Introduction
Glioma is the most common type of primary malignant
cerebral tumor and possesses a high cancer-associated mortality
rate (1). Despite advances in
surgical techniques, chemotherapy drugs and radiotherapy, the
prognosis for glioma patients is poor (2). Malignant tumors may form
immunosuppressive environments to evade immunological attacks
(3). Programmed cell death protein-1
(PD-1) and its ligand programmed death-ligand 1 (PD-L1) have been
widely studied as a crucial pathway in the creation of the tumor
immunosuppressive microenvironment, which causes exhaustion or
dysfunction of antitumor T lymphocytes. A blockade of this pathway
by specific antibodies has resulted in impressive clinical
responses in patients with solid tumors of various origins
(4,5).
T cell immunoglobulin (Ig) and
mucin-domain-containing molecule 3 (Tim-3) is an inhibitory
receptor expressed on the surface of T cells and is critical for
the inhibition of T cell responses against tumors (6). Galectin-9 has been identified as a
ligand for Tim-3, and the binding of galectin-9 to Tim-3 results in
the apoptosis of T cells and a negative regulation of T cell
immunity (7). An in vivo study
demonstrated that administration of Tim-3-Ig partly abrogated
immune tolerance and reversed the suppressed function of immune
cells in patients with chronic hepatitis B (8). Numerous studies support that a blockade
of the galectin-9-Tim-3 pathway may have the potential to restore
the function of tumor-infiltrating lymphocytes in human cancer.
Gliomas have been revealed to induce local and systemic
immunosuppression that limits the immune defense towards the tumor
by the upregulation of anti-inflammatory cytokines and expansion of
immunosuppressive effector cells, including regulatory T cells.
Gliomas may also express immunosuppressive ligands at the cell
surface, such as PD-L1 and B7-H3, an immune checkpoint molecule
(9,10). However, the expression of Tim-3 and
galectin-9 in gliomas has not been well studied. The present study
assessed the expression of Tim-3 on CD4+ and
CD8+ T cells, and galectin-9 expression in tumor tissues
of glioma patients. The significance of the expression of the
galectin-9-Tim-3 pathway in gliomas was evaluated by analyzing the
association between the expression of Tim-3 and galectin-9 and the
clinical indices of glioma patients.
Materials and methods
Patient samples
Peripheral blood and glioma tissues were collected
from 53 patients during surgical resections performed at the Sanbo
Brain Hospital (Beijing, China) between June 2012 and June 2014.
All the patients had been previously diagnosed with glioma and had
not undergone any cancer therapy prior to resection. Patients with
other diseases, including other cancers, were excluded. For control
tissues, 5 non-cancerous brain tissue samples were collected from
patients with severe head injuries, who were undergoing surgery.
All glioma tissue was pathologically graded according to the World
Health Organization (WHO) glioma classification (11). Clinical information, including gender,
age and the Karnofsky Performance Status (KPS) scores, was obtained
from the medical records of the patients.
The present study was authorized by the Ethical
Board of the Sanbo Brain Hospital, and the Institutional Review
Board of Institute of Biophysics (Beijing, China). Written informed
consent was obtained from all the patients.
Cell isolation
Tumor-infiltrating lymphocytes (TILs) were isolated
from glioma patients, as previously described (12,13).
Briefly, the fresh glioma tissues were minced with sterile scissors
and digested with 0.1 mg/ml hyaluronidase (catalog no., H3506;
Sigma-Aldrich, St Louis, MO, USA), 1 mg/ml collagenase IV (catalog
no., 17104-019; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
and 0.01 mg/ml DNase I (catalog no., D5025; Sigma-Aldrich) in
sterile phosphate-buffered saline (PBS; catalog no., P3563;
Sigma-Aldrich) at 37°C for 60 min. The resulting tissue suspension
was filtered through a BD Falcon 100-µm cell strainer (BD
Biosciences, Franklin Lakes, NJ, USA), and layered on 22% Percoll
solution (catalog no., P1644; Sigma-Aldrich) in myelin buffer
[produced according to a previously described protocol (12)], which removed the myelin and cell
debris following continuous centrifugation for 20 min at 950 × g.
The pellet of cells was additionally isolated by continuous density
gradient centrifugation with 25 and 75% Percoll solution for 25 min
at 800 × g. The cells between the 2 layers of Percoll solution were
collected, and they consisted of microglia, tumor infiltrated
macrophages and lymphocytes. Peripheral blood mononuclear cells
(PBMCs) were isolated from venous blood samples by continuous
density Ficoll gradient (Tianjin Haoyang Biology Manufacture Co.,
Ltd., Tianjin, China) centrifugation for 20 min at 800 × g. The
white layer between the plasma and the Ficoll solution was
collected and washed with sterile PBS.
