Introduction
In eukaryotic species, the actin cytoskeleton is
essential for numerous cellular functions, including maintenance of
morphology, motility, division, adhesion, endocytosis,
intracellular transport and signal transduction (1–5). The
varied and complex activities of the actin cytoskeleton are
dynamically regulated by actin-binding proteins (ABPs) (2–4,6,7). Gelsolin
(GSN) is one of the most abundant ABPs, and has been found to be a
multifunctional regulator of physiological and pathological
cellular processes (7,8).
Previous studies have indicated that GSN may be a
tumor suppressor that exerts a crucial role in the carcinogenic
process (7,9). However, biphasic expression of GSN in
oral precancerous lesions and oral cancers has been observed, which
revealed a downregulation in GSN between oral precancerous lesions
and oral cancers, and demonstrated upregulation of GSN in the
stages of oral cancer progression (8). The biphasic expression indicated that
GSN may perform a more complicated role in oral cancer biology.
In order to study the biological roles in oral
cancer development in the present study, GSN was overexpressed in
oral cancer Tca8113 cells, and the effect of GSN on the
proliferation, apoptosis, cell cycle, migration and invasion of
these cells was investigated, which may contribute to the present
understanding of the biological actions of GSN.
Materials and methods
Tca8113 cell culture
The human oral squamous cell carcinoma Tca8113 cell
line was provided by the Shanghai Ninth People's Hospital, Medical
School of Shanghai Jiao Tong University (Shanghai, China). The
cells were cultured in RPMI 1640 medium (Gibco-BRL, Carlsbad, CA,
USA) containing 10% fetal bovine serum (FBS), 100 µg/ml
streptomycin and 100 units/ml penicillin, at 37°C in a 5%
CO2 atmosphere.
Stable transfection
To transiently transfect GSN into the Tca8113 cells,
0.5×105 cells/well were seeded into a 24-well plate
(Corning, Inc., Corning, NY, USA) one day prior to transfection
with 0.5 µg plasmid DNA, using the GBfectene-Elite Transfection
Reagent (Genebank Biosciences, Inc., Zhangjiagang, Jiangsu, China).
The transfection rate was monitored by a fluorescence microscope
(Eclipse Ti; Nikon Corporation, Tokyo, Japan). A total of 400 µg/ml
G418 (Invitrogen Life Technologies, Carlsbad, CA, USA) was applied
to the stable transfectants, and the level of GSN expression was
determined by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and western blotting. β-actin and glyceraldehyde
3-phosphate dehydrogenase were used as reference genes for RT-qPCR
and western blotting, respectively. Cells transfected with the
empty pcDNA3.1 vector served as negative controls. The Tca8113
cells transfected with the pcDNA3.1-GSN and pcDNA3.1 vectors were
respectively termed Tca8113-GSN and Tca8113-Vec.
Proliferation assay
The cell counting kit-8 (CCK-8) proliferation assay
(Dojindo Molecular Technologies, Kumamoto, Japan) was used to
analyze the growth of Tca8113 cells. According to the
manufacturer's instructions, 100 µl RPMI 1640 medium, containing
1×103 cells/well, was added to 96-well plates (Corning,
Inc.). The plates were incubated for 24, 48, 72 or 96 h in a
humidified incubator. Subsequently, 10 µl CCK-8 solution was added
to the wells and five replicate wells were used for each time
point. Following incubation for 1 h, the absorbance was measured
using the EnSpire Multimode Plate Reader (PerkinElmer, Inc.,
Waltham, MA, USA) at a wavelength of 450 nm.
Cell apoptosis assay
Cell apoptosis was analyzed using the Annexin
V-phycoerythrin (PE)/7-amino-actinomycin D (7-AAD) Apoptosis
Detection Kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China)
according to the manufacturer's instructions. Briefly, the cells
were digested with trypsin and washed twice with phosphate-buffered
saline (PBS) and 5×105 cells were resuspended in 500 µl
binding buffer (Nanjing KeyGen Biotech Co., Ltd.). The suspension
was stained with 1 µl Annexin V-PE in the dark for 10 min, at room
temperature. Next, 5 µl 7-AAD was added to the suspension and left
for 10 min at room temperature, in the dark. Cell apoptosis was
analyzed using the BD FACSAria II cell sorter (BD Biosciences,
Franklin Lakes, NJ, USA).
Cell cycle assay
The cell cycle was analyzed using the propidium
iodide (PI) Detection kit (Nanjing KeyGen Biotech Co., Ltd.)
according to the manufacturer's instructions. The cells were
collected and washed once in 1X buffer A (Nanjing KeyGen Biotech
Co., Ltd.). The cells were resuspended at a concentration of
1×106 cells/ml in 1X buffer A. The suspension was mixed
with 70% ethanol at a ratio of 1:9 and was left for ≥12 h at −20°C.
