Introduction
Oral squamous cell carcinoma (OSCC) is a devastating
disease, which affects humans. Despite the development of surgical
techniques and prosthetic repair, defects in the facial structures
lead to severe aesthetic and functional problems (1–4).
Uncontrollable and persistent loco-regional and distant recurrences
with invasive disposition result in a low overall long-term
survival rate. Thus, the valuable predictive factors of OSCC are
required to prevent the tumorigenesis, progression and recurrence
of the disease (5,6). The present study investigated the
expression levels of cyclooxygenase-2 (COX-2), peroxiredoxin 1
(PRDX1), PRDX6 and nuclear factor-κB (NF-κB), which have previously
been found to be highly expressed in various neoplasms (7).
COX is the rate-limiting enzyme in the conversion of
arachidonic acid to prostaglandins (PGs) of which two isoforms,
COX-1 and COX-2, have been described (8). COX-1 is constitutively expressed in a
number of tissues and generates the PGs necessary for normal
physiological function (7). Although
COX-2 cannot normally be detected, proinflammatory or mitogenic
stimuli cause its rapid induction (9,10). Studies
have indicated that COX-2 is involved in the process of
carcinogenesis via a range of different mechanisms, including
angiogenesis, cell proliferation and the prevention of apoptosis.
Elevated COX-2 expression has been noted in several human
malignancies, including colonic, breast, gastric, bronchial,
esophageal and prostatic carcinomas, and head and neck cancers
(11).
The PRDX antioxidant protein family is found in a
large range of species and is important in the protection of cells
against oxidants. Since 1998, the crystal structures of six PRDXs
in mammals have been published, including four typical 2-Cys PRDXs
(PRDX1, PRDX1I, TryP and AhpC), one atypical 2-Cys PRDX (PRDX V)
and one 1-Cys PRDX (PRDX6) (12).
This endogenous defense system is highly expressed in certain
cancers. The overexpression of PRDX1 has been noted in follicular
thyroid neoplasms, thyroiditis, lung cancer, malignant mesothelioma
and breast cancer (13–16). Furthermore, PRDX2, 3, 5 and 6 levels
were shown to be increased in malignant mesothelioma, while PRDX2
and 3 were overexpressed in breast cancer specimens compared with
normal tissues (14,16). The proliferation-associated gene
(Pag), human PRDX1, was first isolated by the differential
screening of cDNA libraries formed from untransformed and
ras-transformed human mammary epithelial cells. As the
proliferation of these cells stops during the commitment phase of
differentiation, the higher Pag expression levels were correlated
with cell proliferation (17). Also,
PRDX1 takes part in the growth factor and tumor necrosis factor-α
(TNF-α) signaling cascades by regulating the intracellular
concentration of H2O2 (18). These data indicate that the activity
of PRDX1 may be associated with not only proliferation, but also
the formation of reactive oxygen species (ROS), which participate
in carcinogenesis in all stages, including initiation, promotion
and progression (19).
NF-κB is found in an inactive form in the majority
of cells. Active NF-κB complexes are dimers of a variety of
combinations of the Rel family of polypeptides, consisting of p-50
(NF-κB1), p52 (NF-κB2), c-Rel, v-Rel, Rel-A (p-65) and Rel-B. In
resting cells, retention of p-50 and p-65 heterodimers in the
cytoplasm and their binding to the inhibitor κB (IκB) proteins,
blocks the nuclear translocation of NF-κB (18,20,21). IκB
degraded when it is phosphorylated by IκB kinase in response to a
wide range of carcinogens and growth stimuli, including cigarette
smoke, ultraviolet radiation, chemical carcinogens, epidermal
growth factor receptor (EGFR), TNF-α and other cytokines (14,22,23). NF-κB
that has been liberated enters the nucleus and transactivates the
expression of a number of significant oncogenes and proinflammatory
cytokines, including c-myc, cyclin D1, B-cell lymphoma (Bcl)-XL,
Bcl-2, Cox-2, survivin, matrix metalloproteinase-9; vascular
endothelial cell growth factor, and interleukin (IL)-1, 6 and 9,
whose functions are closely associated with abnormal cancer cell
proliferation, survival and invasion (18,20,21). The
constitutive activation of NF-κB occurs in numerous types of
malignancies, including head and neck SCC, and is associated with
the aggressive phenotype of these tumors (18,20,24).
During the last few years, studies on COX-2, PRDX1,
PRDX6 and NF-κB have been performed in OSCC, but these studies have
relied on qualitative immunohistochemistry to assess the levels of
protein. This type of test can be subject to interobserver
variability. In the present study, the expression of COX-2, PRDX1,
PRDX6 and NF-κB in OSCC was quantitatively evaluated using
quantitative polymerase chain reaction (qPCR), and to the best of
our knowledge, this is the first study of its type in OSCC.
Patients and methods
Patients and samples
A total of 50 OSCC tissues from 35 males and 15
females, with a mean age of 61 years (range, 35–82 years), were
obtained during surgery or at biopsy in the Department of Oral and
Maxillofacial Surgery and Otolaryngology, Chungbuk National
University Hospital, (Cheongju, South Korea). The samples were
classified in terms of the patient's age and gender, the primary
site, the T stage of tumor size, the grade of histological cell
differentiation, the presence of a primary or recurrent tumor, and
the administration of prior chemotherapy at the time of obtaining
the samples. T staging and histological grade were assessed
according to the tumor-node-metastasis staging by the American
Joint Committee on Cancer (Table I)
(25). Additionally, 19 normal buccal
mucosa tissues were obtained from healthy non-cancer patients with
consent during minor oral surgical procedures. Each sample was
examined histopathologically, and only those that consisted of
normal squamous epithelial cells were subsequently analyzed.
Samples were snap-frozen in liquid nitrogen immediately after
harvest and stored at −70°C until RNA extraction. This study was
approved by the Institutional Review Board of Chungbuk National
University Hospital.
 | Table I.Clinical and pathological features of
patients with oral squamous cell carcinoma. |
Table I.
