Introduction
Glioma is the most common type of primary brain
tumor, accounting for ~80% of all brain and central nervous system
tumors. Glioma is divided into four grades (grade I-IV) according
to criterion established by the World Health Organization. The
survival rate of low-grade glioma (grade II) is higher than that of
high-grade glioma (anaplastic astrocytomas and glioblastoma
multiforme), at ~47% (1). High-grade
gliomas are the most malignant tumors and are characterized by high
cellularity, mitotic activity, vascular proliferation, central
necrosis and aggressive invasion into normal brain tissue (2). Despite the combination of maximal tumor
resection, concurrent radiotherapy, adjuvant chemotherapy and
molecular targeted therapy for the treatment of high grade glioma,
the life expectancy of patients remains poor, at just 12–14 months
(3–6).
Therefore, the identification of novel, effective therapies for the
treatment of gliomas is required.
Artemisinin is derived from a traditional Chinese
herb, which is used for the treatment of fever and malaria
(7). Previous studies have revealed
that artemisinin is able to inhibit the progression of chronic
myelogenous leukemia, pancreatic adenocarcinoma, colon cancer,
prostate cancer, non-small cell lung cancer and breast malignancies
at low concentrations (8–10). Artemisinin and its derivatives have
also been used in the screening of anticancer drugs by the National
Cancer Institute (NCI) (11).
Dihydroartemisinin (DHA), the major active
metabolite of artemisinin, has been found to inhibit the growth of
cancer cells in a variety of cancers (12–15). Xie
et al (16) reported that the
concentration of DHA was two-fold higher in the brain than in the
plasma, which indicates that DHA is able to penetrate the
brain-blood barrier. DHA triggers the production of reactive oxygen
species (ROS) and inhibits the activity of
glutathione-S-transferase, thereby promoting the radiosensitivity
of glioma cells (17). Furthermore,
DHA has been found to potentiate the cytotoxic effect of
temozolomide (TMZ) in rat C6 glioma cells (18). However, whether DHA is able to enhance
the efficiency of TMZ in human glioma cells has remained to be
elucidated. The present study aimed to assess the effect of DHA on
glioma cell lines as well as whether DHA could enhance the cell
response to TMZ.
Materials and methods
Cell lines and reagents
In total, 10 human glioma cell lines, namely MGR1,
MGR3, SF295, SKMG-1, SKMG-4, T98G, U251, U251/CP2, U373 and U87,
were used in the present study. All cells were maintained in
Dulbecco's modified Eagle's medium /F-12 (Gibco Life Technologies,
Grand Island, NY, USA) supplemented with 10% fetal bovine serum (GE
Healthcare Life Sciences, Chalfont, UK) and 1.0%
penicillin/streptomycin (Gibco Life Technologies) in a humidified
incubator at 37°C with 5% CO2. DHA (Sigma-Aldrich, St.
Louis, MO, USA) was dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich; 20 µg/ml) and stored at −20°C in the dark. TMZ
(Sigma-Aldrich) was dissolved in DMSO, (stock concentration, 0.2 M)
and stored at −20°C.
Cytotoxicity assay
The glioma cells were seeded into 96-well plates at
a density of 5×103 cells/well and incubated overnight at
37°C. Next, the cells were treated with DHA at serial
concentrations (0, 0.3125, 0.625, 1.25, 2.5, 5.0, 10.0 and 20.0
µg/ml) for 72 h. Cell viability was analyzed by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay. In order to detect the effect of DHA on TMZ efficiency, the
cells were treated with 2 µM DHA in combination with various
concentrations of TMZ (ranging between 31.25 and 2,000 µM).
Following 72 h of incubation, the MTT assay was performed and the
half maximal inhibitory concentration (IC50) was
calculated. Briefly, MTT (Sigma-Aldrich) was added to cells
following treatment and incubated for 4 h at 37°C. DMSO was
subsequently added and absorbance was measured by microplate reader
(Multiskan MK3; Thermo Fisher Scientific, Waltham, MA, USA) after
shaking for ~30 sec.
Western blot analysis
The cell lysates were collected using
radioimmunoprecipitation assay lysis buffer (1% NP-40, 1% sodium
deoxycholate, 0.1% SDS, 150 mM NaCl and 10 mM
Na2HPO4; pH 7.2; Beyotime Institute of
Biotechnology, Japan), supplemented with a protease inhibitor
cocktail (Roche Diagnostics GmbH, Mannheim, Germany).
