Introduction
Sulfur is bright yellow crystalline solid or powder
material. In traditional Chinese medicine (TCM), sulfur is widely
used for detoxifying the body and the treatment of scabies
(1,2), healing sores and itching (3). It was confirmed by United States
Pharmacopeia that sublimed sulfur and subsided sulfur may be used
as drugs (4). When absorbed by the
skin, sulfur is metabolized to inorganic sulfide or organic
sulfocompounds, and it is involved in metabolism in vivo
(5). Recently, studies have
reported that sulfocompounds inhibit cancer cell growth. For
example, GYY4137, a sulfocompound, caused the
concentration-dependent killing of various cancer cell lines
(6), and S-propargyl-cysteine
exhibited anticancer effects on gastric cancer cells (7). Furthermore, Allicin, which contains
diallyl disulfide, reduced the risk of a variety of malignant
tumors, including different glioblastoma cells (8) and hepatocellular carcinoma (9) in vitro, and osteosarcoma cells
in vitro and in vivo (10,11).
Prostate cancer (PCa) is the most common malignancy
of the male genitourinary system. Since Huggins first treated PCa
by androgen deprivation treatment (ADT) (12), ADT has been widely used for PCa
treatment. Initially, PCa responds to androgen ablation and disease
progression slows down. However, the androgen ablation is
palliative, and the disease eventually ensues (13). PCa recurs and the surviving cancer
cells become androgen-independent. At this stage, tumors are more
aggressive and usually fatal, and hormone blockade therapy fails
(14). At present, no effective
therapy method has been identified for recurrent PCa and only
traditional treatments are primarily used to provide symptomatic
benefits, and identifying new methods to treat androgen-independent
PCa has been a new area of focus for researchers (15). Recently, the pathways meditated by
an androgen receptor (AR) or to bypass ARs have been researched in
recurrent PCa, additionally treatment pathways and novel therapies
have been investigated to PCa therapy with this potential target
Recently, the pathways meditated by an androgen receptor (AR) or to
bypass ARs have been researched in recurrent PCa (16,17).
Additionally treatment pathways and novel therapies have been
investigated for PCa therapy with this potential target (18,19).
22Rv1 is an androgen-independent PCa epithelial cell line, which
grows independently of androgen, and is representative of clinical
recurrent PCa. However, 22Rv1 cells express the AR and react with
androgens (20). Previous studies
have shown that the progression of androgen-independent PCa is
independent of the androgen, but dependent on the AR signaling
pathway (21). 22Rv1 cells also
secrete prostate-specific antigen (PSA) (22), which is often used to evaluate the
efficiency of PCa treatment. DU-145 and PC-3 cell lines are also
androgen-independent PCa cell lines and, thus, are often used in
the study of PCa. The two cell lines do not express AR and PSA
(23), while the majority of
clinical PCa cases significantly express the two genes.
In the present study, 22Rv1 and DU-145 prostate
tumors were develoepd in nude mice and the aim of the study was to
investigate the inhibitory effect of sulfur on prostate cancer
cells.
Materials and methods
Drugs and animals
Sulfur powder with a purity of ≥99% was purchased
from the Shanghai Chemical Reagent supply station and mixed with
milk powder (Nestlé S.A., Vevey, Switzerland) at 1:30 (w/w).
Specific pathogen-free (SPF) male BALB/c nude mice
aged between six and eight weeks (weight range, 18–25 g) were
purchased from Shanghai SLAC Laboratory Animal Co., Ltd., [License
Number, SCXK (Shanghai) 2007–0002; Shanghai, China)], and fed under
SPF conditions. Mice in the 22Rv1 and DU-145 experiments were
randomly divided into two groups, control and sulfur-treated
groups, with 10 mice in each group. Ethical approval for the study
was obtained from the Animal Ethics committee of Shanghai Institute
of Planned Parenthood Research (Shanghai, China).
