Introduction
Nasopharyngeal carcinoma (NPC) is a type of
malignant tumor derived from nasal epithelial cells (1), and is one of the most common
malignancies in Southern China and Southeast Asia (2–4), with an
incidence rate of 20–30 per 100,000 individuals (2,5,6). NPCs frequently go unnoticed by patients
as pathogenic locations are difficult to detect, primary lesions
are small and symptoms are mild. The biological characteristics and
abundant peripheral lymphoid tissue involvement of NPC make this
malignancy more prone to metastasis and invasion compared with
other head and neck tumors. Partial recurrence and distant
metastasis of NPC are causes of treatment failure (7). Consequently, the majority of patients
succumb to the effects of tumor metastasis rather than to the
primary lesion. Research on the invasion and migration of NPC is
therefore essential.
S100A8 protein (also known as calgranulin A or MRP8
protein) and S100A9 protein (also known as calgranulin B or MRP14
protein) are members of the S100 calcium-binding protein family and
usually form a calcium-dependent S100A8/A9 heterodimer complex
(8–10). S100A8/A9 is predominantly located in
the cytoplasm, with some localized to the nucleus. The complex is
normally expressed in circulating neutrophils and monocytes, and
marrow cells in early differentiation, with no expression in tissue
macrophages. S100A8/A9 is primarily expressed in
well-differentiated tissue cells, including epithelial and skin
cells (11). A number of studies have
suggested that S100A8/A9 expression is upregulated in a variety of
primary and invasive tumors (12,13). Kim
et al (14) reported increased
expression of S100A8/A9 in colorectal carcinoma tissues and serum
in patients with early colorectal cancer. Hermani et al
(15) demonstrated that the
expression level of S100A8/A9 in prostatic intraepithelial
neoplasia and prostatic adenocarcinoma is higher compared with that
of benign prostate hyperplasia tissues. In addition, the serum
level of S100A9 in patients with prostate cancer is higher than in
patients with benign prostatic hyperplasia or in healthy
subjects.
Saha et al (16) reported that treating B6F10 melanoma
cells with S100A8/A9 at 0.2–1 µg/ml induces expression of matrix
metalloproteinases (MMPs), which are involved in the invasion and
migration of tumor cells, promoting cell migration. In a study on
endogenous S100A8/A9, Yong and Moon (17) reported that S100A8/A9 secreted by
tumor cells causes cell invasion and migration.
In another previous study, the plasma concentration
of S100A8 and S100A9 in NPC patients was higher than in healthy
subjects, as indicated by isobaric tags for relative and absolute
quantitation combined with two-dimensional liquid
chromatography/tandem mass spectrometry (18). The present study aimed to clarify the
effects on CNE1 NPC cell migration following knockdown of S100A8
and S100A9 with small interfering RNA (siRNA).
Materials and methods
Cell lines and cell culture
The CNE1 NPC cell line was obtained from the Cell
Collection of the Central South University Xiangya Central
Laboratory (Changsha, Hunan, China). The cells were maintained in
RPMI-1640 culture medium (Hyclone, Logan, UT, USA) supplemented
with 10% fetal bovine serum (FBS; Hyclone), 100 U/ml penicillin and
100 µg/ml streptomycin in a humidified atmosphere of 5%
CO2 at 37°C. The cells were subcultured every three
days, and cells in the logarithmic growth phase were used for the
experiments.
Design and synthesis of S100A8 and
S100A9 siRNAs
siRNA sequences for S100A8 and S100A9 gene silencing
were designed according to mRNA sequences (S100A8 NCBI Reference
Sequence, NM_002964.4; S100A9 NCBI Reference Sequence,
NM_002965.3). The siRNA sequences with S100A8 and S100A9 target
sites, in addition to the negative control siRNA sequences
(Shanghai GenePharma Co., Ltd., Shanghai, China) are listed in
(Table I).
 | Table I.Small interfering RNA sequences used
for cell transfection. |
Table I.
Small interfering RNA sequences used
for cell transfection.
| Group | Sense | Antisense |
|---|
| S100A8-Homo-286 |
5′-AGACCGAGUGUCCUCAGUA-3′ |
5′-UACUGAGGACACUCGGUCU-3′ |
| S100A8-Homo-324 |
5′-GACGUCUGGUUCAAAGAGU-3′ |
5′-ACUCUUUGAACCAGACGUC-3′ |
| S100A8-Homo-374 |
5′-CCAGGAGUUCCUCAUUCUG-3′ |
5′-CAGAAUGAGGAACUCCUGG-3′ |
| S100A9-Homo-61 |
5′-GCAGCUGGAACGCAACAUA-3′ |
5′-UAUGUUGCGUUCCAGCUGC-3′ |
| S100A9-Homo-100 |
5′-CCACCAAUACUCUGUGAAG-3′ |
5′-CUUCACAGAGUAUUGGUGG-3′ |
| S100A9-Homo-267 |
5′-GCUUCGAGGAGUUCAUCAU-3′ |
5′-AUGAUGAACUCCUCGAAGC-3″ |
| Negative control |
5′-GCGACGAUCUGCCUAAGAU-3′ |
5′-AUCUUAGGCAGAUCGUCGC-3″ |
Cell transfection
To determine the transfection efficiency prior to
interference experiments, a negative control siRNA with green
fluorescence was transfected into the CNE1 cells. The conditions
for optimal transfection efficiency were determined by calculating
the percentage of fluorescent cells using a fluorescence microscope
and optimizing the ratio of Lipofectamine 2000 transfection reagent
(Invitrogen Life Technologies, Carlsbad, CA, USA) to siRNA. The
cells were divided into five groups: Experimental S100A8 group
transfected with siRNA; experimental S100A9 group transfected with
siRNA; blank control group containing untreated cells; negative
control group transfected with non-targeting control siRNA; and
mock-treatment group treated with transfection reagent only.
