Introduction
The highly invasive and diffusively infiltrative
nature of glioma, an aggressive malignant cancer of the central
nervous system, is the major cause of conventional treatment
failure and results in a mean survival time of 12 months (1–5). Although
advances have been made in surgery and adjuvant therapy, patients
with malignant glioma have experienced little change in survival
time over recent decades (6,7). However, with the development of
molecular biology, gene therapy is becoming a major focus of tumor
therapy. Therefore, the identification of molecular mechanisms and
novel therapeutic targets to combat tumor invasion are critical for
the treatment of this currently incurable type of cancer.
The ubiquitin proteasome pathway is pivotal for
controlling the degradation of the majority of regulatory proteins
in mammalian cells (8,9). Furthermore, the pathway regulates
various cellular processes by facilitating the prompt destruction
of key regulatory proteins by the 26S proteasome complex (10). Protein ubiquitination in the
proteasome pathway involves the concerted action of an E1
ubiquitin-activating enzyme, an E2 ubiquitin-conjugating enzyme and
an E3 ubiquitin-protein ligase, the latter of which delivers
multiple ubiquitin molecules to the target protein (11–13).
β-transducin repeat-containing E3 ubiquitin protein ligase (β-TrCP)
is characterized by a ~40-amino acid motif. β-TrCP utilizes its
seven WD40 repeats to interact with substrates phosphorylated
within the DSG(X)2+nS destruction motif, and is involved
in the degradation of numerous cell signaling and cell cycle
regulation proteins (14–16).
In recent years, a number of β-TrCP substrates have
been identified, including Bmi1 (17), inhibitor of κB (18), β-catenin (19,20), Emi1
(21), nuclear factor-κB p105 subunit
(22) and cell cycle division 25A
(15). It was previously reported
that β-TrCP is able to promote breast and prostate cancer cell
proliferation and migration (23).
However, β-TrCP may also suppress angiogenesis and thyroid cancer
cell migration (24), as well as
pancreatic cancer cell growth (25).
In addition, β-TrCP appears to inhibit lung cancer cell growth and
invasiveness (26). Although there
have been substantial advances in the understanding of the basic
biology and pathogenesis of β-TrCP, little is known with regard to
the possible role and clinical significance of β-TrCP in glioma.
Therefore, the aim of the present study was to explore the protein
expression levels of β-TrCP in glioma tissues by performing western
blot analysis and immunohistochemical staining. In addition, the
study evaluated the association between the expression of β-TrCP
and survival time during the four-year follow-up period. The aim of
the present study was to provide important data with regard to the
possible role of β-TrCP in human glioma progression and to
determine whether β-TrCP could serve as a novel prognostic marker
for patients with glioma.
Materials and methods
Tissue specimens
Western blot specimens
To detect the protein expression levels of β-TrCP
using western blot analysis, 32 glioma tissue specimens (obtained
during surgical resection) and 16 non-tumorous brain tissue
specimens (obtained from the same brain region during internal
decompression in cases of cerebral trauma) were collected from the
Affiliated Hospital of Xuzhou Medical College (Xuzhou, China).
Sections of the surgically removed tissue specimens were analyzed
to determine a histological diagnosis, and the remaining tissues
were immediately frozen and stored in liquid nitrogen for
subsequent analysis. Clinical staging was performed according to
the 2007 World Health Organization (WHO) classification of tumors
of the central nervous system (28),
revealing six grade I astrocytomas, nine grade II astrocytomas,
eight grade III astrocytomas and nine grade IV glioblastomas.
Written informed consent was obtained from each patient and the
study was approved by the Research Ethics Committee of the
Affiliated Hospital of Xuzhou Medical College.
Immunohistochemical specimens
Paraffin-embedded tissue sections for
immunohistochemical analysis were obtained from the pathology files
of the Department of Pathology at the Affiliated Hospital of Xuzhou
Medical College between March 2007 and January 2010. According to
the 2007 WHO classification of tumors, the 66 glioma samples
included 35 grade I and II astrocytomas, 12 grade III anaplastic
astrocytomas and 19 grade IV glioblastomas. The available clinical
data indicated that none of the tissue specimens were recurrent
glioma, and 56 cases had received radiotherapy and chemotherapy.