The cells were then centrifuged at <500 × g at room temperature
and washed with 1X buffer A and resuspended in 500 µl 1X buffer A.
RNase A (0.25 mg/ml) was added and allowed to react at 37°C for 30
min. A total of 5µl PI was added in the dark and allowed to react
for 30 min at room temperature. The cell cycle data were analyzed
by flow cytometry (BD FACSAria II; BD Biosciences).
Cell migration and invasion assay
A 3422 Transwell chamber (Corning, Inc.) was used to
perform the cell invasion assay. Each Transwell chamber was coated
with Matrigel (BD Biosciences) 24 h prior to use. The cells were
harvested and resuspended at a concentration of 1×106
cells/ml in RPMI 1640 medium containing 1% bovine serum albumin.
Subsequently, 100 µl of the suspension was added to the upper
chamber, while 500 µl of the RPMI 1640 medium containing 20% FBS
was added to the lower chamber. After 48-h incubation, the cells on
the upper surface of the chamber were completely removed. The lower
surface was washed gently with PBS and fixed in 4% formaldehyde
polymerisatum for 20 min. The chamber was stained with DAPI
(Sigma-Aldrich, St. Louis, MO, USA) and quantified under a
microscope (Eclipse Ti; Nikon Corporation). The cell migration
assay was the same as the invasion assay, with the exception of the
Matrigel coating.
Results
Overexpression of GSN in transfected
Tca8113 cells
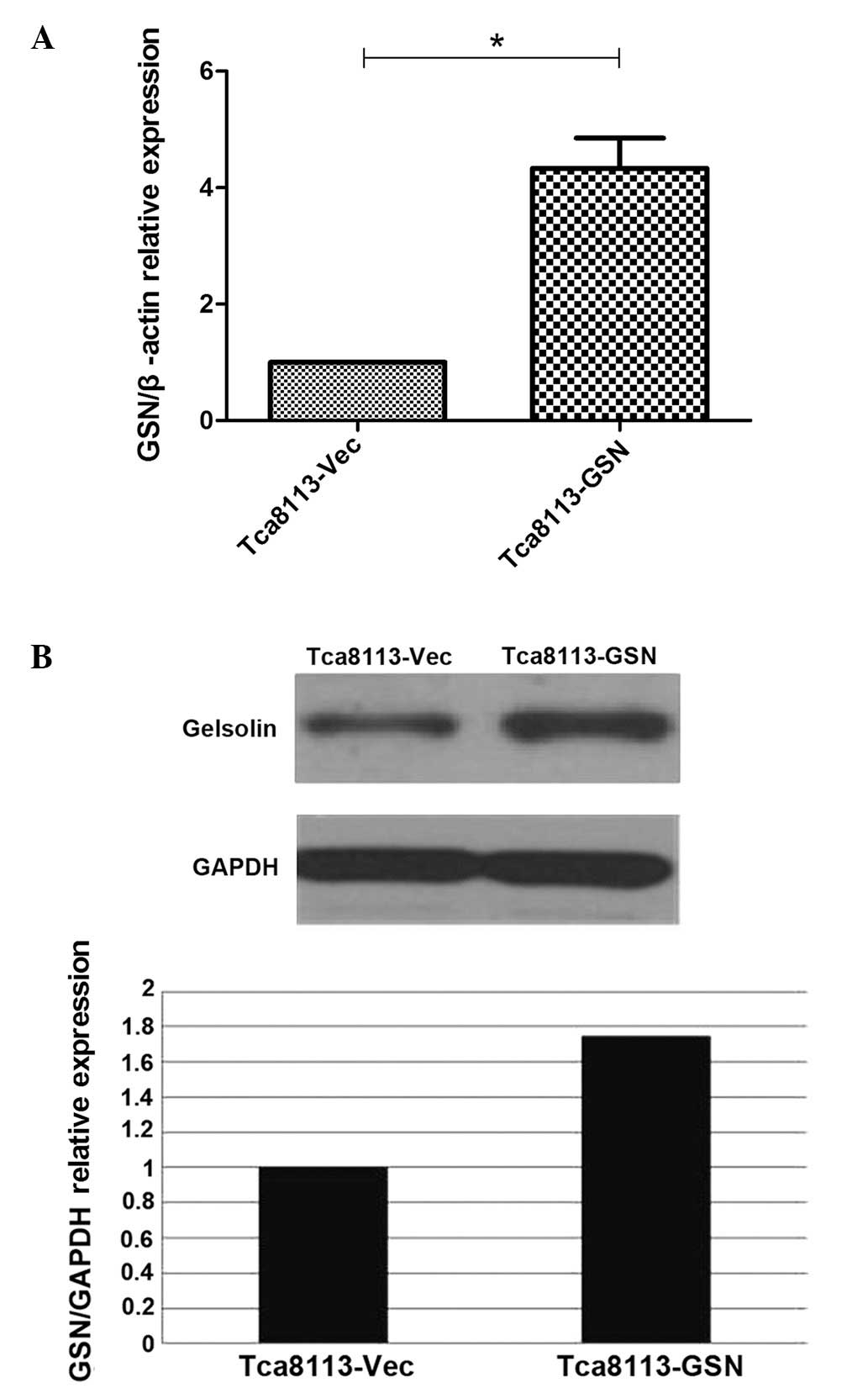
Subsequent to the stable transfectants being
established, RT-qPCR revealed that GSN mRNA expression in the
Tca8113-GSN cells was 4-fold higher compared with the Tca8113-Vec
cells (Fig. 1A). GSN protein
expression was increased by 1.75 times compared with the negative
control cells (Fig. 1B).
