Introduction
Ovarian cancer ranks fifth overall for
cancer-associated mortalities in women, with lung, breast,
colorectal and pancreatic cancer ranking 1–4, respectively, and
accounts for 5% of all cancer-associated mortalities in women
(1). The overall five-year survival
rate is ~44%, and only 27% for patients that are at the distant
stage of the disease at diagnosis (1). The exact cause of epithelial ovarian
cancer has not yet been determined, although follicle-stimulating
hormone (FSH) and luteinizing hormone (LH) are understood to be
associated with ovarian malignancy (2). With apoptosis and apoptosis-linked genes
demonstrating key roles in ovarian cancer tumorigenesis (3,4), it is
important to note that FSH has been previously reported to inhibit
apoptosis in ovarian cancer cells (5). However, little is currently understood
regarding the association between LH and apoptosis in ovarian
cancer, with further investigation required.
Programmed cell death gene 6 (PDCD6), also known as
apoptosis-linked gene-2, is an apoptotic mediator that is required
for apoptosis to occur in response to numerous death stimuli
(6). It has recently been suggested
that PDCD6 is an independent predictor of progression-free survival
in patients with epithelial ovarian cancer (7). However, its effect on ovarian cancer
apoptosis is currently unknown.
To understand the function and regulation of LH on
ovarian cancer apoptosis, the present study investigated the effect
of LH on apoptosis in vitro using flow cytometry. The
protein and mRNA expression levels of PDCD6 were analyzed using
western blotting and quantitative polymerase chain reaction (qPCR).
The signal transduction pathways were also examined using western
blotting.
Materials and methods
Reagents and antibodies
LH, SP600125 [a stress-activated protein
kinase/c-Jun NH2-terminal kinase (SAPK/JNK) inhibitor],
SB203580 [a p38 mitogen-activated protein kinase (MAPK) inhibitor],
LY294002 [a phosphatidylinositol 3-kinase (PI3K) inhibitor] and
U0126 [a p44/42 MAPK (extracellular signal-regulated kinase 1/2;
Erk1/2) inhibitor] were acquired from Sigma-Aldrich (St. Louis, MO,
USA). The PDCD6 rabbit anti-human polyclonal antibody (cat. no.
12303-1-AP; dilution, 1:1,000) was purchased from Proteintech
Group, Inc. (Chicago, IL, USA) for western blotting. The antibodies
against phospho-protein kinase B (pAKT; rabbit anti-human
monoclonal antibody; cat. no. 4060; dilution, 1:1,000), protein
kinase B (AKT; rabbit anti-human monoclonal antibody; cat. no.
9272; dilution, 1:1,000), phospho-c-Jun (pJUN; rabbit anti-human
monoclonal antibody; cat. no. 3270; dilution, 1:1,000), c-Jun
(rabbit anti-human monoclonal antibody; cat. no. 9165; dilution,
1:1,000), phospho-p38 MAPK (rabbit anti-human monoclonal antibody;
cat. no. 4511; dilution, 1:1,000), p38 MAPK (rabbit anti-human
monoclonal antibody; cat. no. 9212; dilution, 1:1,000),
phospho-p44/42 MAPK (pErk1/2; rabbit anti-human monoclonal
antibody; cat. no. 4370; dilution, 1:1,000), and extracellular
signal-regulated protein kinase 1 and 2 (Erk1/2; rabbit anti-human
monoclonal antibody; cat. no. 9102; dilution, 1:1,000) were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
The glyceraldehyde 3-phosphate dehydrogenase (GAPDH) antibody
(mouse anti-human monoclonal antibody; cat. no. KC-5G5; dilution,
1:10,000) was purchased from Kangchen Bioengineering Corporation
(Shanghai, China).
Cell culture
The ovarian cancer cell lines, OVCAR-3 and SKOV-3,
were purchased from the American Type Culture Collection (Manassas,
VA, USA) and cultured according to the company's protocols. SKOV-3
cells were cultured at 37°C with 5% CO2 in McCoy's 5a
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 0.1% gentamicin sulfate (Gemini Bio Products,
West Sacramento, CA, USA), while OVCAR-3 cells were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS and 0.1% gentamicin sulfate. All
experiments were performed with cell lines at 60% to 80%
confluence. To investigate the expression of PDCD6, the cells were
treated for up to 24 h with LH (40 U/l) in the absence of FBS. To
investigate signal transduction, the cells were treated for up to
120 min with LH (40 U/l) in the absence of FBS. To evaluate the
effects of the specific inhibitors, the cells were pre-treated with
each inhibitor for 30 min, and LH was then added for an additional
30 min or 24 h without FBS.