The other 10 cases had not received radiotherapy or chemotherapy
due to the patient's poor physical condition following surgery
(Table I). Non-tumorous brain
specimens were acquired from 25 patients undergoing surgery for
internal decompression in cerebral trauma. Written informed consent
was obtained from each patient and the study was approved by the
Research Ethics Committee.
 | Table I.Clinical features and overall survival
of 66 glioma patients with different β-TrCP expression levels. |
Table I.
Clinical features and overall survival
of 66 glioma patients with different β-TrCP expression levels.
|
|
| β-TrCP
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristic | Patients, n | High (n=29) | Low (n=37) | P-value |
|---|
| Gender |
|
|
| 0.803 |
| Male | 42 | 19 | 23 |
|
|
Female | 24 | 10 | 14 |
|
| Age, years |
|
|
| 0.599 |
|
<60 | 45 | 21 | 24 |
|
|
≥60 | 21 | 8 | 13 |
|
| KPS |
|
|
| 0.068 |
|
<80 | 22 | 6 | 16 |
|
|
≥80 | 44 | 23 | 21 |
|
| Post-operative
treatment strategy |
|
|
| 0.493 |
|
Radiotherapy plus
chemotherapy | 56 | 26 | 30 |
|
| No
radiochemotherapy | 10 | 3 | 7 |
|
| Overall
survival |
|
|
| 0.006 |
|
Mortalities, n | 38 | 11 | 27 |
|
|
Censoreda, n | 28 | 18 | 10 |
|
Antibodies and reagents
Rabbit polyclonal anti-β-TrCP antibody (cat no.
ab71753) was purchased from Abcam (Hong Kong, China) and rabbit
monoclonal anti-β-actin antibody (cat no. 04–1116) was purchased
from EMD Millipore (Billerica, MA, USA). The bound antibodies were
detected using a rabbit streptavidin-peroxidase kit and
diaminobenzidine (DAB; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd., Beijing, China).
Western blot analysis
The 32 glioma and 16 non-tumorous human brain
tissues were weighed and ground into small pieces. For protein
analysis, the tissues were homogenized (Pro 200 homogenizer;
Promega Corporation, Madison, WI, USA) in lysis buffer [0.1% SDS,
50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM Triton X-100 and 2 mM
EDTA] containing complete protease inhibitor (0.5 mM
phenylmethylsulfonyl fluoride, 1 mM Na3VO4
and 1 mM NaF). The protein lysates were then concentrated using a
bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Waltham, MA, USA). After the tissues were lysed for 30 min, equal
quantities of protein lysate were subjected to 10% SDS-PAGE, then
transferred to polyvinylidene difluoride membranes with a 0.45-µm
pore size (EMD Millipore). The membranes were saturated with 3%
bovine serum albumin (BSA) and incubated for 2 h at 37°C.
Subsequently, the membranes were incubated with primary rabbit
polyclonal anti-β-TrCP antibody and rabbit monoclonal anti-β-actin
antibody at 4°C overnight. Subsequent being was hed three times
with 0.1% PBS (10 min each), the membranes were incubated with
anti-rabbit secondary antibody at 37°C for 2 h. After was hing
three times with 0.1% PBS (10 min each), the bound antibodies were
detected using Pierce ECL Plus Western Blotting Substrate (Thermo
Fisher Scientific) and exposed to X-ray film. Band densities were
quantified using ImageJ software (National Institutes of Health,
Bethesda, MD, USA). The relative quantity of protein was determined
by normalizing the densitometry value of interest to that of the
internal loading control (β-actin).
Immunohistochemistry (IHC)
Following collection, the paraffin-embedded tissue
blocks were cut in a microtome to a thickness of 4 µm and affixed
onto slides. IHC was performed using a rabbit
streptavidin-peroxidase kit and DAB, according to the
manufacturer's instructions. Briefly, the tissue sections were
dewaxed in xylene and rehydrated through graded alcohol
concentrations using standard procedures. The sections were
subsequently submerged in EDTA (pH 8.0) and autoclaved at 121°C for
5 min to retrieve antigenicity. After was hing three times in
phosphate-buffered saline (PBS; 0.1 M (pH 7.4)] for 5 min,
endogenous peroxidase was blocked by incubation in 3% hydrogen
peroxide for 15 min at room temperature. Next, incubation with
β-TrCP antibody (Abcam), at a dilution of 1:150 in PBS containing
0.5% BSA, was conducted overnight at 4°C. After was hing the
sections in PBS three times (5 min each), the bound antibodies were
detected by applying the streptavidin-peroxidase kit for 30 min at
room temperature. Subsequently, the sections were was hed with PBS
and DAB coloration was applied, followed by the application of a
DAB solution until the color developed. Staining was monitored
under a bright-field microscope (XSP-17C; Shanghai Changfang
Optical Instrument Co., Ltd., Shanghai, China) and the reaction was
terminated by was hing with distilled water. The slides were then
counterstained with hematoxylin, dehydrated with ethanol and
xylene, and covered with coverslips.
