Introduction
Non-small cell lung cancer (NSCLC) is one of the
leading causes of cancer-related mortality worldwide, despite
considerable progress in surgery, chemotherapy, radiotherapy and
biological targeted therapy (1). In
the previous decade, NSCLC research has demonstrated that these
therapies predominantly function to improve the patient’s quality
of life and that the overall five-year survival rate for such
tumors is <15% (2). Previous
studies indicate that various solid tumors, such as brain gliomas
(4,5), breast (6), prostate (7), colon (8) and liver cancers (9), contain a small population of cancer
stem cells (CSCs) that are responsible for tumor maintenance and
dissemination. CSCs exhibit unlimited proliferative potential, the
ability to self-renew, an elevated capacity to induce malignancy,
and may be associated with the initiation and progression of
malignancies, chemotherapy and radiotherapy resistance, as well as
tumor recurrence and metastasis (10–14).
Since the characteristics of these tumor cells are similar to those
of classic stem cells, they have been termed CSCs. It is proposed
that therapies specifically targeting the stem cell signaling
pathways utilized by CSCs may be beneficial in combating specific
types of cancer (15–16).
Initial research into lung CSCs (LCSCs) has been
undertaken in recent years. For example, Kim et al (17) identified a group of cells at the
bronchioalveolar duct junction carrying Clara and alveolar markers,
which commenced division following naphthalene
administration-induced damage. This cluster of stem cell antigen-1
(Sca-1)+/cluster of differentiation (CD)34+
cells was enriched by fluorescence-activated cell-sorting, and
demonstrated an enhanced capacity for self-renewal and
differentiation in vitro. Activation of the oncogenic
protein K-ras boosted the proliferation of the double-positive
cells and accelerated tumorigenicity. Thus, the
Sca-1+/CD34+ cells were termed
bronchioalveolar stem cells (BASC) and may be the origin of
adenocarcinomas. Furthermore, Ho et al (18) used flow cytometry and the Hoechst
33342 dye efflux assay to isolate and characterize side population
(SP) cells from six human lung cancer cell lines (H460, H23,
HTB-58, A549, H441 and H2170) and sixteen clinical lung cancer
samples. The study performed xenograft experiments to determine
that SP cells were enriched in tumor-initiating capability compared
with non-SP cells, as well as Matrigel invasion assays to
demonstrate that SP cells exhibit a higher potential for
invasiveness. In addition, SP cells displayed elevated expression
levels of ATP-binding cassette superfamily G member 2 (ABCG2), as
well as other ABC transporters, and exhibited resistance to
multiple chemotherapeutic agents. These findings indicate that SP
cells are an enriched source of lung tumor-initiating cells with
stem cell properties. Recent study has demonstrated that SCLC and
NSCLC contain cells that express the glycoprotein prominin-1
(CD133), a cancer stem cell marker, which is essential for tumor
cell propagation and metastasis (19). The proliferative capacity of
CD133+ cells is yet to be determined; however, it is
hypothesized that these cells serve as a reservoir for generating
further cancer cells that are capable of tumorigenesis, leading to
metastasis (19).
In the present study, a population with a
CD133+ phenotype cells from a single cell suspension of
lung adenocarcinoma tissues was isolated using magnetic activated
cell sorting (MACS) technology and enriched in a serum-free
culture. Furthermore, the self-renewal, differentiation and
tumorigenicity of CD133+ cells in NOD/SCID mice were
investigated.
Materials and methods
Cell culture and lung cancer single cell
suspension preparation
Eight fresh lung cancer specimens were obtained
(Table I) from patients who
underwent surgery at the Department of Throacic Surgery, Taizhou
People’s Hospital (Taizhou, China) between February and April 2013.
The specimens were cut into 0.5-mm sections following the removal
of visible blood vessels and necrotic tissue. The tissue specimens
were washed numerous times with D-Hank’s solution (Beijing Huamaike
Biotechnology Co., Ltd., Beijing, China) and left overnight in
Dulbecco’s modified Eagle’s medium with Ham’s nutrient mixture F-12
(DMEM-F12) supplemented with high doses of penicillin/streptomycin
and amphotericin B to avoid contamination. The specimens were
enzymatically digested in 50 ml BD Falcon™(Becton-Dickinson,
Franklin Lakes, NJ, USA) supplemented with collagenase IV (final
concentration, 0.1%; Nanjing Sunshine Biotechnology Co., Ltd.,
Nanjing, China) and hyaluronidase (final concentration, 0.1%;
Nanjing Sunshine Biotechnology Co., Ltd.) for 1 h under 5%
CO2 at 37°C. The remaining cell debris was removed by
passing the cells through a 70μm-diameter disposable cell mesh
filter and centrifuging for 15 min at a speed of 400 × g. Finally,
the primary human lung cancer cells were cultured in RPMI-1640
supplemented with small airway growth medium (SAGM) SingleQuots™
kit (Lonza, Basel, Switzerland) medium with
penicillin/streptomycin, and features of the growth pattern were
observed. Flow cytometry was performed three times using an
EPICS® XL™ flow cytometer (Beckman Coulter, Brea, CA,
USA) to quantify the expression of CD133 (Miltenyi Biotec, Inc.,
Auburn, CA, USA) on the surface of primary human lung cancer cells.
The present study was approved by the Ethics Committee of Taizhou
People’s Hospital (Taizhou, China) and was performed according to
the Declaration of Helsinki. Written informed consent was obtained
from the family of each patient.
 | Table ICase description and CD133 expression
in eight NSCLC patients. |
Table I
Case description and CD133 expression
in eight NSCLC patients.