Immunoblotting was performed as previously described (19). Rabbit anti-human antibodies against
LC3-B (polyclonal; catalog no. ab48394; dilution, 1:1,000; Abcam,
Cambridge, MA, USA), Beclin-1 (monoclonal; catalog no. ab51031;
dilution, 1:750; Abcam) and Caspase-3 (polyclonal; catalog no.
ab90437; dilution, 1:100; Abcam) were added and incubated overnight
at 4°C. A mouse anti-human β-actin antibody (catalog no. AA128;
dilution, 1:1,000; Beyotime Institute of Biotechnology) was loaded
as the internal control. The membranes were then washed three times
with phosphate-buffered saline (PBS). Horseradish
peroxidase-conjugated goat anti-mouse (dilution, 1:1,000; catalog
no. A0216; Beyotime Institute of Biotechnology) and goat
anti-rabbit (dilution, 1:1,000; catalog no. 7074; Cell Signaling
Technology Inc., Danvers, MA, USA) secondary antibodies were
applied for 30 min at room temperature. The membranes were then
washed three times with PBS and incubated with enhanced
chemiluminescence reagent (Millipore, Billerica, MA, USA) according
to the manufacturer's instructions. Next, the membranes were
exposed to ECL sensitive films (Hyperfilm ECL; GE Healthcare
Bio-Sciences, Pittsburgh, PA, USA), and developed using an OPTIMAX
X-Ray film processor (Protec GmbH & Co. KG, Oberstenfeld,
Germany). All protein bands were normalized to β-actin, which was
used as the internal loading control.
Monodansylcadaverine (MDC)
fluorescence staining
The glioma SKMG-4 cells, which are sensitive to DHA,
were seeded into a 30-mm culture dish and treated with 2 µg/ml DHA
for 24 h. MDC (Sigma-Aldrich), at a concentration of 0.05 mM, was
then added for 10 min. Next, the cells were fixed in 4%
paraformaldehyde (Sigma-Aldrich) for 15 min and washed twice with
phosphate-buffered saline. The cytoplasmic autophagic vacuoles were
then observed under a fluorescence microscope (Olympus BX61;
Olympus Corp., Tokyo, Japan).
In vivo glioma formation assay
The U251 cells were maintained in standard culture
medium, adjusted to 5×106 cells/ml. Next, 300 µl cell
suspension was injected into the axillary region of the nude mice
(age, 4–5 weeks; mean weight, 22.6 g). Mice were maintained in
specific-pathogen-free grade room, with 6 mice per cage, at 18–29°C
and with light from 7:30 am to 19:30 pm. Two weeks later, the tumor
masses were resected, cut into sections measuring 2 mm3
and transplanted subcutaneously into the backs of a second set of
experimental nude mice. These mice were randomly separated into
four groups (n=10) and treated for 5 continuous days with either
DMSO (control), 50 mg/kg DHA only, 50 mg/kg TMZ only, or 50 mg/kg
DHA plus 50 mg/kg TMZ. The tumor sizes were measured using a
vernier caliper every three days during the following month. The
tumor volumes (TV) were calculated according to the following
formula: TV = 1/2 × length × width2 (20). The present study was approved by the
ethics committee of Sun Yat-sen University Cancer Center
(Guangzhou, China) according to Institutional Animal Care and Use
Committee protocols. Chloral hydrate was applied for anesthesia
(10%; 80 µl per mouse), and mice were sacrificed by cervical
dislocation.
Statistical analysis
The data is expressed as the mean ± standard error
of the mean. Student's t-test was used to determine any
statistically significant differences. P-values are two sided, with
P<0.05 considered to indicate a statistically significant
difference. Statistical analysis was performed using SPSS 16.0
(SPSS, Inc., Chicago, IL, USA).