Immunohistochemical staining
evaluation
Three independent experienced pathologists, who were
blinded to the clinicopathological data, evaluated the
immunostained slides. The proportion of stained tumor cells in each
selected field was determined by counting individual tumor cells in
three randomly-selected high-magnification fields (magnification,
x400) using light microscopy (IX71+DP721; Olympus Corporation,
Tokyo, Japan). Expression was quantified using a visual grading
system based on the extent and intensity of the staining. The
percentage of positively-stained tumor cells was scored as follows:
0, no positive tumor cells; 1, ≤25% positive tumor cells; 2, 26–50%
positive tumor cells; 3, 51–75% positive tumor cells; and 4, ≥76%
positive tumor cells. Staining intensity was scored as follows: 0,
no staining; 1, very weak staining; 2, weak staining; 3, moderate
staining; and 4, strong staining. The final score was calculated by
multiplying the positive percentage score by the staining intensity
score. A score of 0–3 was considered to indicate low expression and
a score of ≥4 was considered to indicate high expression.
Statistical analysis
SPSS software (version 16.0; SPSS, Inc., Chicago,
IL, USA) was used to perform all statistical analyses. In all
analyses, quantitative data were obtained from a minimum of three
independent experiments and are expressed as the mean ± standard
error of the mean. One-way analysis of variance was used to compare
β-TrCP protein expression levels between glioma and non-tumorous
tissue specimens, and a χ2 test was used to compare
clinical features between the low and high β-TrCP expression
groups. In addition, overall survival (OS) was calculated from the
day of surgery to the date of last follow-up or mortality.
Postoperative survival curves were plotted using the Kaplan-Meier
method and differences in survival rates were assessed by
performing the log-rank test. P<0.05 was considered to indicate
a statistically significant difference.
Results
β-TrCP protein expression levels in
glioma and non-tumorous human brain tissue samples, as determined
by western blotting
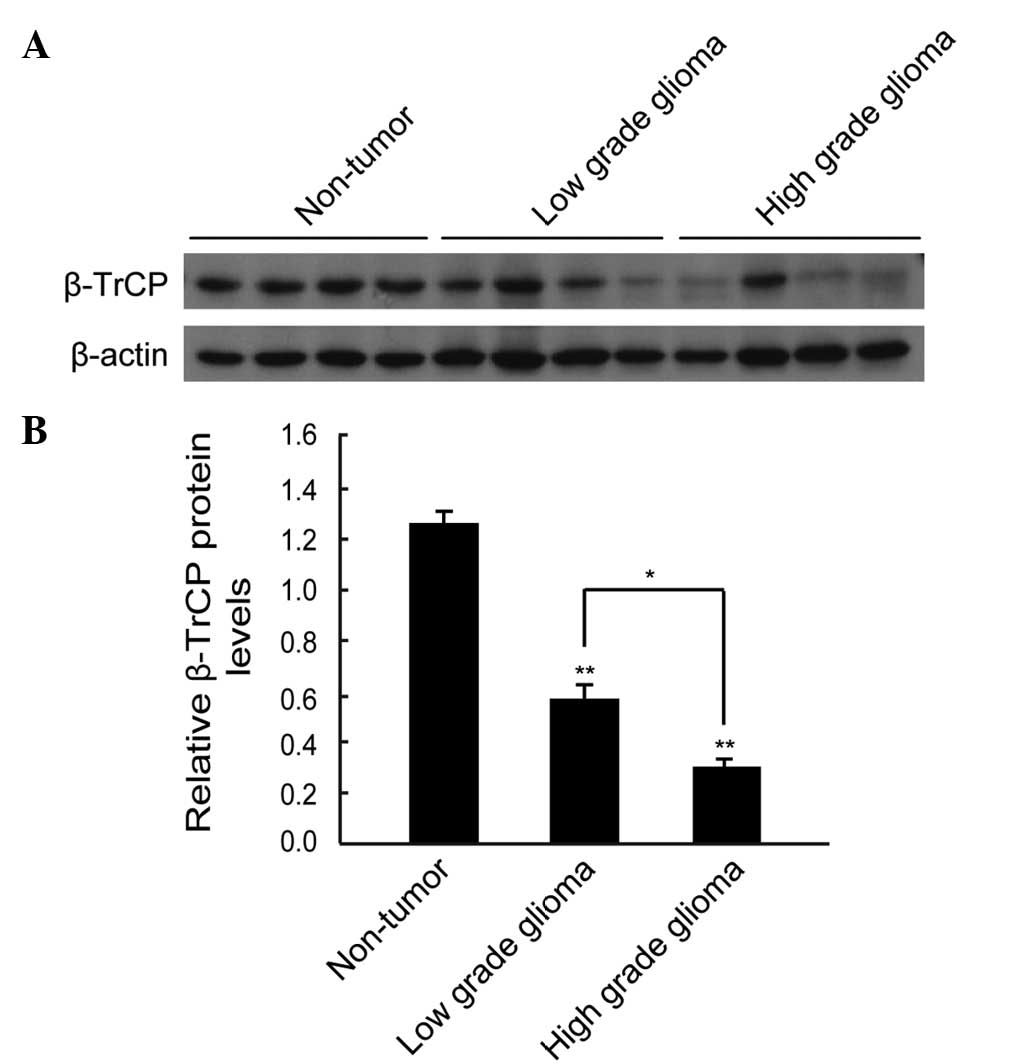
Western blot analysis identified that the expression
level of β-TrCP protein was lower in the majority of the glioma
samples compared with the non-tumorous specimens (Fig. 1A). By performing quantitative
analysis, it was determined that the expression level of β-TrCP
protein was significantly lower in the glioma tissues compared with
the non-tumorous human brain tissues (P<0.01), and the
expression level of β-TrCP protein in high-grade glioma (grades III
and IV) was significantly lower than that in low-grade glioma
(grades I and II; P<0.05) (Fig.
1B).
Immunohistochemical detection of the
expression and location of β-TrCP protein in glioma and
