Introduction
Gastric cancer is the most common malignancy that
occurs in the stomach mucosa, ranking first in incidence for
malignant tumors of the digestive tract. Gastric cancer is among
the five most common cancers in the world and is the second most
prevalent cause of cancer-related mortality (1). Although it has been shown that the
incidence of gastric cancer is now following a decreasing trend,
the mortality rate and incidence of gastric cancer remain high in
the majority of developing countries (2). The occurrence and development of
gastric cancer are known to involve the imbalanced expression of
proto-oncogenes and oncogenes, which are post-transcriptionally
controlled by microRNA (miR/miRNA) (3,4).
Therefore, the misexpression of miRNAs may affect the development
of the gastric cancer by up- or downregulating the target genes.
miR-449 may affect proliferation, differentiation, apoptosis and
the cell cycle by regulating those target genes directly or
indirectly (5). The
cyclin-dependent kinase (CDK)6 protein is a regulator of the cell
cycle, which can lead to cell cycle disorders, and is closely
associated with cancer occurrence and development.
In the present study the association between
miR-449a, the CDK6 protein and gastric carcinoma are identified. An
miR-449a expression group is also established in order to observe
how miR-449a affects gastric carcinoma cells, and the ability of
miR-449a to regulate the expression of the CDK6 protein is
discussed.
Materials and methods
Subject investigated and samples
A total of 66 patients, who were diagnosed with
gastric cancer in the Department of Gastroenterology of Guangzhou
General Hospital of Guangzhou Military Command (Guangdong, China)
between January 2011 and October 2012, were enrolled in the present
study. Gastric carcinoma and tumor-adjacent normal tissue samples
were obtained during the surgery for the carcinoma of the stomach,
and stored at −80°C in an ultra low temperature freezer (Samsung
Electronics Co., Gyeonggi-do, Korea) All patients had not received
any drug or chemotherapy treatments prior to surgery, and there was
no significant difference in age or gender. The study was approved
by the ethics committee of Guangzhou General Hospital of Guangzhou
Military Command (Guangzhou, China). Patients provided written
informed consent.
Main experimental material
Lipofectamine 2000 transfection reagent was
purchased from Invitrogen Life Technologies (Carlsbad, CA, USA),
while TRIzol reagent, Takara reverse transcription kit, Takara
real-time PCR kit, Dulbecco’s modified Eagle’s medium (DMEM; high
glucose), fetal bovine serum (FBS), trypsin-EDTA, miR-449a reverse
transcription and PCR primer, U6snRNA reverse transcription and PCR
primer, has-miR-449a mini and has-miR-449a inhibitor were purchased
from Guangzhou RiboBio Co., Ltd. (Guangzhou, Guangdong, China).
Mouse monoclonal anti-rabbit CDK6 and rabbit monoclonal anti-human
β-actin rhesus antibodies were purchased from Abcam Co. (Cambridge,
UK), and E-Plate 16 was obtained from Roche (Basel, Switzerland).
The MGC-803 cell line was supplied by the Department of Medical
Experiments, Guangzhou General Hospital of Guangzhou Military
Command.
Model construction of the differential
expression of miR-449a
The MGC-803 cells were cultured using DMEM with 10%
FBS. Subsequent to the resuscitation of the frozen cells, the cells
were provided with a 37°C, 5% CO2 environment. The cells
in the logarithmic phase were used for the experiments. Prior to
the experiments, 5 μl Lipofectamine 2000 was blended with 10 μl
has-miR-449a mimic (100 nM) to prepare the has-miR-449a
minic-Lipofectamine 2000 transfection reagent intermixture (termed
HML). In addition, 5 μl Lipofectamine 2000 was belended with 10 μl
has-miR-449a inhibitor (100 nM) to prepare the has-miR-449a
inhibitor-Lipofectamine 2000 transfection reagent intermixture
(termed HIL). The MGC803 cells were inoculated in cell culture
plates (6-well plates), and HML was added to establish the miR-449a
upregulation group, while HIL was added to establish the miR-449a
downregulation group.