Results were obtained by the comparative analysis of experimental
groups to control groups. The CNE1 cells in each group were seeded
into six-well plates at a density of 3×105 cells/well in
RPMI-1640 medium containing 10% FBS without antibiotics for 24 h.
The cells were transfected with siRNA in serum-free RPMI-1640
medium, following the manufacturer's instructions for the use of
Lipofectamine 2000. Following incubation at 37°C in an atmosphere
of 5% CO2 for 6 h, the medium was replaced with
RPMI-1640 medium supplemented with 10% serum, and the cells were
incubated for 24 h prior to use.
Detection of S100A8 and S100A9
expression by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR)
Total RNA was isolated from the cells using total
RNA extraction kits (Corning Life Sciences, Axygen® Inc., Union
City, CA, USA), and RNA integrity and concentration were determined
by spectrophotometry (OD260/OD280 value, 1.9;
concentration, 980 ng/µl). cDNA was obtained using reverse
transcription kits (Takara Biotechnology Co., Ltd., Dalian, China)
following the removal of genomic DNA. qPCR was performed using SYBR
Green master mix (Roche Applied Science, Penzberg, Upper Bavaria,
Germany) and Mastercycler® ep realplex (Eppendorf, Hauppauge, NY,
USA). Primer sequences for S100A8, S100A9 and β-actin are listed in
(Table II). The relative expression
level was normalized to β-actin. Experiments were performed in
triplicate with a no-template control. PCR conditions were as
follows: 95°C for 10 min, followed by 40 cycles of 15 sec at 95°C
and 60 sec at 60°C. S100A8-Homo-374 siRNA and S100A9-Homo-267 siRNA
were used in the experiments, as they were identified to be the
most effective for interference based on multiple comparisons among
the siRNAs for the three groups.
| Table II.Primer sequences used in reverse
transcription-quantitative polymerase chain reaction
experiments. |
Table II.
Primer sequences used in reverse
transcription-quantitative polymerase chain reaction
experiments.
| Gene | Sense | Antisense |
|---|
| S100A8 |
5′-GCTAGAGACCGAGTGTCCTCAG-3′ |
5′-GCCCATCTTTATCACCAGAATG-3′ |
| S100A9 |
5′-TGGAGGACCTGGACACAAATG-3′ |
5′-TCGTCACCCTCGTGCATCTT-3 |
| β-actin |
5′-CAGGCACCAGGGCGTGAT-3′ |
5′-TAGCAACGTACATGGCTGGG-3′ |
| Matrix
metalloproteinase-7 |
5′-GGAACAGGCTCAGGACTATCTC-3′ |
5′-CAACATCTGGCACTCCACA-3′ |
Cell migration assays
Cell migration was determined using scratch
wound-healing assays. Five equally spaced lines were made on the
backs of six-well plates, and the cells were seeded at a density of
3×105 cells/well. Cells were transfected, and after 24
h, a sterile 200-µl pipette tip was used to scratch the cells
perpendicular to the lines to form a wound. The cells were washed
three times with phosphate-buffered saline to remove dead cells and
cultured with serum-free medium. Width values from at least three
scratches were then measured at more than three positions per
scratch. Images were captured at 0, 24, 48 and 72 h after the
scratches were made. The width of scratches was measured at each
time-point to compare migration ratios among the groups.
Detection of MMP7 expression by
RT-qPCR
The primers used to detect MMP7 are listed in
(Table II). The detection method
used was the same as that used for the detection of S100A8 and
S100A9 expression.
Statistical analysis
Statistical analyses were conducted using SPSS
version 16.0 (SPSS, Inc., Chicago, IL, USA). Data are presented as
the mean ± standard deviation, and significance was determined by
one-way analysis of variance (ANOVA); multiple comparisons between
two groups were determined by a least significant difference
analysis in the ANOVA. P<0.05 was considered to indicate a
statistically significant difference.