Results
DHA is cytotoxic to glioma cells in
vitro
The IC50 of ten glioma cell lines treated
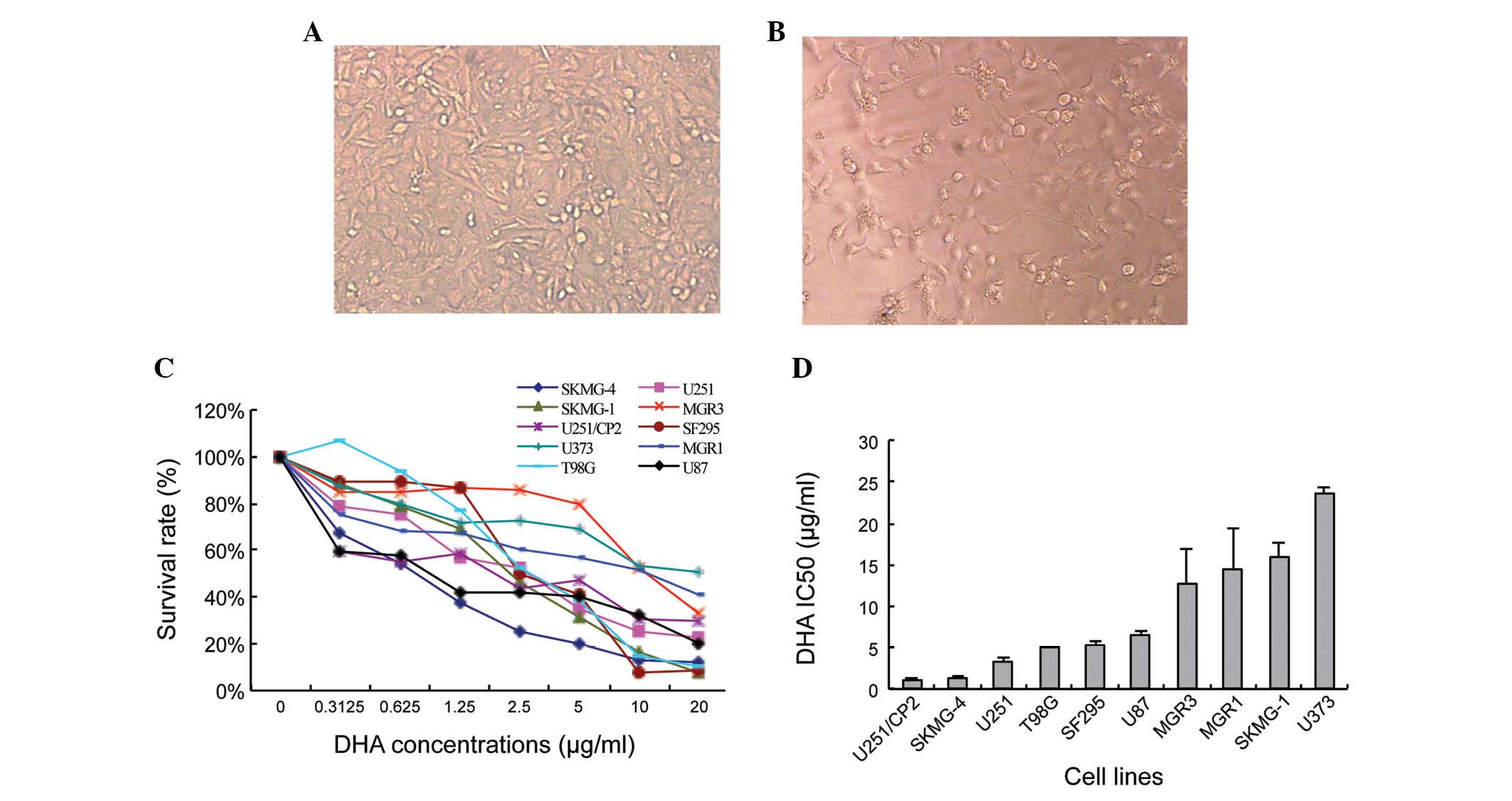
with serial concentrations DHA was calculated. Following treatment,
the SKMG-4 cells appeared to be low in density and exhibit abnormal
morphologies compared with the non-treated cells (Fig. 1A and B). The extent of DHA toxicity
was mild and differed amongst the glioma cell lines (Fig. 1C). The IC50 also varied
between the ten human glioma cell lines, from 1.17±0.08 µg/ml in
U251/CP2 cells to 23.57±0.80 µg/ml in U373 cells (Fig. 1D).
DHA activates autophagy, rather than
apoptosis, in SKMG-4 cells
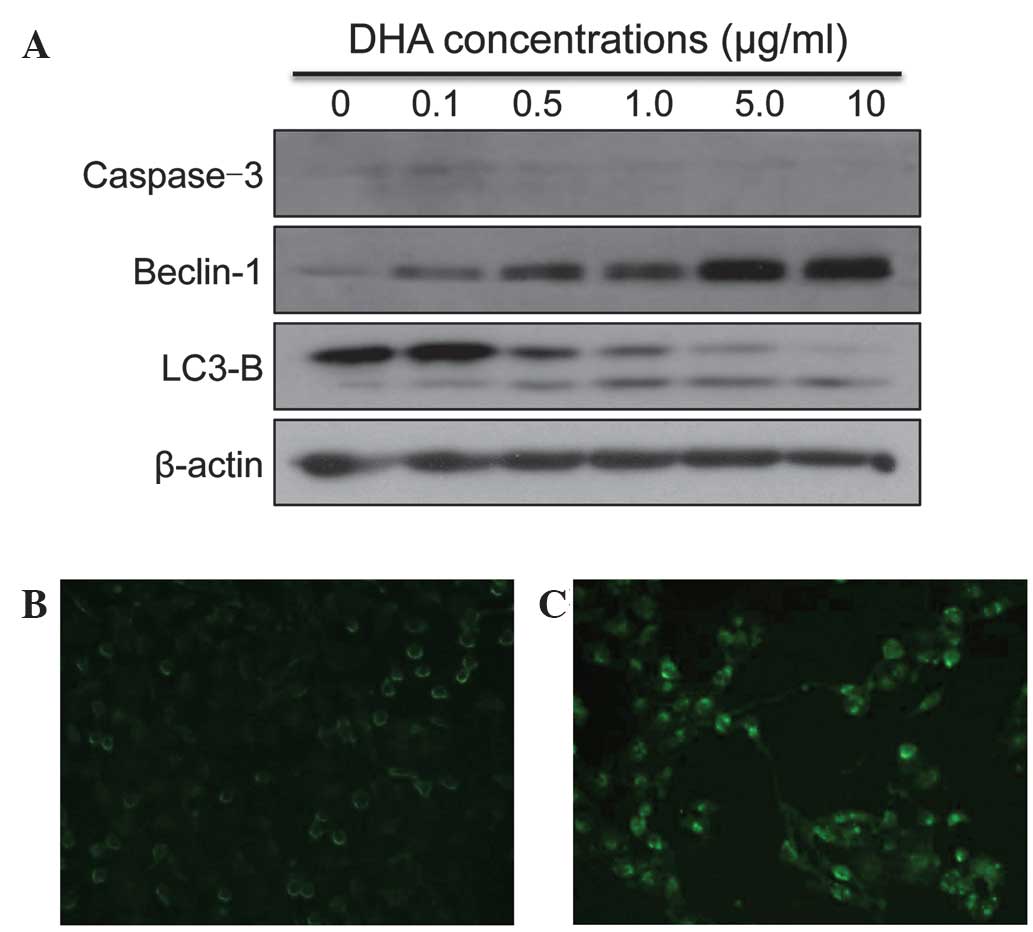
The cell lysates were collected from SKMG-4 cells
following treatment with DHA for 72 h. Markers of apoptosis and
autophagy were then detected by western blotting. There were no
significant alterations to the expression of Caspase-3 following
DHA treatment. However, the expression of the autophagy markers,
Beclin-1 and LC3-B, increased upon DHA treatment in a
dose-dependent manner (Fig. 2A). The
autofluorescent reagent MDC, a specific autophagic vacuole marker,
was also applied in order to monitor the process of autophagy.
Following culture with DHA and MDC, the SKMG-4 cells were fixed and
observed under a fluorescent microscope. It was revealed that MDC
accumulated in DHA-treated cells, but not in untreated control
cells (Fig. 2B and C). The data
therefore indicated that autophagy was induced by DHA in glioma
cell lines.