Detection of miR-449a by quantitative
(q)PCR
In total, 1 μl RNA template (500–550 ng/μl), 4 μl
miR-449a RT Primer (62.5 nM) and 14 μl RNase-free H2O
(Takara Bio, Inc., Shiga, Japan) were placed into PCR tubes. The
PCR was performed under the following conditions: 70°C for 10 min
and 0°C for 2 min. Next, the sample was mixed, and 2.5 μl 5X
PrimeScript buffer, 10μl PrimeScript RT enzyme mix, 10 μl random
6-mers (100 μM), 19 μl template and 8.5 μl RNase-free
H2O (Takara Bio, Inc.) were placed into PCR tubes. The
PCR was performed under the following conditions: Reverse
transcription reaction at 37°C for 15 min; and denaturation at 85°C
for 5 sec. For the qPCR, 9 μl SYBR Green Mix, 2 μl miR-449a forward
primer, 2μl miR-449a reverse primer, 2 μl template cDNA and 5 μl
RNase-free H2O (Takara Bio, Inc.) were placed into the
PCR tubes and underwent qPCR under the following conditions: 40
cycles of predegenration at 95°C for 20 sec; denaturation at 95°C
for 10 sec; annealing at 60°C for 20 sec; prolongation (primer as a
starting point, extending along the template 5′ to 3′ direction) at
70°C for 10 sec. Melting curve analysis was performed through a
temperature range of 70–95°C at a rate of 0.4°C/sec.
Detecting the CDK6 protein by western
blot analysis
The protein samples were heated at 95°C for 10 min
with the sample buffer (250 mM Tris-HCl, 4% sodium dodecyl sulfate,
2% β-mercaptoethanol, 10% glycerol and 0.003% bromophenol blue) and
separated by 12% sodium dodecyl sulfate polyacrylamide gel
electrophoresis. The protein samples were then transferred onto a
polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA),
which were blocked with 5% skimmed dry milk in Tris-buffered saline
(TBS) with 0.1 % Tween 20 (TBS-T) for 1 h. The membranes were then
incubated overnight at 4°C with primary antibody diluted in 0.3%
bovine serum albumin (BSA)-TBS-T. The membranes were incubated with
the primary antibodies (CDK6, 1:1,000 and β-actin, 1:400; Abcam,
Cambridge, UK) at 4°C for 12 h, and then with the horseradish
peroxidase-linked secondary antibodies (1:1,000, goat anti-mouse
monoclonal CD151 and goat anti-rabbit monoclonal β-actin; Abcam) at
37° for 1 h. The membranes were incubated with ECL Plus reagent
(Amersham Biosciences, Uppsala, Sweden) and scanned using the Storm
imaging system (Amersham Biosciences). Immunoreactive products were
quantified using Quantity One software (Bio-Rad, Hercules, CA, USA)
by determining the optical density of the protein bands.
Real-time cell analysis
The cells of the miR-449a upregulation, miR-449a
downregulation and control groups were seeded into the E-plate and
plated in 37°C incubators with 5% CO2. The experimental
specimens were scanned, and data extraction was performed by
xCELLigence real-time cell analysis (RTCA; Roche).
4′-6-diamidino-2-phenylindole (DAPI)
staining test
The cells of the miR-449a upregulation, miR-449a
downregulation and control groups were seeded into the 24-well
plates and plated in 37°C incubators with 5% CO2. The
cells were stained by 0.1% DAPI-phosphate-buffered saline (PBS)
subsequent to being fixed by 4% paraformaldehyde and incubated on
an agitator for 30 min. Images were captured using a inverted
fluorescence microscope (Olympus IX71; Olympus, Tokyo, Japan)
subsequent to being mounted using glycerin.
Cell scratch test
The cells of the miR-449a upregulation, miR-449a
downregulation and control groups were seeded into the 6-well
plates and plated in 37°C incubators with 5% CO2,
diluted at 1×106/ml. A scratch was formed on a single
layer of cells using ultra-high temperature sterilized toothpicks,
then the cells were washed three times with PBS. Images were
captured using the inverted fluorescence microscope subsequent to
being mounted using glycerin.