Cell culture
22Rv1 cells were purchased from Shanghai Institute
of Cell Life Sciences Resource Center (Shanghai, China). The cells
were maintained in RPMI-1640 medium (Hyclone; Thermo Fisher
Scientific, Rockford, IL, USA) containing 50 ml/l fetal bovine
serum (FBS; Gibco-BRL, Carlsbad, CA, USA), 100,000 units/l
penicillin and 100 mg/l streptomycin, at 37°C with an atmosphere of
5% CO2.
Xenograft tumor development in nude
mice
22Rv1 cells and DU-145 cells were harvested at the
exponential growth stage, washed and suspended in
phosphate-buffered saline (PBS). The cells were counted, then the
cell suspension was subcutaneously injected into the flanks of mice
with 2×106 cells in 0.1 ml PBS. A Trypan-blue exclusion
assay was performed to ensure cell viability (>99%) prior to
inoculation.
Each mouse in the sulfur-treated group was provided
with 0.62 g/day sulfur-milk powder one day following inoculation
with 22Rv1 and DU-145 cells, while mice in the control group were
provided with milk powder. Tumor size was measured in two diameters
every other day (22Rv1 tumor) or every week (DU-145 tumor). Tumor
volume (cm3) was calculated using the following formula:
Tumor volume = a*b2/2 (a, longer diameter; b,
shorter diameter).
Following the experiment, mice with 22Rv1 tumors
were narcotized with 0.2 ml 1% sodium pentobarbital and blood was
then collected from the heart. Tumors were dissected and
weighed.
Cells separation from 22Rv1 tumors
22Rv1 tumors were cut into sections between 1 and 2
mm3, and tumor sections in the same group were mixed,
washed with cold PBS containing 200 units/ml penicillin and 200
mg/l streptomycin, digested with trypsin for 5 min at 37°C, and
then the digested cells were suspended by RPMI-1640 medium with 5%
FBS. The cell suspension was filtrated using a 200-mesh sieve, and
then centrifuged at 175 × g for 5 min. The supernatant was then
discarded and the cells were suspended with fresh medium, and
incubated at 37°C in an atmosphere of 5% CO2.
Clone-forming capability of tumor
cells
A total of 300 tumor cells from the sulfur-treated
and control groups were seeded, respectively, into 24-well plates
in RPMI-1640 medium and incubated for two weeks. The medium was
then discarded, cell clones were washed three times with PBS, fixed
with 5% formaldehyde for 20 min, stained with crystal violet
solution for 5 min, washed for several times, dried and
counted.
The cell growth of the tumor cells was measured
using the MTT method demonstrated by Meletiadis et al
(21).
Analysis of gene expression by
quantitative polymerase chain reaction (qPCR)
Total RNA was extracted using TRIzol reagent
(Invitrogen Life Technologies, Inc., Carlsbad, CA, USA) according
to the manufacturer’s instructions. Total RNAs in the same group
were mixed in the same quantity. First-strand cDNA was synthesized
from 2 μg of RNA mixture using Quant Reverse Transcriptase (Toyobo
Co., Ltd., Osaka, Japan). Gene expression in 22Rv1 tumors was
measured using qPCR. The sequences of the primers used are shown in
Table I.
 | Table IPrimers used for qPCR. |
Table I
Primers used for qPCR.
| Gene | Primer sequences | Annealing temperature
(°C) | Product length
(bp) |
|---|
| β-actin | F:
5′-CCTGTACGCCAACACAGTGC-3′
R: 5′-ATACTCCTGCTTGCTGATCC-3′ | 58 | 211 |
| AR | F:
5′-TTCCCTCCCTATCTAACCCTC-3′
R: 5′-TCTAAACTTCCCGTGGCATAA-3′ | 58 | 202 |
| PSA | F:
5′-AGTCTGCGGCGGTGTTCT-3′
R: 5′-GTGGCTGACCTGAAATACCTG-3′ | 58 | 139 |
| NKX3.1 | F:
5′-AGAAAGGCACTTGGGGTCTT-3′
R: 5′-TCCGTGAGCTTAGGTTCTT-3′ | 60 | 210 |
Hydrogen sulfide (H2S) levels
in mice serum
Blood was collected from the hearts of the mice and
the serum was separated. H2S levels in the serum were
analyzed using the mouse serum H2S Elisa kit according
to the manufacturer’s instructions (Shanghai Feng Xiang Biological
Technology Co., Ltd., Shanghai, China).