DHA promotes the cytotoxicity of TMZ
in glioma cells
The present study aimed to investigate whether
combined treatment with DHA and TMZ was able to increase cell
sensitivity to TMZ. In total, four glioma cell lines, namely
SKMG-4, U251, U373 and T98G, were used for the combination
treatment due to the differences in their sensitivity to DHA and
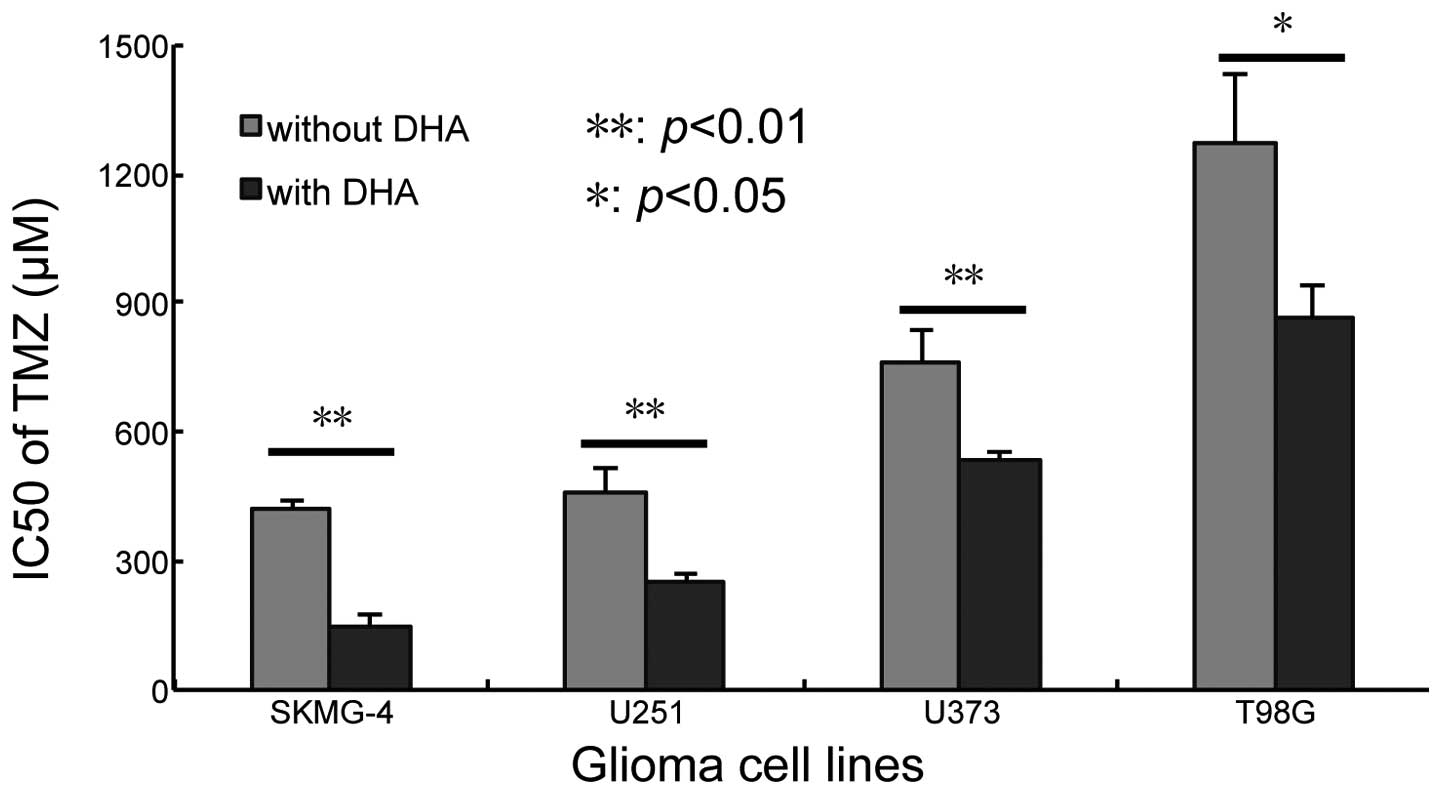
their genetic background. The 10% inhibition concentration of TMZ
(IC10) was calculated according to the results of the
cytotoxicity assay (Fig. 1C). The
concentration of DHA was fixed to the IC10 of each cell
line and then combined with serial concentrations of TMZ. The
IC50 of TMZ was calculated following co-treatment for 72
h. It was revealed that the application of DHA reduced the
IC50 of TMZ in all four cell lines; from 417.36±17.92 to
143.90±28.74 µM in SKMG-4 cells (P<0.01), 455.06±64.39 to
246.84±26.14 µM in U251 cells (P<0.01), 759.03±76.96 to
537.15±11.88 µM in U373 cells (P<0.01) and 1274.72±159.70 to
863.20±73.26 µM in T98G cells (P<0.05) (Fig. 3). These data indicated that DHA
sensitized glioma cell lines to TMZ.
DHA strengthens the tumor inhibitory
effect of TMZ
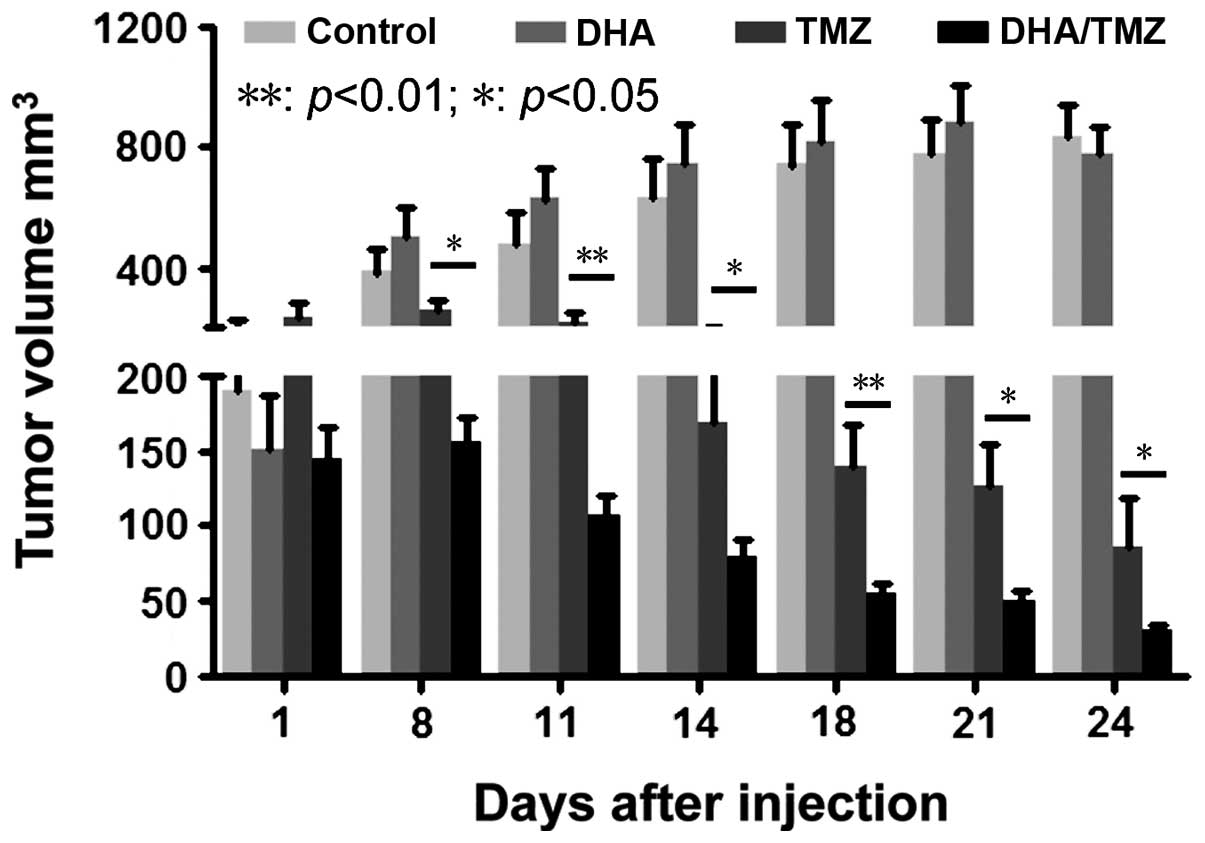
The glioma U251 cell line was used for the in
vivo tumor formation and inhibition assay, due to its ability
to form tumors in xenografts in vivo and its sensitivity to
DHA (low IC50) in vitro. The tumor size was
monitored every 3 days, following continuous treatment for 5 days
with DMSO, DHA, TMZ and DHA+TMZ. It was revealed that 50 mg/kg DHA
