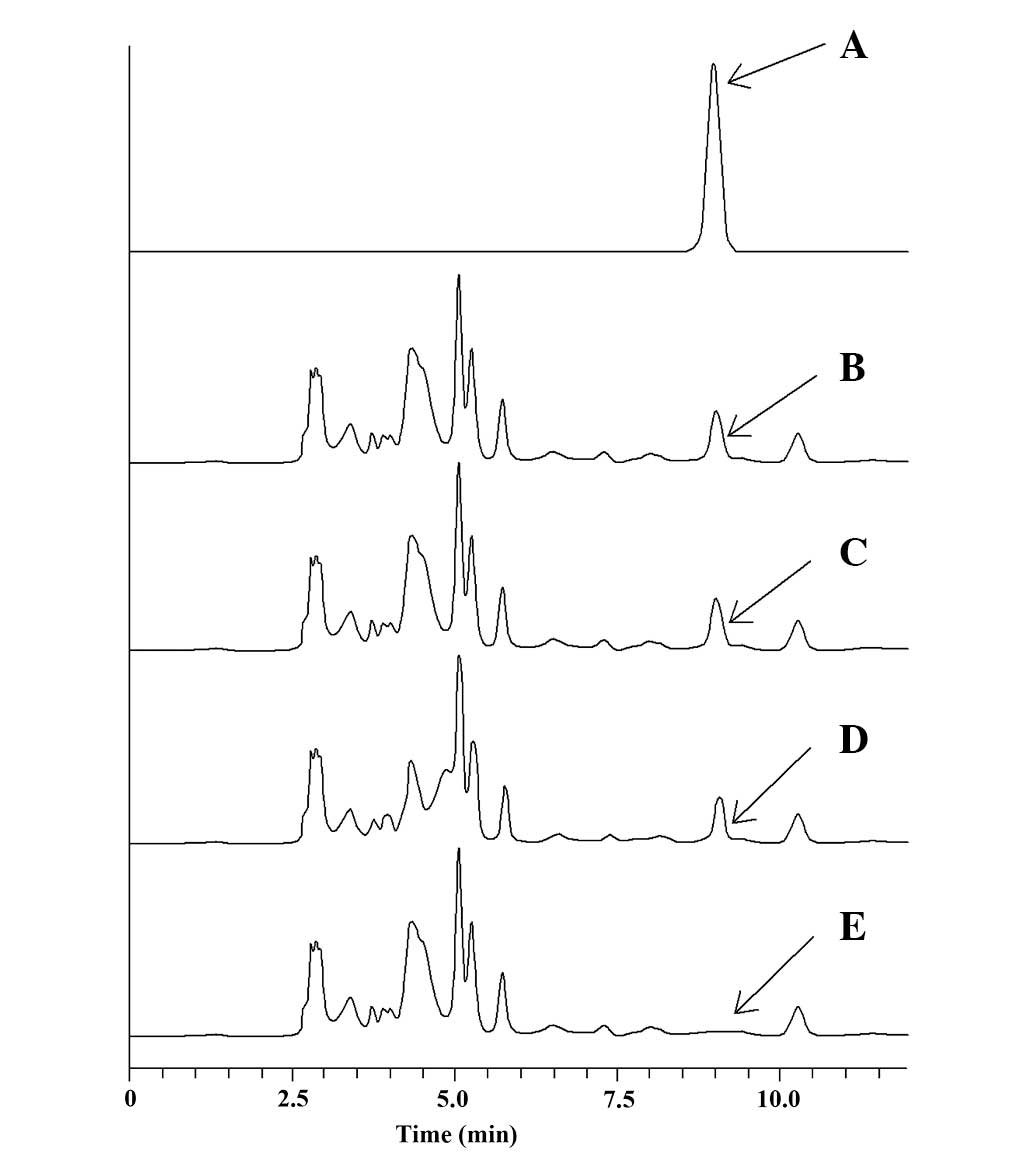
Intracellular cordycepin level was stable
in EA.hy926 cells
Based on the standard curve of cordycepin, the
stability of intracellular cordycepin is shown in Fig. 6A. EA.hy926 cells were incubated with
the culture medium (DMEM plus 10% FBS) containing 125 μg/ml
cordycepin from 0.5 to 3 h (Fig.