Statistical analysis
Data were analyzed using SPSS software, version 11.5
(SPSS, Inc., Chicago, IL, USA). The continuous variables of tumor
weight, tumor volume and H2S level were compared using
one-way analysis of variance. Consequently, the analyzed results
are presented as the mean ± standard deviation (SD). In addition,
the clones were also presented as the mean ± SD. P<0.05 was
considered to indicate a statistically significant difference.
Results
Sulfur inhibits the growth of 22Rv1 and
DU-145 tumors
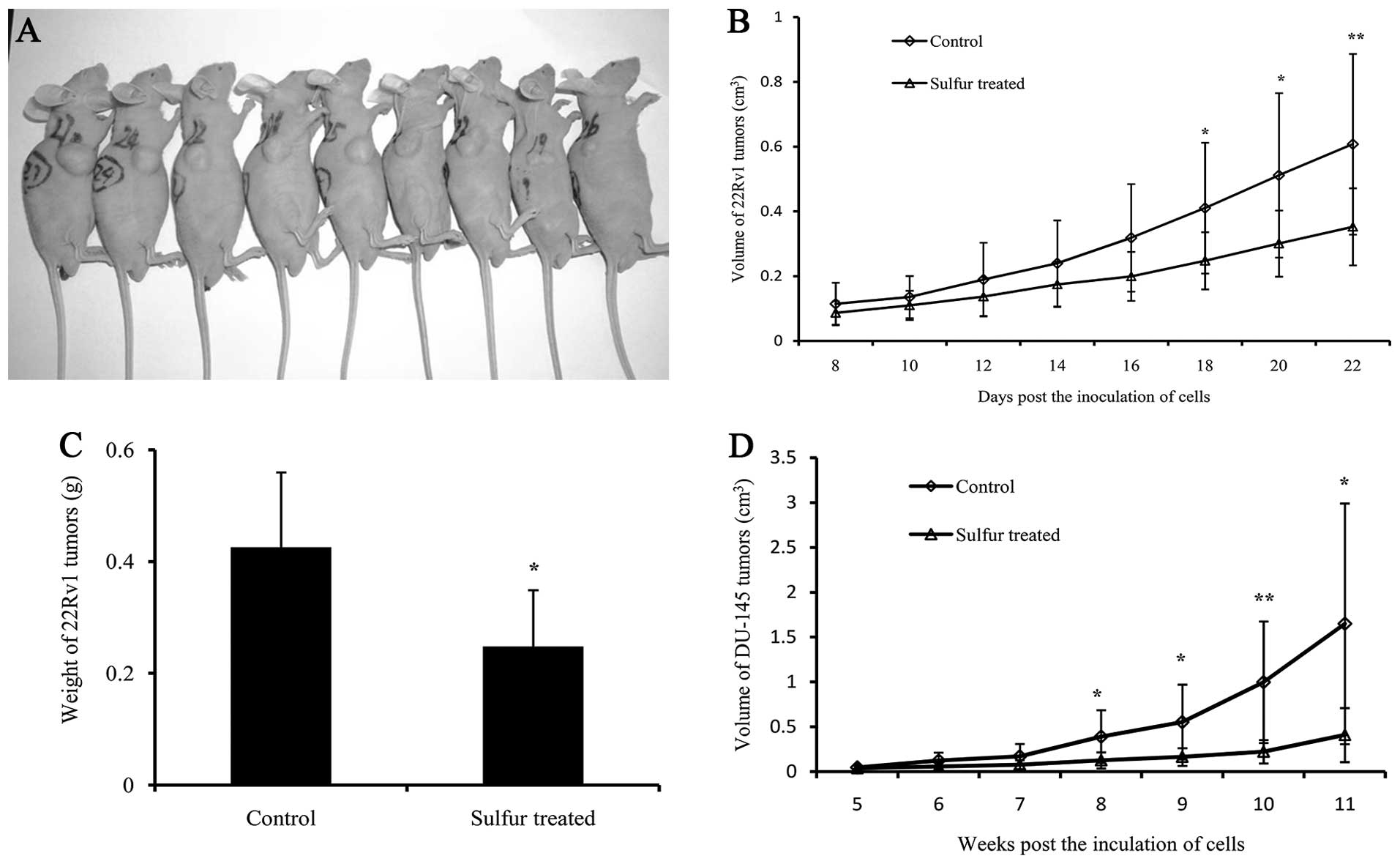
Approximately eight days following inoculation of
the 22Rv1cells, the tumor sizes were measurable (Fig. 1A). The growth of the tumors obtained
from sulfur-treated mice gradually slowed down, and a significant
difference was identified 18 days following inoculation when
compared with that of the control group. The mean tumor volume of
the sulfur-treated group was 0.35±0.12 cm3 after 22
days, which was 41.99% smaller than that of the control group
(0.61±0.28 cm3) (P<0.01; Fig. 1B). The mean tumor weight of the
sulfur-treated group was 0.25±0.10 g, which was significantly
decreased by 41.78% when compared with that of the control group
(0.43±0.13 g) (P<0.05; Fig. 1C).