alone had no effect on tumor growth in vivo compared with
the DMSO-treated control cells. By contrast, TMZ significantly
suppressed tumor growth compared with that of the DMSO-treated
control cells. Co-treatment with DHA and TMZ significantly reduced
the tumor size compared with the effects of TMZ alone (Fig. 4; P<0.01, P<0.05). This indicated
that DHA was able to strengthen the tumor inhibitory effect of TMZ
in vivo. Taken together, the data demonstrated that DHA
inhibited tumor cell proliferation and sensitized glioma cells to
TMZ in vitro and in vivo, potentially through the
activation of autophagy.
Discussion
Glioma is the most common primary malignant brain
tumor amongst adults. The median survival time of patients with
high-grade glioma is generally <12 months, even with surgical
resection, radiotherapy and chemotherapy (21–23).
Therefore, the identification of novel, effective treatment
strategies is necessary. Adjuvant chemotherapeutic reagents, as
well as various combination therapies, have been extensively
studied. Drugs used for combination therapies usually have clear
pharmacokinetics, which leads to improved therapeutic efficiencies
and tolerance, and reduces drug resistance and side effects
(19,24,25).
Due to their efficiency and few side effects,
traditional Chinese medicines have been extensively studied in
anti-cancer research. Artemisinin is a Chinese herb derived from
the plant, Artemisia annua L., and has been used in the