Immunofluorescence test
The cells that reached 60–70% confluence prior to
the test were seeded into the 6-well plates in 37°C incubators with
5% CO2. The cells of the miR-44a upregulation and
downregulation groups, as well as the control group were stained by
0.1% TritonX-100-PBS after being fixed by 4% paraformaldehyde and
incubated in an agitator for 10 min at room-temperature. The cells
were then blocked with 3% BSA-PBS for 30 min and incubated with the
primary antibodies (CDK6, 1:10; Abcam) at 4°C for 12 h, and then
incubated with the secondary antibody (1:500, polyclonal goat
anti-mouse IgG-H&L; Abcam) at 37°C for 1 h. Images were
captured using the inverted fluorescence microscope subsequent to
being mounted using glycerin.
Statistical analysis
Statistical analyses were performed using the SPSS
13.0 software package (SPSS, Inc., Chicago, IL, USA). Normal
distributions were analyzed using the one sample Kolmogorov-Smirnov
test for goodness of fit and the homogeneity test of variance was
performed among the groups, suggesting homogeneity of variance
(P>0.05). Differences between two groups were compared using
one-way analysis of variance, while multiple comparisons were
compared using Fisher’s least significant difference test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression of miR-449a and the CDK6
protein in the gastric carcinoma tissue
The expression levels of miR-449a were compared
between the gastric carcinoma and tumor-adjacent normal tissues
using relative quantification, with the expression of the U6 gene
as a reference. The qPCR showed that the relative expression of
miR-449a was 24.665±1.557 in the gastric carcinoma tissue and
49.207±13.433 in the tumor-adjacent normal tissue, which showed
that the expression of miR-449a was downregulated in the gastric
carcinoma tissue (P<0.001; Fig.
1A). Western blotting was also used to test the different
expression levels of the CDK6 protein in the gastric carcinoma and
tumor-adjacent normal tissues; the relative expression of the CDK6
protein was upregulated in the gastric carcinoma tissue compared
with the tumor-adjacent normal tissue (P<0.001; Fig. 1B and C). The relative expression of
the CDK6 protein was found to be negatively associated with the
relative expression of miR-449a by Spearman’s correlation analysis
(Fig. 1D).
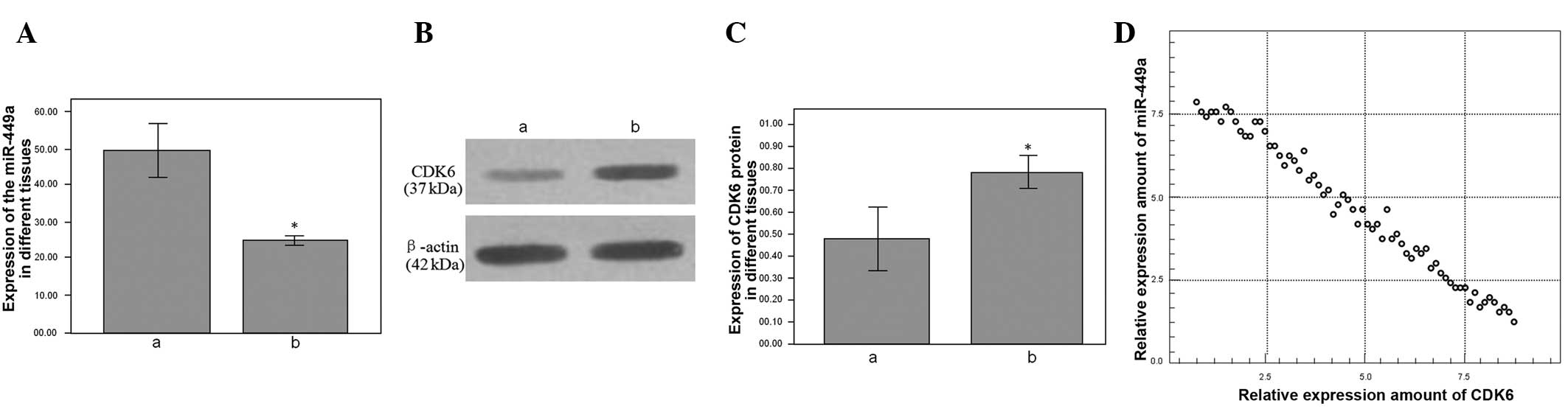 | Figure 1(A) Relative expression of miR-449a in
gastric carcinoma and tumor-adjacent normal tissues, as determined
by qPCR. ‘a’ represents the tumor-adjacent normal tissue; relative
expression, 24.665±1.557. ‘b’ represents the gastric carcinoma
tissue; relative expression, 49.207±13.433, P<0.001. (B) The
relative expression of the CDK6 protein in the gastric carcinoma
and tumor-adjacent normal tissues, as determined by western blot
analysis. (C) ‘a’ represent the tumor-adjacent normal tissue;
relative expression, 0.482±0.290. ‘b’ represents the gastric
carcinoma tissue; relative expression, 0.792±0.165, P=0.003. (D)
The relative expression of CDK6 protein was negatively correlated
with the relative expression of miR-449a, as determined by
Spearman’s correlation analysis, r(tumor-adjacent normal
tissue)=-0.614; r(gastric carcinoma tissue)=-0.694, P<0.001.