Immunohistochemistry
Out of the 53 glioma tissues, only those that were
large enough underwent immunohistochemical analysis. Therefore, a
total of 40 glioma tissues and 5 non-cancerous brain tissues, which
were obtained from severely brain-injured patients as controls
during surgery, were used in the present study. The tissue sections
were fixed in 4% formalin, embedded in paraffin and were prepared
into 5 µm continuous sections. A streptavidin-biotin complex
immunohistochemistry method was performed to determine the
expression of galectin-9 using the DAB kit (catalog no., ZLI-9018;
ZSBIO, Beijing, China), following the manufacturer's protocols,
with the rabbit anti-human galectin-9 polyclonal antibody
(dilution, 1:500; catalog no., ab183965; Abcam, Cambridge, UK) for
immunolabeling. Each tissue section was viewed at a ×400
magnification through 10 randomly selected fields of view, and the
percentage of positive staining for galectin-9 and the staining
intensity was analyzed by Image-Pro® Plus 6.0 software (Media
Cybernetics, Inc., Rockville, MD, USA). The expression levels of
galectin-9 were scored based on the staining intensity and
distribution using the immunoreactive score (IRS), as follows: IRS
= staining intensity (SI) × percentage of positive cells (PP). The
SI was determined as follows: Absent, 0; weak, 1; moderate, 2; and
strong, 3. The PP was scored as follows: 0%, 0; 0–25%, 1; 25–50%,
2; 50–75%, 3; and 75–100%, 4 (14,15).
Flow cytometry
TILs and PBMCs were resuspended with
fluorescence-activated cell sorting buffer (PBS with 3% fetal
bovine serum; Gibco; Thermo Fisher Scientific Inc.) and blocked
with 10% human health serum (Beijing Red Cross Blood Center,
Beijing, China) for 20 min at 4°C, and then stained with
fluorescence-conjugated antibodies for 30 min at 4°C in the dark.
Flow cytometry analysis was performed using BD FACSCalibur™ (BD
Biosciences). In total, ~10,000 cells were collected, and the data
were analyzed using FlowJo software (FlowJo, LLC, Ashland, OR,
USA). The fluorescence-conjugated antibodies were as follows:
Phycoerythrin (PE)-conjugated human anti-Tim-3 rat IgG2a monoclonal
antibody (catalog no., FAB2365P; dilution, 50 µg/ml−1;
R&D Systems China Co., Ltd., Shanghai, China);
PerCp-Cy5.5-conjugated anti-human cluster of differentiation (CD)3
mouse monoclonal antibody (catalog no., 45-0037-42; dilution, 10
µg/ml−1; eBioscience, San Diego, CA, USA);
allophycocyanin-conjugated anti-human CD8 mouse monoclonal antibody
(catalog no., 17-0086-42; dilution, 10 µg/ml−1;
eBioscience); and PE-conjugated rat IgG2a isotype control antibody
(catalog no. IC006P; dilution, 50 µg/ml−1; R&D
Systems China Co., Ltd.).
Statistical analysis
All data was analyzed using GraphPad Prism 5
software (GraphPad Software, Inc., La Jolla, CA, USA). Two-tailed
Mann-Whitney U test was used to assess galectin-9 and Tim-3
expression in different glioma tissues and the association between
the expression and the WHO glioma grade. Spearman's correlation
analysis was used to calculate the correlation between the KPS
score and the expression of galectin-9 and Tim-3 on T cells.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Patient characteristics
In total, 53 glioma patients (30 men and 23 women)
that possessed different grades of malignant glioma and were
undergoing radical resection were enrolled in the present study.
The characteristics of the patients are reported in Table I. The gliomas of the patients were
classified according to the WHO classification of glioma, as
follows: grade II, 7 patients; grade III, 15 patients; and grade
IV, 31 patients. The KPS score was calculated according to the
clinical records of the patients, as follows: ≥80, 22 patients;
60–80, 8 patients; and ≤60, 13 patients.
 | Table I.Characteristics of glioma and control
patients. |
Table I.
Characteristics of glioma and control
patients.
| Characteristic | Glioma patients, n
(%) | Control patients, n
(%) |
|---|
| Total | 53 (100.0) | 15 (100.0) |
| Gender |
|
|
| Male | 30 (56.6) | 7
(46.7) |
|
Female | 23 (43.4) | 8 (53.3) |
| Age, years |
|
|
|
18–29 | 8
(15.1) | 4 (26.7) |
|
30–50 | 33 (62.3) | 8 (53.3) |
| ≥51
years | 12 (22.6) | 3 (20.0) |
| WHO grade |
|
|
|
II–III | 22 (41.5) | N.A. |
| IV | 31 (58.5) | N.A. |
| KPS |
|
|
|
≥80 | 22 (41.5) | N.A. |
|
60–80 | 18 (34.0) | N.A. |
|
<60 | 13 (24.5) | N.A. |
Tim-3 expression is upregulated on
CD4+ and CD8+ T cells in the peripheral blood
and tumors of glioma patients
The expression levels of Tim-3 in T cells in tumor
tissues and the peripheral blood of glioma patients, as well as in
the peripheral blood of the control patients, was explored. TILs
and PBMCs were analyzed directly by flow cytometry using the human
anti-Tim-3, anti-human CD3 and the anti-human CD8 antibodies.