treatment of fevers for >1,000 years (26). Rasheed et al (27) reported that artesunate, a derivative
of artemisinin, was able to inhibit the proliferation and
metastasis of non-small cell lung carcinomas in a chicken embryo
model. A further study, which included 55 cancer cell lines, found
that artesunate demonstrated anti-tumor activities in several types
of tumor cell, including leukemia, as well as breast, ovarian,
cervical, stomach, liver, pancreatic, colon and lung cancer
(28). Artemisinin was therefore
enrolled by the NCI in their drug-screening program. Previous data
has suggested that artemisinin may be involved in multiple
pathways. Screening approaches have established that artemisinin is
involved in the inhibition of proliferation, the promotion of
apoptosis and the inhibition of angiogenesis (28). Further studies have revealed that
artemisinin contributes to the generation of ROS, which leads to
heme-mediated endoperoxide cleavage, DNA damage and apoptosis
(29). In addition, it has been found
that artemisinin enhances the sensitivity of glioma cells to β,
γ-rays (17).
As the major active metabolite of artemisinin, DHA
has advantages over the original compound. The anti-cancer effects
of DHA have been previously reported in several types of cancer,
but not glioma. The present study identified that DHA mildly
inhibited the growth of glioma cells. It was hypothesized that the
inhibition of proliferation exerted by DHA may be cell-type
specific, since its efficiency in the 10 human glioma cell lines
evaluated was varied.