6B–D). The cytosol of EA.hy926 cells without cordycepin is
shown in Fig. 6E. The cellular
content was assayed by HPLC. Under the chromatographic conditions
used, cordycepin had a retention time of 8.96 min. The results
demonstrated that cordycepin was able to permeate the cell membrane
of EA.hy926 cells and was stable during the 3 h of incubation.
Discussion
The present study demonstrated that cordycepin
extracted from C. militaris inhibited HepG2 cell
proliferation, migration and invasion. Simultaneously, cordycepin
also inhibited vascular endothelial EA.hy926 cell proliferation,
migration and angiogenesis, and induced apoptosis. Therefore,
cordycepin targeting tumor and endothelial cells may promote the
efficacy of therapy in HCC.
C. militaris, from which cordycepin is
extracted, has long been used in traditional Chinese medicine
(9). Cordycepin exerts numerous
pharmacological actions, such as suppression of cell proliferation,
activation of apoptosis, and inhibition of cell migration and
invasiveness in different tumor cell lines (15,32–35).
Cordycepin reduced metastatic nodule formation in mice (34) and has therefore been proposed as an
antimetastatic agent. The effects of cordycepin are mainly due to
the inhibition of polyadenylation and the activation of
AMP-activated protein kinase in the mTOR signaling pathway, in
doses over 200 μM (24,36). However, only a few reports have
focused on the effects of cordycepin on cell proliferation,
migration and invasion in HCC cells. The ability of HCC cells to
endlessly proliferate is mainly associated with the deregulation of
the cell cycle and promotion of invasion. Previous studies
suggested that cordycepin reduces lipid deposition and cholesterol
levels in HepG2 cells, but has no effect on cell proliferation, and
suggested that cordycepin may have a protective effect on the liver
(37,38). In an additional study, pure
cordycepin at concentrations of 100 μM had no inhibitory effects on
HepG2 cells and no potent in vitro cytotoxicity (39). However, studies performed in other
HCC cell lines, such as BEL-7402 (21), Hep3B (22) and rat H4 (23) showed results similar to those
observed in the present study. Our results also indicated that
cordycepin exerts an anti-invasive cytotoxic action in HepG2 cells,
and that this effect may contribute, at least in part, to the
antimetastatic effect observed in previous studies.
A number of studies have indicated that blood vessel
proliferation in a tumor is a hallmark of tumor growth and
metastatic spread (40,41). HCC tumor vasculature shows irregular
diameter and an abnormal vascular branching pattern; these tumor
vessels also typically lack a complete basal membrane and are
incompletely covered by pericytes and are therefore leaky (7). Cancer cells can spontaneously fuse
with endothelial cells to form hybrid cells, facilitating the
invasion of the endothelial barrier to form metastases (42). Since HCC is a hypervascular tumor,
uncontrolled angiogenesis plays an important role in HCC
development, and thereby anti-angiogenic agents became one of the
most promising therapeutic strategies in HCC (43). In our study, we explored the effect
of cordycepin on angiogenesis of immortalized human umbilical vein
endothelial cells (EA.hy926). These cells are the product of the
fusion between human umbilical vein cells and a
thioguanine-resistant A549 clone. These cells show morphological,
phenotypic and functional characteristics of human endothelial
cells, without the limited lifespan and the inter-donors
variability. These cells are considered good models for cancer drug
screening (44,45). As expected, the results of the
present study demonstrated that cordycepin effectively inhibits
vascular endothelial cell growth and induces apoptosis. Moreover,
it was observed that the anticancer effects of cordycepin are
likely to be associated with inhibition of endothelial cell
migration and tube formation. Therefore, our results suggested that
cordycepin has potential antiangiogenic activity. The effects
observed at low doses may be due to decreased polyadenylation of
mRNAs, while the effects observed at high doses may be due to the
activation of the mTOR pathway (24,36).