Effect of GSN overexpression on the
proliferation of Tca8113 cells
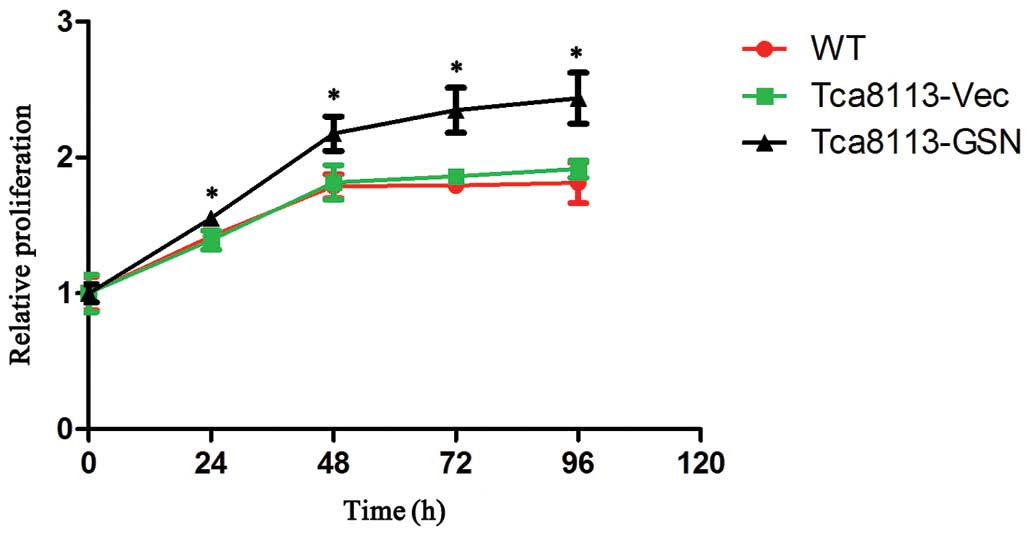
CCK-8 assays were performed to examine the
proliferation of wild type Tca8113 (Tca8113-WT), Tca8113-GSN and
Tca8113-Vec cells in vitro. As shown in Fig. 2, the growth of Tca8113-GSN cells was
significantly increased when compared with Tca8113-Vec or wild type
Tca8113 cells.
Effect of GSN on the apoptosis and
cell cycle of Tca8113 cells
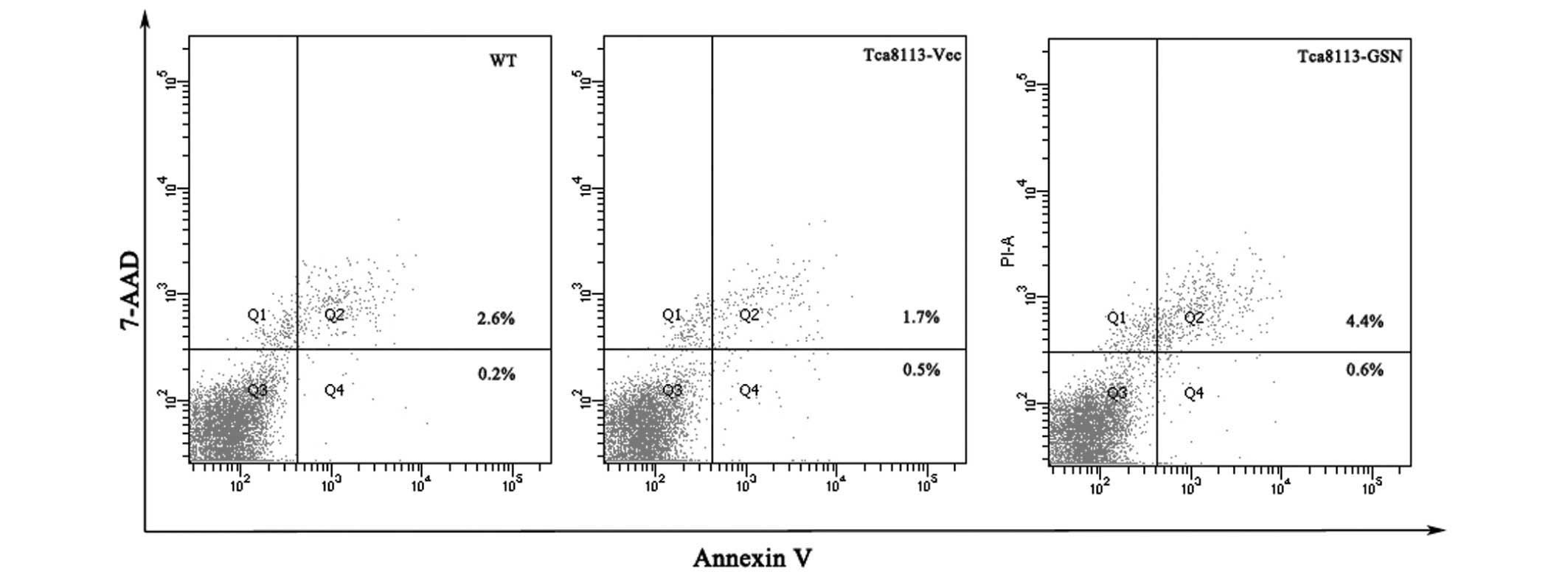
The cellular apoptosis of Tca8113-WT, Tca8113-GSN