Clinical and pathological features of
patients with oral squamous cell carcinoma.
| Parameter | Value |
|---|
| Gender, n |
|
|
Male | 35 |
|
Female | 15 |
| Age, years | 60.57±9.98 |
| Stage, n |
|
| T1 | 9 |
| T2 | 13 |
| T3 | 2 |
| T4 | 26 |
| Differentiation,
n |
|
|
Well | 39 |
|
Moderate | 8 |
|
Poor | 3 |
| Tumor presence,
n |
|
|
Primary | 44 |
|
Recurrence | 6 |
| Chemotherapy,
n |
|
| No
prior chemotherapy | 36 |
| Prior
chemotherapy | 14 |
RNA extraction and reverse
transcription
Total RNA was isolated from the tissues with TRIzol
reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according
to the manufacturer's instructions. cDNA was then prepared from 1
µg total RNA by random priming using a First-Strand cDNA Synthesis
kit (Amersham Biosciences Europe GmbH, Freiburg, Germany) according
to the manufacturer's instructions.
qPCR
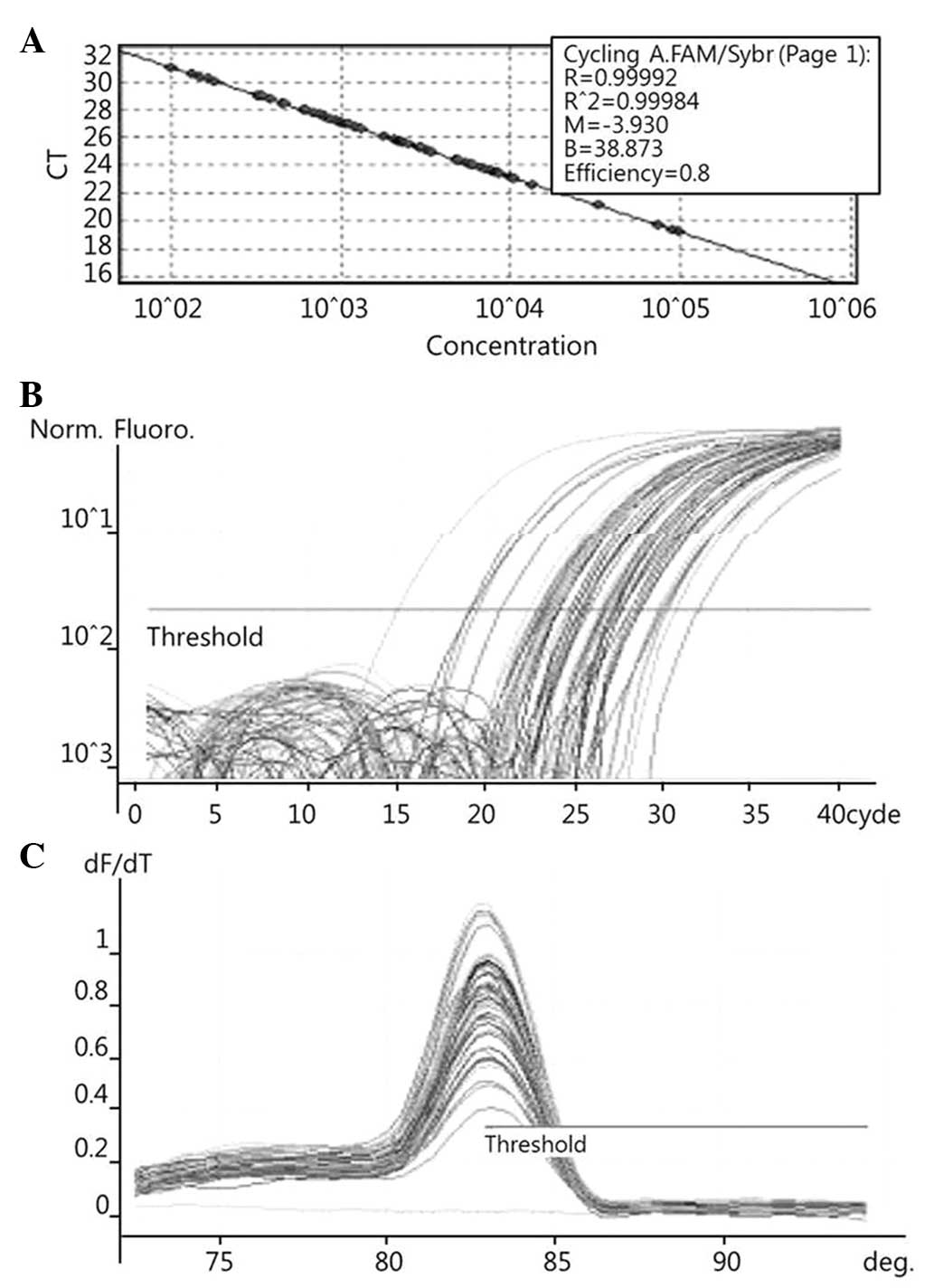
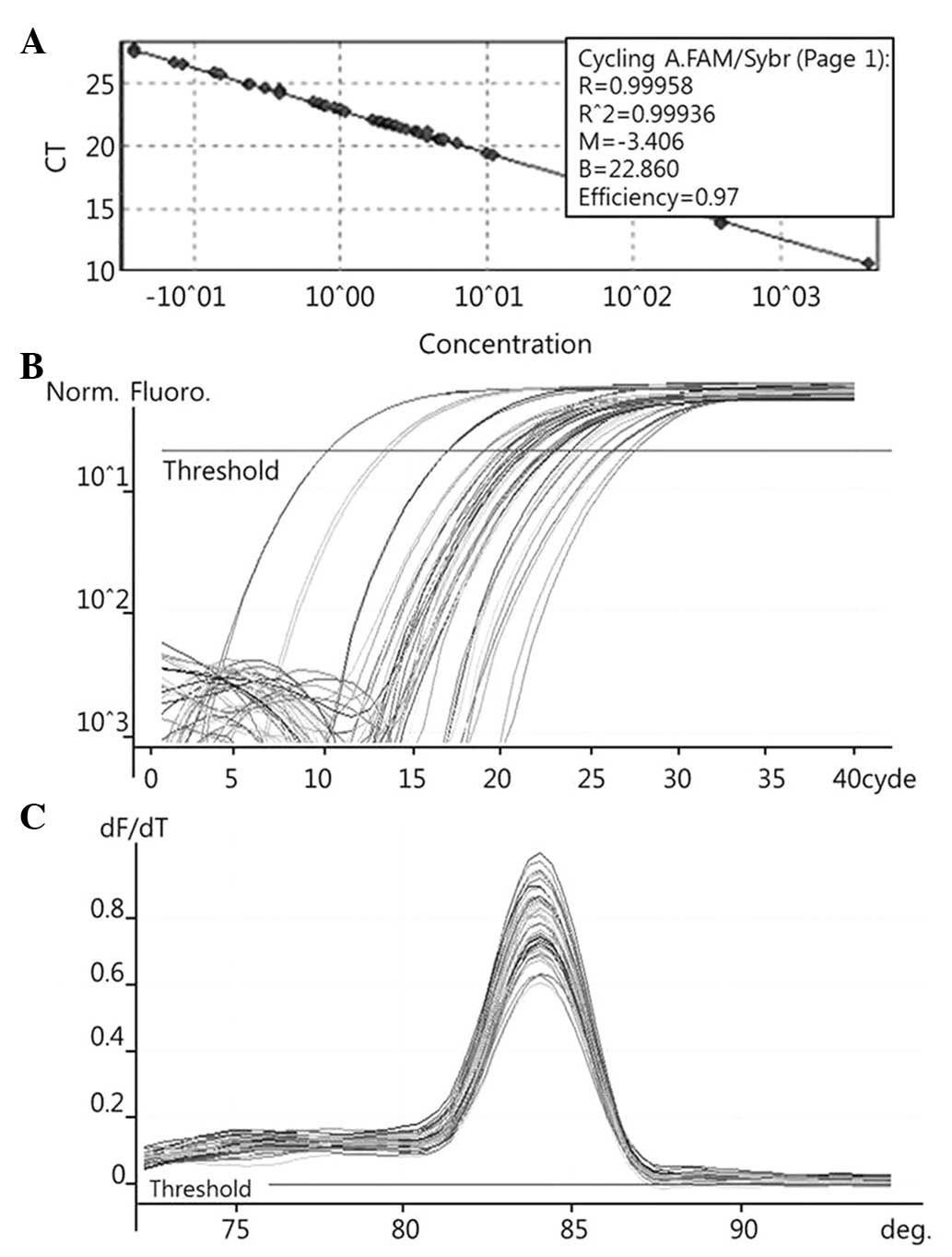
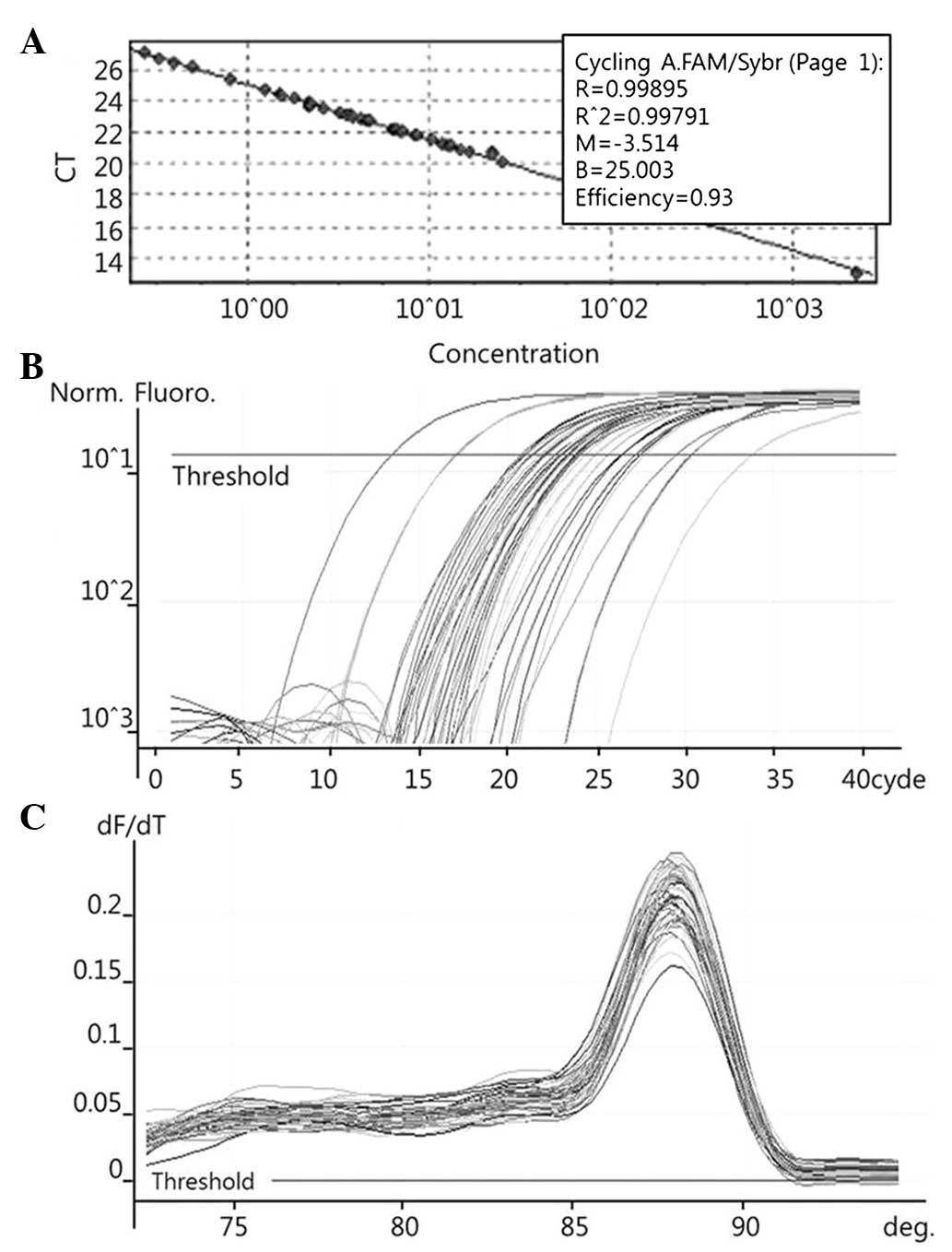
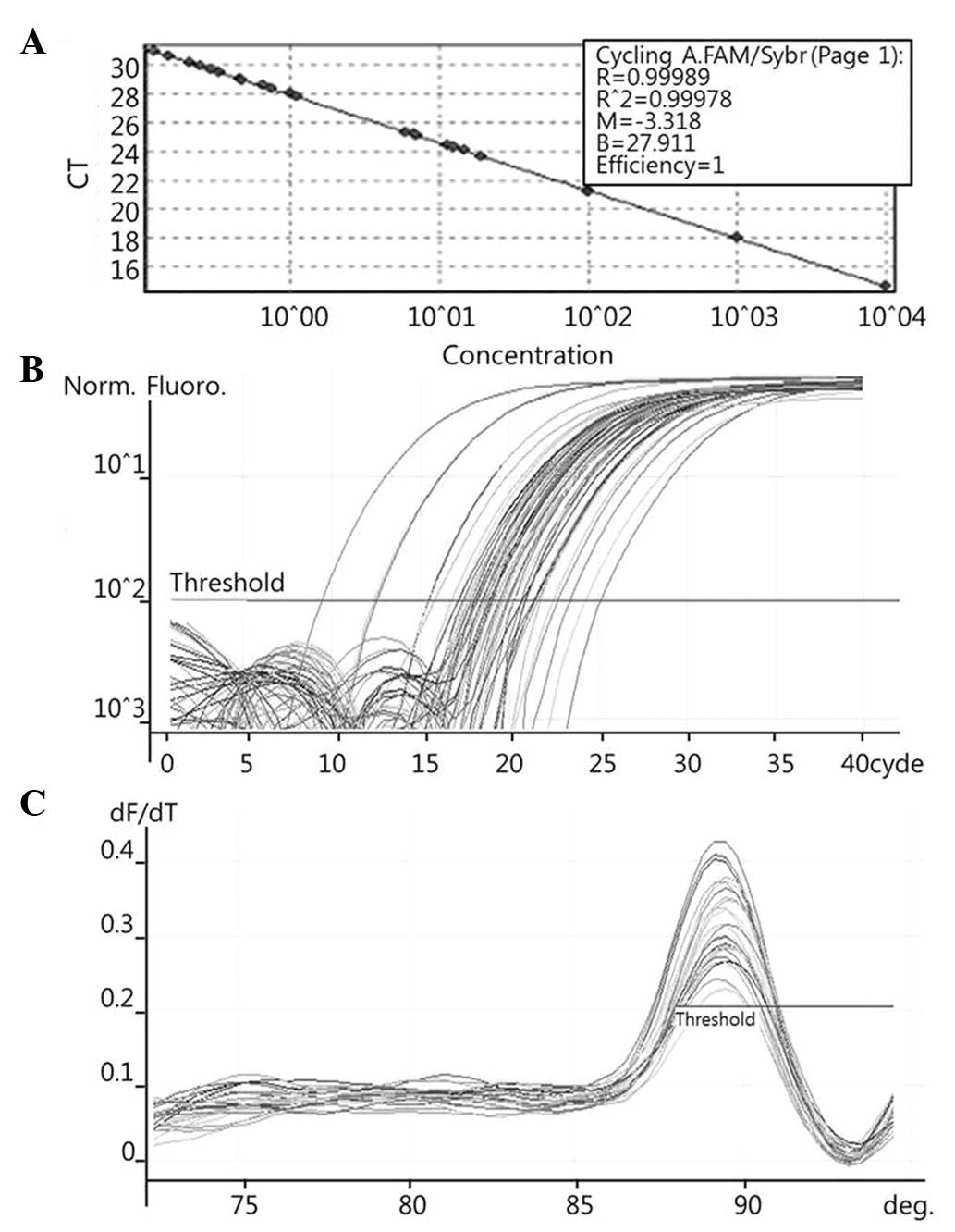
To quantify the expression levels of COX-2 (Fig. 1), PRDX1 (Fig. 2), PRDX6 (Fig. 3) and NF-κB (Fig. 4), qPCR amplification was performed
using the Rotor Gene 3000 (Corbett Research, Mortlake, Australia)
PCR machine. qPCR assays were performed using SYBR Premix EX Taq
(Takara Bio Inc., Otsu, Japan) in a micro reaction tube (Corbett
Research). The PCR was performed with a final volume of 10 µl, the