The mean body weights of the tumor-bearing mice were 21.11±1.62 and
24.91±1.56 g in the sulfur-treated and control groups,
respectively. However, no significant difference in body weight was
identified. The results demonstrated that sulfur significantly
inhibits the growth of 22Rv1 tumors in vivo.
DU-145 tumors were measurable five weeks following
inoculation, and the growth of tumors of the sulfur-treated group
gradually slowed down and a significant difference was identified
eight weeks after inoculation, when compared with that of the
control group. The mean tumor volume of the sulfur-treated group
was 0.41±0.30 cm3 11 weeks after inoculation, which was
75.16% smaller than that of the control group (1.64±1.34
cm3) (P<0.05; Fig.
1D). The results demonstrated that sulfur significantly
inhibits androgen-independent prostate tumor growth.
Since 22Rv1 cells, but not DU-145 cells, express AR
and PSA, 22Rv1 tumors were selected for further study.
Sulfur decreases the clonogenicity of
22Rv1 tumor cells
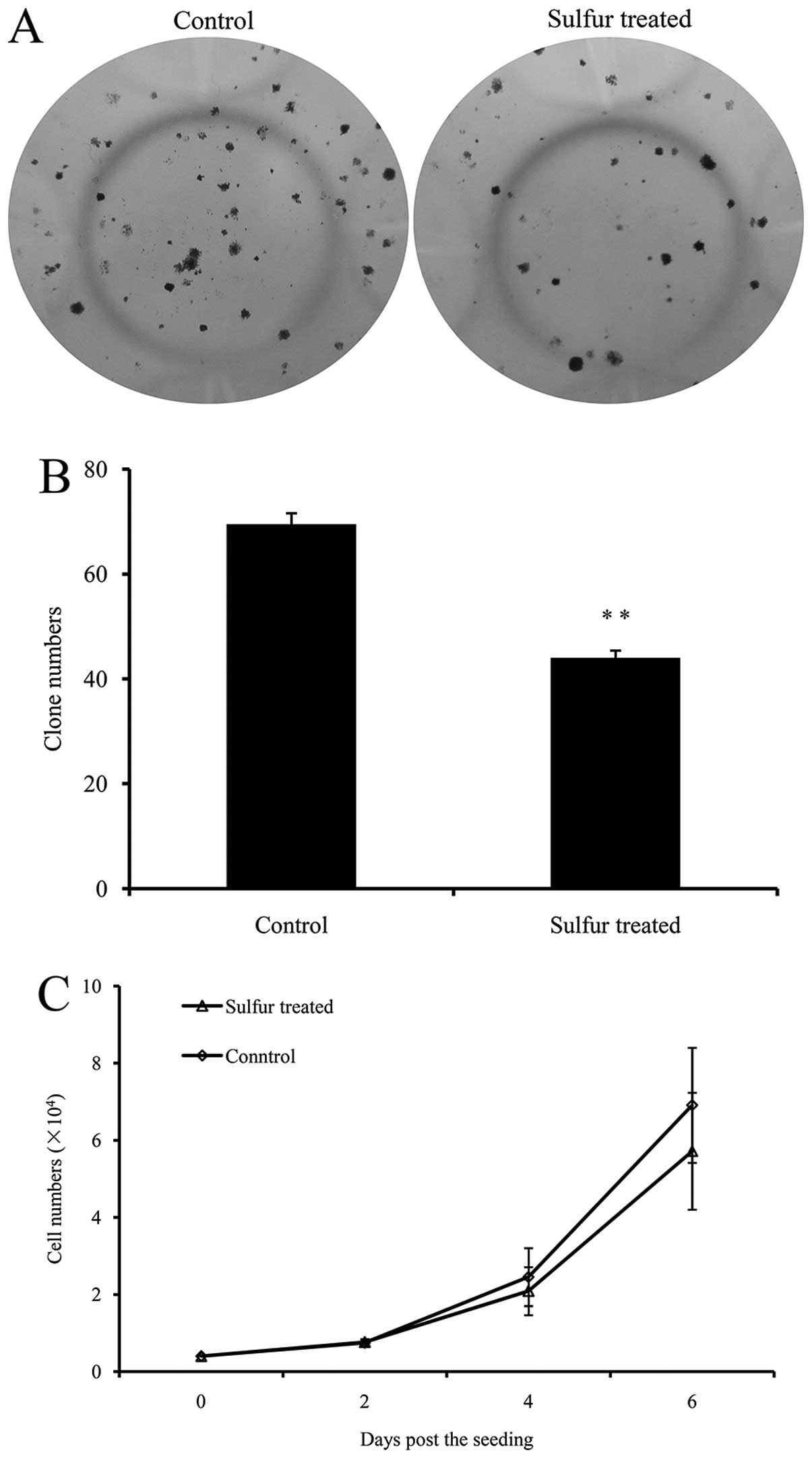
In order to further investigate the inhibitory
effects of sulfur on PCa tumor growth in vivo, 22Rv1 cells
were separated from the xenograft tumors, and maintained in culture