CD4+ T cells were identified as
CD3+/CD8− cells during flow cytometry
analysis. As revealed in Fig. 1A, the
expression of Tim-3 was slightly detected on T cells in PBMCs of
the control patients, in which the mean ± standard error of the
mean (SEM) of the frequencies of Tim-3+ cells among
CD4+ and CD8+ T cells were 1.92±0.26% and
3.43±0.46%, respectively. The frequency of Tim-3+ cells
was ~2-fold higher on CD4+ (3.73±0.27%; P<0.05) and
CD8+ T cells (6.46±0.52%; P<0.05) in PBMCs from the
glioma patients compared with the control individuals. In addition,
Tim-3 expression was increased ~4-fold on the CD4+ T
cells (5.83±1.02%; P<0.0001) and ~2-fold on the CD8+
T cells (12.27±0.85%; P<0.0001) of TILs compared with the
expression in the PBMCs obtained from the glioma patients (Fig. 1B and C). Similar observations were
made when Tim-3 expression was analyzed as the mean fluorescence
intensity (Fig. 1D and E). No
differences were detected between Tim-3 expression on the
CD4+/CD8+ T cells of PBMCs or TILs in male
and female patients or different age groups (data not shown). These
results demonstrate that Tim-3 expression is upregulated on the T
cells of glioma patients; therefore, the glioma microenvironment
may be important in regulating Tim-3 expression on T cells.
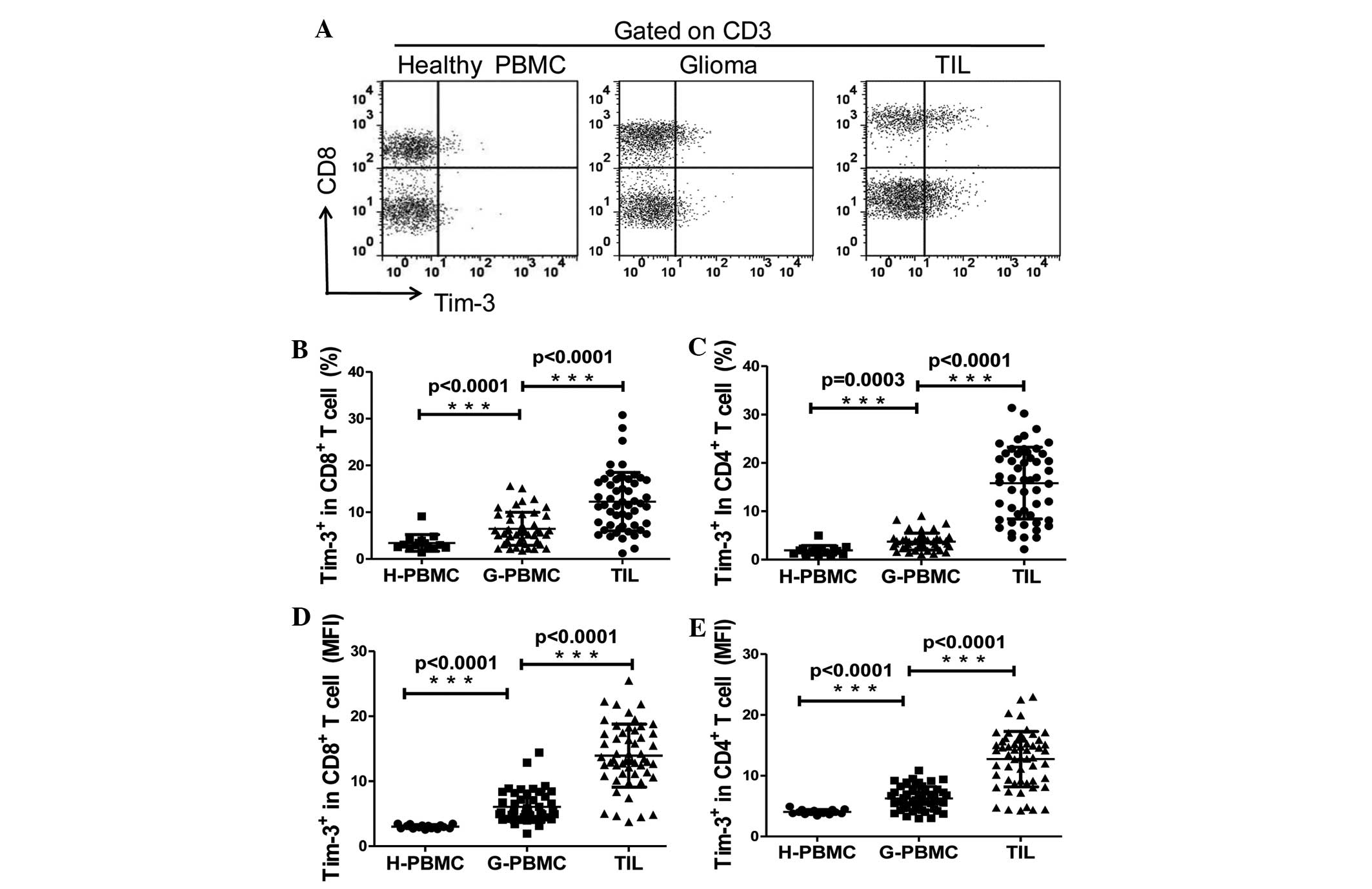 | Figure 1.Expression of Tim-3 on T cells in PBMC
and TILs. (A) A representative flow cytometry analysis of
lymphocytes of PBMCs from control and glioma patients, and TILs
from glioma patients, stained with CD3, CD8 and Tim-3 antibodies.