Cell apoptosis analysis
Following a 48-h incubation with LH, with or without
cisplatin (10 µM), the number of apoptotic cells was determined
using the Annexin V-FITC Apoptosis Detection kit (BD Pharmingen,
San Diego, CA, USA) followed by flow cytometry. The apoptotic cells
were determined using a FACScan cytofluorometer from BD Biosciences
(Franklin Lakes, NJ, USA) with Cell Quest software version 5.1,
also from BD Biosciences. The early apoptotic [Annexin-V positive,
propidium iodide (PI) negative] and late apoptotic (Annexin-V
positive, PI positive) cells were included in cell death
determinations.
qPCR
Total RNA was prepared using the RNAprep Pure Cell
kit (Tiangen Biotech Co., Ltd., Beijing, China) according to the
manufacturer's protocols. The primers utilized for the SYBR Green
(Tiangen Biotech Co., Ltd.) qPCR were as follows: PDCD6,
5′-GGATGATCGATAAGAACGAGCTGAA-3′ (forward) and
5′-ATGAGGATGTCGTGGAACTGGTC-3′ (reverse); and GAPDH,
5′-ATGGAAATCCCATCACCATCTT-3′ (forward) and 5′-CGCCCCACTTGATTTTGG-3′
(reverse). The reaction mixture was composed of 12.5 µl One Step
SYBR RT-PCR Buffer III, 0.5 µl Takara Ex Taq HS DNA Polymerase (5
U/µl), 0.5 µl PrimeScript RT Enzyme Mix II, 0.5 µl Forward PCR
Primer (10 µM), 0.5 µl Reverse PCR Primer (10 µM), 2 µl total RNA
(100 ng) and 8.5 µl RNase Free dH2O (Thermo Fisher
Scientific, Inc.), all obtained from Takara Bio, Inc. (Otsu,
Japan). The qPCR conditions in an Applied Biosystems 7500 series
qPCR system (Thermo Fisher Scientific, Inc.) were as follows: 42°C
for 5 min, followed by 95°C for 10 sec, then 40 cycles of 95°C for
5 sec and 60°C for 30 sec. RNase-Free dH2O without RNA
was set as a negative control. The relative mRNA expression levels
were calculated and normalized using the qPCR and the
2−ΔΔcq method.
Western blotting
Following cell lysis, 30 µg total protein was
separated on 8–12% sodium dodecyl sulfate-polyacrylamide gels,
transferred to nitrocellulose membranes (GE Healthcare Life
Sciences, Uppsala, Sweden) and immunoblotted with specific primary
antibodies (PDCD6, GAPDH, pAKT, AKT, pJUN, c-Jun, phospho-p44/42
MAPK, p38 MAPK, phospho-p38 MAPK, pErk1/2, and Erk1/2) at 4°C
overnight. Subsequently, secondary antibody (sheep anti-rabbit IgG
secondary antibody; cat. no. KC-RB-035; dilution, 1:5,000; or sheep
anti-mouse IgG secondary antibody; cat no. KC-MM-1302; dilution,
1:1,000; Kangchen Bioengineering Co.) incubations were performed at
room temperature for 60 min. The signals were detected using the
Amersham ECL Western Blotting Detection kit (GE Healthcare Life
Sciences, Uppsala, Sweden). GAPDH was used for the loading control.
Densitometry (Image J; National Institutes of Health, Bethesda, MD,
USA) was used to assess the differences in the results.
Statistical analysis
The data averages were based on three individual
experiments that were performed in triplicate. Results for the
experiments were analyzed using Student's t-test. P<0.05 was
considered to indicate a statistically significant difference. SPSS
software, version 11.0 (SPSS, Inc., Chicago, IL, USA), was used for
all statistical analyses.