| Patient | Age, years | Gender | Tumor subtype | TNM stage | CD133 proportion,
% |
|---|
| 1 | 73 | Male | SCC | IIA | None detected |
| 2 | 56 | Female | AdC | IIIA | 1.9 |
| 3 | 65 | Female | AdC | IIA | None detected |
| 4 | 61 | Male | AdC | IIB | 2.1 |
| 5 | 57 | Male | AdC | IIB | None detected |
| 6 | 74 | Male | AdC | IIIA | 1.3 |
| 7 | 69 | Male | SCC | IIB | None detected |
| 8 | 58 | Male | AdC | IIA | 0.8 |
CD133 cell sorting using immunomagnetic
beads
A single cell suspension of ~1×107 lung
cancer cells was used for cell sorting. Cells were incubated with
CD133/l rabbit anti-human polyclonal immunomagnetic beads (Miltenyi
Biotec, Inc.) for 30 min at 4°C. For magnetic separation, a MACS
column (Miltenyi Biotec, Inc.) was used to retain the positive
cells linked with the beads. The CD133+ cells obtained
from the column were centrifuged and resuspended in serum-free
DMEM-F12 medium containing 50 μg/ml insulin, 100 μg/ml
apo-transferrin, 10 μg/ml putrescine, 0.03 mM sodium selenite (all
from Sigma-Aldrich, St. Louis, MO, USA), 2 mM progesterone (Pure
Chemistry Scientific Inc., Sugarland, TX, USA), 0.6% glucose (LGM
Pharma, Nashville, TN, USA), 5 mM HEPES (Nanjing Search Biotech
Co., Ltd., Nanjing, China), 0.1% sodium bicarbonate (Nanjing Search
Biotech Co., Ltd.), 0.4% bovine serum albumin (BSA; Wuhan Boster
Bio-Engineering Co., Ltd., Wuhan, China), glutamine (Amresco LLC,
Solon, OH, USA) and antibiotics (Gibco-BRL, Carlsbad, CA, USA)
supplemented with 20 μg/ml epidermal growth factor (EGF; PeproTech
EC Ltd., London, UK) and 10 μg/ml basic fibroblast growth factor
(bFGF; PeproTech EC Ltd.). The purity of the CD133+ and
CD133− cells was evaluated using standard flow
cytometric analysis. The CD133+ and CD133−
cells were harvested, and sphere formation, tumorigenicity and
differentiation activity were determined.
Cell surface marker analysis by flow
cytometry
Cells (1×105)were resuspended in 100 μl
phosphate-buffered saline (PBS) supplemented with 0.5% BSA and 2 mM
EDTA, and incubated with 10 μl polyclonal mouse anti-human CD133-PE
conjugated antibody (1:100; Miltenyi Biotec, Inc.), monoclonal
mouse anti-human cytokeratin (CK)8 (1:100) and mouse anti-human
CK18 (1:100; Dako, Glostrup, Denmark) for 10 min at 4°C. Following
washing with PBS, the cells were resuspended in a solution of PBS
and 2 μl 7-amino-actinomycin D (7-AAD), and analyzed using a
EPICS® XL™ flow cytometer (Beckman Coulter).
Immunofluorescence
Slides containing CD133+ tumor spheres
and CD133− cells were collected and immersed in PBS for
5 min, permeabilized in 0.1% Triton X-100 (Bebco Industries Inc.,
La Marque, TX, USA) for 10 min and washed with PBS 3 times for 5
min. Following blocking with 5% BSA at 37°C for 30 min, the slides
were incubated overnight with rabbit polyclonal anti-human CD133
(dilution, 1:300; Abcam, Cambridge, UK) at a temperature of 4°C.
Subsequently, the cells were incubated with Cy3-conjugated
monoclonal goat anti-rabbit IgG secondary antibody (1:2,000; Wuhan
Boster Bio-Engineering Co., Ltd.) diluted with 1% BSA at 37°C for 1
h. Finally, the cell nuclei were stained with DAPI (dilution,
1:200). Images were captured and visualized using fluorescence
microscopy (AF6000; Leica, Mannheim, Germany).
Sphere-forming assay
CD133+ tumor spheres and
CD133− cells were dissociated into single-cell
suspensions, and transferred to 96-well plates. The cells were
cultured in serum-free DMEM-F12 medium containing 50 μg/ml insulin,
100 μg/ml apo-transferrin, 10 μg/ml putrescine, 0.03 mM sodium
selenite (all from Sigma-Aldrich), 2 μM progesterone (Pure
Chemistry Scientific Inc.), 0.6% glucose (LGM Pharma), 5 mM HEPES
(Nanjing Search Biotech Co., Ltd.), 0.1% sodium bicarbonate
(Nanjing Search Biotech Co., Ltd.), 0.4% BSA (Wuhan Boster
Bio-Engineering Co., Ltd.), glutamine (Amresco LLC) and
antibiotics, supplemented with 20 ng/ml EGF and 10 ng/ml bFGF.
Wells containing greater than one cell or no cells were marked and
dismissed from statistical analysis. The cells were cultured in 5%
CO2 at 37°C for 2–3 weeks, with the medium replaced or
supplemented with fresh growth factors twice a week. Wells that
contained spheres were counted using inverted phase contrast
microscopy (DMI 6000B; Leica) and the percentage of cells
exhibiting sphere-forming capacity was calculated.
Differentiation
CD133+ tumor spheres and primary lung
cancer cells were cultured in 24-well plates. To allow cell
attachment and differentiation, the stem cell medium was replaced
with RPMI-1640 supplemented with SAGM SingleQuots kit medium with
penicillin/streptomycin and 10% fetal bovine serum (FBS; GE
Healthcare Life Sciences, Logan, UT, USA). The acquisition of
differentiation markers and the loss of stem cell markers was
evaluated using flow cytometry before and after cell attachment, as
described above. The experiment was repeated three times and the
mean values were calculated.
Chemotherapy resistance studies
Cells (1×103) obtained from
CD133+ tumor spheres and CD133− cell
dissociation were plated in 96-well flat-bottomed plates. The
chemotherapeutic agents gemcitabine and cisplatin (Jiangsu Hansoh
Pharmaceutical Co., Ltd., Lianyungang, China) were added at final
concentrations of 250 mM and 5 mg/ml, respectively. Following six
days of treatment, cell viability was evaluated using an MTT and
Trypan blue (Nanjing Search Biotech Co., Ltd.) exclusion assay.
Data are expressed as the mean of three independent experiments
performed with the two experimental procedures.
Tumorigenicity in NOD/SCID mice
CD133− cells and CD133+ tumor
spheres were mechanically dissociated to obtain single cell
suspensions and diluted in growth factor-containing medium prior to
subcutaneous injection. Serial dilutions (102,
103, 104 and 105 cells) of the
cells were subcutaneously injected into the abdominal wall of 20
four-week-old NOD/SCID mice (five mice per group; Beijing Vital
River Laboratory Animal Technology Co., Ltd., Beijing, China).
Tumor size was measured using calipers and tumor volume was
calculated using the equation: Tumor volume = (π × maximum length ×
maximum width × maximum height)/6. Immunohistochemistry (IHC), as
well as hematoxylin and eosin staining (H&E), were performed to
analyze the tumor histology and to compare mouse xenografts with
patient tumors.
IHC
Paraffin-embedded tissue blocks were cut into 4-μm
sections and representative sections were analyzed
immunohistochemically (EliVision™ Plus IHC kit; Wuhan Boster
Biological Engineering Co., Ltd., Wuhan, China) for mouse
anti-human polyclonal CK8 and CK18 (dilution, 1:200; Miltenyi
Biotec, Inc.). Briefly, the sections were dewaxed in xylene and
rehydrated in ethanol using graded concentrations of alcohol.