Relative expression is measured as a percentage and
*P<0.05 vs. tumor-adjacent normal tissues.. miR,
microRNA; qPCR, quantitative PCR; CDK6, cyclin-dependent kinase
6. |
Effects of different expression levels of
miR-449a in the MGC-803 cells
The xCELLigence RTCA system is an electronic cell
sensor array that has been newly developed and is currently being
tested for the dynamic monitoring of cell attachment,
proliferation, damage and death (6). The expression of the miR-449a of the
MGC-803 cell line was upregulated or downregulated by Lipofectamine
2000 transfection to establish the different miR-449a expression
cell groups for the experiment. The qPCR result showed the
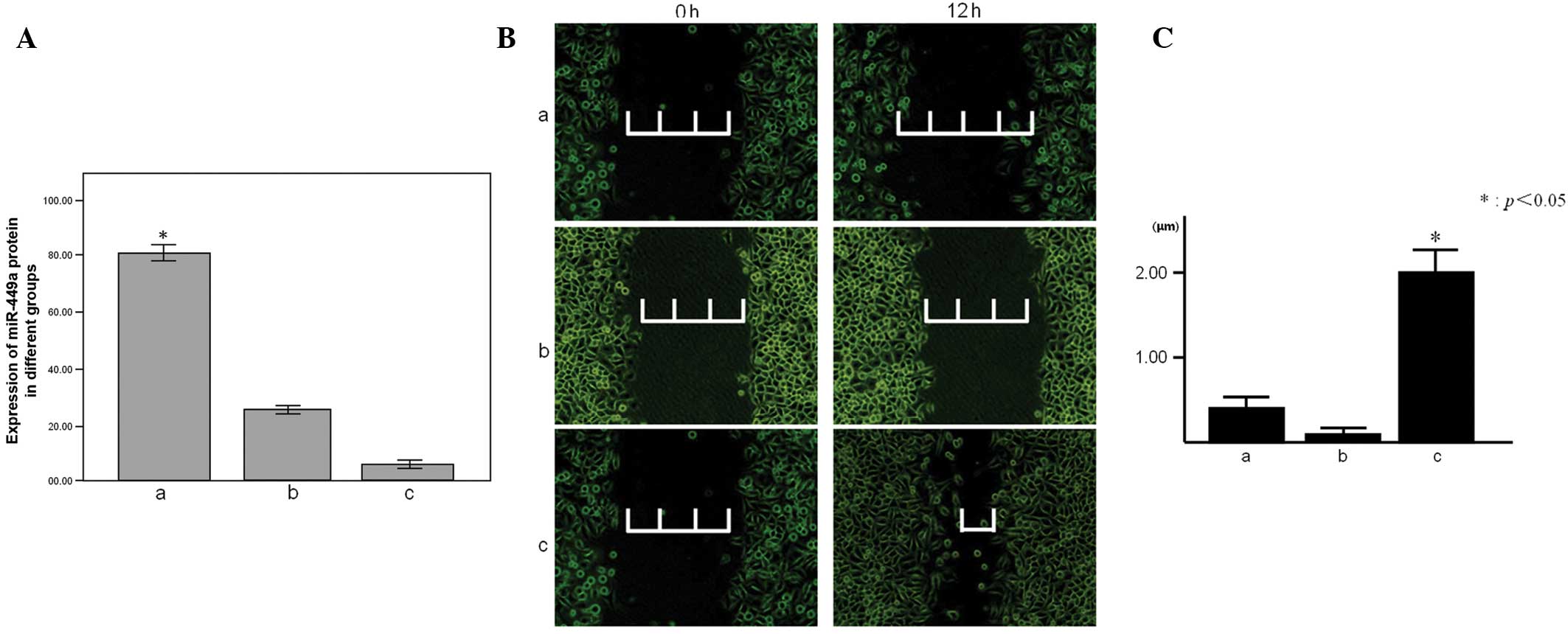
different expression levels of the groups (Fig. 2A). The proliferative capacity of the
MGC803 cell of the miR-449a upregulation group was shown to be
decreased compared with the miR-449a downregulation and control
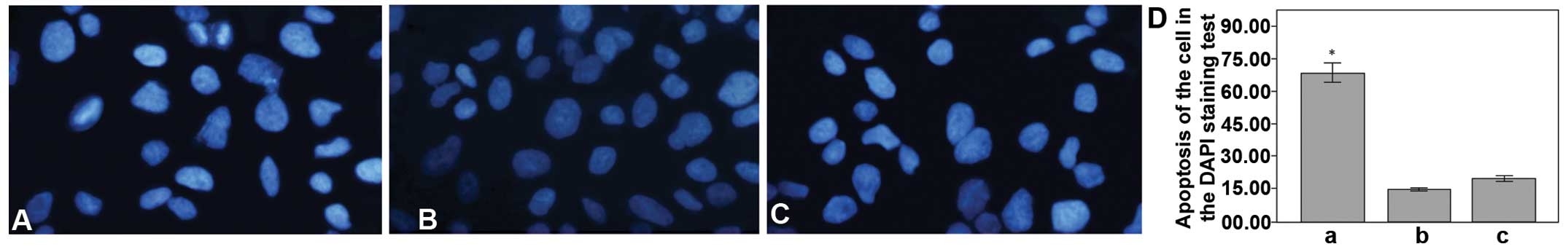
groups by RTCA (Fig. 3A). DAPI is a
useful tool that can be used to monitor the apoptosis of the cell
lines, by exhibiting a strong fluorescence when DAPI becomes bound
to natural double-stranded DNA; this was determined from the
nuclear morphology of the MGC-803 cell line in the present
experiment. The normal cell nuclei were round in shape and staining
was evenly distributed. When the cell became apoptotic, the cell
nuclei became deformed due to the aggregation of the DNA. The
number of apoptotic cells was higher in the miR-449a upregulation
group than in the downregulation and control groups (Fig. 4). The cell scratch test is a useful
and straightforward way to test the migration and invasion of
cells. The degree of cell migration and invasion can be determined
by the mensuration of the change in distance between the edges of
the scratch. In the present study, the miR-449a downregulation
group was found to exhibit more cell migration and invasion than
the groups, as this distance had decreased after 12 h (Fig. 2B and C).