Results
Effect of LH on ovarian cancer cell
apoptosis
Due to the prevalence of ovarian cancer in
post-menopausal women with ~40 U/l LH, this concentration was
chosen for the present study (8). As
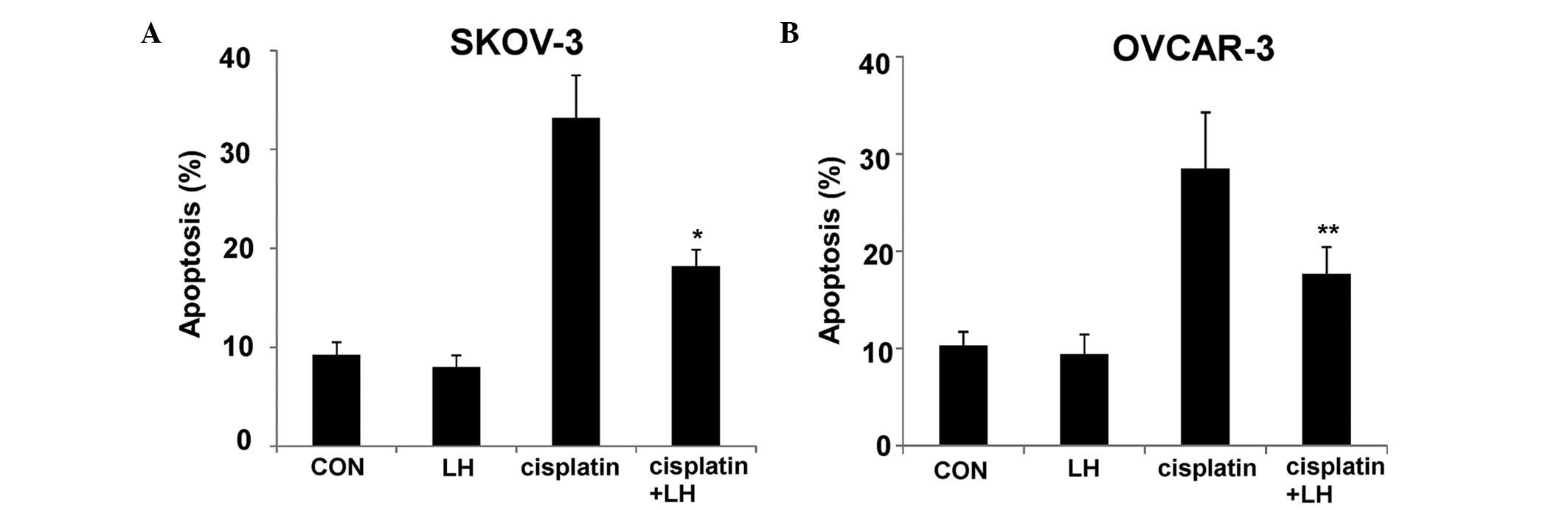
presented in Fig. 1, LH treatment
resulted in a slight decrease in the rate of apoptosis when
compared with controls in the SKOV-3 and OVCAR-3 cells, however,
the difference was not significant (P>0.05). To determine
whether LH could block apoptosis induced by cisplatin, the SKOV-3
and OVCAR-3 cells were treated with 10 µM cisplatin, or a
combination of 40 U/l LH and 10 µM cisplatin. As presented in
Fig. 1, the rate of apoptosis induced
by cisplatin was significantly suppressed by LH (P<0.05).
PDCD6 expression is inhibited by
LH
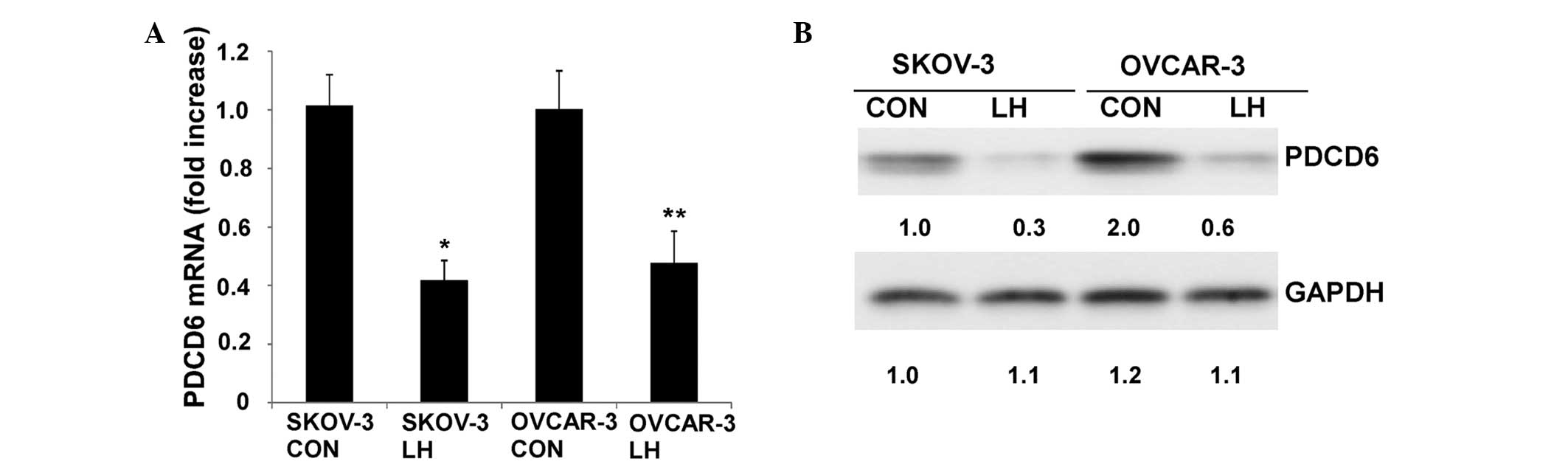
The SKOV-3 and OVCAR-3 cells were treated with 40
U/l LH for up to 24 h. qPCR indicated that PDCD6 mRNA was
significantly downregulated when treated with LH for 16 h
(P<0.05; Fig. 2A). Additionally,
western blotting analysis demonstrated that the protein expression
of PDCD6 decreased when cells were treated with LH for 24 h
(P<0.05) (Fig. 2B).