Endogenous peroxidase activity was blocked by incubating the
sections in 5% hydrogen peroxide and absolute methanol at room
temperature for 10 min, and antigen retrieval was performed in a
microwave oven for two cycles of 10 min each. Primary antibodies
were applied for 1 h at room temperature, the sections were washed
three times with 0.05 M Tris-buffered saline (TBS; pH 7.2) and 50
μl IgG/horseradish peroxidase secondary antibody (Wuhan Boster
Biological Engineering Co., Ltd.) was added, followed by incubation
for 30 min at room temperature. The sections were washed three
times with TBS and the reaction products were visualized using a
diaminobenzidine (DAB) kit (Wuhan Boster Biological Engineering
Co., Ltd.). The sections were counterstained with H&E,
dehydrated and evaluated under a light microscope (DM 3000;
Leica).
Statistical analysis
Statistical analysis was performed using SPSS
software (version, 13.0; SPSS, Inc., Chicago, IL, USA). All
experiments were performed a minimum of three times and
representative results are presented as the mean values ± standard
deviation. Statistical analysis was performed by one-way analysis
of variance and comparisons among groups were achieved using
independent sample t-tests. P<0.05 indicated a statistically
significant difference.
Results
CD133+ cells in primary lung
cancer cells
CD133+ cells were detected in 4/8 primary
human lung cancer samples using the EPICS XL flow cytometer
(Beckman Coulter). The proportion of CD133+ cells was
1.9%, 2.1%, 1.3% and 0.8% in each respective case, and the
pathological types of all four cases were lung adenocarcinoma
(Table I). Immunomagnetic beads
identified two cases of primary lung cancer cell suspension
exhibiting a high percentage of CD133+ cells for cell
sorting. Following immunomagnetic sorting with CD133 beads,
CD133+ tumor cells were cultured in serum-free DMEM-F12
medium supplemented with 20 ng/ml EGF and 10 ng/ml bFGF, and
analyzed using flow cytometry (EPICS XL flow cytometer; Beckman
Coulter). Three measurements of CD133 expression in the
CD133+-sorted population indicated a high mean
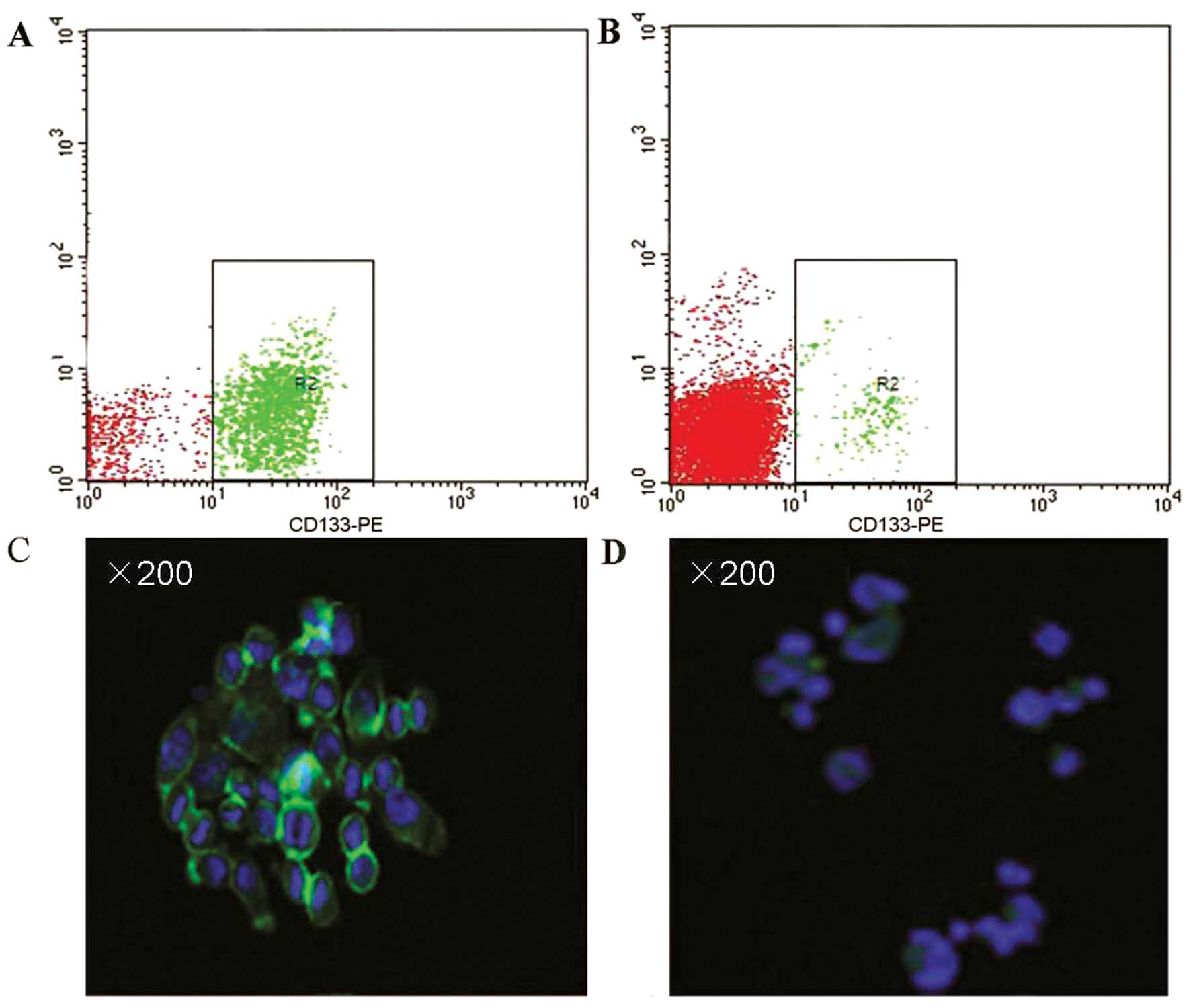
CD133+ expression rate of 89.15±7.80% (Fig. 1A). However, the CD133+
expression rate in the single-cell suspensions of primary lung
cancer was only 2.07±0.21% (Fig.
1B). Additionally, immunostaining assays demonstrated extensive
expression of CD133 in the tumor sphere samples (Fig. 1C), while lower levels of CD133
expression were detected in the primary lung cancer cells (Fig. 1D).