and Tca8113-Vec cells was analyzed using an Annexin V-fluorescein
isothiocyanate assay. The results demonstrated that the apoptosis
rate of Tca8113-GSN cells was 5.0%, compared with 2.2% in
Tca8113-Vec cells and 2.8% in Tca8113-WT cells (Fig. 3). Thus, these results indicated that
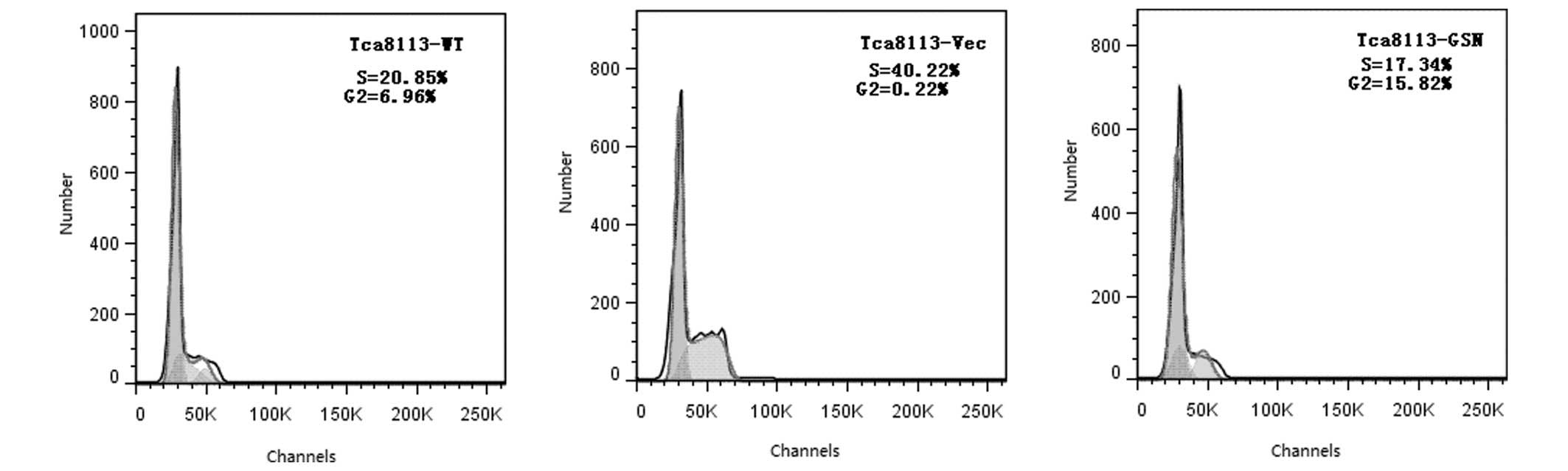
GSN may promote apoptosis in Tca8113 cells. However, cell cycle
analysis revealed that the number of cells in the S and
G2 phases in the Tca8113-WT and Tca8113-GSN groups was
lower compared with the Tca8113-Vec group (Fig. 4).