(B and C) Relative percentage of Tim-3-expressing cells in (B)
CD8+ T cells (CD3+/CD8+; H-PBMC,
3.433±0.4631, n=15; G-PBMC, 6.461±0.5203, n=46; and TILs
12.27±0.8598, n=53). (C) CD4+ T cells
(CD3+/CD8−; H-PBMC, 1.923±0.2562, n=15;
G-PBMC 3.732±0.2574, n=46; and TILs 15.83±1.017, n=53). (D and E)
MFI of Tim-3 expression on (D) CD8+ T cells (H-PBMC,
3.002±0.0788, n=15; G-PBMC 6.065±0.3556, n=46; and TIL
13.93±0.6663, n=53). (E) CD4+ T cells (H-PBMC,
4.041±0.1030, n=15; G-PBMC 6.263±0.2892, n=46; and TILs
12.73±0.6112, n=53). GraphPad Prism 5 software was used to
calculate Mann-Whitney U test. Significance was indicated by
P-values; ***P<0.001. Each symbol represents a single
individual. Data are expressed as the mean ± standard error of the
mean. PBMC, peripheral blood mononuclear cells; CD, cluster of
differentiation; Tim-3, T cell immunoglobulin- and
mucin-domain-containing molecule 3; TILs, tumor-infiltrating
lymphocytes; H-PBMC, control patient PBMC; G-PBMC, glioma patient
PBMC; MFI, mean fluorescence intensity. |
Tim-3 expression on tumor-infiltrating
T cells was associated with clinical pathological
characteristics
Several clinicopathological features have been
clinically used as prognostic factors for glioma, including the WHO
grade of glioma and the KPS score. The KPS score is a standard
method for measuring the ability of cancer patients to perform
ordinary tasks, which is used to evaluate the severity of glioma
(16). A lower KPS score indicates
increased severity of the disease and a reduced survival time of
patients. The WHO grade of glioma (17) and the KPS score have been demonstrated
to be associated with the overall survival rate of patients, since
patients with high-grade glioma and a KPS score of <80 possess a
poorer overall survival rate and a higher risk of mortality
compared with those with a low WHO grade and a KPS score of ≥80. To
investigate the significance of Tim-3 expression on the T cells of
glioma patients, the present study analyzed the association between
the expression level of Tim-3 in T cells and the WHO grade of the
glioma and KPS score of the glioma patients. Tim-3 expression on
the CD4+ and CD8+ T cells of TILs was
increased in grade IV gliomas compared with grade II and III
gliomas, suggesting that Tim-3 expression on T cells in tumors was
significantly associated with the WHO grade of glioma (P<0.0001;
Fig. 2A and B). The frequency of
Tim-3+ cells in CD4+ and CD8+ T
cells of TILs was negatively correlated with the KPS score of
glioma patients (P<0.001, r=−0.6723 and P=0.0005, r=−0.4642,
respectively; Fig. 2C and D). This
data indicates that the expression of Tim-3 on TILs was associated
with glioma severity. The Tim-3 expression on the CD4+ T