for at least two weeks prior to their application in the following
approach. A total of 200 cells from each group were seeded in
24-well plates, respectively, and incubated to form clones
(Fig. 2A). Cells from the
sulfur-treated tumors formed 44.00±1.41 clones per well, which was
36.69% less than that of the control tumors (69.50±2.12 clones per
well) (Fig. 2B), indicating that
the clonogenicity of 22Rv1 PCa cells was significantly decreased by
sulfur.
In addition, the inhibition of 22Rv1 tumor growth
was maintained when sulfur was removed. The MTT method was used to
measure the growth rate of 22Rv1 tumor cells; however, no
significant difference was identified between cells from the two
groups (Fig. 2C).
Sulfur inhibits the expression of AR, PSA
and NKX3.1 in 22Rv1 tumors
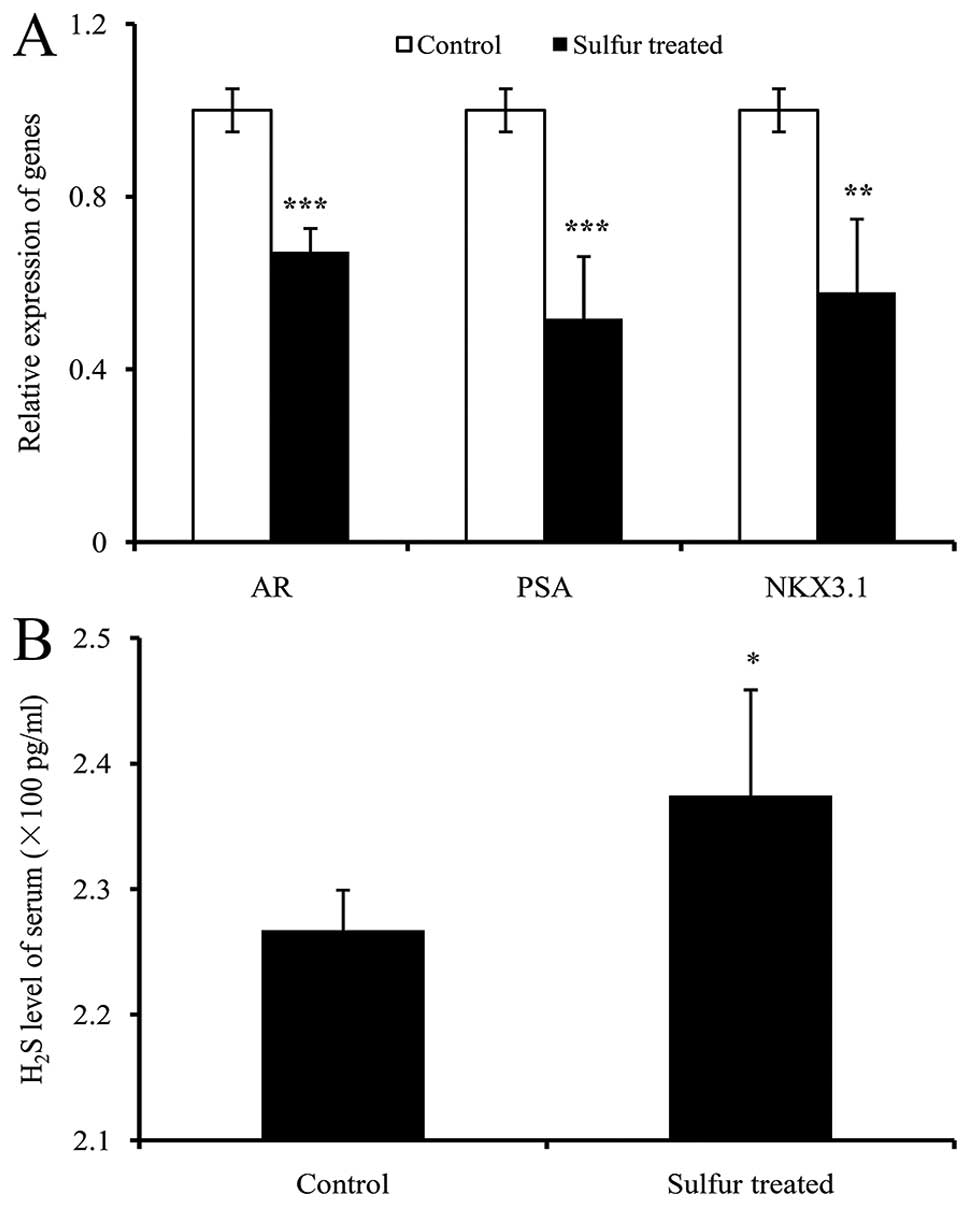
AR is involved in PCa progression (21), and is expressed in 22Rv1 cells, in
addition to AR-regulated genes PSA (25) and NKX3.1 (26). The expression of these genes was
analyzed using qPCR. As shown in Fig.
3, the expression of AR, PSA and NKX3.1 were significantly
decreased in sulfur-treated tumors by 32.8, 48.2 and 42.2%,
respectively (Fig. 3A). These
results indicated that the downregulation of the AR signaling
pathway contributes to the inhibitory effect of sulfur on PCa
growth in vivo.
Sulfur marginally increases
H2S levels in mice serum
Mice serum H2S levels were measured by
ELISA. The H2S level in sulfur-treated mice was
marginally increased by 4.73% (237.46±8.40 pg/ml) in the
sulfur-treated group compared with 226.74±3.18 pg/ml in the control
group (Fig. 3B), and a significant
difference was identified. The results indicated that sulfur
marginally increases H2S levels in mice serum.
Discussion
The present study demonstrated that sulfur
significantly inhibits PCa growth in xenograft models (Fig. 1). In previous studies, inorganic
sulfur has been demonstrated to inhibit the cell proliferation of
breast cancer cells in vitro when dissolved in methanol,
which is toxic to the body and difficult to use in vivo
(27). As sulfur is not dissolved
in water saline solution, it cannot be used directly for the
evaluation of its anticancer effects in cell culture. In the
present study, clone-forming analysis of cells separated from
tumors also demonstrated the inhibitory effect of sulfur on cancer
cells (Fig. 2A), and possibly
contributed to its inhibitory effect observed in vivo.