A previous study demonstrated that cordycepin exerts
its effects at doses over 200 μM (24). Additionally, pharmacokinetic data
demonstrated that cordycepin has a short half-life and is
metabolized in a short period of time. As an adenosine analog, the
metabolic pathway for cordycepin may be similar to adenosine.
Cordycepin is rapidly deaminated by adenosine deaminase, and is
promptly metabolized to an inactive metabolite,
3′-deoxy-hypoxanthinosine (46,47).
The half-life of cordycepin in rat blood is 1.6±0.0 min after
administration, and the measurable concentration of cordycepin in
rat blood vanishes within 30 min. Cordycepin-induced compounds
appear in the blood and liver for over 2 h after administration
(11). To overcome the problem of
rapid elimination, a high dosage must be administered; otherwise,
intracellular concentrations would be sub-therapeutic. Hence, the
present study used cordycepin at doses ranging from 125 to 2,000
μg/ml, which allowed detection and therapeutic effects. Using a
HPLC method, we showed that cordycepin is able to permeate the
EA.hy926 cell membrane within 0.5 h and was stable for the whole
studied period (3 h). Thus, local administration of high doses of
cordycepin may be sufficiently potent and specific to destroy
enough tumor vasculature to starve the entire tumor.
In conclusion, our results indicate that cordycepin
possesses anticancer properties, which are not solely a result of
direct cytotoxicity in a HCC cellular model, but also of inhibition
of angiogenesis in vascular endothelial cells. Our results also
suggest that a dose greater than 500 μg/ml is required in order to
observe therapeutic effects. Cordycepin may be a potential
anti-angiogenic candidate for cancer therapy in HCC; however, its
mechanisms and adverse effects require further investigation.
Acknowledgements
The authors would like to thank Professor Li (South
China Normal University, Guangzhou, China) for generously providing
cordycepin. This study was supported by grants from the specialized
research fund for the doctoral program of higher education in China
(no. 20120171110073) and the Science and Technology Planning
Project of Guangdong Province (2010B031600222).
References
|
1
|
McGlynn KA and London WT: Epidemiology and
natural history of hepatocellular carcinoma. Best Pract Res Clin
Gastroenterol. 19:3–23. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
3
|
Llovet JM, Burroughs A and Bruix J:
Hepatocellular carcinoma. Lancet. 362:1907–1917. 2003. View Article : Google Scholar
|
|
4
|
Azam F and Koulaouzidis A: Hepatitis B
virus and hepatocarcinogenesis. Ann Hepatol. 7:125–129.
2008.PubMed/NCBI
|
|
5
|
Bosch FX, Ribes J, Diaz M and Cleries R:
Primary liver cancer: worldwide incidence and trends.
Gastroenterology. 127(5 Suppl 1): S5–S16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Flamme I, Frolich T and Risau W: Molecular
mechanisms of vasculogenesis and embryonic angiogenesis. J Cell
Physiol. 173:206–210. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar
|
|
8
|
Lopez PM, Villanueva A and Llovet JM:
Systematic review: evidence-based management of hepatocellular
carcinoma - an updated analysis of randomized controlled trials.
Aliment Pharmacol Ther. 23:1535–1547. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cunningham KG, Manson W, Spring FS and
Hutchinson SA: Cordycepin, a metabolic product isolated from
cultures of Cordyceps militaris (Linn.) Link. Nature.
166:9491950. View
Article : Google Scholar
|
|
10
|
Lennon MB and Suhadolnik RJ: Biosynthesis
of 3′-deoxyadenosine by Cordyceps militaris. Mechanism of
reduction. Biochim Biophys Acta. 425:532–536. 1976.
|
|
11
|
Tsai YJ, Lin LC and Tsai TH:
Pharmacokinetics of adenosine and cordycepin, a bioactive
constituent of Cordyceps sinensis in rat. J Agric Food Chem.
58:4638–4643. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sugar AM and McCaffrey RP: Antifungal
activity of 3′-deoxyadenosine (cordycepin). Antimicrob Agents
Chemother. 42:1424–1427. 1998.
|
|
13
|
Jagger DV, Kredich NM and Guarino AJ:
Inhibition of Ehrlich mouse ascites tumor growth by cordycepin.