non-tumorous human brain tissues samples
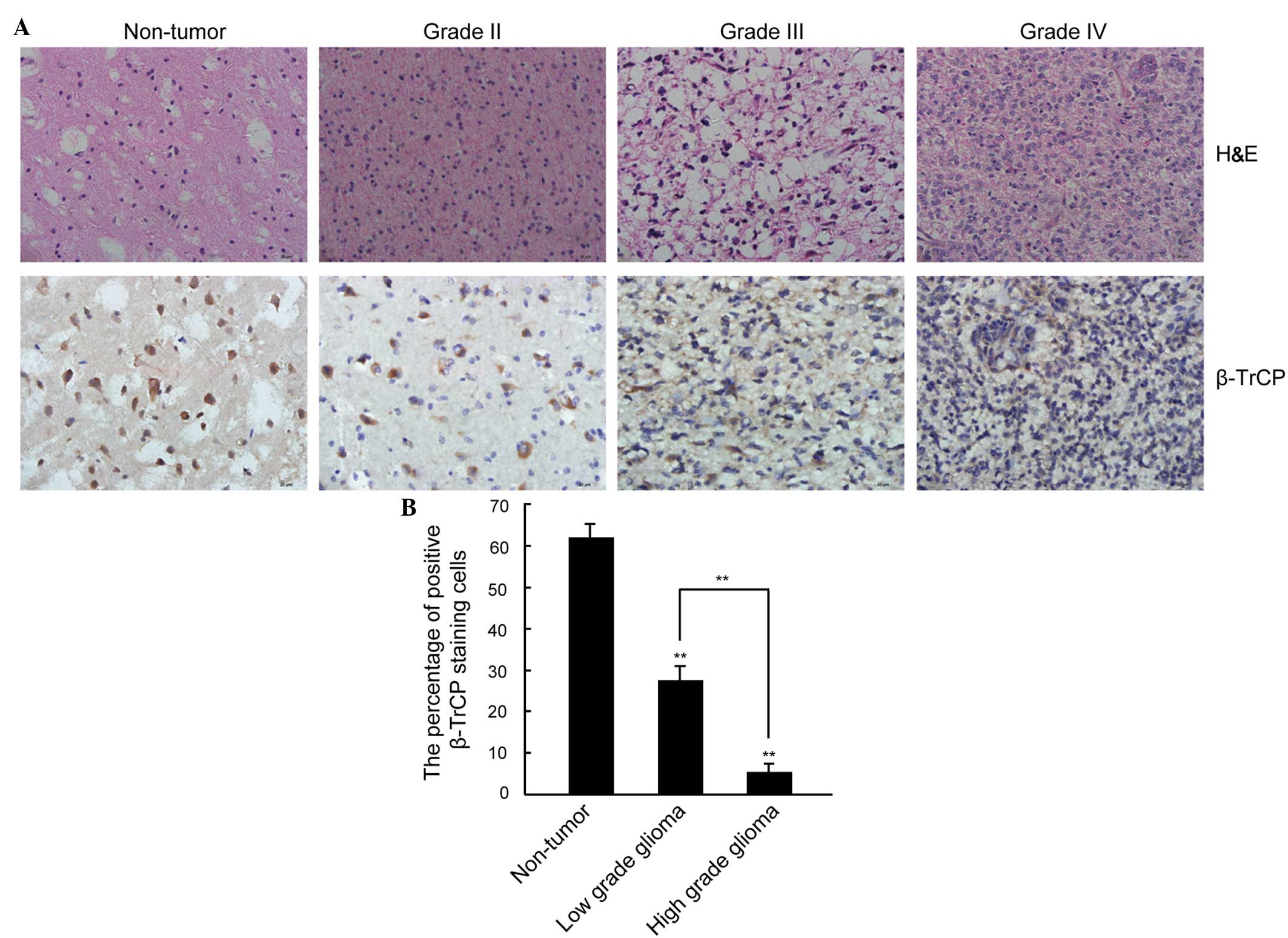
To clarify the association between β-TrCP expression
and the pathological grade of glioma, the expression level and
intracellular location of β-TrCP protein was analyzed in glioma and
non-tumorous human brain tissues by immunohistochemical staining.
As indicated in Fig. 2A, β-TrCP
staining appeared as brown particles and was predominantly located
in the cytoplasm. Furthermore, the expression level of β-TrCP was
significantly lower in the glioma tissue samples compared with in
the non-tumorous brain tissue samples. In addition, the high grade
glioma was associated with a significantly lower level of β-TrCP
expression compared with low grade glioma, with weak expression of
β-TrCP only identified in a small number of cell nuclei
(non-tumorous nuclei, 62.09±2.36% positive β-TrCP staining; low
grade nuclei, 27.65±2.75% positive β-TrCP staining; high grade
nuclei, 5.41±1.94% positive β-TrCP staining; P<0.01; Fig. 2B).
Impact of β-TrCP expression on the OS
of patients with glioma
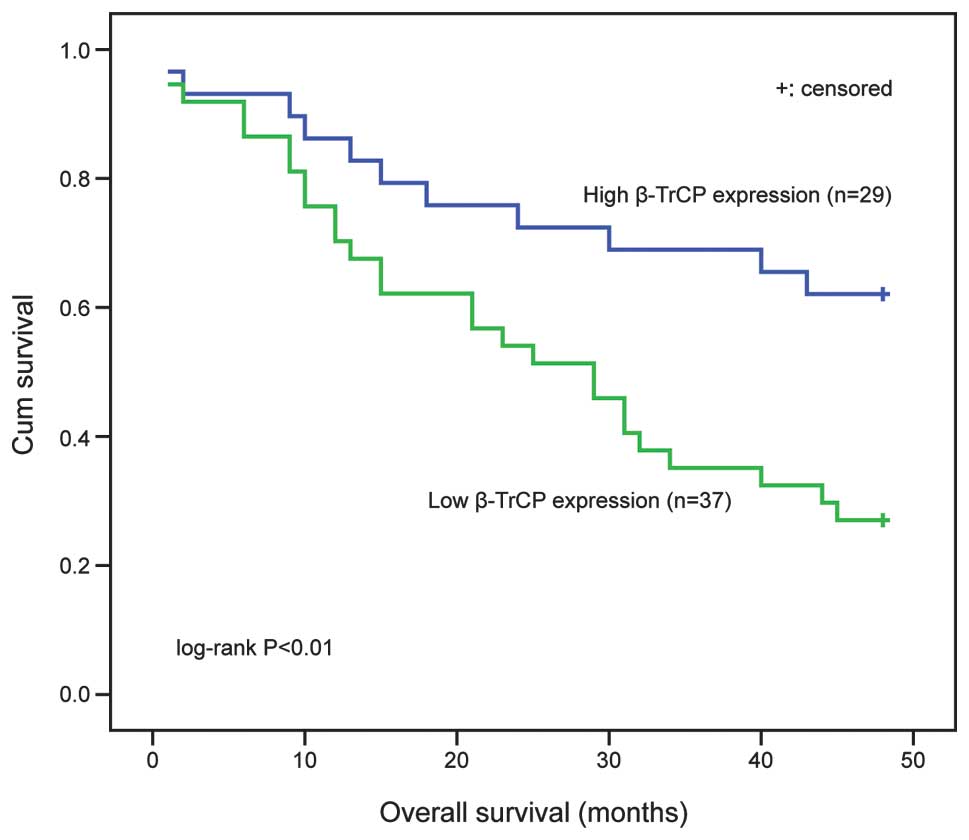
The prognostic value of β-TrCP expression on the OS
of patients with glioma was evaluated in the present study. There
were no statistical differences in the pre-operative clinical data
and postoperative treatment strategies between the low and high
β-TrCP expression groups (Table I).