reaction mixture consisting of 5 µl of 2X SYBR Premix EX Taq
buffer, 0.5 µl of each of 5′ and 3′ primer (10 pmol/µl) and 1 µl of
sample cDNA.
For amplification, the following primers were used:
COX-2, forward, 5′-CCATGGGGTGGACTTAAA-3′ and reverse,
5′-ACAGCAAACCGTAGATGC-3′ (186 bp); PRXDX1, forward,
5′-CCAACTTCAAAGCCACAG-3′ and reverse, 5′-GTCTGATACCAAAGGAATG-3′
(284 bp); PRDX6, forward, 5-CTTTGAGGCCAATACCACCG-3 and reverse,
5-AGATGGTCCTCAACACTGTC-3 (201 bp); and NF-κB, forward,
5′-AAGACCCACCCCACCATCAA-3′ and reverse,
5′-AAACTGTGGATGCAGCAGCGGTC-3′ (173 bp). The products were purified
using a QIAquick Gel Extraction kit (Qiagen, Hilden, Germany),
quantified by spectrophotometer (MBA2000; Perkin Elmer Inc.,
Walther, MA, USA) and the product sequences confirmed by automated
laser fluorescence sequencer (ABI PRISM 3100 Genetic Analyzer;
Applied Biosystems Life Technologies, Foster City, CA, USA).
The known concentrations of the COX-2, PRDX1, PRDX6
and NF-κB PCR products were serially diluted (10-fold) from 1 pg/µl
to 0.1 fg/µl, from 4 pg/µl to 0.4 fg/µl, from 22.5 pg/µl to 2.5
fg/µl, and from 10 pg/µl to 0.1 fg/µl, respectively. The dilution
series of the PCR products were used for establishing standard
curves of the qPCR (Figs. 1A,
2A and 3A). The qPCR conditions were as follows: 1
cycle at 96°C for 1 min, followed by 40 cycles of 96°C for 2 sec,
56°C for 20 sec and 72°C for 20 sec for COX-2; 1 cycle at 95°C for