Results
Detection of fluorescence in
transfected cells
The cells were observed under a fluorescence
microscope. The CNE1 cells transfected with the negative control
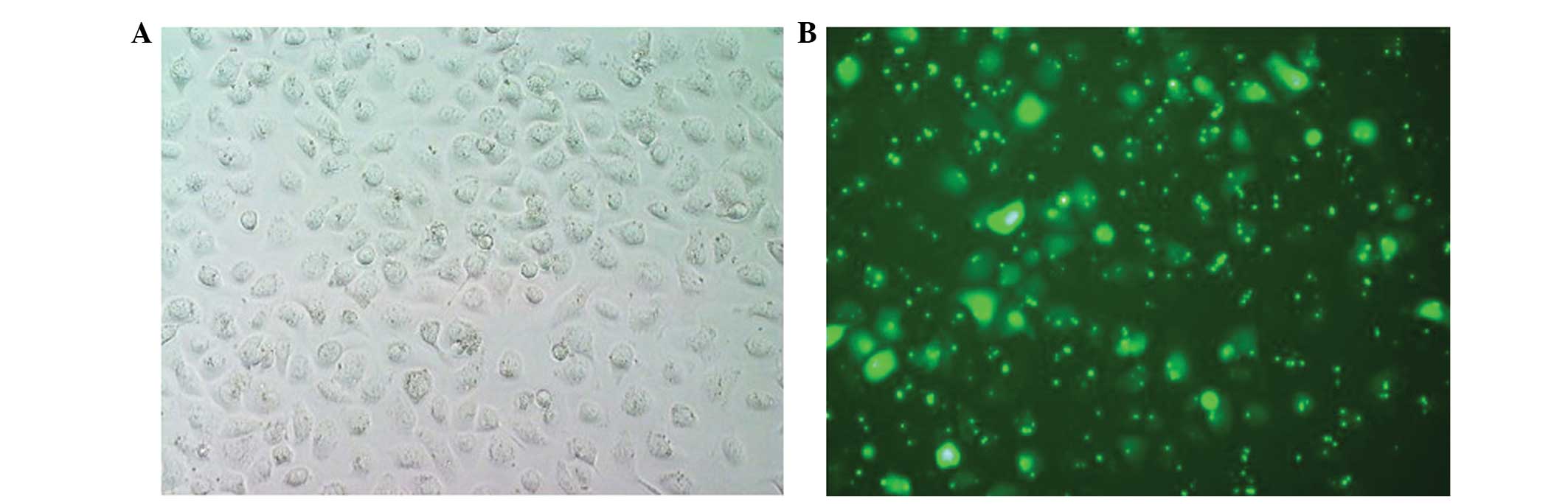
siRNA exhibited a green fluorescence (Fig. 1), whilst untransfected cells did not.
Transfection efficiencies were high using transfection agent to
siRNA ratios of 1:2 and 1:3; a ratio of 1:2 was selected for use in
subsequent transfections. Five randomly selected fields of ×20
magnification revealed ∼80% of cells with fluorescence, indicating
the successful transfection of the siRNA.
Interference effects of S100A8 and
S100A9 siRNAs
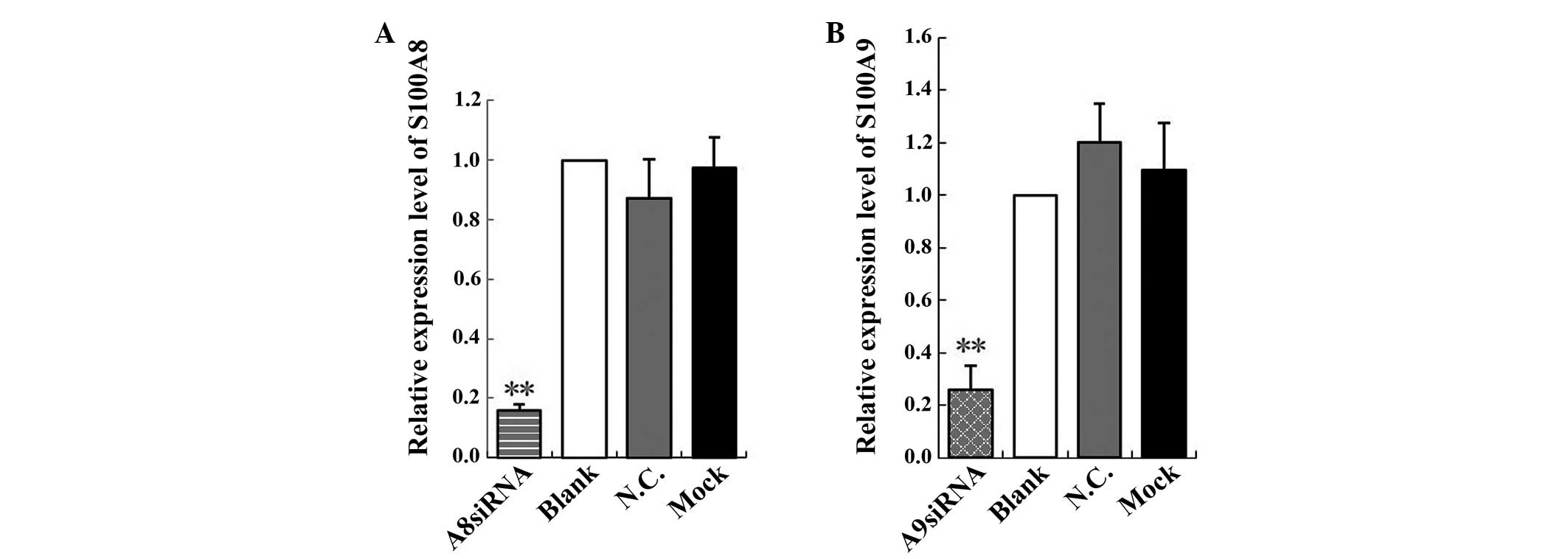
The relative expression of S100A8 was 0.16, which
was significantly lower compared with that of the blank control,
negative control and mock-treatment groups (F=69.95, P<0.01). No
significant difference was observed among the blank control,
negative control and mock-treatment groups (P>0.05; Fig. 2A). The relative expression of S100A9
was 0.26, and was significantly lower compared with that of the
blank control, negative control and mock-treatment groups (F=34.91,
P<0.01). No significant difference was observed among the blank
control, negative control and mock-treatment groups (P>0.05;
Fig. 2B). These results indicated
that the S100A8 and S100A9 siRNAs effectively inhibited S100A8 and
S100A9 expression.