Kaplan-Meier and log-rank analysis determined that glioma patients
with high β-TrCP expression had significantly improved OS compared
with those with low expression (P<0.01; Fig. 3). These results indicate that β-TrCP
protein expression levels are downregulated in human glioma tissue
and that β-TrCP expression levels are significantly associated with
the malignant state of glioma. These data provide early evidence
for β-TrCP as an important factor in the maintenance of normal
conditions in brain tissue and as a negative factor in the
development of human glioma.
Discussion
Previous studies have identified that β-TrCP
recruits phosphorylated substrates to the Skp, Cullin, F-box
ubiquitin ligase complex (18,29).
Considering the diversity in its substrates, β-TrCP may be
responsible for oncogenesis or the inhibition of tumorigenesis. For
example, previous studies have determined that β-TrCP is involved
in the oncogenesis of gastric, prostate and breast cancer cell
lines (30–32), with overexpression of β-TrCP mRNA and
protein expression levels observed in colorectal cancer (33). β-TrCP has been identified to act as a
tumor suppressor gene in various types of solid cancer, such as
thyroid and lung cancer (24,26). However, to the best of our knowledge,
no studies have thus far investigated β-TrCP expression levels in
human glioma tissue.
In the present study, western blot analysis
identified that the levels of β-TrCP protein expression in the
glioma tissues were significantly lower than those in the
non-tumorous tissues, and that an increase in glioma grade was
significantly associated with a gradual reduction in the overall
expression of β-TrCP protein. IHC supported the observation that
the level of β-TrCP was significantly lower in the glioma tissues
compared with the non-tumorous brain tissues, and identified that
β-TrCP was predominantly located in the cytoplasm. Furthermore,
weak β-TrCP expression was observed in a small number of cell
nuclei. Warfel et al (34)
identified higher β-TrCP1 protein expression levels in the cytosol
compared with the nucleus in astrocyte and astrocytoma cell lines.
However, in contrast to the present study, the expression level of
β-TrCP1 protein was lower in the cytosol of glioblastoma cells
compared with the nuclei. These findings indicate that β-TrCP may
be an important factor for the maintenance of normal conditions in
glioma tissue, and that its nuclear deficiency in astrocytes may
contribute to glioma formation and progression.
Furthermore, in the present study, Kaplan-Meier
survival analysis determined that the OS period of patients
exhibiting tumors with low β-TrCP expression was significantly
worse than that of patients with high β-TrCP expression. These
results indicated that the detection of decreased β-TrCP expression
may facilitate the identification of glioma patients with a poor
prognosis. Thus, β-TrCP may be a novel prognostic marker for
patients with glioma. However, due to the limited sample size and
short duration of patient follow-up in the present study, more
studies with larger sample sizes are required to clarify these
findings. A previous study identified that β-TrCP regulates Bmi-1
protein turnover via ubiquitination and degradation in a human
osteosarcoma cell line (17).
Furthermore, Bmi-1 appears to be highly expressed in patients with
glioma, and promotes glioma cell invasion, migration and
proliferation (35,36). The high expression of Bmi-1 also
predicted poor OS in patients with glioma (37–39).