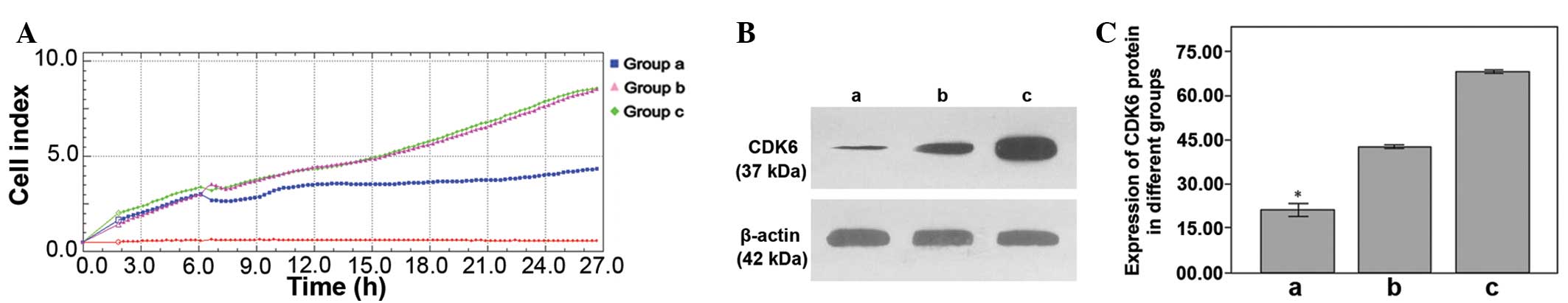 | Figure 3(A) Proliferation of the MGC-803 cells
in 3 groups, as observed by RTCA. ‘a’ represents the miR-449a
upregulation group; proliferation coefficient, 4.282±0.386,
P<0.001. ‘b’ represents the control group; proliferation
coefficient, 9.002±0.066. ‘c’ represents the miR-449a
downregulation group; proliferation coefficient, 9.049±0.073. (B)
The relative expression of the CDK6 protein in 3 groups. (C) ‘a’
represents the miR-449a upregulation group; relative expression of
CDK6 protein, 21.243±4.097, P<0.001. ‘b’ represents the control
group; relative expression, 41.831±1.647. ‘c’ represents the
miR-449a downregulation group; relative expression, 67.418±1.369.
Relative expression is measured as a percentage and
*P<0.05. RTCA, real-time cell analysis; miR,
microRNA; CDK6, cyclin-dependent kinase 6. |
Effect of different expression levels of
miR-449a on the expression of the CDK6 protein in the MGC-803
cells
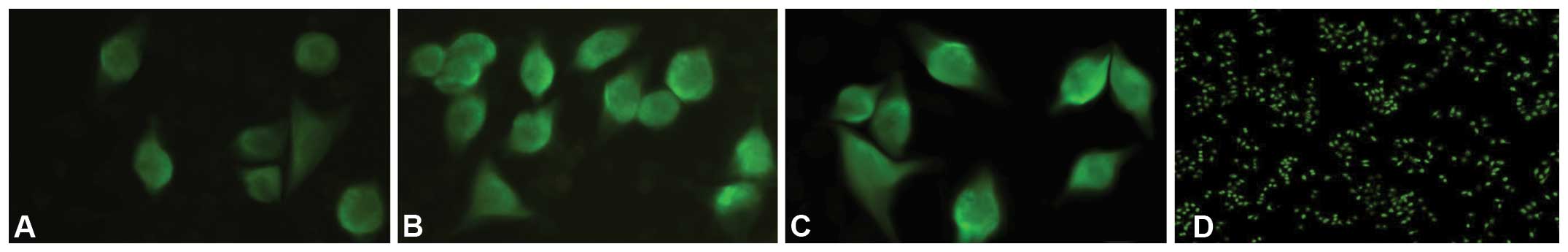
Immunofluorescence and western blot analyses are
common, powerful and useful techniques that are based on the
antigen-antibody reaction. In the present study, immunofluorescence
and western blot analyses were used to detect the expression level
of the CDK6 protein in the MGC-803 cells. It was shown that the
fluorescence intensity decreased in the miR-449a upregulation group
(Fig. 5) and that the relative
expression of the CDK6 protein was downregulated in the miR-449a
upregulation group (Fig. 3B and
C).
Discussion
More attention is currently being focused on the
association between miRNA and the gastric carcinoma, in association
with the pace of research on non-coding nucleic acids (3,7,8). It
has been found that miR-449 is closely correlated with the
occurrence and development of gastric cancer. Studies have found
that miR-449 can not only induce apoptosis by affecting the E2F-p53
negative feedback loop system, but that it can also inhibit cell
proliferation and cell carcinogenesis by activating the Notch
signaling pathway (5). Certain
studies have shown that miR-449 could directly target and switch
off the expression of GMNN, MET, CCNE2 and another cancer-related
genes, the majority of which are associated with the cell cycle, by
syncretizing with the 3′UTR of target genes. If the miR-449 are
downregulated in the gastric mucosal cell, it may ultimately lead
to gastric cancer through the indefinite proliferation of the
gastric mucosal cell (9). In the
present study, gastric carcinoma and tumor-adjacent normal tissues
were obtained from gastric cancer patients during surgery. qPCR was
utilized to analyze the expression of miR-449a in the different
tissues, and it was shown that the expression of miR-449a was
downregulated in the gastric carcinoma tissue, which indicated that
the downregulation of miR-449a was correlated with the occurrence
of gastric cancer. This provide further evidence that miR-449a has
a similar function to that of a cancer suppressor gene.