Effects of S100A8 and S100A9 siRNAs on
cell migration
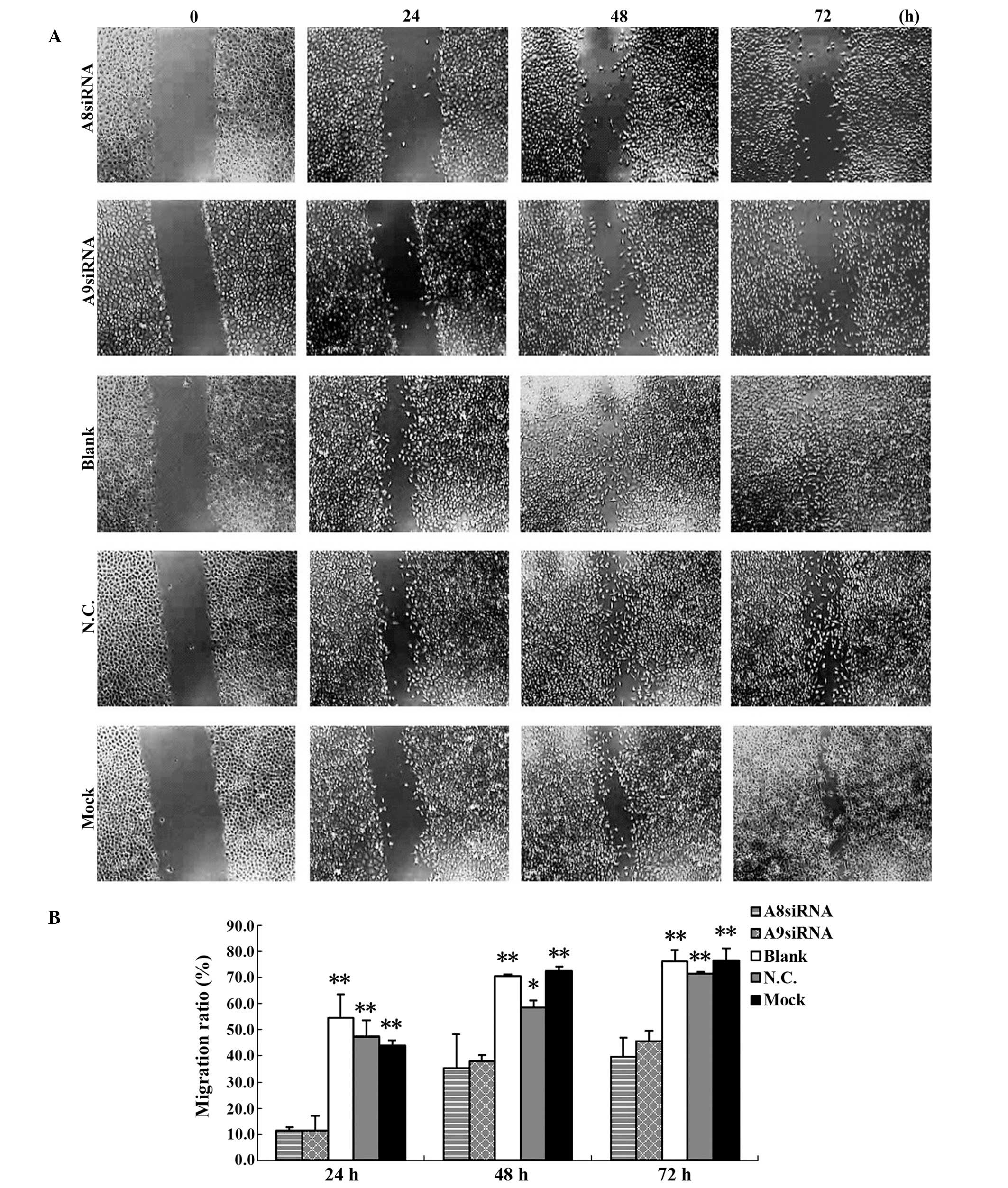
No evident restoration was observed in the
experimental groups transfected with S100A8 and S100A9 siRNAs at 24
h after scratching. By contrast, scratches were narrowed in the
blank control, negative control and mock-treatment groups. At 48 h
and 72 h, after the effects of the siRNAs had diminished, cell
migration in the S100A8 and S100A9 siRNA-transfected experimental
groups were restored to a certain degree, while scratched areas in
the blank control, negative control and mock-treatment groups had
filled with cells. At each time-point after scratching (24, 48 and
72 h), the migration ratio of the S100A8 and S100A9
siRNA-transfected experimental groups was significantly lower
compared with that of the blank control, negative control and
mock-treatment groups (Tables III
and IV). No significant differences
were observed among the blank control, negative control and
mock-treatment groups (P>0.05; Fig.