1 min, followed by 40 cycles of 95°C for 10 sec, 54°C for 15 sec
and 72°C for 20 sec for PRDX1; 1 cycle at 95°C for 1 min, followed
by 40 cycles of 95°C for 10 sec, 62.5°C for 15 sec and 72°C for 20
sec for PRDX6; and 1 cycle at 96°C for 1 min, followed by 40 cycles
of 96°C for 2 sec, 60°C for 20 sec and 72°C for 20 sec for NF-κB.
The melting program was performed at 72–96°C for COX-2 and NF-κB,
and at 72–95°C for PRDX1 and PRDX6, with a heating rate of 1°C
every 45 sec. Spectral data were captured and analyzed using
Rotor-Gene Real-Time Analysis Software 6.0 Build 14 (Corbett
Research) (Figs. 1B and C, 2B and C, and 3B
and C).
Statistical analysis
The COX-2, PRDX1, PRDX6 and NF-κB mRNA expression
levels of the OSCC tissues and normal oral mucosae were compared.
The results were presented as the mean ± standard deviation upon
comparison. Histologically well-differentiated tumors were
classified as low grade, while moderate- and poorly-differentiated
tumors were classified as high grade. The correlations of COX-2,
PRDX1, PRDX6 and NF-κB mRNA expression levels with tumor size were
assessed by Kruskal-Wallis test, and grade, recurrence and the
previous administration of chemotherapy were assessed by the
Mann-Whitney test. All analyses were performed with SPSS for
windows (ver. 12.0.1; SPSS, Inc., Chicago, IL, USA), and P<0.05
was used to indicate a statistically significant difference.