cells was slightly increased compared to the expression on the
CD8+ T cells of TILs (mean ± SEM, 15.83±1.02% vs.
12.27±0.85%; Fig. 1B and C); however,
the Tim-3 expression on tumor-infiltrating CD8+ T cells
demonstrated an increased correlation with the KPS score compared
with tumor-infiltrating CD4+ T cells (Fig. 2B). This data demonstrates that the
expression of Tim-3 on tumor-infiltrating T cells was associated
with the prognosis of glioma patients.
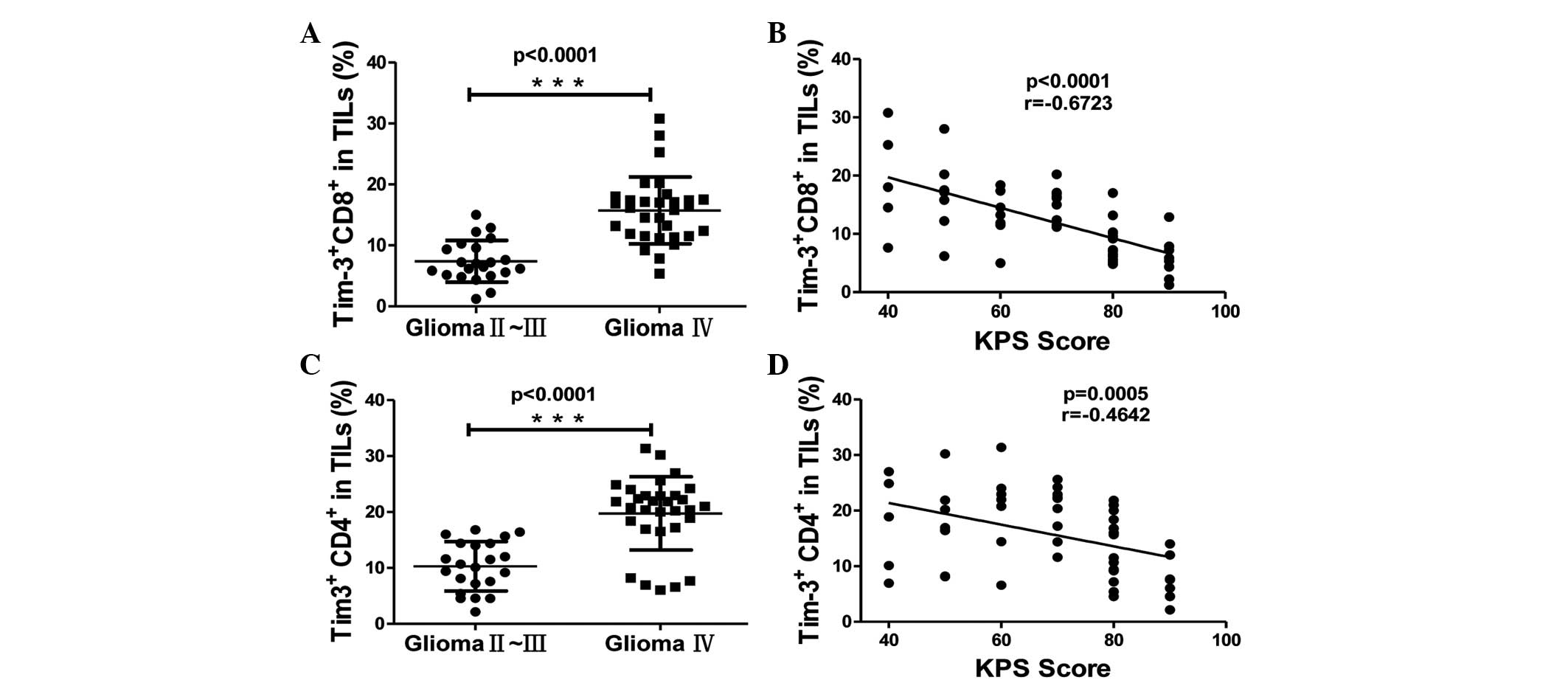 | Figure 2.The association between Tim-3
expression on CD4+ and CD8+ T cells in TILs
and the clinicalpathological characteristics of patients. (A and B)
Association between Tim-3 expression and the World Health
Organisation grade of glioma, as calculated by Mann-Whitney U test,
on (A) CD8+ (glioma grade II–III, 7.391±0.727, n=22; and
glioma grade IV, 15.72±0.9835, n=31) and (B) CD4+ T
cells (glioma grade II–III, 10.29±0.9405, n=22; and glioma grade
IV, 19.76±1.117, n=31). (C and D) Correlation between Tim-3
expression and the KPS score of patients, as calculated by
Spearman's correlation analysis, on (C) CD8+ and (D)
CD4+ T cells. Significance was indicated by P-values,
***P<0.001. Each symbol represents a single individual. Data are
expressed as the mean ± standard error of the mean. CD, cluster of
differentiation; Tim-3, T cell immunoglobulin- and
mucin-domain-containing molecule 3; TILs, tumor-infiltrating
lymphocytes; KPS, Karnofsky Performance Status. |
Galectin-9 expression in tumors was
associated with the WHO grade of glioma and Tim-3 expression on the
T cells of TILs. Galectin-9 is a ligand for Tim-3
The interaction of Tim-3 with galectin-9 induces the
apoptosis of Tim-3+ T cells and inhibits T helper cell
(Th)-1 responses. To assess the significance of Tim-3 signaling in
gliomas, the present study examined the expression of galectin-9 in
tumor cells using immunohistochemistry. Galectin-9 was expressed at
an extremely low level in non-cancerous brain tissues from control
patients, but there was an increased expression in glioma patient
tissues. The expression level of galectin-9 in tumor tissues was
scored as the IRS based on the intensity and distribution of
galectin-9 staining. The IRS of galectin-9 was correlated with the
WHO grade of glioma. The expression of galectin-9 in grade IV
gliomas was significantly increased compared with grade II–III
gliomas (Fig. 3A and B; P=0.008 vs.
P=0.0002, respectively). In addition, galectin-9 expression in
glioma tissue was associated with Tim-3 expression on
CD4+ and CD8+ T cells of TILs (Fig. 3C and D; P=0.0094, r=0.406 and P=0.001,
r=0.502, respectively). This data indicates that the
galectin-9/Tim-3 pathway may be critical in the progression of
glioma.