CD133+ cell enrichment in the
serum-free cultures
According to the CSC theory, only a small number of
cells in the tumor exhibit CSC characteristics. These stem-like
cells are able to grow in serum-free medium and are innately
resistant to chemotherapy, due to their ability to pump out toxic
agents. The CD133+ cells obtained via immunomagnetic
cell sorting were harvested and cultured in serum-free medium
containing various growth factors. Following three days of
culturing, numerous individual cells in the CD133+
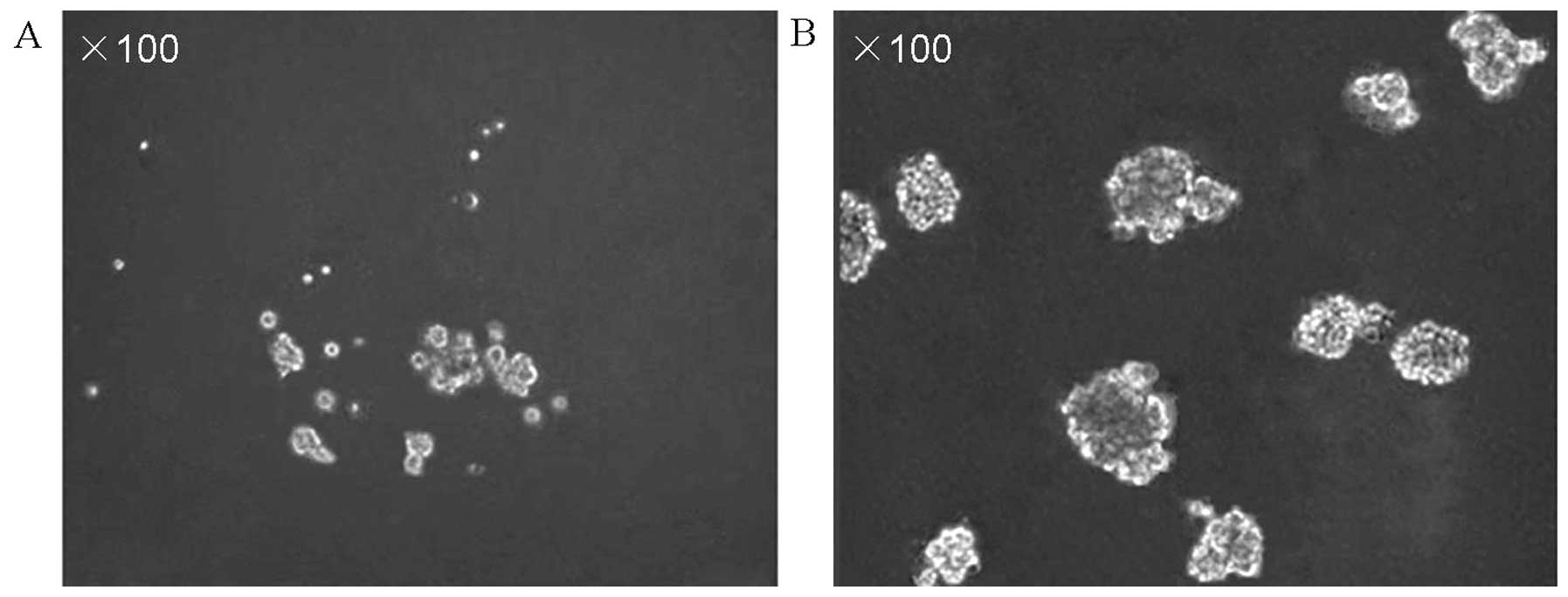
suspension culture were observed to survive and proliferate. After
approximately one week, these cells gradually formed sphere
colonies of various sizes and irregular shapes (Fig. 2A). After approximately one month,
these cell cultures were exclusively formed by cellular clusters
resembling tumor spheres (Fig. 2B).
However, in standard stem cell medium cultures, almost no clear
sphere colonies were observed; the majority of CD133−
cells died within two weeks, and just a small number of
CD133− cells adhered to the wall and grew slowly.
Sphere-forming assay
The primary lung cancer cell suspension,
CD133− cells and CD133+ tumor spheres were
examined for their ability to form new spheres following initial
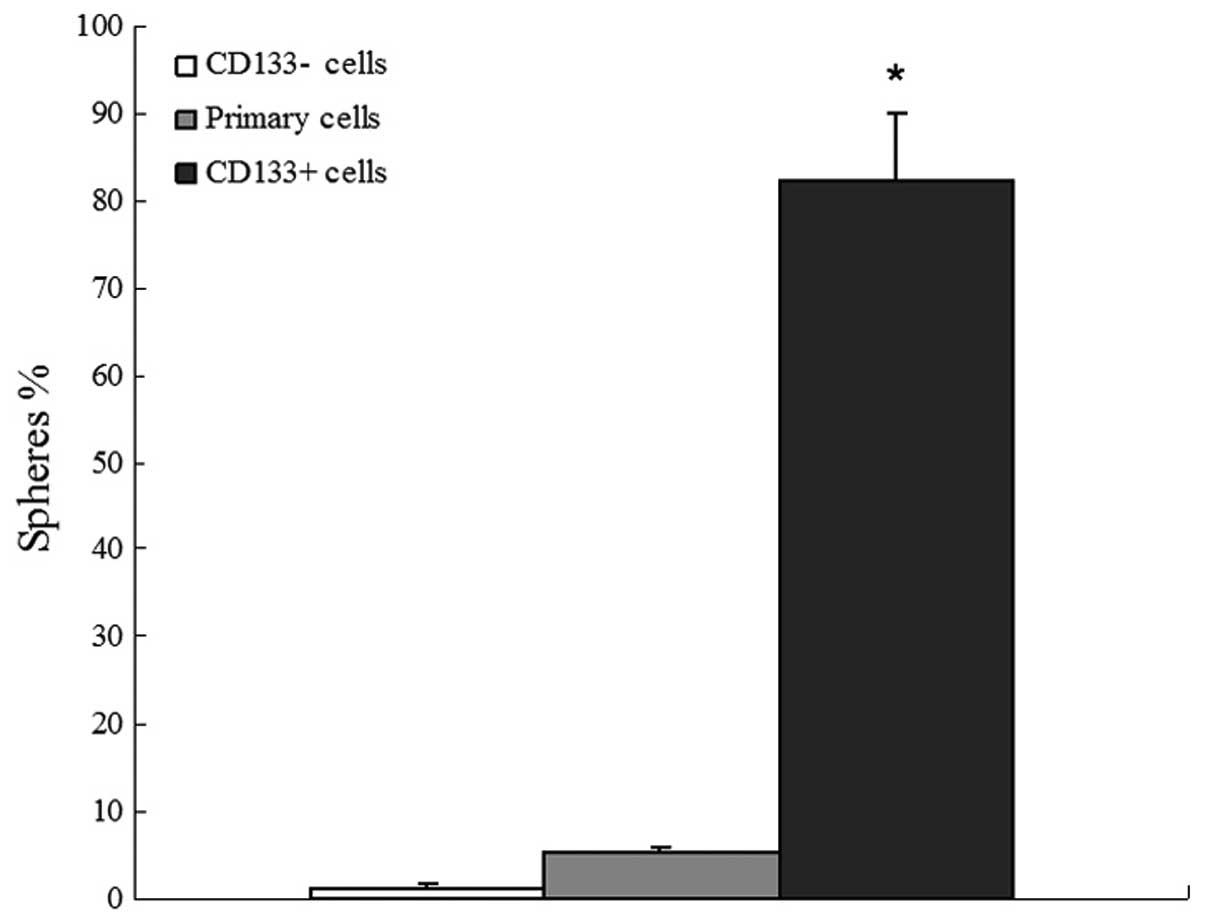
culturing as single cells. Subsequent to two weeks of culturing,
82.37±7.6% of single cell wells derived from CD133+
tumor spheres formed a new set of spheres; however, only 5.23±0.71%
of wells with a single cell derived from the primary lung cancer
cell suspension formed spheres. Additionally, almost no obvious
sphere colonies derived from CD133− cells were
identified (Fig. 3).