3; Tables III and IV). This result indicated that the
expression of S100A8 and S100A9 was downregulated by the siRNAs,
and that the migration of the CNE1 NPC cells had decreased.
| Table III.Migration ratio of each group in
scratch wound healing assays. |
Table III.
Migration ratio of each group in
scratch wound healing assays.
| Migration ratio,
% |
|---|
|
|---|
| Group | 24 h | 48 h | 72 h |
|---|
| S100A8 siRNA | 11.41±1.30 |
35.27±13.03 | 39.57±7.29 |
| S100A9 siRNA | 11.53±5.50 | 37.99±2.28 | 45.71±4.04 |
| Blank | 54.61±8.87 | 70.64±0.66 | 76.06±4.49 |
| Negative control | 47.36±6.32 | 58.59±2.69 | 71.45±0.79 |
| Mock | 43.83±2.22 | 72.71±1.58 | 76.67±4.55 |
| F-value | 27.56 | 16.88 | 28.64 |
| P-value | 0.001 | 0.004 | 0.001 |
| Table IV.P-values for the difference in
migration ratios between each treatment group at 24, 48 and 72 h
after scratching, determined by one-way analysis of variance. |
Table IV.
P-values for the difference in
migration ratios between each treatment group at 24, 48 and 72 h
after scratching, determined by one-way analysis of variance.
| P-value |
|---|
|
|---|
| Time | A8siRNA | A9siRNA | Blank | N.C. | Mock |
|---|
| 24 h |
|
|
|
|
|
|
A8siRNA | - | 0.984 | 0.001 | 0.001 | 0.002 |
|
A9siRNA | 0.984 | - | 0.001 | 0.001 | 0.002 |
|
Blank | 0.001 | 0.001 | - | 0.250 | 0.111 |
|
N.C. | 0.001 | 0.001 | 0.250 | - | 0.555 |
|
Mock | 0.002 | 0.002 | 0.111 | 0.555 | - |
| 48 h |
|
|
|
|
|
|
A8siRNA | - | 0.674 | 0.002 | 0.012 | 0.002 |
|
A9siRNA | 0.674 | - | 0.003 | 0.020 | 0.002 |
|
Blank | 0.002 | 0.003 | - | 0.104 | 0.748 |
|
N.C. | 0.012 | 0.020 | 0.104 | - | 0.068 |
|
Mock | 0.002 | 0.002 | 0.748 | 0.068 | - |
| 72 h |
|
|
|
|
|
|
A8siRNA | - | 0.249 | 0.001 | 0.001 | 0.001 |
|
A9siRNA | 0.249 | - | 0.001 | 0.003 | 0.001 |
|
Blank | 0.001 | 0.001 | - | 0.373 | 0.902 |
|
N.C. | 0.001 | 0.003 | 0.373 | - | 0.318 |
|
Mock | 0.001 | 0.001 | 0.902 | 0.318 | - |
Impact of S100A8 and S100A9 siRNAs on
MMP7 expression
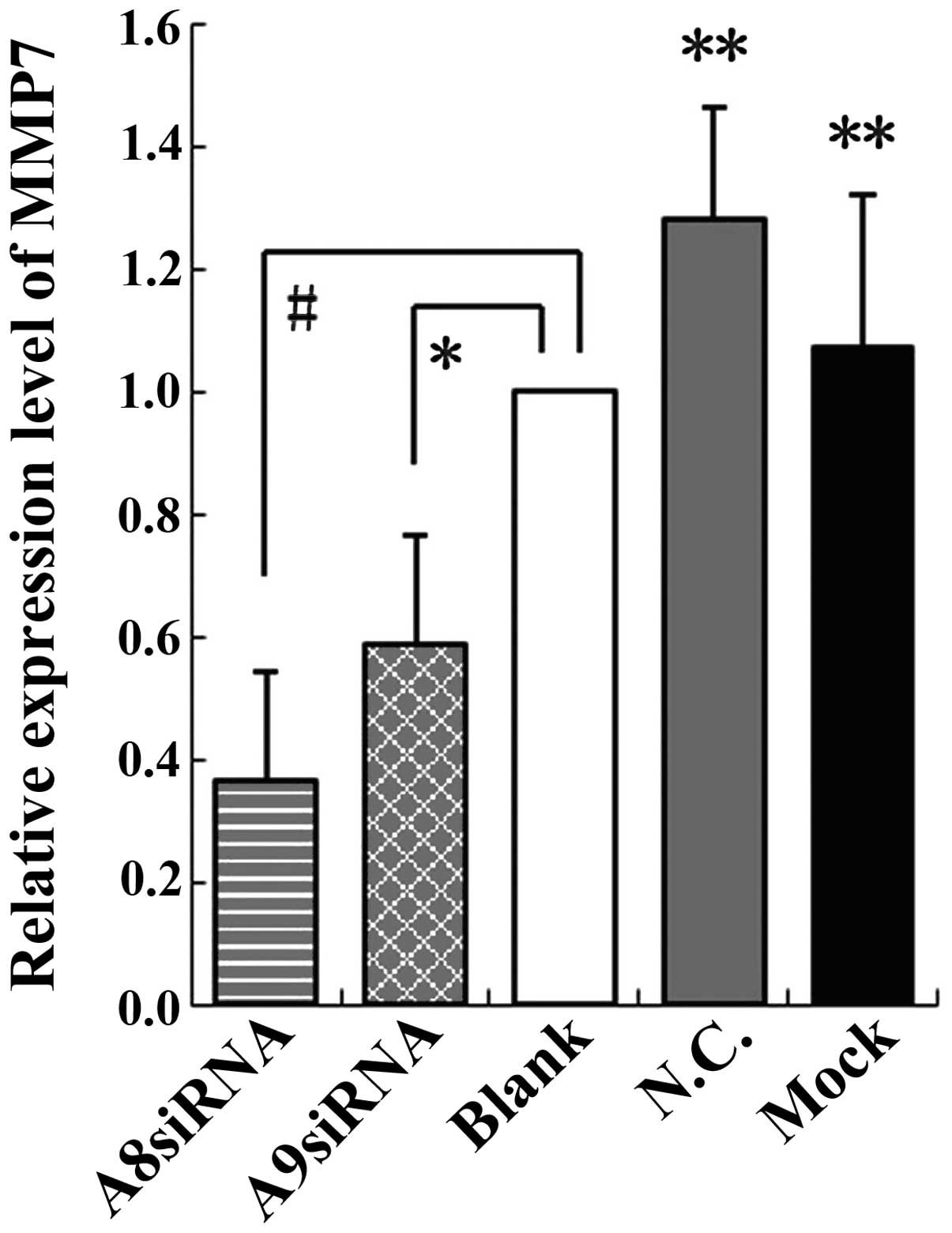
The relative expression of MMP7 was 0.3643 for the