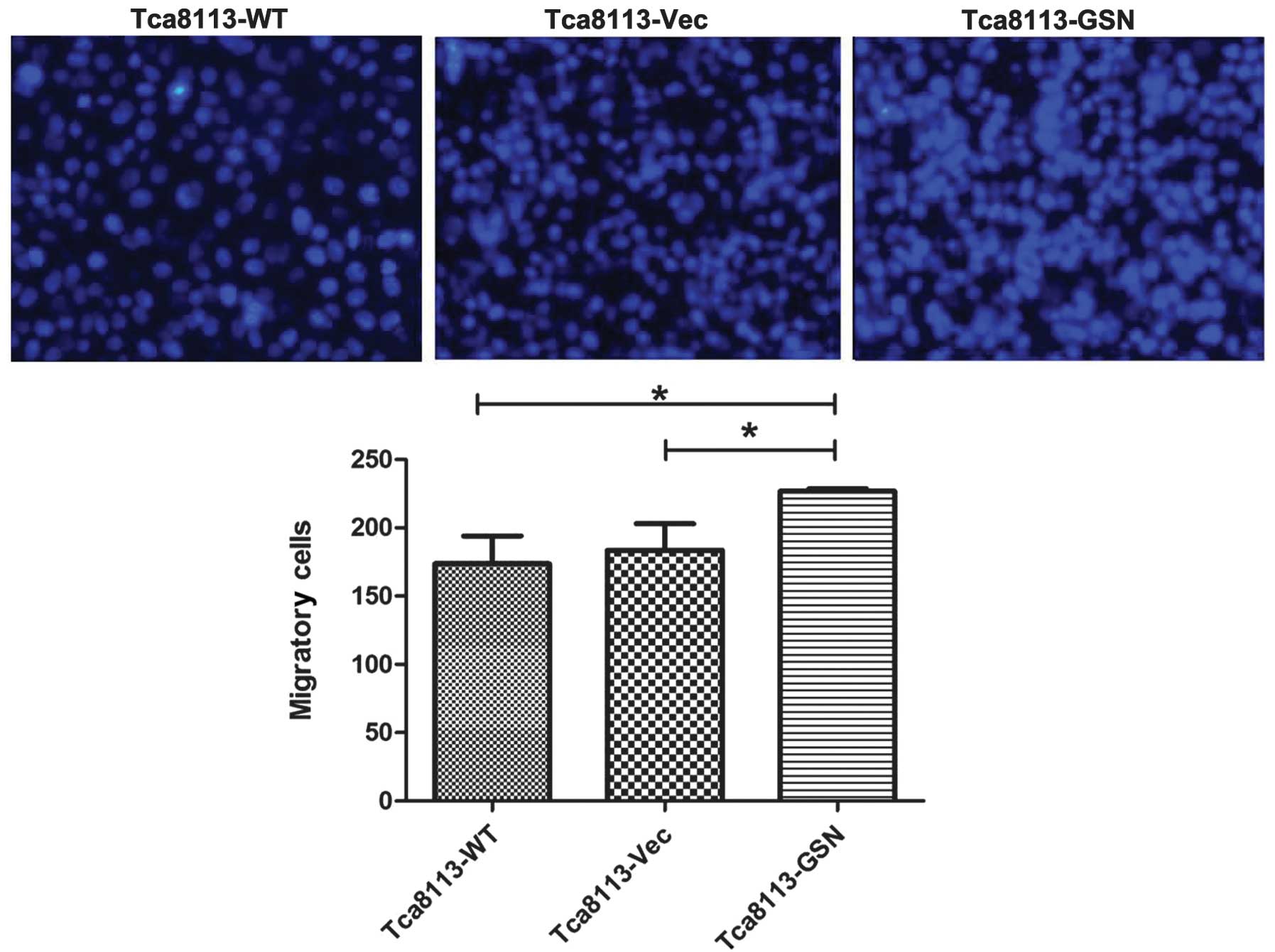
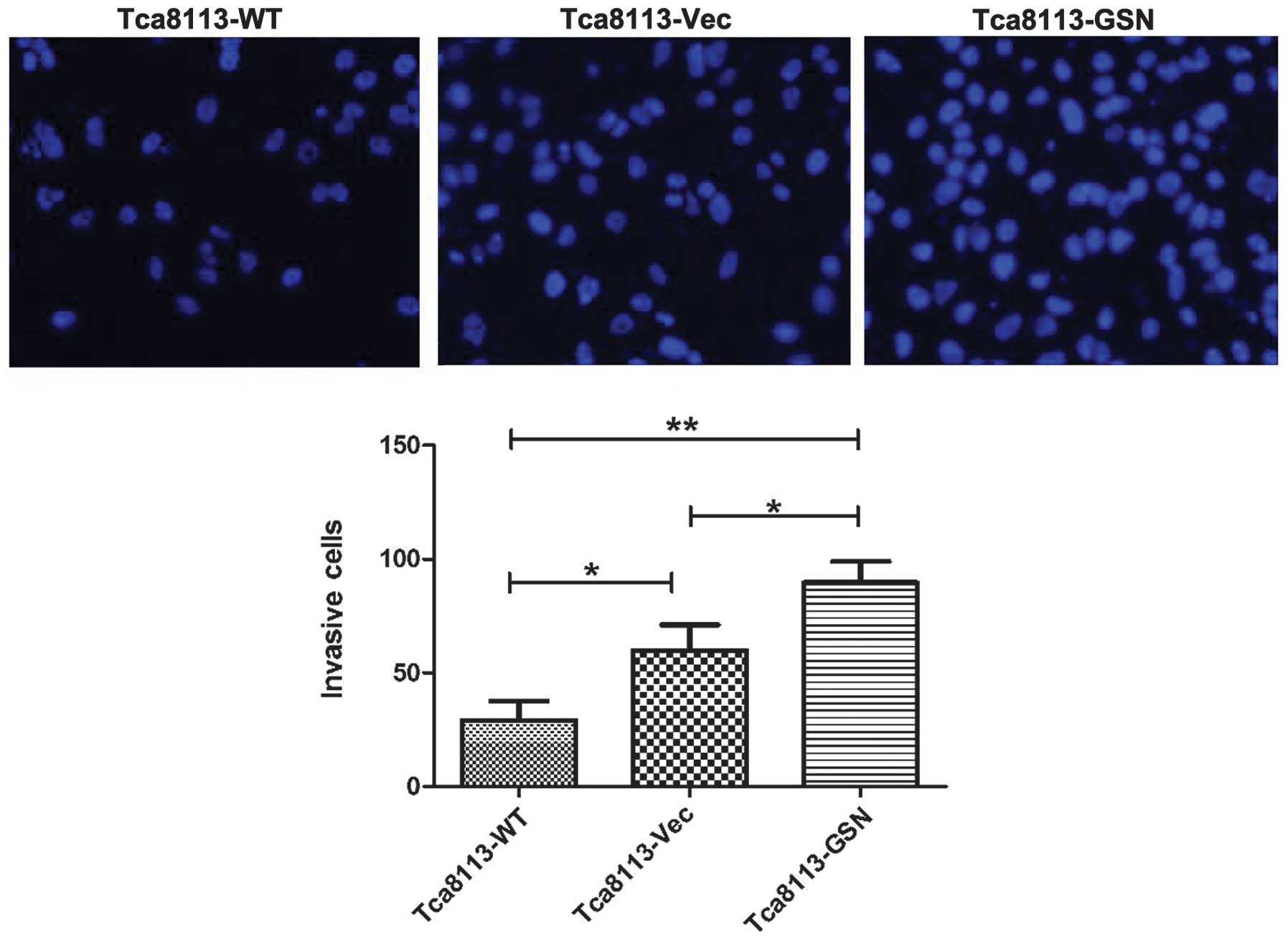
GSN overexpression correlates with the
promotion of migration and invasion in Tca8113 cells
Increased migration and invasion was closely
associated with carcinoma progression. To determine the role of GSN
in oral cancer cell migration and invasion, Transwell assays were
performed. The results revealed that Tca8113-GSN cells exhibited
significantly increased migratory and invasive abilities (Figs. 5 and 6).
Discussion
Oral cancer is the eighth most common cancer
globally with ∼270,000 novel cases and 130,000 mortalities
annually, and a five-year survival rate of ∼63% (10–14). Oral
cavity cancer is characterized by high aggression, early recurrence
and frequent lymph node metastasis (15).
The early diagnosis of oral cancer is extremely
important as, early stage oral cancer treated with surgery or
radiation is usually associated with a good prognosis. Advanced
cancers are routinely treated by surgery and post-operative
radiotherapy, but the prognosis is often poor (14.) However, numerous patients possess
advanced-stage disease at the time of diagnosis. The five-year
survival rates are 41 and 9% for patients with stage III and IV
disease, respectively, which are significantly lower compared with
patients possessing stage I or II disease, with a five-year
survival rate of 85 and 66%, respectively (16). It has been shown that early diagnosis
is critical for improving oral cancer treatment. A number of
studies have been devoted to identifying appropriate biomarkers for