Differentiation of CD133+
cells
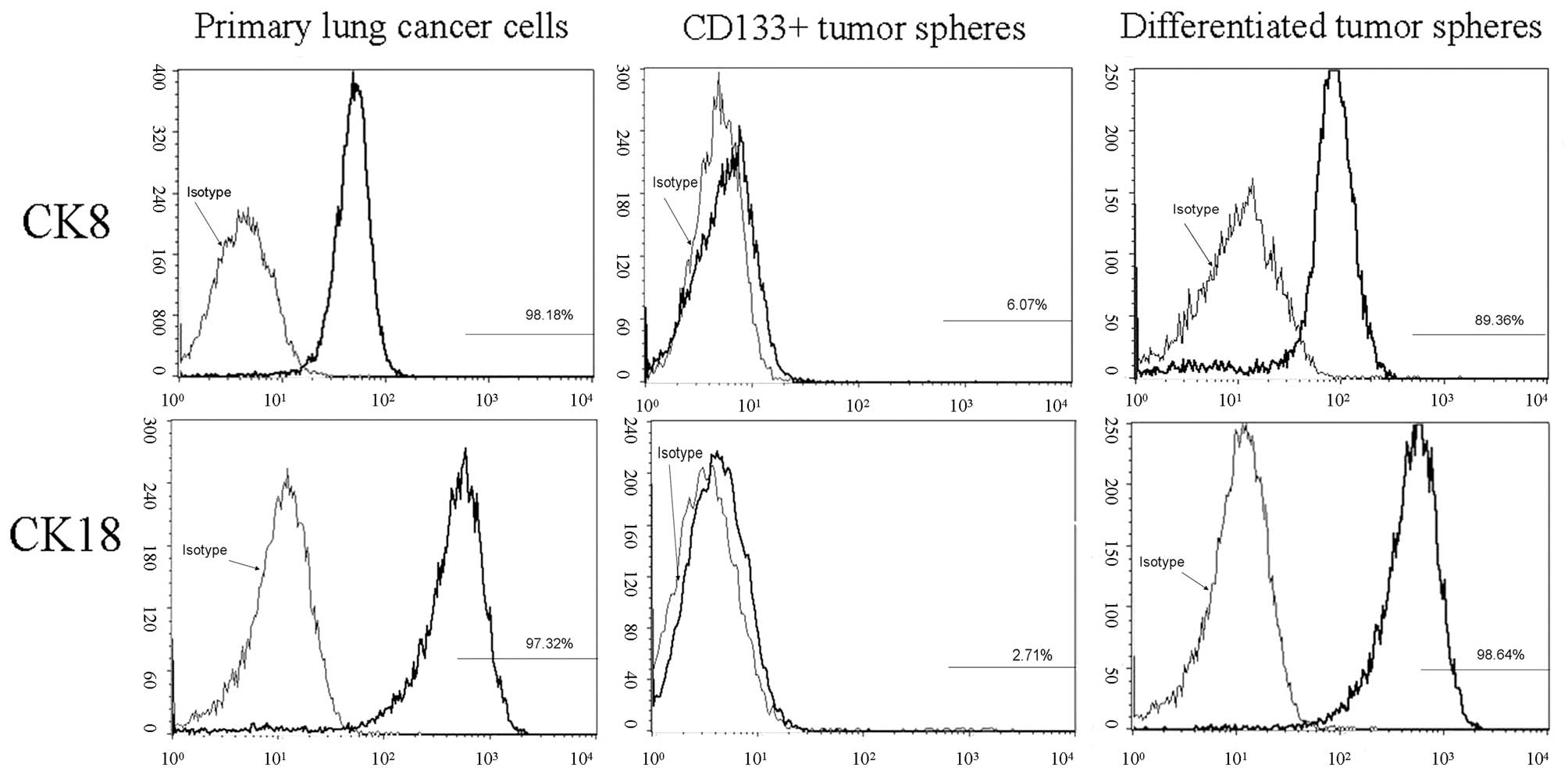
The expression of CK8 and CK18 in primary lung
cancer cells, and before and after differentiation of
CD133+ tumor spheres, was observed using flow cytometry.
CK8 and CK18 expression increased from 6.07±0.32 to 89.36±9.08%
(P<0.01) and 2.71±0.18 to 98.64±10.13% (P<0.01),
respectively, following cell adherence and tumor sphere
differentiation in the culture system supplemented with 10% FBS.
Similarly, the CK8 and CK18 expression of primary lung cancer cells
was 98.18±12.59 and 97.32±11.22%, respectively. This finding
indicates that CD133+ cells have the potential to
differentiate into various cell types (Fig. 4).