However, no significant inhibitory effect of sulfur was identified
on the growth of tumor cells in the present study. The anticancer
effect of sulfur may be temporary, as the growth of tumor cells was
inhibited only in the environment of sulfur in vivo. In our
study, tumors were removed from the sulfur-treated mice and the
anticancer effect of sulfur was diminished. However, cells with a
lower growth rate may be eliminated during long term culturing,
such as in the present study.
The androgen-receptor is involved in PCa progression
(21), and is an important target
of numerous drugs used for PCa therapy. The present study
demonstrated that sulfur significantly inhibited AR expression in
22Rv1 prostate tumors. Possibly as a result of reduced AR
expression, the expression of PSA and NKX3.1, two AR-targeted
genes, were also inhibited by sulfur. These results indicated that
the downregulation of the AR signaling pathway was involved with
the anticancer effect of sulfur. However, sulfur also markedly
inhibits the growth of PCa tumors in an AR-independent way, since
it also inhibited the growth of DU-145 tumors, which lacked AR
expression. Novel approaches are required for understanding the
AR-independent mechanism of the anticancer effect of sulfur.
As previously reported, a number of sulfocompounds
inhibited the growth of various cancer cells (6–11). The
anticancer capabilities were dependent (6,7) or
independent (8–11) on their ability to release
H2S. In the current study, only a marginally increased
level of H2S was detected in the serum of sulfur-treated
mice, indicating that H2S was not the key intermediate
metabolite of sulfur. The characterization of metabolites of sulfur
that exhibit anticancer activity may aid the development of
anti-PCa drugs.
In conclusion, the present study revealed that
sulfur significantly inhibited the growth of androgen-independent
tumors. Since sulfur has been used in the clinical treatment of
various diseases and is also a component of TCM, this study
supports the application of sulfur in the treatment of clinical
PCa, particularly recurrent PCa.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 81270760) and the National
Basic Research Program of China (grant no. 2014CB943104).
References
|
1
|
Pruksachatkunakorn C, Damrongsak M and
Sinthupuan S: Sulfur for scabies outbreaks in orphanages. Pediatr
Dermatol. 19:448–453. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kenawi MZ, Morsy TA, Abdalla KF and el
Hady HM: Treatment of human scabies by sulfur and permethrin. J
Egypt Soc Parasitol. 23:691–696. 1993.PubMed/NCBI
|
|
3
|
Gupta AK and Nicol K: The use of sulfur in
dermatology. J Drugs Dermatol. 3:427–431. 2004.PubMed/NCBI
|
|
4
|
United States Pharmacopeia. USP35-NF30.
The Unites States Pharmacopeial Convention. 4727:2012.
|
|
5
|
Parcell S: Sulfur in human nutrition and
applications in medicine. Altern Med Rev. 7:22–44. 2002.PubMed/NCBI
|
|
6
|
Lee ZW, Zhou J, Chen CS, et al: The
slow-releasing hydrogen sulfide donor, GYY4137, exhibits novel
anti-cancer effects in vitro and in vivo. PLoS One. 6:e210772011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma K, Liu Y, Zhu Q, et al: H2S donor,
S-propargyl-cysteine, increases CSE in SGC-7901 and cancer-induced
mice: evidence for a novel anti-cancer effect of endogenous H2S?
PLoS One. 6:e205252011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Das A, Banik NL and Ray SK: Garlic
compounds generate reactive oxygen species leading to activation of
stress kinases and cysteine proteases for apoptosis in human
glioblastoma T98G and U87MG cells. Cancer. 110:1083–1095. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang ZM, Zhong N, Gao HQ, et al: Inducing
apoptosis and upregulation of Bax and Fas ligand expression by
allicin in hepatocellular carcinoma in Balb/c nude mice. Chin Med J
(Eng). 119:422–425. 2006.
|
|
10
|
Zeng JX, Sun JJ, Mo JC, Ma Y and He XS:
Inhibition of Allicin on Growth of Neuroblastoma in Vitro and in
Vivo [J]. Journal of Sun Yat-Sen University (Medical Sciences).