the early diagnosis, which may increase understanding of the
pathogenesis and may aid in the identification of therapeutic
targets (12,15).
In eukaryotic species, it has been found that ABPs
are not only involved in normal cell metabolism, but also perform
an extremely important role in numerous pathological processes
(1–3,5,7,17). As an
extremely abundant eukaryotic ABP, GSN severs, caps and nucleates
actin filaments, and then dynamically regulate the cytoskeleton.
Therefore, GSN exerts numerous physiological effects, including
modulation of cell apoptosis, mediation of signal transduction and
regulation of transcriptional coactivation (7).
The importance of GSN in the initiation and
progression of malignancies remains unclear. To date, the altered
expression of GSN has been identified in specific cancers,
including human bladder, ovarian, lung, breast, gastric,
pancreatic, cervical and oral carcinomas (8,18–25). The majority of carcinomas exhibited
downregulation of GSN, but GSN overexpression was observed in
cervical carcinoma (8,18–25). In
contrast to other tumors, a biphasic expression of GSN in oral
precancerous lesions and cancers has been observed (8). The downregulation of GSN was observed
between oral normal mucosa and precancerous lesions, whereas GSN
upregulation in oral cancer predicated an increased tumor size and
invasive growth (8). The dual effects
indicate that GSN may serve as a tumor suppressor in oral cancer,
while serving as a promoter in oral cancer progression.