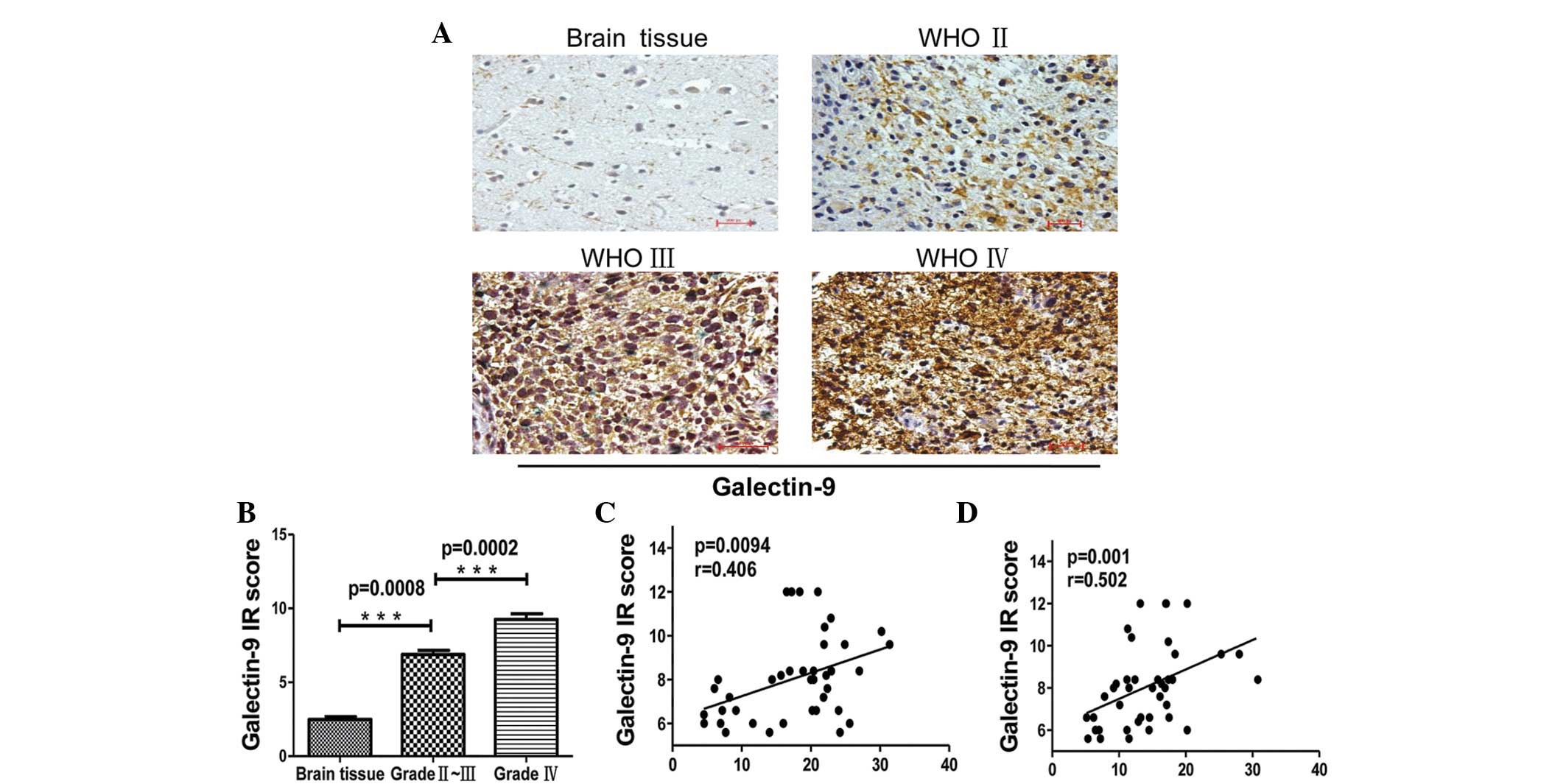 | Figure 3.Immunohistochemical staining of glioma
tissues for galectin-9 expression. (A) Paraffin-embedded tissue
sections from injured brain tissue and different grades of glioma
tissues were stained with galectin-9-specific monoclonal
antibodies. Representative patterns of galectin-9 expression in
brain tissues and different grades of glioma (magnification, ×200).
(B) Plot of the scores of galectin-9 expression vs. WHO grade of
glioma and IR scores of different grades of glioma (brain tissue,
2.480±0.1855, n=5; glioma grade II–III, 6.880±0.2704, n=20; and
glioma grade IV, 9.260±0.3899, n=20). (C and D) Association between
galectin-9 in glioma tissues and Tim-3 expression in (C)
CD4+ and (D) CD8+ T cells in
tumor-infiltrating lymphocytes. P-values were determined by
Mann-Whitney U test and Spearman's correlation analysis,
***P<0.001.. Data are expressed as the mean ± standard error of
the mean. WHO, World Health Organisation; IR, immunoreactive score;
Tim-3, T cell immunoglobulin- and mucin-domain-containing molecule
3; CD, cluster of differentiation. |
Discussion
Tim-3 is a receptor expressed on Th1 cells and
CD8+ T cytotoxic type 1 cells (18). Galectin-9 was identified as a ligand
of Tim-3, and is expressed by numerous cells (7). Galectin-9 expression may be induced by
interferon (IFN)-γ from numerous tissue types (19,20). When
galectin-9 binds to Tim-3 the apoptosis of T cells may be induced
(18) and T cell responses may be
inhibited (21). The Tim-3-galectin-9
pathway is considered to be a negative regulator for T
cell-mediated immune responses (22).
The present study investigated the expression of Tim-3 and
galectin-9 in glioma tissues and identified an association between
the expression of Tim-3 and galectin-9 and the malignancy of
gliomas.