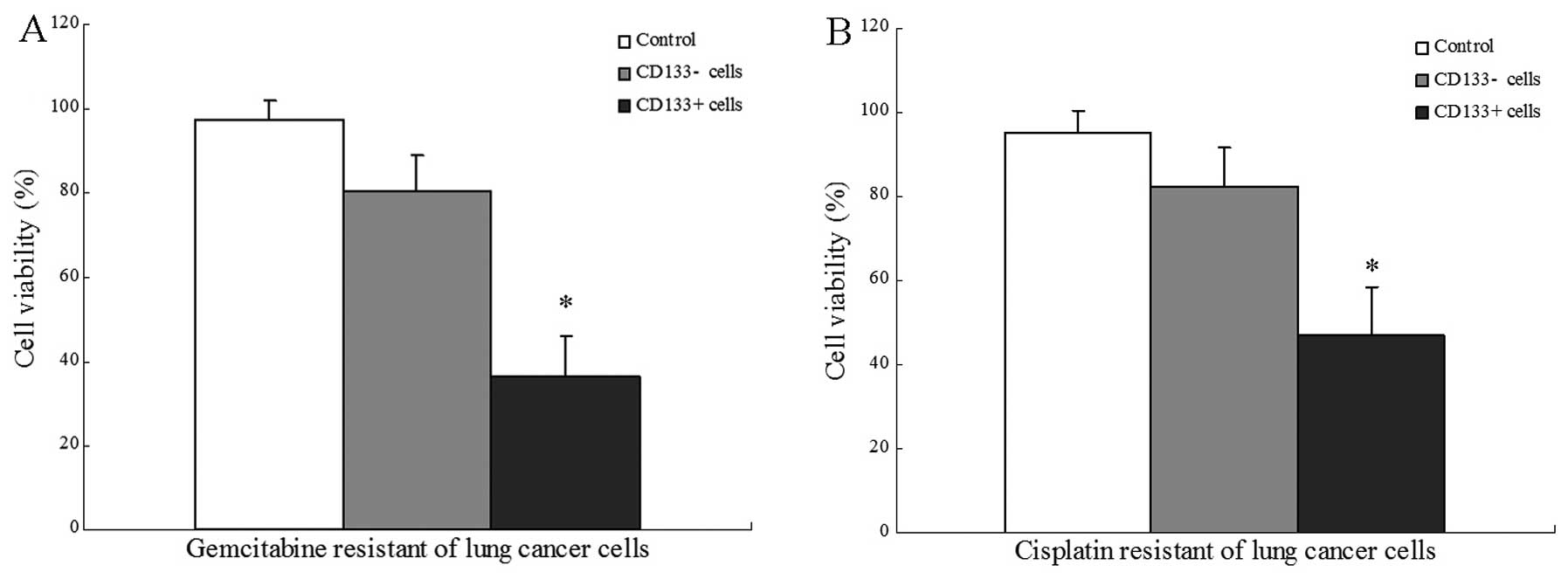
CD133+ are cells resistant to
conventional chemotherapy
Gemcitabine and cisplatin were administered at doses
comparable with the higher plasma levels obtained in treated
CD133+ and CD133− cells. The results
demonstrated that the gemcitabine (Fig.
5A) and cisplatin (Fig. 5B)
resistance of CD133+ cells was significantly stronger
than that of CD133− cells, in vitro.
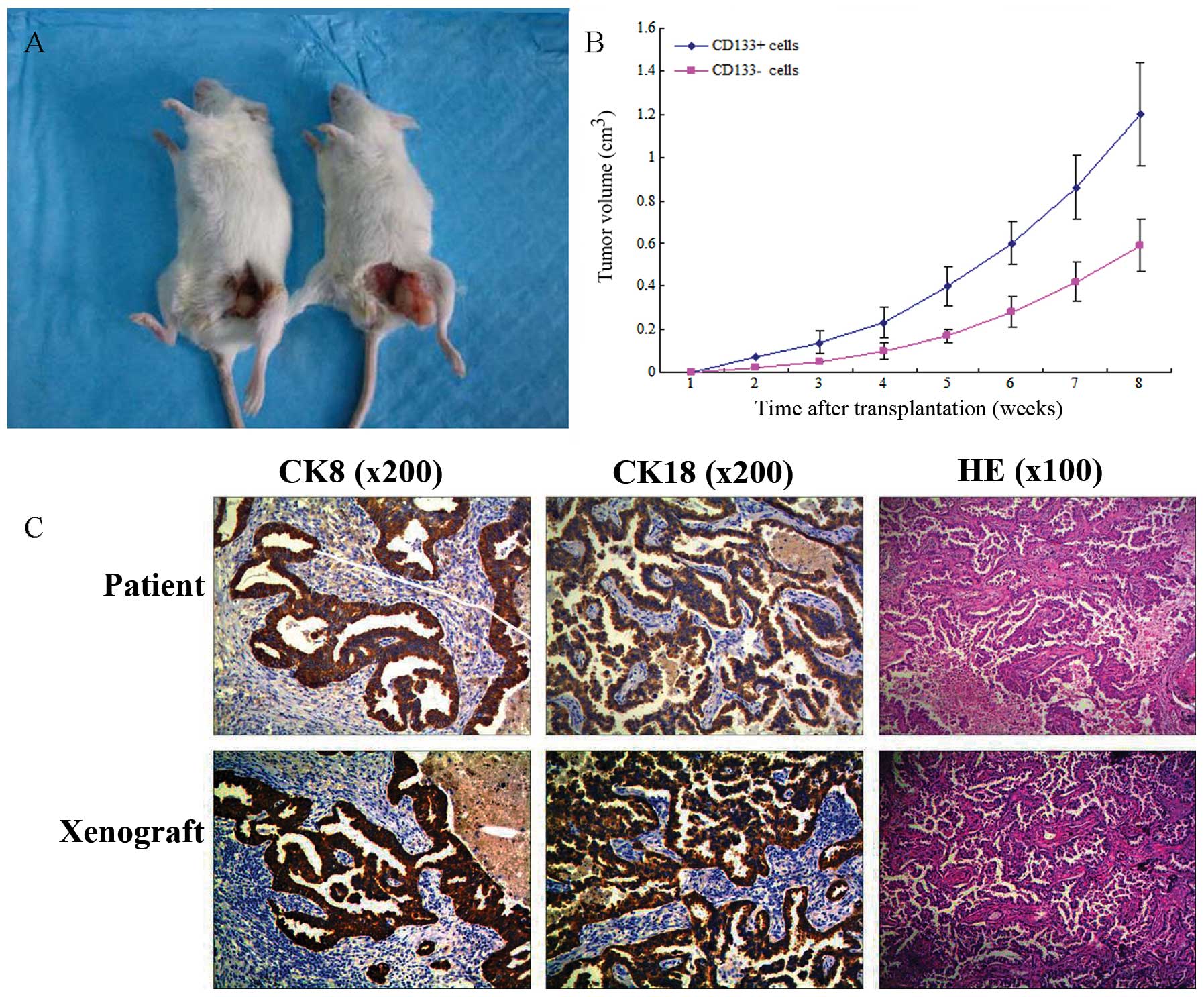
Tumorigenic potential of
CD133+ tumor spheres in NOD/SCID mice
Accumulating evidence indicates that CSCs exhibit
powerful tumorigenicity. To evaluate the hypothesis that
CD133+ tumor spheres are more tumorigenic due to their
enhanced stem-like properties, CD133+ tumor spheres and
CD133− cells were subcutaneously injected into NOD/SCID
mice in a limiting dilution assay (102, 103,
104 and 105 cells; Fig 6A). The results demonstrated that
CD133+ cells possess more powerful tumorigenicity
compared with CD133− cells, in vitro (Table II; Fig
6B). In order to further compare the size of the xenograft
tumor of the two groups cells in vitro, the tumors induced
by 105 CD133+ and CD133− cells
were observed; the tumor volume induced by CD133+ cells
was significantly greater than that induced by CD133−
cells. Additionally, H&E staining and immunohistochemical
markers indicated similar histology in the xenograft and primary
tumors. This finding indicates that CD133+ cells exhibit
powerful tumorigenicity in vitro (Fig. 6C).