Results
Comparison of COX-2, PRDX1, PRDX6 and
NF-κB mRNA expression levels in OSCC and normal oral mucosa
tissues
The COX-2 mRNA expression levels of 50 OSCC tissues
and 19 normal oral mucosa tissues were 7.753±2.294 and 2.179±0.464
pg/ml, respectively. The COX-2 mRNA expression levels in the OSCC
tissues were significantly higher than that in the normal oral
mucosa tissues (P=0.021) (Table
II).
| Table II.COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression levels in OSCC tissues and normal oral mucosae. |
Table II.
COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression levels in OSCC tissues and normal oral mucosae.
|
| mRNA expression,
pg/ml |
|
|---|
|
|
|
|
|---|
| Genes | OSCC (n=50) | Normal oral mucosae
(n=19) | P-value |
|---|
| COX-2 |
7.753±2.294 |
2.179±0.464 | 0.021 |
| PRDX1 |
10.962±1.641 |
6.420±4.072 | 0.218 |
| PRDX6 |
3.186±0.490 |
3.460±0.595 | 0.760 |
| NF-κB |
10.098±1.602 |
8.979±1.097 | 0.687 |
The PRDX1 mRNA expression levels of the 50 OSCC
tissues and 19 normal oral mucosa tissues were 10.962±1.641 and
6.420±4.072 pg/ml, respectively. The PRDX1 mRNA expression levels
were not significantly different between the OSCC tissues and the
normal oral mucosa tissues (P=0.218) (Table II).
The PRDX6 mRNA expression levels of the 50 OSCC
tissues and 19 normal oral mucosa tissues were 3.186±0.490 and
3.460±0.595 pg/ml, respectively. The PRDX6 mRNA expression levels
were not significantly different between the OSCC tissues and the
normal oral mucosa tissues (P=0.760) (Table II).
The NF-κB mRNA expression levels of the 50 OSCC
tissues and 19 normal oral mucosa tissues were 10.098±1.602 and
8.979±1.097 pg/ml, respectively. The NF-κB mRNA expression levels
were not significantly different between the OSCC tissues and the
normal oral mucosa tissues (P=0.687) (Table II).
COX-2, PRDX1, PRDX6 and NF-κB mRNA expression levels
of OSCC tissues according to the tumor stage. The COX-2, PRDX6 and
NF-κB mRNA expression levels of the OSCC tissues did not show a
significant correlation with tumor size (P=0.147, P=0.288 and
P=0.147, respectively; Table III).
However, the PRDX1 mRNA expression levels of the 28 high-stage (T3
and T4) and 9 low-stage (T1) OSCC tissues were 13.981±2.509 and
5.234±3.078 pg/ml, respectively. The PRDX1 mRNA expression levels
in the high-stage OSCC tissues were significantly higher than those
in the low-stage OSCC tissues (P=0.047) (Table III).
| Table III.COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression level of OSCC tissues according to stage. |
Table III.
COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression level of OSCC tissues according to stage.
|
| OSCC mRNA
expression, pg/ml |
|
|---|
|
|
|
|
|---|
| Genes | Low-stage tumor
(T1) (n=9) | High-stage tumor
(T3 and T4) (n=28) | P-value |
|---|
| COX-2 |
5.240±1.586 |
12.511±2.778 | 0.147 |
| PRDXI |
5.234±3.078 |
13.981±2.509 | 0.047 |
| PRDX6 |
2.290±0.771 |
3.872±0.752 | 0.288 |
| NF-κB |
2.373±0.965 |
10.970±4.115 | 0.147 |
Comparison of COX-2, PRDX1, PRDX6 and
NF-κB mRNA expression levels of OSCC tissues according to the
degree of histological tumor cell differentiation
The COX-2, PRDX1 and PRDX6 mRNA expression levels of
the OSCC tissues did not show a significant correlation with tumor
differentiation (P=0.788, P=0.171 and P=0.343, respectively;
Table IV). However, the NF-κB mRNA
expression levels of 11 high-grade (moderate- and
poorly-differentiated) OSCC tissues and 39 low-grade
(well-differentiated) OSCC tissues were 17.218±5.228 and
8.488±1.490 pg/ml, respectively. The NF-κB mRNA expression levels
in the high-grade OSCC tissues were significantly higher than those
in the low-grade OSCC tissues (P=0.048) (Table IV).
| Table IV.COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression level of OSCC tissues according to differentiation. |
Table IV.
COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression level of OSCC tissues according to differentiation.
|
| OSCC mRNA
expression, pg/ml |
|
|---|
|
|
|
|
|---|
| Genes | Low-grade tumor
(G1) (n=39) | High-grade tumor
(G2 and G3) (n=11) | P-value |
|---|
| COX-2 |
7.245±2.482 |
10.469±6.887 | 0.788 |
| PRDXI |
9.848±1.907 |
15.368±3.591 | 0.171 |
| PRDX6 |
3.036±0.532 |
4.378±1.371 | 0.343 |
| NF-κB |
8.488±1.490 |
17.218±5.228 | 0.048 |
Comparison of COX-2, PRDX1, PRDX6 and
NF-κB mRNA expression levels of OSCC tissues according to tumor
recurrence
The COX-2, PRDX1, PRDX6 and NF-κB mRNA expression
levels of 44 primary OSCC tissues and 6 recurrent OSCC tissues did
not show a correlation with tumor recurrence (P=0.439, P=0.850,
P=0.590 and P=0.189, respectively, Table
V). COX-2 level tended to be higher in the recurrent tumors,
but this difference was not statistically significant.
| Table V.COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression levels of primary and recurrent tumors. |
Table V.
COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression levels of primary and recurrent tumors.
|
| OSCC mRNA
expression, pg/ml |
|
|---|
|
|
|
|
|---|
| Genes | Primary tumor
(n=44) | Recurrent tumor
(n=6) | P-value |
|---|
| COX-2 |
5.547±1.824 |
25.609±14.165 | 0.439 |
| PRDXI |
10.908±1.813 |
12.196±5.306 | 0.850 |
| PRDX6 |
3.449±0.563 |
2.468±1.122 | 0.590 |
| NF-κB |
11.042±1.859 |
5.763±2.990 | 0.189 |
Comparison of COX-2, PRDX1, PRDX6 and
NF-κB mRNA expression levels of OSCC tissues according to the prior
administration of chemotherapy
The COX-2, PRDX1, PRDX6 and NF-κB mRNA expression
levels of 14 OSCC tissues previously administered chemotherapy and
36 OSCC tissues not administered chemotherapy were examined. There
was no correlation with the administration of previous chemotherapy
in each group (P=0.105, P=0.218, P=0.364 and P=0.290, respectively;
Table 6).
| Table 6.COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression levels of OSCC tissues following chemotherapy. |
Table 6.