Therefore, it is proposed that the function of β-TrCP in glioma may
be mediated by Bmi-1. Additional studies are required to clarify
the specific molecular mechanism underlying the association between
β-TrCP expression and Bmi-1.
In conclusion, the data obtained in the current
study indicates that the β-TrCP expression level is low in glioma
and is associated with the poor prognosis of glioma patients.
Therefore, β-TrCP may serve as a novel prognostic marker for
patients with glioma.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (no. 81072072) and
Xuzhou Medical College (no. 09KJZ18).
References
|
1
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Buonerba C, Di Lorenzo G, Marinelli A, et
al: A comprehensive outlook on intracerebral therapy of malignant
gliomas. Crit Rev Oncol Hematol. 80:54–68. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sherman JH, Hoes K, Marcus J, Komotar RJ,
Brennan CW and Gutin PH: Neurosurgery for brain tumors: update on
recent technical advances. Curr Neurol Neurosci Rep. 11:313–319.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Onishi M, Ichikawa T, Kurozumi K and Date
I: Angiogenesis and invasion in glioma. Brain Tumor Pathol.
28:13–24. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim CS, Jung S, Jung TY, Jang WY, Sun HS
and Ryu HH: Characterization of invading glioma cells using
molecular analysis of leading-edge tissue. J Korean Neurosurg Soc.
50:157–165. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ohgaki H and Kleihues P: Population-based
studies on incidence, survival rates, and genetic alterations in
astrocytic and oligodendroglial gliomas. J Neuropathol Exp Neurol.
64:479–489. 2005.PubMed/NCBI
|
|
7
|
Rutka JT, Taylor M, Mainprize T, Langlois
A, Ivanchuk S, Mondal S and Dirks P: Molecular biology and
neurosurgery in the third millennium. Neurosurgery. 46:1034–1051.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
DeSalle LM and Pagano M: Regulation of the
G1 to S transition by the ubiquitin pathway. FEBS Lett.
490:179–189. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pickart CM: Mechanisms underlying
ubiquitination. Annu Rev Biochem. 70:503–533. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ciechanover A, Orian A and Schwartz AL:
Ubiquitin-mediated proteolysis: biological regulation via
destruction. BioEssays. 22:442–451. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou P: Targeted protein degradation. Curr
Opin Chem Biol. 9:51–55. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hershko A and Ciechanover A: The ubiquitin
system. Annu Rev Biochem. 67:425–479. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maniatis T: A ubiquitin ligase complex
essential for the NF-kappaB, Wnt/Wingless and Hedgehog signaling
pathways. Genes Dev. 13:505–510. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu C, Kato Y, Zhang Z, Do VM, Yankner BA
and He X: beta-Trcp couples beta-catenin
phosphorylation-degradation and regulates Xenopus axis formation.
Proc Natl Acad Sci USA. 96:6273–6278. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Busino L, Donzelli M, Chiesa M, et al:
Degradation of Cdc25A by beta-TrCP during S phase and in response
to DNA damage. Nature. 426:87–91. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Watanabe N, Arai H, Nishihara Y, Taniguchi
M, Watanabe N, Hunter T and Osada H: M-phase kinases induce
phospho-dependent ubiquitination of somatic Weel by SCFbeta-TrCP.
Proc Natl Acad Sci USA. 101:4419–4424. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sahasrabuddhe AA, Dimri M, Bommi PV and
Dimri GP: βTrCP regulates BMI1 protein turnover via ubiquitination
and degradation. Cell Cycle. 10:1322–1330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fuchs SY, Chen A, Xiong Y, Pan ZQ and
Ronai Z: HOS, a human homolog of Slimb, forms an SCF complex with
Skp1 and Cullin1 and targets the phosphorylation-dependent
degradation of IkappaB and beta-catenin. Oncogene. 18:2039–2046.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hart M, Concordet JP, Lassot I, et al: The
F-box protein beta-TrCP associates with phosphorylated beta-catenin
and regulates its activity in the cell. Curr Biol. 9:207–210. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Latres E, Chiaur DS and Pagano M: The
human F box protein beta-Trcp associates with the Cul1/Skp1 complex
and regulates the stability of beta-catenin. Oncogene. 18:849–854.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Margottin-Goguet F, Hsu JY, Loktev A,
Hsieh HM, Reimann JD and Jackson PK: Prophase destruction of Emi1
by the SCF (betaTrCP/Slimb) ubiquitin ligase activates the anaphase
promoting complex to allow progression beyond prometaphase. Dev
Cell. 4:813–826. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lang V, Janzen J, Fischer GZ, et al:
betaTrCP-mediated proteolysis of NF-kappaB1 p105 requires
phosphorylation of p105 serines 927 and 932. Mol Cell Biol.