In the present study, GSN expression was increased
in oral squamous cell carcinoma cells, which was demonstrated by
GSN protein expression in Tca8113-GSN cells being double that of
Tca8113-Vec cells. In this study, the overexpression of GSN
promoted the proliferation and apoptosis of Tca8113 cells. However,
the mechanism of the promotion of apoptotic activity in Tca8113
cells was not investigated and thus requires additional
investigation. However, previous studies have indicated that GSN
functions as a growth promoter and apoptosis inhibitor (26). GSN overexpression may inhibit the
activation of caspases-3, -8 and -9 by preventing the release of
cytochrome c from mitochondria, and thus exert its function
in anti-apoptotic mechanisms. Additional studies have revealed that
GSN may exert anti-apoptotic effects by blocking actin-dependent
voltage-dependent anion-selective channels in
mitochondrial-dependent cell death, and repress the p53-mediated
apoptosis in HepG2 cells (7,27–34).
The results of the present study revealed that GSN
was involved in cell motility. Increased cell motility is important
in tumor progression, particularly in the multistep process of
invasion and metastasis (35). A
previous study found that the expression of GSN was closely
associated with oral carcinoma progression (8). The current study indicated that the
upregulation of GSN increased Tca8113 cell migration and invasion
in vitro, which caused enhanced motility. In addition, it
has been indicated that GSN may affect motility in human colon
cancer and melanoma cells (30,36–40). In
human colon cancer cells, the overexpression of GSN was accompanied
by the promotion of migration capacity. In addition, the decreased
expression of GSN decreased the migratory potential of melanoma
cells (30,36–40).
Therefore, GSN overexpression may result in positive effects that
promote invasion and metastasis in oral cancer progression.
In conclusion, the results of the present study
demonstrate that GSN significantly promotes oral squamous cell
carcinoma Tca8113 cell proliferation, migration and invasion in
human oral cancer. GSN may exert an important role in cell growth
and motility, which may provide novel insights for understanding
the potential mechanisms of oral cancer progression.
Acknowledgements
This study was supported by the Nanjing Medical
Science and Technique Development Foundation (grant no. QYK11129),
the Natural Science Foundation of Jiangsu Province (grant no.
BK2012523) and the National Key Disciplines Constructional Project
Funding.
References
|
1
|
Uribe R and Jay D: A review of actin
binding proteins: new perspectives. Mol Biol Rep. 36:121–125. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
dos Remedios CG, Chhabra D, Kekic M, et
al: Actin binding proteins: regulation of cytoskeletal
microfilaments. Physiol Rev. 83:433–473. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Winder SJ and Ayscough KR: Actin-binding
proteins. J Cell Sci. 118:651–654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dominguez R: Actin-binding proteins - a
unifying hypothesis. Trends Biochem Sci. 29:572–578. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Janmey PA and Chaponnier C: Medical
aspects of the actin cytoskeleton. Curr Opin Cell Biol. 7:111–117.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pollard TD and Borisy GG: Cellular
motility driven by assembly and disassembly of actin filaments.
Cell. 112:453–465. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li GH, Arora PD, Chen Y, McCulloch CA and
Liu P: Multifunctional roles of gelsolin in health and diseases.
Med Res Rev. 32:999–1025. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shieh DB, Chen IW, Wei TY, et al: Tissue
expression of gelsolin in oral carcinogenesis progression and its
clinicopathological implications. Oral Oncol. 42:599–606. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mullauer L, Fujita H, Ishizaki A and
Kuzumaki N: Tumor-suppressive function of mutated gelsolin in
ras-transformed cells. Oncogene. 8:2531–2536. 1993.PubMed/NCBI
|
|
10
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics. 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar
|
|
12
|
Nomura H, Uzawa K, Ishigami T, et al:
Clinical significance of gelsolin-like actin-capping protein
expression in oral carcinogenesis: an immunohistochemical study of
premalignant and malignant lesions of the oral cavity. BMC Cancer.
8:392008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Petersen PE: Oral cancer prevention and
control - the approach of the World Health Organization. Oral
Oncol. 45:454–460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tsantoulis PK, Kastrinakis NG, Tourvas AD,
Laskaris G and Gorgoulis VG: Advances in the biology of oral
cancer. Oral Oncol. 43:523–534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
da Silva SD, Ferlito A, Takes RP, et al:
Advances and applications of oral cancer basic research. Oral
Oncol. 47:783–791. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sciubba JJ: Oral cancer. The importance of
early diagnosis and treatment. Am J Clin Dermatol. 2:239–251. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ayscough KR: In vivo functions of
actin-binding proteins. Curr Opin Cell Biol. 10:102–111. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tanaka M, Mullauer L, Ogiso Y, et al:
Gelsolin: a candidate for suppressor of human bladder cancer.