| Table IIIncidence of tumors in non-obese
diabetic/severe combined immunodeficiency mice serially
transplanted with CD133+ and CD133−
cells. |
Table II
Incidence of tumors in non-obese
diabetic/severe combined immunodeficiency mice serially
transplanted with CD133+ and CD133−
cells.
| Cells, n |
|---|
|
|
|---|
| Transplanted
cells | 102 | 103 | 104 | 105 |
|---|
|
CD133− | 0/5 | 0/5 | 2/5 | 4/5 |
|
CD133+ | 2/5 | 4/5 | 5/5 | 5/5 |
Discussion
According to the CSC theory, lung cancer may be
described as a stem cell disease originating from the malignant
transformation of adult lung stem cells; such transformed adult
stem cells are also known as LCSCs. To date, three types of LCSC
have been reported. Ho et al (18) reported that LCSCs expressing ABC
membrane transporter proteins such as ABCG2, multidrug
resistance-associated protein 1 and multidrug resistance protein 1
were predominantly localized in SP cells, while Eramo et al
(19) reported that LCSCs are a
subset of CD133+ cells. By contrast, Dong et al
(21) identified that BASC-like
LCSCs were present in human pulmonary adenocarcinoma samples. These
cells were characterized by a
CD24+/IGF−IR+ phenotype and
expressed a variety of genes that comprise the backbone of
embryonic and lung stem cells; consequently, these cells were
highly invasive and tumorigenic.
Research into LCSCs is in the preliminary stages and
LCSCs are not yet commercialized; therefore, they are difficult to
purchase and studies of LCSCs generally require an initial
isolation step. There are currently four methods for the separation
or enrichment of CSCs: i) Separation of SP cells (18); ii) separation using flow cytometry
or magnetic beads with recognized cell surface markers (19); iii) suspension of the culture in
serum-free medium; and iv) selection based on the drug resistance
of LCSCs, which increases the purity of stem cells by promoting the
apoptosis of other SP cells through in vitro or in
vivo treatment with therapeutic agents (22). However, these four separation
methods have certain limitations. For example, Hoechst 33342 is
used to separate SP cells based on the characteristics of CSCs,
resulting in cellular toxicity, thus, restricting its further
application. In addition, no recognized cell surface markers of
LCSCs are currently available, therefore, the reliability of cell
surface marker separation using flow cytometry or magnetic beads is
yet to be confirmed. Furthermore, the sorting process may cause
damage to the cells and the number of sorted cells may be small due
to a low proportion of positive cells in the sorted tissues.
Moreover, suspension of the culture in serum-free medium alone may
only achieve preliminary enrichment of CSCs. Thus, other SP cells
may be present, resulting in a culture with reduced LCSC purity and
concentration that is unsuitable for further research.
Additionally, cell cultures are time-consuming and costly. Finally,
the CSC theory is in the exploratory stage; the process by which
LCSCs initiate tumorigenesis, as well as the molecular mechanisms
of their formation and existence, particularly CSC self-regulation
mechanisms, have yet to be investigated. Hence, the induction of
resistant cells may affect the biological properties of CSCs
(23–25).
Considering that the undifferentiated LCSCs are more
resistant to apoptosis and more able to withstand serum-free
conditions than human lung cancer cells, the present experiment
initially used flow cytometry to detect the proportion of
CD133+ cells in single cell suspensions of sorted lung
cancer tissue samples. Subsequently, the single cell suspensions
containing the highest proportion of CD133+ cells were
selected and the CD133+ cells were isolated using
immunomagnetic beads. Utilizing the characteristics of LCSCs, such
as resistance to serum-free culture conditions and apoptosis
tolerance, a serum-free suspension culture was used to enrich the
CD133+ cells in the population. Following greater than
one week of culturing, the cells were observed to form a ball. The
two techniques proved complementary, producing a novel method that
should be further investigated, due to the short culture period,
low cost and high efficiency.
The functions of the cultured cells were
predominantly used to determine whether they possessed the
characteristics of LCSCs. The present study identified that,
following enrichment, the CD133+ tumor spheres exhibited
strong self-renewal capacities compared with the CD133−
cells. In the serum-free culture, 82.3% of CD133+ cells
formed balls of stem cells, which was significantly different from
the CD133− cells (1.21%; P<0.01). In addition, the
CD133+ cells possessed multipotent differentiation
capacity. The expression patterns of CK8 and CK18 following cell
differentiation were similar to those observed in the primary cell
culture. CK8 and CK18 are important protein components of the
cytoskeleton, and are the most widely expressed members of the 21
intermediate filament types of epithelial and tumor cells (26). CK8 and CK18 function in the
determination and maintenance of cell morphology, and the
regulation of spatial organization of proteins in cell organelles
and the cytoplasm. Furthermore, CK8 and CK18 participate in cell
movement, cell division and cytoplasmic transport (27). Positive expression of CK8 and CK18
is a diagnostic indicator of pulmonary adenocarcinoma in the
pathological diagnosis of lung cancer (28). The results of the present study
demonstrated that chemotherapeutic agent resistance was
significantly higher in CD133+ cells compared with in
CD133− cells and, thus, is a characteristic of CSCs
(29).
The gold standard for determining whether LCSCs are
present in an enriched cell population is the tumorigenicity
experiment in NOD/SCID mice (19).