S100A8 siRNA group and 0.5864 for the S100A9 siRNA group when
compared simultaneously with that of the blank control, negative
control and mock-treatment groups (F=13.193, P<0.01); no
significant differences were observed among the controls
(P>0.05; Fig. 4). These results
indicated that inhibition of MMP7 by S100A8 siRNA was more
effective than inhibition by S100A9 siRNA. Following the knockdown
of S100A8 and S100A9, the expression of MMP7 was downregulated to
different extents compared with the control groups.
Discussion
S100A8/A9 has a strong chemotactic effect on
leukocytes surrounding inflammatory lesions, which produce
inflammatory cytokines of neutrophils in inflammatory disease.
Hermani et al (19) reported
that S100A8/A9 at 10 µg/ml promotes migration of PNT1A prostate
cells. Hiratsuka et al (20)
demonstrated that S100A8/A9 at extremely low concentrations (S100A8
at 100 pg/ml and S100A9 at 1 ng/ml) promotes the migration of Lewis
lung carcinoma and B16 melanoma cells. Mounting evidence indicates
that MMP family members are involved in tumor invasion and
metastasis (17). MMPs are
proteolytic enzymes capable of degrading extracellular matrix
proteins (21), whose activity is
implicated in a number of key normal and pathological processes,
including tumor growth, progression, and metastasis and
dysregulated angiogenesis (22).
Enzymatic degradation of the extracellular matrix is a crucial step
in cancer invasion and metastasis. Another previous study supports
the hypothesis that S100A8 is more closely associated with MMP9
expression mediated by the extracellular-signal-regulated kinase
pathway, whilst S100A9 has a major role in MMP2 upregulation that
is dependent on p38 mitogen-activated protein kinase (MAPK)
signaling (23). A further study
reported that S100A8 and S100A9 contribute to colorectal carcinoma
cell survival and migration via the Wnt/β-catenin pathway. S100A8
and S100A9 increase total β-catenin levels and promote
transcription of its target genes (c-myc and MMP7), resulting in
the upregulation of the Wnt/β-catenin pathway (24).