Signaling transduction pathways
induced by LH
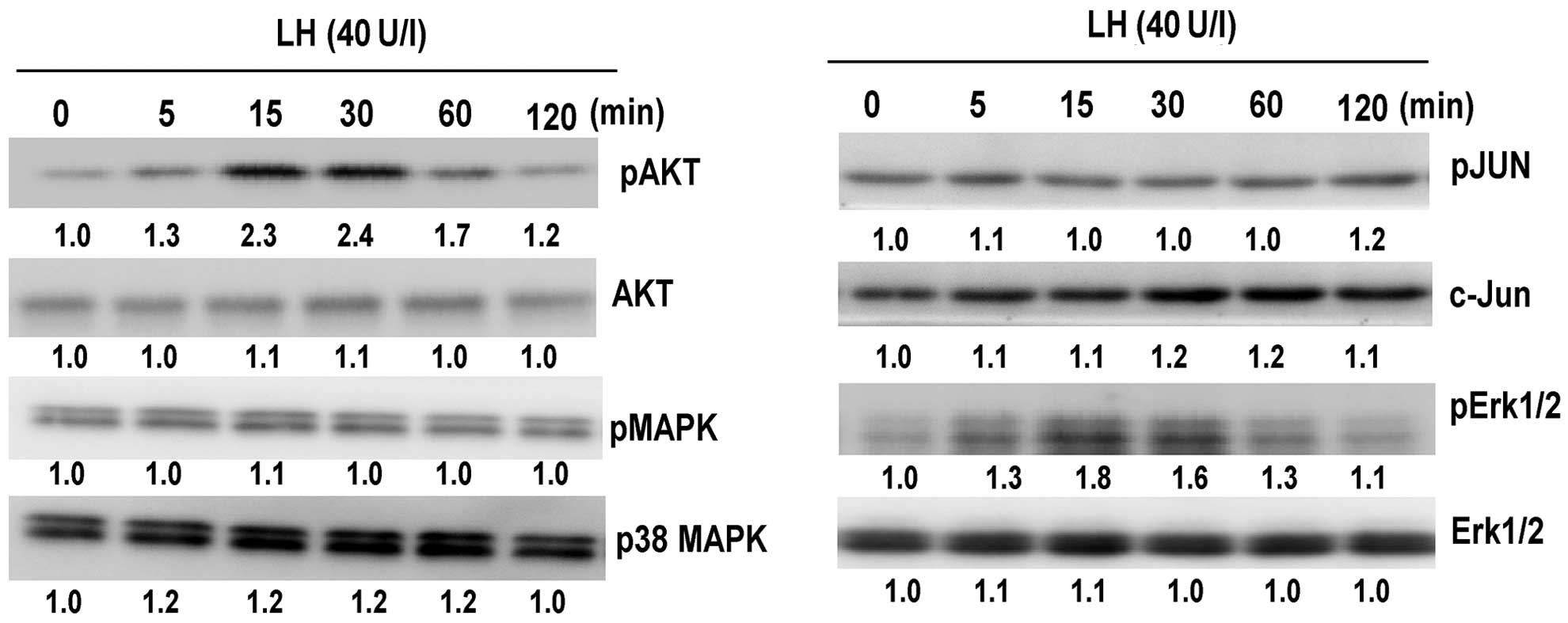
The present study investigated whether the SAPK/JNK,
p38 MAPK, PI3K/AKT and Erk1/2 signaling pathways are involved in
LH-mediated apoptosis. Due to LH exerting similar effects on each
of the cell lines, signaling was only analyzed in the SKOV-3 cells.