COX-2, PRDXI, PRDX6 and NF-κB mRNA
expression levels of OSCC tissues following chemotherapy.
|
| OSCC mRNA
expression, pg/ml |
|
|---|
|
|
|
|
|---|
| Genes | Prior chemotherapy
(n=14) | No prior
chemotherapy (n=36) | P-value |
|---|
| COX-2 |
11.437±6.312 |
6.600±2.349 | 0.105 |
| PRDXI |
13.618±3.038 |
10.068±2.044 | 0.218 |
| PRDX6 |
3.596±0.834 |
3.228±0.638 | 0.364 |
| NF-κB |
10.944±2.258 |
10.200±2.185 | 0.290 |
Discussion
The identification of a biological marker that
correlates with a specific tumor would provide more accurate
information and enable molecular-targeted therapy. There are a
number of potential molecular markers in OSCC that have drawn
considerable interest.
Selective COX-2 overexpression has been found in
head and neck, bronchial, breast, pancreatic, colon and gastric
cancers. Particularly, a higher expression level of COX-2 has been
identified in head and neck OSCC when compared with other cancers
(11,26–29). Tumor
specimens with confirmed cervical lymph node metastasis have a
significantly higher level of COX-2 expression, as do advanced and
poorly-differentiated head and neck SCCs. Peng et al noted
the overexpression of COX-2 in N1-N3 tumors compared with N0 tumors
in a series of 23 hypopharyngeal SCC cases (28). Shibata et al noted labeling
indices of COX-2 that correlated with the histological grade of
dysplasia, being highest for the severe dysplasias on four human
OSCC cell lines (11). Another study
also showed that COX-2 expression is involved in the regulation of
cell proliferation and in the progression from normal tissue to SCC
(30). One previous study by Tsujii
and DuBois examined the association between apoptosis and COX-2,
and found that COX-2 gene transfection into rat intestinal
epithelial cells decreases the susceptibility of the cells to
apoptosis and upregulates antiapoptotic Bcl-2 protein expression
(31). Although the present study
showed a good correlation between COX-2 mRNA expression and OSCC
(P=0.021), COX-2 mRNA expression was not significantly correlated
with other factors, such as tumor size, differentiation, recurrence
and previous chemotherapy. COX-2 tended to be expressed at a higher
level in recurrent tumors, although without a significant
correlation (primary tumor (n=44), 5.547±1.824 pg/ml; recurrent
tumor (n=6), 25.609±14.165 pg/ml; P=0.439); this may be as the
recurrent tumor group was small in number. In addition, it has been
demonstrated that COX-2 overexpression has a critical period. Zhi
et al noted that strong COX-2 expression was detected as
early as the stage of dysplasia and invasive SCC, while being
weakly expressed in the esophageal squamous epithelium (32). Kase et al demonstrated a peak
of expression in high-grade dysplasia and carcinoma in situ,
then a stable level with a declining trend (33). This may indicate that COX-2
facilitates tumorigenesis and progression in the early stage,
independent of its expression level during progression.
Recently, a number of studies reported the increased
expression of PRDX in several cancers, and since then, PRDX has
received considerable attention as a novel marker. The studies
using a PRDX1-knockout mouse indicated that PRDX1 in normal cells
plays a direct role in tumor suppression by eliminating ROS and
preventing oxidative damage, and PRDX1 expression was associated
with the occurrence of malignant neoplasms (24). Similarly, consumedly proliferative
tumor cells may express PRDX to protect themselves from oxidative
damage. In malignant mesothelioma, PRDX1 and 6 are overexpressed,
and tumor tissues with strong PRDX1 expression tended to experience
an increased survival time (14). In
primary bladder cancer, the PRDX1 and 6 expression levels were
significantly elevated in non-progressed cases compared to the
progressed cases (34). The
overexpression of PRDX was also reported in human breast and lung
cancer (16,35). In OSCC, Yanagawa et al studied
PRDX1 expression for the first time and found low PRDX1expression
levels to be associated with larger tumor masses, lymph node
metastases and poorly-differentiated tumors (36). This data is contrary to previous
studies and the present result showing that the PRDX1 mRNA
expression was elevated in larger tumors (P=0.047). Wen and Van
Etten revealed that the paradoxical mechanism may be explained by
the fact that Pag is a physiological inhibitor of c-Abl (37). c-Abl is an oncogene product with
tyrosine kinase activity, which inhibits cell proliferation when
activated by interacting with central elements controlling the cell
cycle (38).
In a subsequent study, Yanagawa et al
reported that PRDX1 expression was significantly associated with
local and lymph node recurrence (39), which was in contrast to the present
results. These results indicated that PRDX1 expression predicts a
2.8- to 2.9-fold more frequent incidence of recurrence. PRDX1 was
originally isolated from proliferating cells and the overexpression
of PRDX1 has been found in the proliferative type of tumors.
Additionally, an elevated level of PRDX1 appears to abrogate the
induced cell-cycle block and is able to act as a tumor suppressor
through the reduction of ROS. Taken together, these data indicate
that the PRDX expression level appears to vary depending on the
type or stage of the cancer. It is also reasonable to suggest that
the augmented expression of PRDX provides increased antioxidative
activity, which may be responsible for its resistance to apoptosis
as a tumor suppressor. An increased PRDX level in larger tumors
would be a result of the antiapoptotic features of PRDX, providing
a growth advantage to tumor cells.
NF-κB and NF-κB inducible gene activation has been
detected in melanomas, prostate and breast cancer, and lymphoma
(40,41). In the head and neck, Takada et
al showed enhanced IκB kinase (IKK) activity in carcinoma cells
(42). Wolf et al revealed
that IL-1 may function as an autocrine growth factor that is able
to induce constitutive NF-κB activation (43), while Bancroft et al
demonstrated that EGFR activation could result in NF-κB activation
(44). Cumulative evidence in
malignancies is compatible with the hypothesis that NF-κB is
constitutively activated and has a major function in the
pathogenesis of cellular and host alterations. NF-κB activation has
been suggested to be a common pathway of widespread importance, and
it has been suggested to be involved in promoting the expression of
the phenotypic changes and genes involved in inhibition of
programmed and therapeutic cell death, proliferation,
tumorigenesis, angiogenesis, invasiveness and metastatic potential
(41,45). Nakayama et al noted that high
expression levels of NF-κB (p65) and IKK contribute to malignant
behavior and antiapoptotic activity in OSCC (45). Furthermore, molecular profiling has
revealed that NF-κB may regulate a diverse repertoire of genes in
SCC. Loercher et al obtained evidence with regard to the
differential expression of known NF-κB-associated genes and found
that NF-κB pathway modulation directly or indirectly alters a
significant portion of these and other genes in the changed
molecular profile, with malignant progression in a syngeneic model
of SCC (41). This study confirmed
that IκB-α phosphorylation mutant expression suppressed NF-κB
reporter and DNA binding activity, and modulated the protein and
mRNA expression of a number of putative NF-κB target genes that are
differentially-expressed in SCC, including Trp53, cyclin D1, IAP-1
and β-catenin. Consistent with the diversity and putative functions
of a number of these genes, NF-κB inhibition was noted to inhibit
the proliferation, survival, migration, angiogenesis and
tumorigenesis of cells. These results indicate that NF-κB is a
significant modulator of the gene expression profile and malignant
phenotype in SCC. In the present study, NF-κB mRNA expression was
correlated with the histopathological grade. The level was
significantly elevated in the high-grade OSCC tissues (P=0.048),
which agrees with the results previously published stating that the
activation of NF-κB is associated with an aggressive tumor
phenotype.