The findings of the present study indicated that the
mechanisms underlying DHA-induced proliferation arrest may involve
autophagy activation, since the expression of autophagic molecular
markers increased following DHA treatment. Autophagy is a
self-digestive process that degrades intracellular structures in
response to oxidative stresses, ultimately leading to cell survival
(30). However, cells die following
prolonged autophagic activity. It has been established that cancer
cells produce higher levels of ROS than those of normal cells, as a
result of metabolic stress and proliferative requirements. A
previous study reported that DHA induced autophagy in the
mitochondria of leukemia K562 cells (31). A further study found that the
sensitivity of cancer cells to artemisinin was attenuated following
the overexpression of oxidative stress associated enzymes (32). The detailed mechanisms underlying the
role of DHA in glioma cell autophagy will be investigated in future
studies.
TMZ is an alkylating agent, whose therapeutic
benefit depends upon its ability to alkylate DNA at the
N7 or O6 positions of guanine residues. The
low cytotoxicity, high lipid solubility and small molecular weight
of TMZ facilitates its passage through the blood-brain barrier.
These properties established TMZ as an effective, first-line
chemotherapeutic drug for glioma patients. Kanzawa et al
(33) identified that TMZ
cytotoxicity resulted from the induction of autophagy, rather than
apoptosis, in malignant glioma cells as the blockage of autophagy
significantly decreased the anti-tumor effect of TMZ. The results
of the present study revealed that DHA interacted with TMZ to
enhance the anti-cancer effect in vitro and in
vivo.
In conclusion, DHA may inhibit the growth of glioma
cells through the induction of autophagy. Furthermore, cell
response to TMZ may be enhanced significantly by co-treatment of
DHA, both in vitro and in vivo. However, the
mechanisms underlying the interaction between DHA and TMZ in
autophagy and other signaling pathways require further
investigation.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (no. 81372685), the
National High Technology Research and Development Program 863 (no
2012AA02A508), the Department of Science and Technology of
Guangdong Province (no. 2011B031800178), the Natural Science
Foundation of Guangdong Province (no. 10151040701000035), the Hong
Kong Scholars Program (no. XJ2012059), the China Postdoctoral
Science Foundation (no. 2013M540678) and the Fundamental Research
Funds for Central Universities (no. 12ykpy49).
References
|
1
|
Smoll NR, Gautschi OP, Schatlo B, Schaller
K and Weber DC: Relative survival of patients with supratentorial
low-grade gliomas. Neuro Oncol. 14:1062–1069. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nakazato Y: The 4th edition of WHO
classification of tumours of the central nervous system published
in 2007. No Shinkei Geka. 36:473–491. 2008.(In Japanese).
PubMed/NCBI
|
|
3
|
Stupp R, Mason WP, van den Bent MJ, et al:
National Cancer Institute of Canada Clinical Trials Group:
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Davis FG, McCarthy BJ, Freels S, Kupelian
V and Bondy ML: The conditional probability of survival of patients
with primary malignant brain tumors: Surveillance, epidemiology and
end results (SEER) data. Cancer. 85:485–491. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Giese A, Bjerkvig R, Berens ME and
Westphal M: Cost of migration: Invasion of malignant gliomas and
implications for treatment. J Clin Oncol. 21:1624–1636. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stupp R, Hegi ME, Mason WP, et al:
European Organisation for Research and Treatment of Cancer Brain
Tumour and Radiation Oncology Groups; National Cancer Institute of
Canada Clinical Trials Group: Effects of radiotherapy with
concomitant and adjuvant temozolomide versus radiotherapy alone on
survival in glioblastoma in a randomised phase III study: 5-year
analysis of the EORTC-NCIC trial. Lancet Oncol. 10:459–466.
2009.PubMed/NCBI
|
|
7
|
Klayman DL: Qinghaosu (artemisinin): An
antimalarial drug from China. Science. 228:1049–1055. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qaderi A, Dadgar N, Mansouri H, Alavi SE,
Esfahani MK and Akbarzadeh A: Modeling and prediction of
cytotoxicity of artemisinin for treatment of the breast cancer by
using artificial neural networks. Springerplus. 2:3402013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim SH, Chun SY and Kim TS:
Interferon-alpha enhances artemisinin-induced differentiation of
HL-60 leukemia cells via a PKC alpha/ERK pathway. Eur J Pharmacol.
587:65–72. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sadava D, Phillips T, Lin C and Kane SE:
Transferrin overcomes drug resistance to artemisinin in human
small-cell lung carcinoma cells. Cancer Lett. 179:151–156. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schmuck G and Haynes RK: Establishment of
an in vitro screening model for neurodegeneration induced by
antimalarial drugs of the artemisinin-type. Neurotox Res. 2:37–49.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ontikatze T, Rudner J, Handrick R, Belka C
and Jendrossek V: Dihydroartemisinin is a hypoxia-active
anti-cancer drug in colorectal carcinoma cells. Front Oncol.