The present study confirmed that S100A8 and S100A9
is vital for the migration of CNE1 NPC cells, and indicated that
MMP7 may be involved. RT-qPCR suggested different inhibitory
effects for the three pairs of siRNA sequences for S100A8 and
S100A9. The results revealed that S100A8-Homo-374 and
S100A9-Homo-267 were the most effective, with inhibition ratios of
84.10 and 74.15%, respectively; these siRNAs were used for
subsequent experiments. A previous study indicated that S100A8 and
S100A9 were knocked down in SNU484 gastric cancer cells; immunoblot
analysis suggested that S100A8 protein levels decreased by 47% and
S100A9 decreased by 85% (17).
Although the S100A8 and S100A9 siRNA sequences used in the previous
study differ from those used in the current study, the two siRNA
sequences inhibited S100A8 and S100A9 expression effectively.
Scratch wound healing assays performed after
different time periods revealed that, according to the migration
ratio ((Table III)), the migration
of the CNE1 cells was inhibited by S100A8 siRNA by 88.59% at 24 h,
64.73% at 48 h and 60.43% at 72 h. Marked inhibition of migration
was also observed when S100A9 expression was reduced by siRNA
(88.47% at 24 h, 62.01% at 48 h and 54.29% at 72 h). Moon et
al (23) performed Transwell
migration assays to demonstrate that H-Ras-mediated human breast
epithelial cell migration was inhibited by 57% by the knockdown of
S100A8, and by 80% by the knockdown of S100A9. Therefore, the
scratch wound healing assays and Transwell migration assays
illustrated that silencing S100A8 and S100A9 with siRNA inhibits
CNE1 cell migration.
In the present study, when MMP7 expression was
detected following the application of the most effective siRNAs for
S100A8 and S100A9, MMP7 expression was inhibited by 63.57% in
S100A8-downregulated cells and 41.36% in S100A9-downregulated
cells, indicating that the treatment with siRNA for S100A8 exerted
a greater inhibitory effect on MMP7 expression compared with that
of S100A9. This suggests that S100A8, and to a lesser extent
S100A9, may be required for MMP7 expression in CNE1 cells. However,
the difference in these results may also be due to greater
inhibitory effects of S100A8 siRNA on S100A8 expression, compared
with that of S100A9 siRNA on S100A9 expression. Further
investigation is required to clarify the differential roles of
S100A8 and S100A9 on MMP7 expression in CNE1 cells. Yong and Moon
(17) demonstrated that expression
level of MMP2 was decreased by 83% with S100A9 siRNA and by 52%
with S100A8 siRNA, indicating that S100A8/A9 may increase MMP2 and
MMP7, although this is inconsistent with the results reported by
Kwon et al (21).
In conclusion, the present study demonstrated that
the expression of S100A8 and S100A9 was effectively suppressed by
siRNA against these two genes, and that the migration of cells was
inhibited. In addition, MMP7 expression was somewhat reduced,
indicating that endogenous S100A8 and S100A9 promoted the migration
of CNE1 NPC cells; this is consistent with the conclusion that
S100A8/A9 promotes colon tumor cell metastasis, as reported by
Ichikawa et al (25). S100A8
and S100A9 may promote cell migration and invasion through p38
MAPK-dependent nuclear factor-κB activation (21). Further investigation is required to
determine the molecular mechanisms underlying the promotion of CNE1
NPC cell migration by endogenous S100A8 and S100A9.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation (no. 81260405), the Natural
Science Foundation of Guangxi (no. 2011GXNSFA018233), and the
Guangxi Science and Technology Agency project (no.
GK2013-A-02-02).
Abbreviations:
|
siRNA
|
small interfering RNA
|
|
RT-qPCR
|
real-time quantitative polymerase
chain reaction
|
|
NPC
|
nasopharyngeal carcinoma
|
|
MMPs
|
matrix metalloproteinases
|
References
|
1
|
Sham JS, Wei WI, Zong YS, Choy D, Guo YQ,
Luo Y, Lin ZX and Ng MH: Detection of subclinical nasopharyngeal
carcinoma by fibreoptic endoscopy and multiple biopsy. Lancet.
335:371–374. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chang ET and Adami HO: The enigmatic
epidemiology of nasopharyngeal carcinoma. Cancer Epidemiol
Biomarkers Prev. 15:1765–1777. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shin HR, Curado MP, Ferlay J, Heanue M,
Edwards B and Storm H: Comparability and Quality of DataCancer
Incidence in Five Continents. Parkin DM and Muir CS: IARC Sci Publ;
pp. 45–173. 1992
|
|
4
|
Nielsen NH, Mikkelsen F and Hansen JP:
Nasopharyngeal cancer in Greenland. The incidence in an Arctic
Eskimo population. Acta Pathol Microbiol Scand A. 85:850–858.
1977.PubMed/NCBI
|
|
5
|
Yu MC and Yuan JM: Epidemiology of
nasopharyngeal carcinoma. Semin Cancer Biol. 12:421–429. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gu AD, Mo HY, Bei JX, Xie YB, Chen LZ,
Feng QS, Kang T and Zeng YX: Evaluation of antibodies against
different Epstein-Barr virus nuclear antigen 1 peptides in
diagnosis of nasopharyngeal carcinoma. Clin Vaccine Immunol.