It was observed that LH induced the phosphorylation of AKT and
Erk1/2 (P<0.05; Fig. 3), however,
it did not affect the phosphorylation of p38 MAPK and c-Jun
(P>0.05.
LH-induced phosphorylation is blocked
by inhibitors
The SKOV-3 and OVCAR-3 cells were pre-treated with
the inhibitors for 30 min, and LH was then added for an additional
30 min. A pre-treatment with LY294002 or U0126 blocked LH-induced
phosphorylation (Fig. 4).
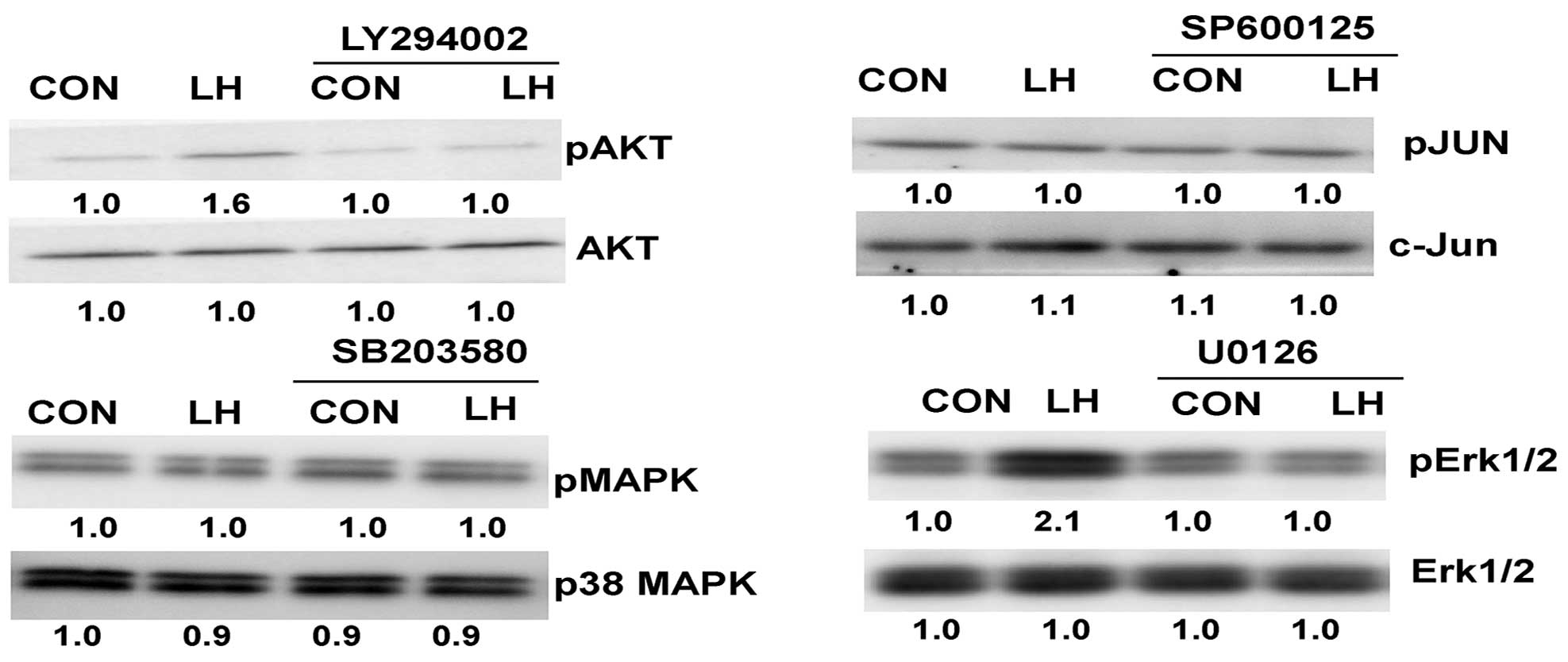 | Figure 4.Western blot analysis demonstrating
suppression of phosphorylation by the inhibitors. Following a
30-min pre-treatment with 10 µM LY294002 (PI3K inhibitor), 10 µM
SP600125 (SAPK/JNK inhibitor), 10 µM U0126 (Erk1/2 inhibitor) or 20
µM SB203580 (p38 MAPK inhibitor), the cells were treated with 40
U/l LH for an additional 30 min. LH, luteinizing hormone; pAKT,
phospho-AKT; MAPK, mitogen-activated protein kinase; pMAPK,
phospho-p38 MAPK; pJUN, phosphorylated c-Jun; Erk1/2, extracellular
signal-regulated kinase 1/2; pErk1/2, phospho-Erk1/2; CON,
control. |
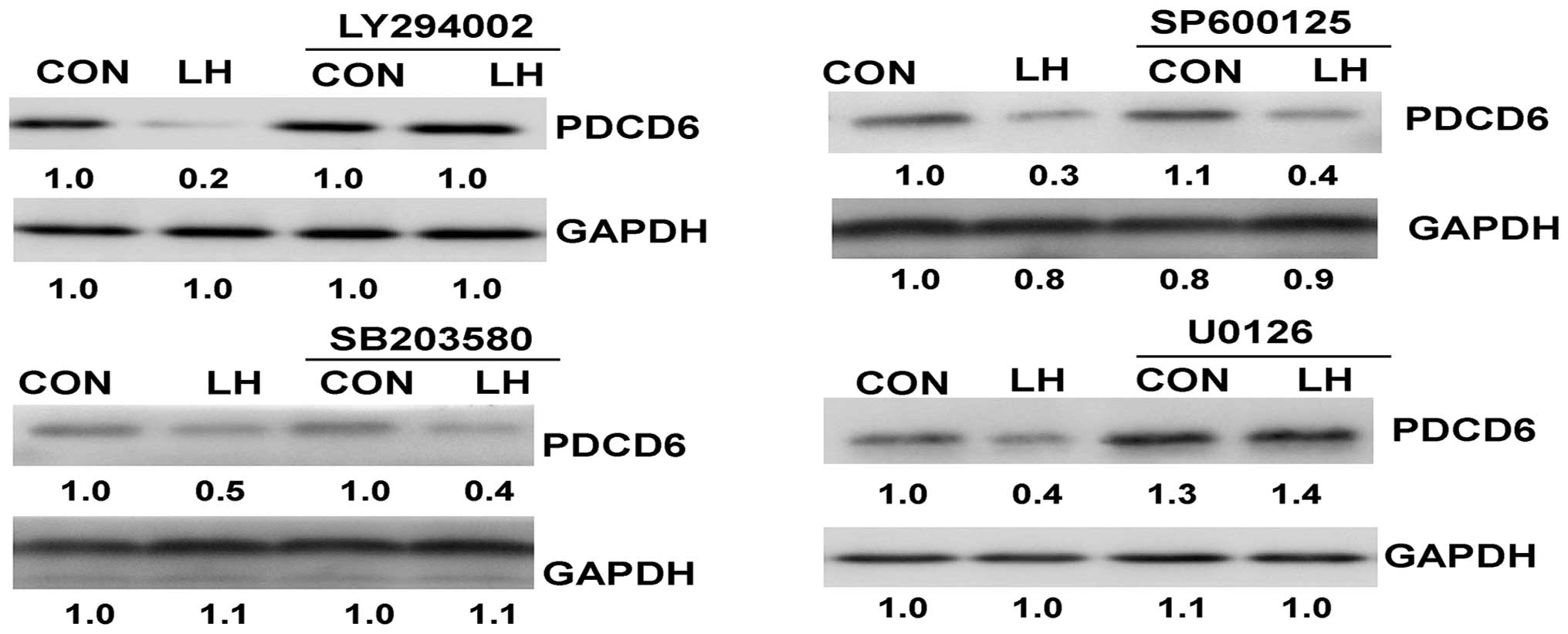
The LH-induced PDCD6 protein
inhibition is neutralized by the inhibitors
The SKOV-3 and OVCAR-3 cells were pre-treated with
the inhibitors for 30 min, and LH was then added for an additional
24 h. LY294002 and U0126 significantly neutralized LH-induced PDCD6
protein inhibition (P<0.05; Fig.
5), however, SP600125 and SB203580 did not have the same effect
(P>0.05).
Discussion
The majority of ovarian tumors develop in
post-menopausal women presenting with high gonadotropin levels.
Thus, gonadotropins are regarded as possible risk factors for the
formation of ovarian tumors (2). In
1992, Ohtani et al (9)
observed that FSH stimulated proliferation in ovarian cancer cells;
subsequent to this, it was reported by Zheng et al (10) that ovarian epithelial tumor growth was
promoted by FSH and inhibited by LH. Choi et al (11) demonstrated that gonadotropins
upregulated epidermal growth factor receptors (EGFRs) through the
activation of MAPK and PI3K in human ovarian surface epithelial
cells. In a further study by Choi et al (12), it was observed that gonadotropins
activated proteolysis and increased invasion through the protein
kinase A (PKA) and PI3K pathways in human epithelial ovarian cancer
cells. Apoptosis serves an important role in the progression of
ovarian cancer (13), and certain
papers have suggested that human chorionic gonadotropin or LH may
inhibit cisplatin-induced apoptosis in ovarian cancer cells
(14,15).