In general, the glioma environment may remold and
immunoedit immune cells that infiltrate the tumor, resulting in the
tumor cells escaping from immune surveillance (23). Several mechanisms of tumor cell immune
evasion have been studied. One mechanism is the expression of
immune checkpoint molecules on activated T cells infiltrating the
tumor, including PD-1, cytotoxic T-lymphocyte-associated protein-4
and Tim-3 (24). Tim-3 is often
co-expressed with PD-1 on exhausted T cells, which are T cells with
a decreased ability to express cytotoxic cytokines, including IFN-γ
and tumor necrosis factor-α, when they are continuously exposed to
antigens (22). Co-blockade of the
two pathways has been demonstrated to be an effective way of
reversing the exhaustion of CD8+ T cells (25). Tim-3 may be highly upregulated on
tumor-infiltrated-lymphocytes compared with T cells in the
peripheral blood of tumors (26,27). In
addition, the present study detected a low expression level of
Tim-3 on healthy PBMCs, and a low expression of galectin-9 on
non-cancerous brain tissues. However, Tim-3 and galectin-9 were
expressed at an increased level in TILs and glioma tissues,
respectively, and the expression was associated with tumor
malignancy. The present study suggests that malignant tumors may
create an immunosuppressive environment by increasing the
expression of immunosuppressive molecules, including galectin-9, in
tumors. When activated T cells infiltrate the tumors, the
interaction of Tim-3 on T cells with galectin-9 expressed by tumors
induces dysfunction or exhaustion of the tumor-infiltrating T
cells.
Tumor progression is a complex process with numerous
genes and multiple signaling pathways becoming activated. It is
challenging to identify particular molecules that are associated
with the severity of the clinical diagnosis and prognosis of
glioma. The KPS score is widely used to evaluate the severity of
glioma (16). The present study
investigated the correlation between the Tim-3 expression on TILs
and the KPS score. The expression of Tim-3 on CD4+ and
CD8+ T cells was negatively correlated with the KPS
score. This data indicates that Tim-3 may be involved in the
progression of glioma.
Glioma is a fatal disease of the nervous system, and
there are numerous challenges in treating this disease.
Immunotherapy is a promising method for the treatment of malignant
tumors (28). However, the mechanism
by which glioma cells escape immunological surveillance is unknown,
and therefore it is critical to identify efficient immune methods
to treat the disease. The present study systemically investigated
the expression of Tim-3 in CD4+ and CD8+ T
cells in TILs, the expression of the Tim-3 ligand galectin-9 in
glioma tissue, and the association of these two proteins with tumor
malignancy and clinical pathological characteristics of patients.
The data from the present study suggests that Tim-3-galectin-9
signaling may be a critical pathway in the immunoevasion of glioma
cells. A blockade of the Tim-3 pathway may delay the exacerbation
of glioma and enhance the KPS score, therefore improving clinical
symptoms and prolonging the life of the patient.
Acknowledgements
The authors would like to thank Dr Xuexiang Du from
the Key Laboratory of Infection and Immunity, Institute of
Biophysics, University of Chinese Academy of Sciences (Beijing,
China), for technical assistance. The present study was finished
under the guidance of Professor SD Wang, and was supported in part
by the National Natural Science Foundation of China (grant no.
81302200).
References
|
1
|
Liu Y, Shete S, Hosking F, Robertson L,
Houlston R and Bondy M: Genetic advances in glioma: Susceptibility
genes and networks. Curr Opin Genet Dev. 20:239–244. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: European Organisation for Research and Treatment of
Cancer Brain Tumor and Radiotherapy Groups; National Cancer
Institute of Canada Clinical Trials Group: Radiotherapy plus
concomitant and adjuvant temozolomide for glioblastoma. N Engl J
Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Okada H, Kohanbash G, Zhu X, Kastenhuber
ER, Hoji A, Ueda R and Fujita M: Immunotherapeutic approaches for
glioma. Crit Rev Immunol. 29:1–42. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Flies DB, Sandler BJ, Sznol M and Chen L:
Blockade of the B7-H1/PD-1 pathway for cancer immunotherapy. Yale J
Biol Med. 84:409–421. 2011.PubMed/NCBI
|
|
5
|
Topalian SL, Hodi FS, Brahmer JR,
Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD,
Sosman JA, Atkins MB, et al: Safety, activity, and immune
correlates of anti-PD-1 antibody in cancer. N Engl J Med.
366:2443–2454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koguchi K, Anderson DE, Yang L, O'Connor
KC, Kuchroo VK and Hafler DA: Dysregulated T cell expression of
TIM3 in multiple sclerosis. J Exp Med. 203:1413–1418. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu C, Anderson AC, Schubart A, Xiong H,
Imitola J, Khoury SJ, Zheng XX, Strom TB and Kuchroo VK: The Tim-3
ligand galectin-9 negatively regulates T helper type 1 immunity.
Nat Immunol. 6:1245–1252. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ju Y, Hou N, Meng J, Wang X, Zhang X, Zhao
D, Liu Y, Zhu F, Zhang L, Sun W, et al: T cell immunoglobulin- and
mucin-domain-containing molecule-3 (Tim-3) mediates natural killer
cell suppression in chronic hepatitis B. J Hepatol. 52:322–329.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lemke D, Pfenning PN, Sahm F, Klein AC,
Kempf T, Warnken U, Schnölzer M, Tudoran R, Weller M, Platten M and
Wick W: Costimulatory protein 4IgB7H3 drives the malignant
phenotype of glioblastoma by mediating immune escape and
invasiveness. Clin Cancer Res. 18:105–117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wainwright DA, Chang AL, Dey M,
Balyasnikova IV, Kim CK, Tobias A, Cheng Y, Kim JW, Qiao J, Zhang
L, et al: Durable therapeutic efficacy utilizing combinatorial
blockade against IDO, CTLA-4, and PD-L1 in mice with brain tumors.