16:592–593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chiesa F and De Paoli F: Distant
metastases from nasopharyngeal cancer. ORL J Otorhinolaryngol Relat
Spec. 63:214–216. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Korndörfer IP, Brueckner F and Skerra A:
The crystal structure of the human (S100A8/S100A9)2 heterotetramer,
calprotectin, illustrates how conformational changes of interacting
alpha-helices can determine specific association of two EF-hand
proteins. J Mol Biol. 370:887–898. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ott HW, Lindner H, Sarg B, Mueller-Holzner
E, Abendstein B, Bergant A, Fessler S, Schwaerzler P, Zeimet A,
Marth C, et al: Calgranulins in cystic fluid and serum from
patients with ovarian carcinomas. Cancer Res. 63:7507–7514.
2003.PubMed/NCBI
|
|
10
|
Donato R: S100: A multigenic family of
calcium-modulated proteins of the EF-hand type with intracellular
and extracellular functional roles. Int J Biochem Cell Biol.
33:637–668. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kong JP, Ding F, Zhou CN, Wang XQ, Miao
XP, Wu M and Liu ZH: Loss of myeloid-related proteins 8 and
myeloid-related proteins 14 expression in human esophageal squamous
cell carcinoma correlates with poor differentiation. World J
Gastroenterol. 10:1093–1097. 2004.PubMed/NCBI
|
|
12
|
Gebhardt C, Németh J, Angel P and Hess J:
S100A8 and S100A9 in inflammation and cancer. Biochem Pharmacol.
72:1622–1631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salama I, Malone PS, Mihaimeed F and Jones
JL: A review of the S100 proteins in cancer. Eur J Surg Oncol.
34:357–364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim HJ, Kang HJ, Lee H, Lee ST, Yu MH, Kim
H and Lee C: Identification of S100A8 and S100A9 as serological
markers for colorectal cancer. J Proteome Res. 8:1368–1379. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hermani A, Hess J, De Servi B, et al:
Calcium-binding proteins S100A8 and S100A9 as novel diagnostic
markers in human prostate cancer. Clin Cancer Res. 11:5146–5152.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Saha A, Lee YC, Zhang Z, Chandra G, Su SB
and Mukherjee AB: Lack of an endogenous anti-inflammatory protein
in mice enhances colonization of B16F10 melanoma cells in the
lungs. J Biol Chem. 285:10822–10831. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yong HY and Moon A: Roles of
calcium-binding proteins, S100A8 and S100A9, in invasive phenotype
of human gastric cancer cells. Arch Pharm Res. 30:75–81. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Han RR, Huang YJ, Chen L, Xiao XL, Yi X,
Cai HW and Wu YH: Determination of S100A8 and S100A9 protein in
plasma of nasopharyngeal carcinoma patients and its clinical
significance. Chin J Clin Lab Sci. 32:252–254. 2014.[(In
Chinese)].
|
|
19
|
Hermani A, De Servi B, Medunjanin S,
Tessier PA and Mayer D: S100A8 and S100A9 activate MAP kinase and
NF-kappaB signaling pathways and trigger translocation of RAGE in
human prostate cancer cells. Exp Cell Res. 312:184–197. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hiratsuka S, Watanabe A, Aburatani H and
Maru Y: Tumour-mediated upregulation of chemoattractants and
recruitment of myeloid cells predetermines lung metastasis. Nat
Cell Biol. 8:1369–1375. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kwon CH, Moon HJ, Park HJ, Choi JH and
Park Y: S100A8 and S100A9 promotes invasion and migration through
p38 mitogen-activated protein kinase-dependent NF-κB activation in
gastric cancer cells. Mol Cells. 35:226–234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roy R, Yang J and Moses MA: Matrix
metalloproteinases as novel biomarkers and potential therapeutic
targets in human cancer. J Clin Oncol. 27:5287–5297. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moon A, Yong HY, Song JI, Cukovic D,
Salagrama S, Kaplan D, Putt D, Kim H, Dombkowski A and Kim HR:
Global gene expression profiling unveils S100A8/A9 as candidate
markers in H-ras-mediated human breast epithelial cell invasion.
Mol Cancer Res. 6:1544–1553. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Duan L, Wu R, Ye L, Wang H, Yang X, Zhang
Y, Chen X, Zuo G, Zhang Y, Weng Y, et al: S100A8 and S100A9 are
associated with colorectal carcinoma progression and contribute to
colorectal carcinoma cell survival and migration via Wnt/β-catenin
pathway. PLoS One. 8:e620922013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ichikawa M, Williams R, Wang L, Vogl T and
Srikrishna G: S100A8/A9 activate key genes and pathways in colon
tumor progression. Mol Cancer Res. 9:133–148. 2011. View Article : Google Scholar : PubMed/NCBI
|