Cell cycle control is becoming one of the main areas
of study in the field of cancer prevention. Cell cycle disorder,
due to the imbalance of cell cycle element secretion, is one of the
characteristics of tumor cells. CDKs and cyclins are closely
associated with the development and cancerization of tissues by
regulating the cell cycle, two areas that are current research
hotspots (10). CDK6, a member of
the CDK family, can express the CDK6 protein, which is a
significant cell cycle controlling factor (11). It has been reported that the CDK6
protein is closely associated with the occurrence and development
of gastric cancer (12). Cam et
al (13) found that the
expression of the CDK6 protein was maladjusted in the tumor cells.
The upregulated expression of the CDK6 protein makes the
G1 phase longer in the cells and generates a positive
change in the proliferation rate. The increased cell proliferation
or lessened cell apoptosis are the beginning of cancerization. The
present study showed that there was downregulation of miR-449a and
upregulation of the CDK6 protein in the clinical gastric cancer
tissue samples. The association between the downregulation of
miR-449a and the upregulation of CDK6 protein in gastric carcinoma
was consequently identified, along with the association between the
downregulation of miR-449a and the proliferation, apoptosis and
migration of the gastric cancer MGC-803 cell line. The study showed
that increased apoptosis and decreased proliferation occurred if
the expression of miR-449a was upregulated in the MGC-803 cells,
while cell proliferation and migration were increased the
expression was downregulated. The study indicates that the
expression level of miR-449a may affect the clinical pattern of
cancer development gastric cancer patients.
miR-449a, which functions like a cancer suppressor
gene, is closely associated with a great variety of cell cycle
control genes. A previous study found that miR-449 could regulate
the cell cycle-related CDK gene family (14) not only by regulating CDK-rb-e2F1
through an auto-regulatory feedback circuit (15), but also by targeting, identifying
and regulating the target gene expression directly (7). Therefore, the gastric mucosal cell may
have unlimited cell proliferation and finally develop gastric
cancer due to the disorder of the cell cycle caused by
downregulating miR-449, and miR-449a is the most common subtype of
the miR-449 family (16). In the
present study, the different expression levels of CDK6, affected by
the different miR-449a expression levels, were analyzed using
immunofluorescence and western blot analyses. The higher the
expression level of miR-449a, the weaker the cell fluorescence was
in the immunofluorescence analysis, and the western blotting
results also showed that the expression level of miR-449a was
negatively correlated with the CDK6 protein expression level. From
this, it was indicated that miR-449a could regulate the expression
of the CDK6 protein, and that this association may be closely
correlated with the occurrence and development of gastric cancer.
The abnormal expression of certain miRNAs, such as miR-196b, could
indicate the occurrence of certain tumors, and miR-196b had thus
been defined as a significant specific marker by scientists
(17,18). As the expression of miRNA in plasma
has tissue specificity, monitoring specific miRNA expression in the
plasma may be a means for the early screening for cancer in
high-risk groups (19).
The result of the present study have validated the
fact that the downregulation of miR-449a and the upregulation of
CDK6 protein participate in the occurrence and development of
gastric cancer, and have also added to the data on the association
between miR-449a and the CDK6 protein. We hypothesize that the
correlation of gastric cancer and miRNA will become a novel
direction in the future research on gastric cancer prevention,
which will be good for the early diagnosis of gastric cancer by
miRNA expression level screening. The present study aimed to
accumulate related basic research data for cancer prevention and
control through research into miR-449a, in order to improve the
early diagnosis of gastric cancer patients and improve the gastric
cancer survival rate.
Acknowledgements
The authors would like to thank the Department of
Medical Experiment (General Hospital of Guangzhou Military Command)
for the MGC-803 cell line. The study was supported by Science and
Technology Planning Project of Guangdong Province, China (grant no.