Cancer Res. 21:216–220. 1961.PubMed/NCBI
|
|
14
|
Rich MA, Meyers P, Weinbaum G, Cory JG and
Suhadolnik RJ: Inhibition of human tumor cells by cordycepin.
Biochim Biophys Acta. 95:194–204. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakamura K, Yoshikawa N, Yamaguchi Y, et
al: Antitumor effect of cordycepin (3′-deoxyadenosine) on mouse
melanoma and lung carcinoma cells involves adenosine A3 receptor
stimulation. Anticancer Res. 26:43–47. 2006.
|
|
16
|
Wehbe-Janek H, Shi Q and Kearney CM:
Cordycepin/hydroxyurea synergy allows low dosage efficacy of
cordycepin in MOLT-4 leukemia cells. Anticancer Res. 27:3143–3146.
2007.PubMed/NCBI
|
|
17
|
Lee SJ, Kim SK, Choi WS, Kim WJ and Moon
SK: Cordycepin causes p21WAF1-mediated G2/M cell-cycle arrest by
regulating c-Jun N-terminal kinase activation in human bladder
cancer cells. Arch Biochem Biophys. 490:103–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi S, Lim MH, Kim KM, Jeon BH, Song WO
and Kim TW: Cordycepin-induced apoptosis and autophagy in breast
cancer cells are independent of the estrogen receptor. Toxicol Appl
Pharmacol. 257:165–173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee SJ, Moon GS, Jung KH, Kim WJ and Moon
SK: c-Jun N-terminal kinase 1 is required for cordycepin-mediated
induction of G2/M cell-cycle arrest via p21WAF1 expression in human
colon cancer cells. Food Chem Toxicol. 48:277–283. 2010. View Article : Google Scholar
|
|
20
|
Jeong JW, Jin CY, Park C, et al:
Inhibition of migration and invasion of LNCaP human prostate
carcinoma cells by cordycepin through inactivation of Akt. Int J
Oncol. 40:1697–1704. 2012.
|
|
21
|
Shi P, Huang Z, Tan X and Chen G:
Proteomic detection of changes in protein expression induced by
cordycepin in human hepatocellular carcinoma BEL-7402 cells.
Methods Find Exp Clin Pharmacol. 30:347–353. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang BJ, Won SJ, Yu ZR and Su CL: Free
radical scavenging and apoptotic effects of Cordyceps
sinensis fractionated by supercritical carbon dioxide. Food
Chem Toxicol. 43:543–552. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Robertson JB, Williams JR and Little JB:
Enhancement of radiation killing of cultured mammalian cells by
cordycepin. Int J Radiat Biol Relat Stud Phys Chem Med. 34:417–429.
1978. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wong YY, Moon A, Duffin R, et al:
Cordycepin inhibits protein synthesis and cell adhesion through
effects on signal transduction. J Biol Chem. 285:2610–2621. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ni H, Zhou XH, Li HH and Huang WF: Column
chromatographic extraction and preparation of cordycepin from
Cordyceps militaris waster medium. J Chromatogr B Analyt
Technol Biomed Life Sci. 877:2135–2141. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hansen MB, Nielsen SE and Berg K:
Re-examination and further development of a precise and rapid dye
method for measuring cell growth/cell kill. J Immunol Methods.
119:203–210. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Telford WG, King LE and Fraker PJ:
Evaluation of glucocorticoid-induced DNA fragmentation in mouse
thymocytes by flow cytometry. Cell Prolif. 24:447–459. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xing YN, Liang HW, Zhao L and Xu HM: The
antitumor activity of exogenous and endogenous canstatin on
colorectal cancer cells. Asian Pac J Cancer Prev. 12:2713–2716.
2011.PubMed/NCBI
|
|
29
|
Hu J and Verkman AS: Increased migration
and metastatic potential of tumor cells expressing aquaporin water
channels. FASEB J. 20:1892–1894. 2006. View Article : Google Scholar
|
|
30
|
Oikawa T, Sasaki T, Nakamura M, et al: The
proteasome is involved in angiogenesis. Biochem Biophys Res Commun.