The present study used this method to demonstrate that
CD133+ cells exhibit higher tumorigenic capacities than
CD133− cells. The transplantation of only 100
CD133+ cells was sufficient to form tumors under the
abdominal wall of NOD/SCID mice. When the same numbers of
CD133+ and CD133− cells were transplanted
under the abdominal wall of NOD/SCID mice, the tumor volumes formed
by CD133+ cells were significantly greater than those
formed by CD133− cells.
In conclusion, the culture and identification of
LCSCs remains a controversial topic. To the best of our knowledge,
the present study is the first to use flow cytometry to detect the
proportion of CD133+ cells in single cell suspensions of
lung cancer tissue and to use immunomagnetic beads to select the
single cell suspensions with the highest proportions of
CD133+ cells. Used in combination with serum-free
suspension culture, this technique enriched the LCSC population
sufficiently to overcome the limitations of the existing separation
CSC separation techniques: The existence of other SP cells; the
time-consuming characteristic of serum-free suspension culture; and
the difficulties of immunomagnetic separation due to a low
proportion of positive cells in the sorted cell population. The
present method improves the speed of LCSC separation, resulting in
increased purity and concentration of LCSC. Furthermore, the
self-renewal, multipotent differentiation capacity, drug resistance
and tumorigenic capacity of LCSCs were verified experimentally,
providing a foundation for further theoretical research into
LCSCs.
References
|
1
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang L, Parkin DM, Ferlay J, Li L and Chen
Y: Estimates of cancer incidence in China for 2000 and projections
for 2005. Cancer Epidemiol Biomarkers Prev. 14:243–250.
2005.PubMed/NCBI
|
|
4
|
Zhou XD, Wang XY, Qu FJ, et al: Detection
of cancer stem cells from the C6 glioma cell line. J Int Med Res.
37:503–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qiu B, Zhang D, Tao J, Wu A and Wang Y: A
simplified and modified procedure to culture brain glioma stem
cells from clinical specimens. Oncol Lett. 3:50–54. 2012.PubMed/NCBI
|
|
6
|
Hwang-Verslues WW, Kuo WH, Chang PH, et
al: Multiple lineages of human breast cancer stem/progenitor cells
identified by profiling with stem cell markers. PLoS One.
4:e83772009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen W and Wang GM: Gene expression
profiling of cancer stem cells in the Du145 prostate cancer cell
line. Oncol Lett. 3:791–796. 2012.PubMed/NCBI
|
|
8
|
O’Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar
|
|
9
|
Suetsugu A, Nagaki M, Aoki H, Motohashi T,
Kunisada T and Moriwaki H: Characterization of CD133+
hepatocellular carcinoma cells as cancer stem/progenitor cells.
Biochem Biophys Res Commun. 351:820–824. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Varnat F, Duquet A, Malerba M, et al:
Human colon cancer epithelial cells harbour active HEDGEHOG-GLI
signalling that is essential for tumour growth, recurrence,
metastasis and stem cell survival and expansion. EMBO Mol Med.
1:338–351. 2009. View Article : Google Scholar
|
|
11
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sullivan JP, Minna JD and Shay JW:
Evidence for self-renewing lung cancer stem cells and their
implications in tumour initiation, progression, and targeted
therapy. Cancer Metastasis Rev. 29:61–72. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Otto WR: Lung epithelial stem cells. J
Pathol. 197:527–535. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rawlins EL and Hogan BL: Epithelial stem
cells of the lung: privileged few or opportunities for many?
Development. 133:2455–2465. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi Y, Fu X, Hua Y, Han Y, Lu Y and Wang
J: The side population in human lung cancer cell line NCI-H460 is
enriched in stem-like cancer cells. PLoS One. 7:e333582012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meng M, Zhao XH, Ning Q, Hou L, Xin GH and
Liu LF: Tumor stem cells: A new approach for tumor therapy
(Review). Oncol Lett. 4:187–193. 2012.PubMed/NCBI
|
|
17
|
Kim CF, Jackson EL, Woolfenden AE, et al:
Identification of bronchioalveolar stem cells in normal lung and
lung cancer. Cell. 121:823–835. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ho MM, Ng AV, Lam S and Hung JY: Side
population in human lung cancer cell lines and tumors is enriched
with stem-like cancer cells. Cancer Res. 67:4827–4833. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eramo A, Lotti F, Sette G, et al:
Identification and expansion of the tumorigenic lung cancer stem
cell population. Cell Death Differ. 15:504–514. 2008. View Article : Google Scholar
|
|
20
|
Groome PA, Bolejack V, Crowley JJ, et al:
IASLC International Staging Committee; Cancer Research and
Biostatistics; Observers to the Committee; Participating
Institutions: The IASLC Lung Cancer Staging Project: validation of
the proposals for revision of the T, N, and M descriptors and
consequent stage groupings in the forthcoming (seventh) edition of
the TNM clas-sification of malignant tumors. J Thorac Oncol.
2:694–705. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong QG, Yao M, Geng Q, Zhou J and Yan MX:
Isolation and identification of human lung adenocarcinoma stem
cells. Tumor. 28:1–7. 2008.(In Chinese).
|
|
22
|
Bertolini G, Roz L, Perego P, et al:
Highly tumorigenic lung cancer CD133+ cells display stem-like
features and are spared by cisplatin treatment. Proc Natl Acad Sci
USA. 106:16281–16286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dean M: ABC transporters, drug resistance,
and cancer stem cells. J Mammary Gland Biol Neoplasia. 14:3–9.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sung JM, Cho HJ, Yi H, et al:
Characterization of stem cell population in lung cancer A549 cells.
Biochem Biophys Res Commun. 371:163–167. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Levina V, Marrangoni AM, DeMarco R,
Gorelik E and Lokshin AE: Drug-selected human lung cancer stem
cells: cytokine network, tumorigenic and metastatic properties.
PLoS One. 3:e30772008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Omary MB, Coulombe PA and McLean WH:
Intermediate filament proteins and their associated diseases. New
Engl J Med. 351:2087–2100. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fukunaga Y, Bandoh S, Fujita J, et al:
Expression of cytokeratin 8 in lung cancer cell lines and
measurement of serum cytokeratin 8 in lung cancer patients. Lung
Cancer. 38:31–38. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kanaji N, Bandoh S, Fujita J, Ishii T,
Ishida T and Kubo A: Compensation of type I and type II cytokeratin
pools in lung cancer. Lung Cancer. 55:295–302. 2007. View Article : Google Scholar
|
|
29
|
Ghotra VP, Puigvert JC and Danen EH: The
cancer stem cell microenvironment and anti-cancer therapy. Int J
Radiat Biol. 85:955–962. 2009. View Article : Google Scholar : PubMed/NCBI
|