4:1162014.PubMed/NCBI
|
|
13
|
Li YJ, Zhou JH, Du XX, et al:
Dihydroartemisinin accentuates the anti-tumor effects of
photodynamic therapy via inactivation of NF-κB in Eca109 and Ec9706
esophageal cancer cells. Cell Physiol Biochem. 33:1527–1536. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Park J, Lai HC, Singh M, Sasaki T and
Singh NP: Development of a dihydroartemisinin-resistant Molt-4
leukemia cell line. Anticancer Res. 34:2807–2810. 2014.PubMed/NCBI
|
|
15
|
Gu HM, Warhurst DC and Peters W: Uptake of
[3H] dihydroartemisinine by erythrocytes infected with Plasmodium
falciparum in vitro. Trans R Soc Trop Med Hyg. 78:265–270. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xie LH, Li Q, Zhang J and Weina PJ:
Pharmacokinetics, tissue distribution and mass balance of
radiolabeled dihydroartemisinin in male rats. Malar J. 8:1122009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim SJ, Kim MS, Lee JW, et al:
Dihydroartemisinin enhances radiosensitivity of human glioma cells
in vitro. J Cancer Res Clin Oncol. 132:129–135. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang XJ, Li CT, Zhang WP, Lu YB, Fang SH
and Wei EQ: Dihydroartemisinin potentiates the cytotoxic effect of
temozolomide in rat C6 glioma cells. Pharmacology. 82:1–9. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun YC, Wang J, Guo CC, et al: MiR-181b
sensitizes glioma cells to teniposide by targeting MDM2. BMC
Cancer. 14:6112014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Naito S, von Eschenbach AC, Giavazzi R and
Fidler IJ: Growth and metastasis of tumor cells isolated from a
human renal cell carcinoma implanted into different organs of nude
mice. Cancer Res. 46:4109–4115. 1986.PubMed/NCBI
|
|
21
|
Buckner JC: Factors influencing survival
in high-grade gliomas. Semin Oncol. 30(Suppl 19): S10–S14.
2003.
|
|
22
|
Curran WJ Jr, Scott CB, Horton J, et al:
Recursive partitioning analysis of prognostic factors in three
Radiation Therapy Oncology Group malignant glioma trials. J Natl
Cancer Inst. 85:704–710. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
De Angelis LM: Brain tumors. N Engl J Med.
344:114–123. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sai K, Li WY, Chen YS, et al: Triptolide
synergistically enhances temozolomide-induced apoptosis and
potentiates inhibition of NF-κB signaling in glioma initiating
cells. Am J Chin Med. 42:485–503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fu J, Yang QY, Sai K, et al: TGM2
inhibition attenuates ID1 expression in CD44-high glioma-initiating
cells. Neuro Oncol. 15:1353–1365. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Y and Wu YL: An over four millennium
story behind qinghaosu (artemisinin) - a fantastic antimalarial
drug from a traditional Chinese herb. Curr Med Chem. 10:2197–2230.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rasheed SA, Efferth T, Asangani IA and
Allgayer H: First evidence that the antimalarial drug artesunate
inhibits invasion and in vivo metastasis in lung cancer by
targeting essential extracellular proteases. Int J Cancer.
127:1475–1485. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Efferth T, Sauerbrey A, Olbrich A, et al:
Molecular modes of action of artesunate in tumor cell lines. Mol
Pharmacol. 64:382–394. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stockwin LH, Han B, Yu SX, et al:
Artemisinin dimer anticancer activity correlates with
heme-catalyzed reactive oxygen species generation and endoplasmic
reticulum stress induction. Int J Cancer. 125:1266–1275. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Scherz-Shouval R and Elazar Z: ROS,
mitochondria and the regulation of autophagy. Trends Cell Biol.
17:422–427. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Z, Hu W, Zhang JL, Wu XH and Zhou HJ:
Dihydroartemisinin induces autophagy and inhibits the growth of
iron-loaded human myeloid leukemia K562 cells via ROS toxicity.
FEBS Open Bio. 2:103–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Efferth T, Briehl MM and Tome ME: Role of
antioxidant genes for the activity of artesunate against tumor
cells. Int J Oncol. 23:1231–1235. 2003.PubMed/NCBI
|
|
33
|
Kanzawa T, Germano IM, Komata T, Ito H,
Kondo Y and Kondo S: Role of autophagy in temozolomide-induced
cytotoxicity for malignant glioma cells. Cell Death Differ.
11:448–457. 2004. View Article : Google Scholar : PubMed/NCBI
|