246:243–248. 1998. View Article : Google Scholar
|
|
31
|
Liu Z, Tao X, Zhang C, Lu Y and Wei D:
Protective effects of hyperoside (quercetin-3-o-galactoside) to
PC12 cells against cytotoxicity induced by hydrogen peroxide and
tert-butyl hydroperoxide. Biomed Pharmacother. 59:481–490. 2005.
View Article : Google Scholar
|
|
32
|
Chang W, Lim S, Song H, Song BW, Kim HJ,
Cha MJ, Sung JM, Kim TW and Hwang KC: Cordycepin inhibits vascular
smooth muscle cell proliferation. Eur J Pharmacol. 597:64–69. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen LS, Du-Cuny L, Vethantham V, et al:
Chain termination and inhibition of mammalian poly(A) polymerase by
modified ATP analogues. Biochem Pharmacol. 79:669–677. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nakamura K, Konoha K, Yoshikawa N,
Yamaguchi Y, Kagota S, Shinozuka K and Kunitomo M: Effect of
cordycepin (3′-deoxyadenosine) on hematogenic lung metastatic model
mice. In Vivo. 19:137–141. 2005.
|
|
35
|
Wu WC, Hsiao JR, Lian YY, Lin CY and Huang
BM: The apoptotic effect of cordycepin on human OEC-M1 oral cancer
cell line. Cancer Chemother Pharmacol. 60:103–111. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Thomadaki H, Scorilas A, Tsiapalis CM and
Havredaki M: The role of cordycepin in cancer treatment via
induction or inhibition of apoptosis: implication of
polyadenylation in a cell type specific manner. Cancer Chemother
Pharmacol. 61:251–265. 2008. View Article : Google Scholar
|
|
37
|
Lee CW, Wong LL, Tse EY, et al: AMPK
promotes p53 acetylation via phosphorylation and inactivation of
SIRT1 in liver cancer cells. Cancer Res. 72:4394–4404. 2012.
View Article : Google Scholar
|
|
38
|
Guo P, Kai Q, Gao J, et al: Cordycepin
prevents hyperlipidemia in hamsters fed a high-fat diet via
activation of AMP-activated protein kinase. J Pharmacol Sci.
113:395–403. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu JY, Zhang QX and Leung PH: Inhibitory
effects of ethyl acetate extract of Cordyceps sinensis
mycelium on various cancer cells in culture and B16 melanoma in
C57BL/6 mice. Phytomedicine. 14:43–49. 2007.PubMed/NCBI
|
|
40
|
Zetter BR: Angiogenesis and tumor
metastasis. Annu Rev Med. 49:407–424. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lu ZJ, Ren YQ, Wang GP, et al: Biological
behaviors and proteomics analysis of hybrid cell line EAhy926 and
its parent cell line A549. J Exp Clin Cancer Res. 28:162009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nelson NJ: Angiogenesis research is on
fast forward. J Natl Cancer Inst. 91:820–822. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Edgell CJ, Haizlip JE, Bagnell CR, et al:
Endothelium specific Weibel-Palade bodies in a continuous human
cell line, EA.hy926. In Vitro Cell Dev Biol. 26:1167–1172. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Edgell CJ, McDonald CC and Graham JB:
Permanent cell line expressing human factor VIII-related antigen
established by hybridization. Proc Natl Acad Sci USA. 80:3734–3737.
1983. View Article : Google Scholar
|
|
46
|
Adamson RH, Zaharevitz DW and Johns DG:
Enhancement of the biological activity of adenosine analogs by the
adenosine deaminase inhibitor 2′-deoxycoformycin. Pharmacology.
15:84–89. 1977.
|
|
47
|
Farthing D, Sica D, Gehr T, et al: An HPLC
method for determination of inosine and hypoxanthine in human
plasma from healthy volunteers and patients presenting with
potential acute cardiac ischemia. J Chromatogr B Analyt Technol
Biomed Life Sci. 854:158–164. 2007. View Article : Google Scholar
|