PDCD6 is an important apoptotic mediator and a
prognostic marker for gastric cancer (16); it is also an independent predictor of
progression-free survival in patients with epithelial ovarian
cancer (7). Park et al
(17) demonstrated that PDCD6
additively cooperated with anticancer drugs through the activation
of nuclear factor-κB pathways. It was previously reported that FSH
decreased PDCD6 levels and inhibited the rate of apoptosis in
ovarian cancer cells (5). In the
present study, it was observed that LH significantly blocked
cisplatin-induced apoptosis in vitro. Furthermore, LH
significantly reduced PDCD6 expression. Such results suggest that
the downregulation of PDCD6, induced by LH, appears to be involved
in the chemoresistance of ovarian cancer due to the induction of
apoptosis inhibition.
Numerous studies have focused on the signaling
pathways induced by gonadotropins in ovarian cancer cells and in
human ovarian surface epithelium (OSE). Pon et al (18) reported that gonadotropins reduced
N-cadherin expression in OSE cells through the PKA pathway. It has
also been demonstrated that EGFR expression is regulated by
gonadotropins via cyclic adenosine monophosphate, and this does not
signal through PKA in immortalized OSE cells (19). FSH-induced DNA synthesis and
proliferation may be neutralized by a number of protein kinase C
(PKC) inhibitors, implying that the PKC pathway is also associated
with FSH-induced cell growth (20).
It was previously reported that FSH induced vascular endothelial
growth factor through the PI3K/AKT pathway (21). Slot et al (22) first demonstrated that LH protected HEY
cells from Fas-induced apoptosis, occurring via a signaling cascade
involving PKA. The Erk1/2 pathway has been identified to be
involved in LH-induced survivin expression and apoptosis (15). A previous study indicated that FSH
induced PDCD6 expression through activation of the PI3K/AKT and
SAPK/JNK signaling pathways, and that treatment with LY294002, the
specific PI3K inhibitor, antagonized the effects of FSH on pAKT and
PDCD6 expression; furthermore, the study reported that treatment
with SP600125, the specific SAPK/JNK inhibitor, antagonized the
effects of FSH on pJUN and PDCD6 expression (5). It remains unknown whether FSH and LH
regulate PDCD6 expression in ovarian cancer through different or
identical signaling pathways. The results of the present study
indicate that LH inhibits PDCD6 expression in ovarian cancer cells
by activating the PI3K/AKT and Erk1/2 signaling pathways.
In conclusion, the current study demonstrated that
LH inhibited cisplatin-induced apoptosis and suppressed PDCD6 in
ovarian cancer cells. LH inhibited PDCD6 through the activation of
the PI3K/AKT and Erk1/2 signaling pathways. The results suggest
that LH serves a critical role in the chemoresistance of ovarian
cancer based on the anti-apoptotic effect it induces.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mertens-Walker I, Baxter RC and Marsh DJ:
Gonadotropin signalling in epithelial ovarian cancer. Cancer Lett.
324:152–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dong Z, Yang L and Lai D: KLF5 strengthens
drug resistance of ovarian cancer stem-like cells by regulating
survivin expression. Cell Prolif. 46:425–435. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duiker EW, Meijer A, van der Bilt AR,
Meersma GJ, Kooi N, van der Zee AGJ, de Vries EG and de Jong S:
Drug-induced caspase 8 upregulation sensitises cisplatin-resistant
ovarian carcinoma cells to rhTRAIL-induced apoptosis. Br J Cancer.