Clin Cancer Res. 20:5290–5301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
De Groot CJ, Montagne L, Janssen I, Ravid
R, Van Der Valk P and Veerhuis R: Isolation and characterization of
adult microglial cells and oligodendrocytes derived from postmortem
human brain tissue. Brain Res Brain Res Protoc. 5:85–94. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Olah M, Raj D, Brouwer N, De Haas AH,
Eggen BJ, Den Dunnen WF, Biber KP and Boddeke HW: An optimized
protocol for the acute isolation of human microglia from autopsy
brain samples. Glia. 60:96–111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Berns K, Horlings HM, Hennessy BT,
Madiredjo M, Hijmans EM, Beelen K, Linn SC, Gonzalez-Angulo AM,
Stemke-Hale K, Hauptmann M, et al: A functional genetic approach
identifies the PI3K pathway as a major determinant of trastuzumab
resistance in breast cancer. Cancer Cell. 12:395–402. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mack PC, Redman MW, Chansky K, Williamson
SK, Farneth NC, Lara PN Jr, Franklin WA, Le QT, Crowley JJ and
Gandara DR: SWOG: Lower osteopontin plasma levels are associated
with superior outcomes in advanced non-small-cell lung cancer
patients receiving platinum-based chemotherapy: SWOG Study S0003. J
Clin Oncol. 26:4771–4776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang L, He S, Tu Y, Ji P, Zong J, Zhang J,
Feng F, Zhao J, Gao G and Zhang Y: Downregulation of chromatin
remodeling factor CHD5 is associated with a poor prognosis in human
glioma. J Clin Neurosci. 20:958–963. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ji Y, Wei Y, Wang J, Ao Q, Gong K and Zuo
H: Decreased expression of microRNA-107 predicts poorer prognosis
in glioma. Tumour Biol. 36:4461–4466. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Monney L, Sabatos CA, Gaglia JL, Ryu A,
Waldner H, Chernova T, Manning S, Greenfield EA, Coyle AJ, Sobel
RA, et al: Th1-specific cell surface protein Tim-3 regulates
macrophage activation and severity of an autoimmune disease.
Nature. 415:536–541. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Asakura H, Kashio Y, Nakamura K, Seki M,
Dai S, Shirato Y, Abedin MJ, Yoshida N, Nishi N, Imaizumi T, et al:
Selective eosinophil adhesion to fibroblast via IFN-gamma-induced
galectin-9. J Immunol. 169:5912–5918. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park WS, Jung WK, Park SK, Heo KW, Kang
MS, Choi YH, Kim GY, Park SG, Seo SK, Yea SS, et al: Expression of
galectin-9 by IFN-γ stimulated human nasal polyp fibroblasts
through MAPK, PI3K, and JAK/STAT signaling pathways. Biochem
Biophys Res Commun. 411:259–264. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ungerer C, Quade-Lyssy P, Radeke HH,
Henschler R, Königs C, Köhl U, Seifried E and Schüttrumpf J:
Galectin-9 is a suppressor of T and B cells and predicts the immune
modulatory potential of mesenchymal stromal cell preparations. Stem
Cells Dev. 23:755–766. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Anderson AC, Anderson DE, Bregoli L,
Hastings WD, Kassam N, Lei C, Chandwaskar R, Karman J, Su EW,
Hirashima M, et al: Promotion of tissue inflammation by the immune
receptor Tim-3 expressed on innate immune cells. Science.
318:1141–1143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rolle CE, Sengupta S and Lesniak MS:
Mechanisms of immune evasion by gliomas. Adv Exp Med Biol.
746:53–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Anderson AC: Tim-3, a negative regulator
of anti-tumor immunity. Curr Opin Immunol. 24:213–216. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fourcade J, Sun Z, Benallaoua M, Guillaume
P, Luescher IF, Sander C, Kirkwood JM, Kuchroo V and Zarour HM:
Upregulation of Tim-3 and PD-1 expression is associated with tumor
antigen-specific CD8+ T cell dysfunction in melanoma patients. J
Exp Med. 207:2175–2186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sakuishi K, Apetoh L, Sullivan JM, Blazar
BR, Kuchroo VK and Anderson AC: Targeting Tim-3 and PD-1 pathways
to reverse T cell exhaustion and restore anti-tumor immunity. J Exp
Med. 207:2187–2194. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Baitsch L, Baumgaertner P, Devêvre E,
Raghav SK, Legat A, Barba L, Wieckowski S, Bouzourene H, Deplancke
B, Romero P, et al: Exhaustion of tumor-specific CD8+ T cells in
metastases from melanoma patients. J Clin Invest. 121:2350–2360.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Calinescu AA, Kamran N, Baker G, Mineharu
Y, Lowenstein PR and Castro MG: Overview of current
immunotherapeutic strategies for glioma. Immunotherapy.
7:1073–1104. 2015. View Article : Google Scholar : PubMed/NCBI
|