Generally, SCCs exhibit a range of clinical and
histological features according to their stage of epithelial
differentiation. Poorly-differentiated SCCs are known to grow at a
faster pace and spread diffusely, leading to a poor prognosis.
Furthermore, well-differentiated SCCs are chemotherapy- and
radiotherapy-resistant. This suggests that NF-κB is a key molecular
switch of the alterations in genotype and phenotype in the
malignant progression of OSCC.
In conclusion, the expression of COX-2 is strongly
associated with the development of OSCC. Moreover, the enhanced
expression of PRDX1 and NF-κB may function in the progression of
OSCC, which serves as a useful marker for the prognosis of patients
with this disease.
References
|
1
|
Bianchi B, Ferri A, Ferrari S, et al:
Reconstruction of lateral through and through oro-mandibular
defects following oncological resections. Microsurgery. 30:517–525.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thankappan K, Kuriakose MA, Chatni SS,
Sharan R, Trivedi NP, Vijayaraghavan S, Sharma M and Iyer S:
Lateral arm free flap for oral tongue reconstruction: An analysis
of surgical details, morbidity, and functional and aesthetic
outcome. Ann Plast Surg. 66:261–266. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zeno HA, Sternberger SS, Tuminelli FJ,
Billotte M and Kurtz KS: Combination lower lip prosthesis retained
by an intraoral component. J Prosthodont. 22:397–401. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kawase-Koga Y, Mori Y, Saijo H, Hoshi K
and Takato T: Reconstruction of a complex midface defect from
excision of a squamous cell carcinoma, according to regional
aesthetic units. Oral Surg Oral Med Oral Pathol Oral Radiol.
117:e97–e101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chiesa F, Mauri S, Tradati N, Calabrese L,
Giugliano G, Ansarin M, Andrle J, Zurrida S, Orecchia R and Scully
C: Surfing prognostic factors in head and neck cancer at the
millennium. Oral Oncol. 35:590–596. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Brinkman BM and Wong DT: Disease mechanism
and biomarkers of oral squamous cell carcinoma. Curr Opin Oncol.
18:228–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Babu GS, Supriya AN, Kumar Raj NG and
Swetha P: Tumor markers: An overview. J Orofac Sci. 3:87–95.
2012.
|
|
8
|
Sudbø J and Reith A: The evolution of
predictive oncology and molecular-based therapy for oral cancer
prevention. Int J Cancer. 115:339–345. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kujubu DA, Reddy ST, Fletcher BS and
Herschman HR: Expression of the protein product of the
prostaglandin synthase-2/TIS10 gene in mitogen-stimulated Swiss 3T3
cells. J Biol Chem. 268:5425–5430. 1993.PubMed/NCBI
|
|
10
|
Vane J: Towards a better aspirin. Nature.
367:215–216. 1994. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shibata M, Kodani I, Osaki M, Araki K,
Adachi H, Ryoke K and Ito H: Cyclo-oxygenase-1 and-2 expression in
human oral mucosa, dysplasias and squamous cell carcinomas and
their pathological significance. Oral Oncol. 41:304–312. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wood ZA, Schröder E, Robin Harris J and
Poole LB: Structure, mechanism and regulation of peroxiredoxins.
Trends Biochem Sci. 28:32–40. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yanagawa T, Ishikawa T, Ishii T, Tabuchi
K, Iwasa S, Bannai S, Omura K, Suzuki H and Yoshida H:
Peroxiredoxin I expression in human thyroid tumors. Cancer Lett.
145:127–132. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kinnula VL, Lehtonen S, Sormunen R,
Kaarteenahob WR, Kang SW, Rhee SG and Soini Y: Overexpression of
peroxiredoxins III, IIIV and VI in malignant mesothelioma. J
Pathol. 196:316–323. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang JW, Jeon HB, Lee JH, Yoo JS, Chun
JS, Kim JH and Yoo YJ: Augmented expression of peroxiredoxin I in
lung cancer. Biochem Biophys Res Commun. 289:507–512. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Noh DY, Ahn SJ, Lee RA, Kim SW, Park IA
and Chae HZ: Overexpression of peroxiredoxin in human breast
cancer. Anticancer Res. 21:2085–2090. 2001.PubMed/NCBI
|
|
17
|
Prosperi MT, Ferbus D, Karczinski I and
Goubin G: A human cDNA corresponding to a gene overexpressed during
cell proliferation encodes a product sharing homology with amoebic
and bacterial proteins. J Biol Chem. 268:11050–11056.
1993.PubMed/NCBI
|
|
18
|
Kang SW, Chae HZ, Seo MS, Kim K, Baines IC
and Rhee SG: Mammalian peroxiredoxin isoforms can reduce hydrogen
peroxide generated in response to growth factors and tumor necrosis
factor-alpha. J Biol Chem. 273:6297–6302. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Klaunig JE, Xu Y, Isenberg JS, Bachowski
S, Kolaja KL, Jiang J, Stevenson DE and Walborg EF Jr: The role of
oxidative stress in chemical carcinogenesis. Environ Health
Perspect. 106:289–295. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fujii J and Ikeda Y: Advances in our
understanding of peroxiredoxin, a multifunctional, mammalian redox
protein. Redox Rep. 7:123–130. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ishii T, Yamada M, Sato H, Matsue M,
Taketani S, Nakayama K, Sugita Y and Bannai S: Cloning and
characterization of a 23-kDa stress-induced mouse peritoneal
macrophage protein. J Biol Chem. 268:18633–18636. 1993.PubMed/NCBI
|
|
22
|
Liu M, Lawson G, Delos M, Jamart J, Ide C,
Coche E, Weynand B, Desuter G, Hamoir M, Remacle M and Marbaix E:
Predictive value of the fraction of cancer cells immunolabeled for
proliferating cell nuclear antigen or Ki67 in biopsies of head and
neck carcinomas to identify lymph node metastasis: Comparison with
clinical and radiologic examinations. Head Neck. 25:280–288. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Matsumoto A, Okado A, Fujii T, Fujii J,
Egashira M, Niikawa N and Taniguchi N: Cloning of the peroxiredoxin
gene family in rats and characterization of the fourth member. FEBS
Lett. 443:246–250. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Neumann CA, Krause DS, Carman CV, Das S,
Dubey DP, Abraham JL, Bronson RT, Fujiwara Y, Orkin SH and Van
Etten RA: Essential role for the peroxiredoxin Prdx1 in erythrocyte
antioxidant defence and tumour suppression. Nature. 424:561–565.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Greene FL, Page D, Morrow M, et al: Lip
and oral cavity. AJCC Cancer Staging Manual (6th). (New York).