104:1278–1287. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang Y, Jin H, Liu Y, Zhou J, Ding J,
Cheng KW, Yu Y and Feng Y: FSH inhibits ovarian cancer cell
apoptosis by up-regulating survivin and down-regulating PDCD6 and
DR5. Endocr Relat Cancer. 18:13–26. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vito P, Lacanà E and D'Adamio L:
Interfering with apoptosis: Ca(2+)-binding protein ALG-2 and
Alzheimer's disease gene ALG-3. Science. 271:521–525. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Su D, Xu H, Feng J, Gao Y, Gu L, Ying L,
Katsaros D, Yu H, Xu S and Qi M: PDCD6 is an independent predictor
of progression free survival in epithelial ovarian cancer. J Transl
Med. 10:312012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Brodowska A, Laszczyńska M, Brodowski J,
Masiuk M and Starczewski A: Analysis of pituitary gonadotropin
concentration in blood serum and immunolocalization and
immunoexpression of follicle stimulating hormone and luteinising
hormone receptors in ovaries of postmenopausal women. Histol
Histopathol. 27:241–248. 2012.PubMed/NCBI
|
|
9
|
Ohtani K, Sakamoto H and Satoh K:
Stimulatory effects of follicular stimulating hormone on the
proliferation of ovarian cancer cell line in vitro and in
vivo. Nihon Sanka Fujinka Gakkai Zasshi. 44:717–724. 1992.(In
Japanese). PubMed/NCBI
|
|
10
|
Zheng W, Lu JJ, Luo F, Zheng Y, Feng Y,
Felix JC, Lauchlan SC and Pike MC: Ovarian epithelial tumor growth
promotion by follicle-stimulating hormone and inhibition of the
effect by luteinizing hormone. Gynecol Oncol. 76:80–88. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Choi JH, Choi KC, Auersperg N and Leung
PC: Gonadotropins upregulate the epidermal growth factor receptor
through activation of mitogen-activated protein kinases and
phosphatidyl-inositol-3-kinase in human ovarian surface epithelial
cells. Endocr Relat Cancer. 12:407–421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi JH, Choi KC, Auersperg N and Leung
PC: Gonadotropins activate proteolysis and increase invasion
through protein kinase A and phosphatidylinositol 3-kinase pathways
in human epithelial ovarian cancer cells. Cancer Res. 66:3912–3920.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao JX, Liu H, Lv J and Yang XJ:
Wortmannin enhances cisplatin-induced apoptosis in human ovarian
cancer cells in vitro. Eur Rev Med Pharmacol Sci.
18:2428–2434. 2014.PubMed/NCBI
|
|
14
|
Kuroda H, Mandai M, Konishi I, Yura Y,
Tsuruta Y, Hamid AA, Nanbu K, Matsushita K and Mori T: Human
chorionic gonadotropin (hCG) inhibits cisplatin-induced apoptosis
in ovarian cancer cells: possible role of up-regulation of
insulin-like growth factor-1 by hCG. Int J Cancer. 76:571–578.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Z, Liao H, Chen X, Zheng Y, Liu Y,
Tao X, Gu C, Dong L, Duan T, Yang Y, et al: Luteinizing hormone
upregulates survivin and inhibits apoptosis in ovarian epithelial
tumors. Eur J Obstet Gynecol Reprod Biol. 155:69–74. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yoon JH, Choi YJ, Kim SG, Nam SW, Lee JY
and Park WS: Programmed cell death 6 (PDCD6) as a prognostic marker
for gastric cancers. Tumour Biol. 33:485–494. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park SH, Lee JH, Lee GB, Byun HJ, Kim BR,
Park CY, Kim HB and Rho SB: PDCD6 additively cooperates with
anti-cancer drugs through activation of NF-κB pathways. Cell
Signal. 24:726–733. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pon YL, Auersperg N and Wong AS:
Gonadotropins regulate N-cadherin-mediated human ovarian surface
epithelial cell survival at both post-translational and
transcriptional levels through a cyclic AMP/protein kinase A
pathway. J Biol Chem. 280:15438–15448. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Choi JH, Chen CL, Poon SL, Wang HS and
Leung PC: Gonadotropin-stimulated epidermal growth factor receptor
expression in human ovarian surface epithelial cells: Involvement
of cyclic AMP-dependent exchange protein activated by cAMP pathway.
Endocr Relat Cancer. 16:179–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ohtani K, Sakamoto H, Kikuchi A, Nakayama
Y, Idei T, Igarashi N, Matukawa T and Satoh K: Follicle-stimulating
hormone promotes the growth of human epithelial ovarian cancer
cells through the protein kinase C-mediated system. Cancer Lett.
166:207–213. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang Y, Hua K, Zhou X, Jin H, Chen X, Lu
X, Yu Y, Zha X and Feng Y: Activation of the PI3K/AKT pathway
mediates FSH-stimulated VEGF expression in ovarian serous
cystadenocarcinoma. Cell Res. 18:780–791. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Slot KA, de Boer-Brouwer M, Houweling M,
Vaandrager AB, Dorrington JH and Teerds KJ: Luteinizing hormone
inhibits Fas-induced apoptosis in ovarian surface epithelial cell
lines. J Endocrinol. 188:227–239. 2006. View Article : Google Scholar : PubMed/NCBI
|