3:0092009.
|
|
11
|
Karmakar S, Banik NL, Patel SJ and Ray SK:
Garlic compounds induced calpain and intrinsic caspase cascade for
apoptosis in human malignant neuroblastoma SH-SY5Y cells.
Apoptosis. 12:671–684. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huggins C and Hodges CV: Studies on
prostatic cancer: I. The effect of castration, of estrogen and of
androgen injection on serum phosphatases in metastatic carcinoma of
the prostate. 1941. J Urol. 168:9–12. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kozlowski JM, Ellis WJ and Grayhack JT:
Advanced prostatic carcinoma. Early versus late endocrine therapy.
Urol Clin North Am. 18:15–24. 1991.PubMed/NCBI
|
|
14
|
Denmeade SR and Isaacs JT: A history of
prostate cancer treatment. Nat Rev Cancer. 2:389–396. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Petrylak DP: Current state of
castration-resistant prostate cancer. Am J Manag Care. 19:358–365.
2013.
|
|
16
|
Chi KN, Bjartell A, Dearnaley D, et al:
Castration-resistant prostate cancer: from new pathophysiology to
new treatment targets. Eur Urol. 56:594–605. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Debes JD and Tindall DJ: Mechanisms of
androgen-refractory prostate cancer. N Eng J Med. 351:1488–1490.
2004. View Article : Google Scholar
|
|
18
|
Hartel A, Didier A, Pfaffl MW and Meyer
HH: Characterisation of gene expression patterns in 22RV1 cells for
determination of environmental androgenic/antiandrogenic compounds.
J Steroid Biochem Mol Biol. 84:231–238. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim J and Coetzee GA: Prostate specific
antigen gene regulation by androgen receptor. J Cell Biochem.
93:233–241. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sramkoski RM, Pretlow TG II, Giaconia JM,
et al: A new human prostate carcinoma cell line, 22Rv1. In Vitro
Cell Dev Biol Anim. 35:403–409. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ware JL, Paulson DF and Webb KS: 1,
10-Phenanthroline reversibility inhibits proliferation of two human
prostate carcinoma cell lines (PC-3 and DU145). Biochem Biophys Res
Commun. 124:538–543. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Meletiadis J, Meis JF, Mouton JW, Donnelly
JP and Verweij PE: Comparison of NCCLS and
3-(4,5-dimethyl-2-thiazyl)-2, 5-diphenyl-2H-tetrazolium bromide
(MTT) methods of in vitro susceptibility testing of filamentous
fungi and development of a new simplified method. J Clin Microbiol.
38:2949–2954. 2000.PubMed/NCBI
|
|
23
|
Sardana G, Jung K, Stephan C and Diamandis
EP: Proteomic analysis of conditioned media from the PC3, LNCaP,
and 22Rv1 prostate cancer cell lines: discovery and validation of
candidate prostate cancer biomarkers. J Proteome Res. 7:3329–3338.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sciavolino PJ, Abrams EW, Yang L,
Austenberg LP, Shen MM and Abate-Shen C: Tissue-specific expression
of murine Nkx3.1 in the male urogenital system. Dev Dyn.
209:127–138. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ha AW, Hong KH, Kim HS and Kim WK:
Inorganic sulfur reduces cell proliferation by inhibiting of ErbB2
and ErbB3 protein and mRNA expression in MDA-MB-231 human breast
cancer cells. Nutr Res Pract. 7:89–95. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qin X, Zhang H, Yao X, et al: Variations
in circulating sex steroid levels in metastatic prostate cancer
patients with combined androgen blockade: observation and
implication. Andrology. 1:512–516. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nishimura S, Hato M, Hyugaji S, et al:
Glycomics for drug discovery: metabolic perturbation in
androgen-independent prostate cancer cells induced by unnatural
hexosamine mimics. Angew Chem Int Ed Engl. 51:3386–3390. 2012.
View Article : Google Scholar : PubMed/NCBI
|