2011B031800317) and the Undergraduate Innovative Experiment Project
of Guangdong Province, China (grant no. 1057212046).
References
|
1
|
Smith MG, Hold GL, Tahara E, et al:
Cellular and molecular aspects of gastric cancer. World J
Gastroenterol. 12:2979–2990. 2006.
|
|
2
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011.
|
|
3
|
Stenvang J, Petri A, Lindow M, et al:
Inhibition of microRNA function by antimiR oligonucleotides.
Silence. 3:12012.
|
|
4
|
Wu WK, Lee CW, Cho CH, et al: MicroRNA
dysregulation in gastric cancer: a new player enters the game.
Oncogene. 29:5761–5771. 2010.
|
|
5
|
Lizé M, Klimke A and Dobbelstein M:
MicroRNA-449 in cell fate determination. Cell Cycle. 10:2874–2882.
2011.
|
|
6
|
Teng Z, Kuang X, Wang J and Zhang X:
Real-time cell analysis - a new method for dynamic, quantitative
measurement of infectious viruses and antiserum neutralizing
activity. J Virol Methods. 193:364–370. 2013.
|
|
7
|
Bou Kheir T, Futoma-Kazmierczak E,
Jacobsen A, et al: miR-449 inhibits cell proliferation and is
down-regulated in gastric cancer. Mol Cancer. 10:292011.
|
|
8
|
Chan SH, Wu CW, Li AF, et al: miR-21
microRNA expression in human gastric carcinomas and its clinical
association. Anticancer Res. 28:907–911. 2008.
|
|
9
|
Tsujiura M, Ichikawa D, Komatsu S, et al:
Circulating microRNAs in plasma of patients with gastric cancers.
Br J Cancer. 102:1174–1179. 2010.
|
|
10
|
Duman-Scheel M, Weng L, Xin S and Du W:
Hedgehog regulates cell growth and proliferation by inducing Cyclin
D and Cyclin E. Nature. 417:299–304. 2002.
|
|
11
|
Kozar K and Sicinski P: Cell cycle
progression without cyclin D-CDK4 and cyclin D-CDK6 complexes. Cell
Cycle. 4:388–391. 2005.
|
|
12
|
Laman H, Funes JM, Ye H, et al:
Transforming activity of Fbxo7 is mediated specifically through
regulation of cyclin D/cdk6. EMBO J. 24:3104–3116. 2005.
|
|
13
|
Cam EJ, Liu BD, Bjeldanes LF and Firestone
GL: Indole-3-carbinol inhibits CDK6 expression in human MCF-7
breast cancer cells by disrupting Sp1 transcription factor
interactions with a composite element in the CDK6 gene promoter. J
Biol Chem. 276:22332–22340. 2001.
|
|
14
|
Yang XJ, Feng M, Jiang X, et al: miR-449a
and miR-449b are direct transcriptional targets of E2F1 and
negatively regulate pRb-E2F1 activity through a feedback loop by
targeting CDK6 and CDC25A. Genes Dev. 23:2388–2393. 2009.
|
|
15
|
Feng M and Yu Q: miR-449 regulates
CDK-Rb-E2F1 through an auto-regulatory feedback circuit. Cell
Cycle. 9:213–214. 2010.
|
|
16
|
Lizé M, Herr C, Klimke A, et al:
MicroRNA-449a levels increase by several orders of magnitude during
mucociliary differentiation of airway epithelia. Cell Cycle.
9:4579–4583. 2010.
|
|
17
|
Bhatia S, Kaul D and Varma N: Functional
genomics of tumor suppressor miR-196b in T-cell acute lymphoblastic
leukemia. Mol Cell Biochem. 346:103–116. 2011.
|
|
18
|
Guan Y, Mizoguchi M, Yoshimoto K, et al:
MiRNA-196 is upregulated in glioblastoma but not in anaplastic
astrocytoma and has prognostic significance. Clin Cancer Res.
16:4289–4297. 2010.
|
|
19
|
Katsios C, Baltogiannis G and Roukos DH:
Progress, challenges and new genome-based concepts in the
multidisciplinary treatment of gastric cancer. Expert Rev
Anticancer Ther. 11:503–506. 2011.
|