Cancer Res. 55:3228–3232. 1995.PubMed/NCBI
|
|
19
|
Noske A, Denkert C, Schober H, et al: Loss
of Gelsolin expression in human ovarian carcinomas. Eur J Cancer.
41:461–469. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dosaka-Akita H, Hommura F, Fujita H, et
al: Frequent loss of gelsolin expression in non-small cell lung
cancers of heavy smokers. Cancer Res. 58:322–327. 1998.PubMed/NCBI
|
|
21
|
Winston JS, Asch HL, Zhang PJ, Edge SB,
Hyland A and Asch BB: Downregulation of gelsolin correlates with
the progression to breast carcinoma. Breast Cancer Res Treat.
65:11–21. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim JH, Choi YK, Kwon HJ, Yang HK, Choi JH
and Kim DY: Downregulation of gelsolin and retinoic acid receptor
beta expression in gastric cancer tissues through histone
deacetylase 1. J Gastroenterol Hepatol. 19:218–224. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ni XG, Zhou L, Wang GQ, et al: The
ubiquitin-proteasome pathway mediates gelsolin protein
downregulation in pancreatic cancer. Mol Med. 14:582–589. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shieh DB, Godleski J, Herndon JE II, et
al: Cell motility as a prognostic factor in Stage I nonsmall cell
lung carcinoma: the role of gelsolin expression. Cancer. 85:47–57.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liao CJ, Wu TI, Huang YH, et al:
Overexpression of gelsolin in human cervical carcinoma and its
clinicopathological significance. Gynecol Oncol. 120:135–144. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Silacci P, Mazzolai L, Gauci C,
Stergiopulos N, Yin HL and Hayoz D: Gelsolin superfamily proteins:
key regulators of cellular functions. Cell Mol Life Sci.
61:2614–2623. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
An JH, Kim JW, Jang SM, Kim CH, Kang EJ
and Choi KH: Gelsolin negatively regulates the activity of tumor
suppressor p53 through their physical interaction in
hepatocarcinoma HepG2 cells. Biochem Biophys Res Commun. 412:44–49.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Koya RC, Fujita H, Shimizu S, et al:
Gelsolin inhibits apoptosis by blocking mitochondrial membrane
potential loss and cytochrome c release. J Biol Chem.
275:15343–15349. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kusano H, Shimizu S, Koya RC, et al: Human
gelsolin prevents apoptosis by inhibiting apoptotic mitochondrial
changes via closing VDAC. Oncogene. 19:4807–4814. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kwiatkowski DJ: Functions of gelsolin:
motility, signaling, apoptosis, cancer. Curr Opin Cell Biol.
11:103–108. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sakurai N and Utsumi T: Posttranslational
N-myristoylation is required for the anti-apoptotic activity of
human tGelsolin, the C-terminal caspase cleavage product of human
gelsolin. J Biol Chem. 281:14288–14295. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Azuma T, Koths K, Flanagan L and
Kwiatkowski D: Gelsolin in complex with phosphatidylinositol
4,5-bisphosphate inhibits caspase-3 and-9 to retard apoptotic
progression. J Biol Chem. 275:3761–3766. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Galluzzi L and Kroemer G: Mitochondrial
apoptosis without VDAC. Nat Cell Biol. 9:487–489. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Baines CP, Kaiser RA, Sheiko T, Craigen WJ
and Molkentin JD: Voltage-dependent anion channels are dispensable
for mitochondrial-dependent cell death. Nat Cell Biol. 9:550–555.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ke H, Parron VI, Reece J, Zhang JY,
Akiyama SK and French JE: BCL2 inhibits cell adhesion, spreading,
and motility by enhancing actin polymerization. Cell Res.
20:458–469. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
De Corte V, Bruyneel E, Boucherie C,
Mareel M, Vandekerckhove J and Gettemans J: Gelsolin-induced
epithelial cell invasion is dependent on Ras-Rac signaling. EMBO J.
21:6781–6790. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Azuma T, Witke W, Stossel TP, Hartwig JH
and Kwiatkowski DJ: Gelsolin is a downstream effector of rac for
fibroblast motility. EMBO J. 17:1362–1370. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Radwanska A, Litwin M, Nowak D, et al:
Overexpression of lumican affects the migration of human colon
cancer cells through up-regulation of gelsolin and filamentous
actin reorganization. Exp Cell Res. 318:2312–2323. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Litwin M, Nowak D, Mazur AJ, Baczynska D,
Mannherz HG and Malicka-Blaszkiewicz M: Gelsolin affects the
migratory ability of human colon adenocarcinoma and melanoma cells.
Life Sci. 90:851–861. 2012. View Article : Google Scholar : PubMed/NCBI
|