Springer. 23–32. 2002.
|
|
26
|
Wolff H, Saukkonen K, Anttila S,
Karjalainen A, Vainio H and Ristimäki A: Expression of
cyclooxygenase-2 in human lung carcinoma. Cancer Res. 58:4997–5001.
1998.PubMed/NCBI
|
|
27
|
Shamma A, Yamamoto H, Doki Y, Okami J,
Kondo M, Fujiwara Y, Yano M, Inoue M, Matsuura N, Shiozaki H and
Monden M: Up-regulation of cyclooxygenase-2 in squamous
carcinogenesis of the esophagus. Clin Cancer Res. 6:1229–1238.
2000.PubMed/NCBI
|
|
28
|
Peng JP, Su CY, Chang HC, Chai CY and Hung
WC: Overexpression of cyclo-oxygenase 2 in squamous cell carcinoma
of the hypopharynx. Hum Pathol. 33:100–104. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Banerjee AG, Gopalakrishnan VK,
Bhattacharya I and Vishwanatha JK: Deregulated cyclooxygenase-2
expression in oral premalignant tissues. Mol Cancer Ther.
1:1265–1271. 2002.PubMed/NCBI
|
|
30
|
Yu HP, Xu SQ, Liu L, Shi LY, Cai XK, Lu
WH, Lu B, Su YH and Li YY: Cyclo-oxygenase-2 expression in squamous
dysplasia and squamous cell carcinoma of the esophagus. Cancer
lett. 198:193–201. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsujii M and DuBois RN: Alterations in
cellular adhesion and apoptosis in epithelial cells overexpressing
prostaglandin endoperoxide synthase 2. Cell. 83:493–501. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhi H, Wang L, Zhang J, Zhou C, Ding F,
Luo A, Wu M, Zhan Q and Liu Z: Significance of COX-2 expression in
human esophageal squamous cell carcinoma. Carcinogenesis.
27:1214–1221. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kase S, Osaki M, Honjo S, Adachi H,
Tsujitani S, Kaibara N and Ito H: Expression of cyclo-oxygenase-2
is correlated with high intratumoral microvessel density and low
apoptotic index in human esophageal squamous cell carcinomas.
Virchows Arch. 442:129–135. 2003.PubMed/NCBI
|
|
34
|
Quan C, Cha EJ, Lee HL, Han KH, Lee KM and
Kim WJ: Enhanced expression of peroxiredoxin I and 6 correlates
with development, recurrence and progression of human bladder
cancer. J Urol. 175:1512–1516. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lehtonen ST, Svensk AM, Soini Y, Pääkkö P,
Hirvikoski P, Kang SW, Säily M and Kinnula VL: Peroxiredoxins, a
novel protein family in lung cancer. Int J Cancer. 111:514–521.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yanagawa T, Iwasa S, Ishii T, Tabuchi K,
Yusa H, Onizawa K, Omura K, Harada H, Suzuki H and Yoshida H:
Peroxiredoxin I expression in oral cancer: A potential new tumor
marker. Cancer Lett. 156:27–35. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wen ST and Van Etten RA: The pag gene
product, a stress-induced protein with antioxidant properties, is
an abl SH3-binding protein and a physiological inhibitor of c-Abl
tyrosine kinase activity. Genes Dev. 11:2456–2467. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Prospéri MT, Ferbus D, Rouillard D and
Goubin G: The pag gene product, a physiological inhibitor of c-abl
tyrosine kinase, is overexpressed in cells entering S phase and by
contact with agents inducing oxidative stress. FEBS Lett.
423:39–44. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yanagawa T, Omura K, Harada H, Ishii T,
Uwayama J, Nakaso K, Iwasa S, Koyama Y, Onizawa K, Yusa H and
Yoshida H: Peroxiredoxin I expression in tongue squamous cell
carcinomas as involved in tumor recurrence. Int J Oral Maxillofac
Surg. 34:915–920. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Richmond A: NF-kappa B chemokine gene
transcription and tumour growth. Nat Rev Immunol. 2:664–674. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Loercher A, Lee TL, Ricker JL, Howard A,
Geoghegen J, Chen Z, Sunwoo JB, Sitcheran R, Chuang EY, Mitchell
JB, et al: Nuclear factor-kappaB is an important modulator of the
altered gene expression profile and malignant phenotype in squamous
cell carcinoma. Cancer Res. 64:6511–6523. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Takada Y, Singh S and Aggarwal BB:
Identification of a p65 peptide that selectively inhibits NF-kappa
B activation induced by various inflammatory stimuli and its role
in down-regulation of NF-kappaB-mediated gene expression and
up-regulation of apoptosis. J Biol Chem. 279:15096–15104. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wolf JS, Chen Z, Dong G, Sunwoo JB,
Bancroft CC, Capo DE, Yeh NT, Mukaida N and Van Waes C: IL
(interleukin)-1alpha promotes nuclear factor-kappaB and
AP-1-induced IL-8 expression, cell survival and proliferation in
head and neck squamous cell carcinomas. Clin Cancer Res.
7:1812–1820. 2001.PubMed/NCBI
|
|
44
|
Bancroft CC, Chen Z, Yeh J, Sunwoo JB, Yeh
NT, Jackson S, Jackson C and Van Waes C: Effects of pharmacologic
antagonists of epidermal growth factor receptor, PI3K and MEK
signal kinases on NF-kappaB and AP-1 activation and IL-8 and VEGF
expression in human head and neck squamous cell carcinoma lines.
Int J Cancer. 99:538–548. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nakayama H, Ikebe T, Beppu M and Shirasuna
K: High expression levels of nuclear factor kappaB IkappaB kinase
alpha and Akt kinase in squamous cell carcinoma of the oral cavity.
Cancer. 92:3037–3044. 2001. View Article : Google Scholar : PubMed/NCBI
|