23:402–413. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhong J, Shaik S, Wan L, et al: SCF β-TRCP
targets MTSS1 for ubiquitination-mediated destruction to regulate
cancer cell proliferation and migration. Oncotarget. 4:2339–2353.
2013.PubMed/NCBI
|
|
24
|
Shaik S, Nucera C, Inuzuka H, et al: SCF
(β-TRCP) suppresses angiogenesis and thyroid cancer cell migration
by promoting ubiquitination and destruction of VEGF receptor 2. J
Exp Med. 209:1289–1307. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wan M, Huang J, Jhala NC, et al:
SCF(beta-TrCP1) controls Smad4 protein stability in pancreatic
cancer cells. Am J Pathol. 166:1379–1392. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He N, Li C, Zhang X, et al: Regulation of
lung cancer cell growth and invasiveness by beta-TRCP. Mol
Carcinog. 42:18–28. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Peus D, Newcomb N and Hofer S: Appraisal
of the Karnofsky Performance Status and proposal of a simple
algorithmic system for its evaluation. BMC Med Inform Decis Mak.
13:722013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rosenblum MK: The 2007 WHO Classification
of Nervous System Tumors: newly recognized members of the mixed
glioneuronal group. Brain Pathol. 17:308–313. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Scherer DC, Brockman JA, Chen Z, Maniatis
T and Ballard DW: Signal-induced degradation of I kappa B alpha
requires site-specific ubiquitination. Proc Natl Acad Sci USA.
92:11259–11263. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao G, Kun T, Sheng Y, et al: SGT1
regulates Akt signaling by promoting beta-TrCP-dependent PHLPP1
degradation in gastric cancer cells. Mol Biol Rep. 40:2947–2953.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gluschnaider U, Hidas G, Cojocaru G,
Yutkin V, Ben-Neriah Y and Pikarsky E: beta-TrCP inhibition reduces
prostate cancer cell growth via upregulation of the aryl
hydrocarbon receptor. PLoS One. 5:e90602010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tang W, Li Y, Yu D, Thomas-Tikhonenko A,
Spiegelman VS and Fuchs SY: Targeting beta-transducin
repeat-containing protein E3 ubiquitin ligase augments the effects
of antitumor drugs on breast cancer cells. Cancer Res.
65:1904–1908. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ougolkov A, Zhang B, Yamashita K, et al:
Associations among beta-TrCP, an E3 ubiquitin ligase receptor,
beta-catenin and NF-kappaB in colorectal cancer. J Natl Cancer
Inst. 96:1161–1170. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Warfel NA, Niederst M, Stevens MW, Brennan
PM, Frame MC and Newton AC: Mislocalization of the E3 ligase,
β-transducin repeat-containing protein 1 (β-TrCP1), in glioblastoma
uncouples negative feedback between the pleckstrin homology domain
leucine-rich repeat protein phosphatase 1 (PHLPP1) and Akt. J Biol
Chem. 286:19777–19788. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu Z, Wang Q, Wang L, et al: Combined
aberrant expression of Bmi1 and EZH2 is predictive of poor
prognosis in glioma patients. J Neurol Sci. 335:191–196. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tu Y, Gao X, Li G, et al: MicroRNA-218
inhibits glioma invasion, migration, proliferation and cancer
stem-like cell self-renewal by targeting the polycomb group gene
Bmi1. Cancer Res. 73:6046–6055. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vlachostergios PJ and Papandreou CN: The
Bmi-1/NF-ĸB/VEGF story: another hint for proteasome involvement in
glioma angiogenesis? J Cell Commun Signal. 7:235–237. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Y, Hu J, Guan F, et al: Copper induces
cellular senescence in human glioblastoma multiforme cells through
downregulation of Bmi-1. Oncol Rep. 29:1805–1810. 2013.PubMed/NCBI
|
|
39
|
Jiang L, Song L, Wu J, et al: Bmi-1
promotes glioma angiogenesis by activating NF-ĸB signaling. PLoS
One. 8:e555272013. View Article : Google Scholar : PubMed/NCBI
|